



物种丰富度在空间上的分布格局是一系列复杂过程的结果, 反映了物种与环境的关系。因此, 研究物种丰富度空间分布格局的形成机制或影响因子对于解决当今的生态环境问题有着重要的指导意义。物种丰富度空间分布格局受到多种因素的影响, 如气候、生产力、人类活动、生境异质性、海拔等。气候可以直接或间接地影响物种分布。直接影响源于特定生物对于某气候因素的生理耐受性, 气候直接限制了该物种的分布范围(Hawkins et al, 2003)。另外, 气候可以通过影响某个地区生物赖以生存的其他环境因素(如植被类型、食物丰度、栖息地环境等)间接影响物种的空间分布(Davies et al, 2007; Jetz et al, 2009)。生产力假说也是解释物种分布的重要理论, 其认为在生产力高的生态系统中有着充足可靠的可用资源, 因此单位面积上能够维持更大的种群数量, 同时充足的资源也降低了物种灭绝的风险, 即物种丰富度与生产力呈正相关关系(Hurlbert & Stegen, 2014)。而生境异质性假说则认为, 生境异质性高的地方能提供的生态位更多, 有利于多个物种共存, 因而物种多样性更高(Tews et al, 2004; Stein & Kreft, 2015; Stein et al, 2015)。另外, 海拔综合了多种环境因子的梯度效应, 是影响较大区域范围物种丰富度空间分布格局的重要因素之一(Kratochwil, 1999; Virkkala et al, 2010; Hof et al, 2012)。最后, 由于人类活动造成了自然资源量改变、生境面积缩减以及土地覆被类型改变, 对自然生态系统有着不可忽视的作用, 因此人类活动也是影响物种丰富度空间分布格局的重要因素之一(He et al, 2018)。
山地生态系统在全球生态系统中的地位显要, 是研究物种丰富度的代表。山地具有很高的生境异质性, 并且其环境随着地质构造、侵蚀和气候变化过程而不断改变(Badgley et al, 2017; Rahbek et al, 2019b)。持续变化的环境使得物种扩散并产生了地理隔离, 并通过长时间的进化过程导致物种的分化, 最终构建了山地高水平的物种丰富度(Antonelli et al, 2018): 全球大陆山地面积占全球陆地面积的比例不到15%, 但是却承载了全球约87%的两栖类、鸟类和哺乳类物种丰富度(Rahbek et al, 2019a; García- Rodríguez et al, 2021)。因此, 从进化过程上看, 山地是为物种形成提供自然条件的摇篮(Fjeldså et al, 2012; Rangel et al, 2018)。
南岭是我国11个具有国际意义的陆地生物多样性关键地区之一(中国生物多样性国情研究报告编写组, 1998), 属于《中国生物多样性保护战略与行动计划》(2011-2030年)确定的生物多样性保护优先区域(环境保护部, 2011)。同时, 南岭作为中国南方分布范围最广的山脉, 是中国重要的自然地理界线, 也是长江水系、珠江水系的分水岭(庄长伟等, 2021)。虽然南岭具有丰富的生物资源, 但是过去的相关研究以局部区域为主, 如广东车八岭自然保护区的鸟类多样性分析(黄石林等, 2003)、南岭国家级自然保护区3种鸟类海拔梯度上的丰度季节性变化(Zou et al, 2019), 以及广东连平黄牛石自然保护区的兽类初步调查(彭红元等, 2006)。然而, 对于整个南岭山脉哺乳类和鸟类物种丰富度及其主要影响因子的研究依然匮乏。
本研究通过详尽的文献数据收集南岭地区鸟兽物种记录, 结合南岭地区哺乳类和鸟类的地理分布数据构建整个南岭山脉地区上述类群的物种丰富度空间分布格局, 并结合环境因子(气候、生产力、人类活动足迹、生境异质性和海拔), 利用路径分析和空间误差模型探究其潜在影响因子。通过探索南岭地区哺乳类和鸟类物种丰富度的空间分布格局及其主要影响因子, 期望为南岭及其周边区域的保护提供科学依据。
1 材料与方法
1.1 研究区域
南岭山脉位于中国福建、湖南、江西、广东、广西五省(区)交界处, 为东西走向。地球同纬度最大面积的亚热带常绿阔叶林就位于南岭(徐卫等, 2022)。南岭海拔落差明显, 其中主要的山岭如越城岭、都庞岭主峰的海拔均可达2,000 m。整个南岭山脉连续性不高, 在各个高山之间有大面积东西走向的隘口和低海拔的丘陵山地(海拔500-1,200 m), 兼有小面积的盆地、阶地、平原、丘陵(如喀斯特地貌) (缪绅裕等, 2020)。为涵盖地理学意义上的整个南岭山脉, 本研究选择南岭山脉及其周边地区(23°37′-27°14′ N, 109°43′-116°41′ E; 图1)为研究区域, 该区面积约为168,193 km2。
图1
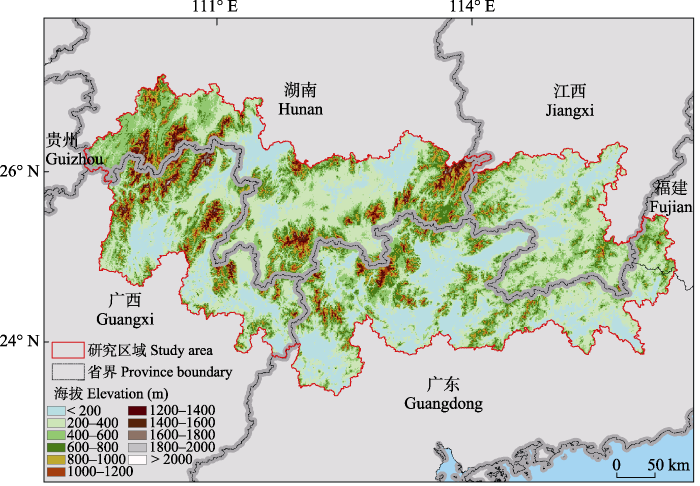
图1
南岭山脉及其周边区域地形图
Fig. 1
Generalized topographic map of the Nanling Mountains and surrounding area
1.2 物种分布数据的获取
我们通过查阅发表时间在1980年至今的相关文献及专著, 构建南岭哺乳类和鸟类的名录。文献获取方式为: 在中国知网、万方文献数据库, 以“南岭”、“南岭五岭” (越城岭、都庞岭、萌渚岭、骑田岭、大庾岭)或南岭五岭周边山地具体地名搭配“哺乳类(兽类)”、“鸟类”为关键词; 在Web of Science数据库, 以“Nanling”、“the five ridges” (Yuechengling, Dupangling, Mengzhuling, Qitianling, Dayuling)或南岭五岭周边山地具体地名(如“Xinfeng County”)搭配“mammal”、“bird”为关键词, 进行文献检索(附录1)。另外, 通过南岭所在地区的地方动物志, 补充南岭区域哺乳类和鸟类的物种丰富度数据。将所有南岭区域物种记录整合, 得到研究区域的哺乳类和鸟类物种名录, 并将其与中国观鸟记录中心网站(
1.3 哺乳类和鸟类物种丰富度空间分布格局的构建
根据上述的南岭地区哺乳类和鸟类物种名录, 我们从世界自然保护联盟濒危物种红色名录(IUCN Red List of Threatened Species,
1.4 影响因子的获取
我们引入8个变量对南岭区域哺乳类和鸟类物种丰富度空间分布格局的影响因子进行分析, 包括气候因子、净初级生产力、人类足迹指数(human footprint index, HFP)、生境异质性(habitat heterogeneity, HH)和海拔(elevation, ELE), 分别对应于气候假说、生产力假说、人类活动的影响、生境异质性和海拔。我们在WorldClim数据库(
1.5 统计分析
根据已有研究, 本研究选用拟合优度指数(goodness-of-fit index, GFI)、比较适配指数(comparative fit index, CFI)和增值拟合指数(incremental fit index, IFI)对模型进行适配性检验, 一般认为, 若GFI、CFI和IFI都大于0.9, 则模型拟合良好(Grace, 2006; 温涵和梁韵斯, 2015; Xie et al, 2020)。另外, 我们计算了直接路径模型各个自变量的方差膨胀因子(variance inflation factor, VIF)以检验变量之间的潜在共线性; 模型的变量VIF值都小于10 (附录3), 其共线性属于可控范围(Dormann et al, 2013)。
考虑到各环境因子(变量)对物种丰富度空间分布格局的解释力可能会受到空间自相关的影响, 因此我们加入空间误差模型(spatial error model, SEM)对路径分析进行补充。我们首先将(正方形)网格外围两圈的24个网格作为邻接要素(neighbor), 计算出该网格中心点距离外围网格中心点的距离, 并创建基于距离范围的空间邻接矩阵; 然后对邻接要素之间的权重按照距离衰减的原则进行赋值, 创建空间权重矩阵; 最后创建7个自变量的空间误差模型。
式中, ε为误差项向量, λ为空间误差参数, W为n × n阶的空间权重矩阵, μ为满足正态分布的随机误差向量。
上述分析均在R 3.4.3中进行, 其中, 路径分析使用lavaan包完成, 方差膨胀因子计算使用car包完成, 空间邻接矩阵和权重矩阵的创建使用spdep包完成, 空间误差模型的构建使用spatialreg包完成。
2 结果
2.1 南岭地区哺乳类和鸟类物种丰富度空间分布格局
南岭地区每个网格(25 km2)内哺乳类的丰富度范围为57-92种, 鸟类为246-351种。哺乳类和鸟类的丰富度热点区域不同, 哺乳类主要分布在西南部而鸟类主要分布在东南部(图2; 哺乳类物种丰富度阈值为89种, 鸟类为328种)。哺乳类的物种丰富度空间分布总体自西南部山地的中心向外围递减, 东西方向的丰富度差异比南北方向的差异显著; 分布热点区域位于南岭地区西南部, 为广西和湖南交界处的都庞岭及其周边地区; 物种丰富度最低的区域为东南部平原地区, 该区距海岸线较近(图2a)。鸟类的物种丰富度空间分布格局总体自东南向西北递减, 且丰富度差异主要体现在南北方向上; 鸟类物种丰富度分布中心为南岭东南部平原, 物种丰富度最低的区域位于南岭西北部(图2b)。另外, 在南岭24°00′ N以南的宽广而平坦的平原地区, 哺乳类和鸟类的物种丰富度都比相邻丘陵和山地的要低(图2)。
图2
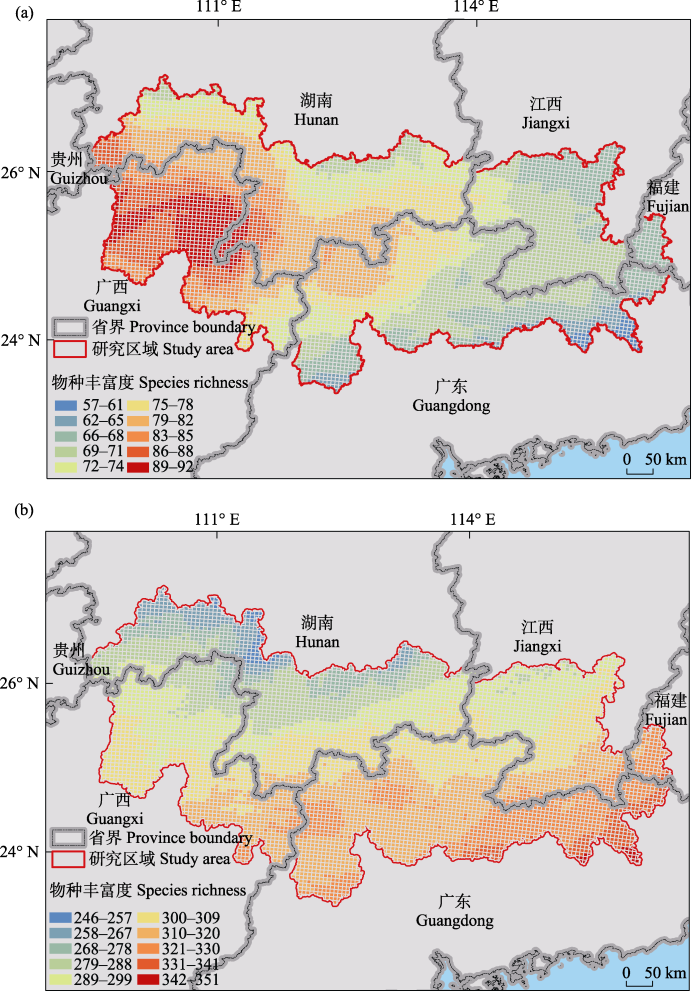
图2
南岭地区物种丰富度空间分布格局(GCS WGS 1984坐标系投影)。a: 哺乳类; b: 鸟类。
Fig. 2
Spatial distribution patterns of species richness in the Nanling Mountain Range (projected with GCS WGS 1984 coordinate system). a, Mammal; b, Bird.
2.2 南岭地区哺乳类和鸟类物种丰富度空间分布格局的影响因子
路径分析的模型拟合度良好, 其中哺乳类模型的GFI = 0.987, CFI = 0.978, IFI = 0.978; 鸟类模型的GFI = 0.997, CFI = 0.989, IFI = 0.989。各变量对南岭地区哺乳类和鸟类物种丰富度空间分布格局的直接和间接效应路径图见图3。各变量对鸟兽物种丰富度空间分布格局的总影响、直接影响和间接影响见表1。对南岭哺乳类物种丰富度空间分布格局而言, 年均温的总影响最大(总影响为-0.537, 其中直接影响为-0.424, 间接影响为-0.113), 其次分别为气温年较差(总影响为0.344, 其中直接影响为0.361, 间接影响为-0.017)、归一化植被指数(总影响为-0.306, 无间接影响)和年均降水量(总影响为0.279, 其中直接影响为0.248, 间接影响为0.031), 而海拔(总影响为-0.157, 直接影响为-0.208, 间接影响为0.051)和生境异质性(总影响为-0.124, 无间接影响)的影响相对较小, 人类足迹指数(总影响为0.056, 其中直接影响为0.017, 间接影响为0.039)的影响力最弱。对南岭鸟类物种丰富度空间分布格局而言, 气温年较差(总影响为-0.597, 其中直接影响为-0.615, 间接影响为0.018)和年均温(总影响为0.525, 其中直接影响为0.502, 间接影响为0.023)的总影响最大, 其次为海拔(总影响为0.209, 其中直接影响为0.201, 间接影响为0.008), 而年均降水量(总影响为0.089, 直接影响为0.094, 间接影响为-0.005)和生境异质性(总影响为0.047, 无间接影响)的影响相对较小, 人类足迹指数(总影响为0.031, 其中直接影响为0.019, 间接影响为0.012)和归一化植被指数(总影响为-0.030, 无间接影响)的影响力最弱。
图3
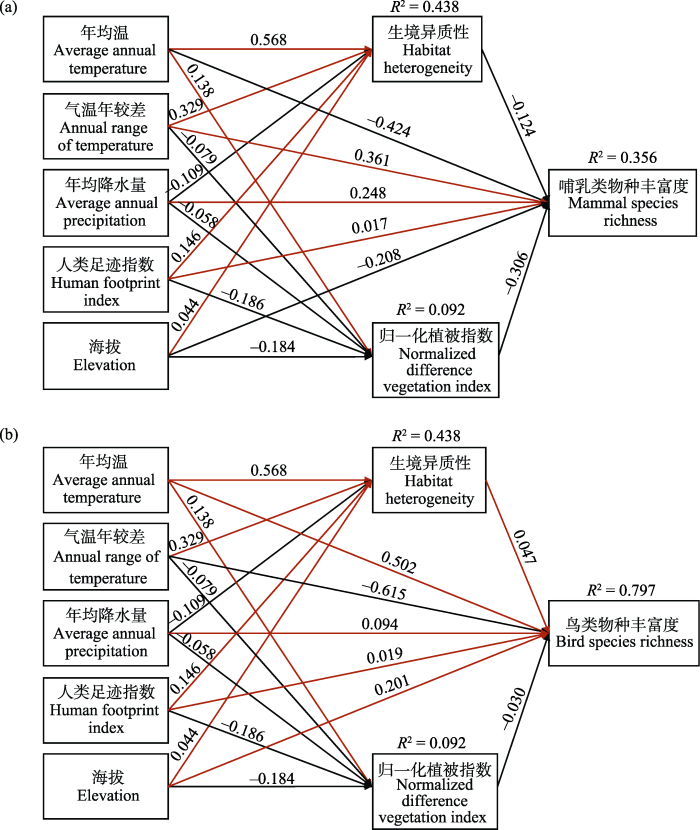
图3
各变量(年均温、气温年较差、年均降水量、人类足迹指数、海拔、生境异质性和归一化植被指数)对南岭地区物种丰富度空间分布格局的直接和间接效应路径图。a: 哺乳类; GFI = 0.987; CFI = 0.978; IFI = 0.978。b: 鸟类; GFI = 0.997; CFI = 0.989; IFI = 0.989。红色和黑色箭头分别表示正效应和负效应; 箭头边的数字表示标准化系数, 且所有系数的P值均小于0.05; 图中标明了三个模型集的R2。
Fig. 3
The path diagram of direct and indirect effects of variables (average annual temperature, annual range of temperature, average annual precipitation, human footprint index, elevation, habitat heterogeneity, normalized difference vegetation index) on the spatial distribution pattern of species richness in the Nanling region. a, Mammal; GFI = 0.987; CFI = 0.978; IFI = 0.978. b, Bird; GFI = 0.997; CFI = 0.989; IFI = 0.989. Red and black arrows indicate positive and negative effects respectively; numbers along the arrows represent standardized coefficients and the P-values of all coefficients were less than 0.05; R2 of the three model sets is given in the figure.
表1 基于路径分析的哺乳类和鸟类物种丰富度空间分布格局影响因子的结果
Table 1
| 类群 Group | 变量 Variables | 总影响 Total effect | 直接影响 Direct effect | 间接影响 Indirect effect |
|---|---|---|---|---|
| 哺乳类 Mammal | 年均温 Average annual temperature | -0.537 | -0.424 | -0.113 |
| 气温年较差 Annual range of temperature | 0.344 | 0.361 | -0.017 | |
| 年均降水量 Average annual precipitation | 0.279 | 0.248 | 0.031 | |
| 归一化植被指数 Normalized difference vegetation index | -0.306 | -0.306 | 0 | |
| 人类足迹指数 Human footprint index | 0.056 | 0.017 | 0.039 | |
| 生境异质性 Habitat heterogeneity | -0.124 | -0.124 | 0 | |
| 海拔 Elevation | -0.157 | -0.208 | 0.051 | |
| 鸟类 Bird | 年均温 Average annual temperature | 0.525 | 0.502 | 0.023 |
| 气温年较差 Annual range of temperature | -0.597 | -0.615 | 0.018 | |
| 年均降水量 Average annual precipitation | 0.089 | 0.094 | -0.005 | |
| 归一化植被指数 Normalized difference vegetation index | -0.030 | -0.030 | 0 | |
| 人类足迹指数 Human footprint index | 0.031 | 0.019 | 0.012 | |
| 生境异质性 Habitat heterogeneity | 0.047 | 0.047 | 0 | |
| 海拔 Elevation | 0.209 | 0.201 | 0.008 |
空间误差模型和路径分析的结果类似, 并且空间误差参数λ很低(λ哺乳类 < 0.01; λ鸟类< 0.01; 表2): 南岭哺乳类物种丰富度空间分布格局的主要影响因子为气温年较差(标准回归系数β = 0.138)和年均温(β = -0.096), 而生境异质性(β = -0.045)、年均降水量(β = -0.039)、归一化植被指数(β = -0.021)和海拔(β = -0.013)的影响相对较小, 人类足迹指数(β < 0.001)的影响力最弱; 南岭鸟类物种丰富度空间分布格局的主要影响因子为气温年较差(β = -0.454)和年均温(β = 0.398), 其次为年均降水量(β = 0.235), 而海拔(β = 0.037)和生境异质性(β = 0.014)的影响力相对较弱, 人类足迹指数(β = 0.006)和归一化植被指数(β = -0.001)的影响力最弱。
表2 基于空间误差模型的哺乳类和鸟类物种丰富度空间分布格局影响因子的结果
Table 2
| 类群 Group | 变量 Variables | 空间误差模型 Spatial error model | ||
|---|---|---|---|---|
| 标准估计值 Standard estimate (β) | 标准误差 Standard error | z检验概率值 Pr(>|z|) | ||
| 哺乳类 Mammal | 年均温 Average annual temperature | -0.096 | 0.009 | < 0.01 |
| 气温年较差 Annual range of temperature | 0.138 | 0.009 | < 0.01 | |
| 年均降水量 Average annual precipitation | -0.039 | 0.010 | < 0.01 | |
| 归一化植被指数 Normalized difference vegetation index | -0.021 | 0.003 | < 0.01 | |
| 人类足迹指数 Human footprint index | 0.000 | 0.003 | 0.92 | |
| 生境异质性 Habitat heterogeneity | -0.045 | 0.004 | < 0.01 | |
| 海拔 Elevation | -0.013 | 0.006 | 0.03 | |
| 鸟类 Bird | 年均温 Average annual temperature | 0.398 | 0.008 | < 0.01 |
| 气温年较差 Annual range of temperature | -0.454 | 0.009 | < 0.01 | |
| 年均降水量 Average annual precipitation | 0.235 | 0.009 | < 0.01 | |
| 归一化植被指数 Normalized difference vegetation index | -0.001 | 0.003 | 0.68 | |
| 人类足迹指数 Human footprint index | 0.006 | 0.003 | 0.03 | |
| 生境异质性 Habitat heterogeneity | 0.014 | 0.004 | < 0.01 | |
| 海拔 Elevation | 0.037 | 0.006 | < 0.01 | |
y = Xβ + ε, ε = λWε + μ, β: 标准回归系数; ε: 误差项向量; λ: 空间误差参数; W: n × n阶的空间权重矩阵; μ: 满足正态分布的随机误差向量。λ哺乳类 < 0.01; λ鸟类 < 0.01。
y = Xβ + ε, ε = λWε + μ, β, Standardized regression coefficient; ε, The error term vector; λ, The spatial error parameter; W, The spatial weight matrix of order n × n; μ, The random error vector satisfying the normal distribution. λmammal < 0.01; λbird < 0.01.
综上, 温度相关的变量(年均温和气温年较差)对于鸟兽物种空间分布格局的解释力最强, 但是年均温与哺乳类物种丰富度呈负相关, 与鸟类却呈正相关; 气温年较差与哺乳类物种丰富度呈正相关, 与鸟类却呈负相关。年均降水量、归一化植被指数、生境异质性和海拔对鸟兽物种空间分布格局的解释力相对较弱, 人类足迹指数对鸟兽物种空间分布格局的解释力最弱。
3 讨论
南岭地区哺乳类和鸟类的物种丰富度空间分布格局差异较大。哺乳类主要分布在南岭地区的西南部, 而鸟类主要分布在南岭地区的东南部。南岭地区哺乳类和鸟类的物种丰富度空间分布格局由南岭独特的地形塑造而成。由于高海拔的山体阻挡冷暖气流的交汇, 导致南岭南北两侧有约4-5℃的温差, 哺乳类和鸟类中许多物种的南界或北界止于南岭的北缘或南缘, 南岭南北的物种组成差异较大(Tian et al, 2018; 吴醇, 2020①(①吴醇 (2020) 气候变化背景下珍稀动物适宜栖息地模拟与预测——以白颈长尾雉为例. 硕士学位论文, 浙江大学, 杭州.))。比如, 权擎等(2018)通过分析南岭山脉周边区域(东部丘陵平原亚区、西部山区高原亚区、闽广沿海亚区)鸟类类群的谱系发现, 南岭山脉北侧和南侧的鸟类物种构成具有明显差异; 同时南岭山脉鸟类群落谱系结构的变化也发生在山脉的东侧和西侧之间。另外, 南岭东南部临近广东沿海地区, 而广东沿海地区位于世界重要的候鸟迁徙路线上(杨锡涛等, 2022), 这是造成南岭东南部鸟类物种丰富度更高的原因之一(候鸟种类占所有鸟类种类的54%; 附录2)。
气候因素, 特别是温度相关因子(年均温和气温年较差), 在南岭地区对哺乳类和鸟类物种丰富度空间分布格局有较大的影响。例如, 温度可以通过限制物种的生长、发育和繁殖等方面以及环境中的资源分布来影响物种的分布范围(Pearman, 2002; Bozinovic et al, 2011; Gonçalves et al, 2017; Howard et al, 2019)。然而, 年均温和气温年较差这两个主要影响因子的效应在哺乳类和鸟类中却呈现出差异。年均温与哺乳类物种丰富度呈负相关, 但与鸟类物种丰富度呈正相关。南岭是典型亚热带地区, 该区总体气候温暖湿润, 直接限制动物生存的极端气温少有出现; 如2008年极端低温冰雪灾害(张建新等, 2008)和2009年的极端干旱(郭纯青等, 2012)。最近的研究也表明温度与物种丰富度在气候温暖的区域呈负相关, 在总体寒冷的区域呈正相关, 这一偏好与动物的生理适应性和行为策略密切相关(Hu et al, 2022)。从生理适应性上看, 较高的气温可能对哺乳类造成一定程度的热应激, 导致它们需要更多的能量来维持适宜的体温, 从而增加了生存的代价(Takahashi, 2012), 而年均温相对较低的地区可能为哺乳类提供更适宜的温度范围, 减少了热应激的风险。从行为策略来看, 哺乳类在面对较高的竞争压力时, 可能更倾向于选择年均温相对较低的地区, 以减轻竞争压力(Edelman & Koprowski, 2007)。相较于哺乳类, 鸟类的移动能力更强, 可以通过飞行来躲避极端气候(Hancock & Hedrick, 2018; Webber & McGuire, 2022)。另外, 温暖的气候条件可以提供更长的繁殖季节, 也延长了食物资源的可获得时间, 这也吸引了更多的鸟类选择在年均温更高的地区筑巢和繁殖(Kawamura et al, 2019)。而且, 鸟类维持体温的能力较强, 在温暖但不过热的气候条件下不需要过多的能量投入来调节体温(Dobrovolski et al, 2012)。
气温年较差对哺乳类和鸟类物种丰富度的影响也不一致, 与哺乳类物种丰富度呈正相关, 但与鸟类物种丰富度呈负相关。对哺乳类而言, 其生理特征(如毛皮、皮肤、脂肪储备等)使它们能够在较冷的季节保持体温稳定并具有较高的体温恒定性, 或者通过冬眠来抵御极端寒冷天气, 因此可以适应较大的气温季节性变化(Li et al, 2001)。对鸟类而言, 它们更倾向于通过季节性迁徙(或短途迁徙)来适应较强烈的气温变化(Abolafya et al, 2013), 所以, 鸟类可能会表现出对气候变化更为敏感的特性。温度对于南岭哺乳类和鸟类物种丰富度空间分布格局的较大影响也说明了全球气候变化可能对南岭地区的生物产生影响, 而且值得注意的是, 这种影响对于不同生物类群的方向不一致。
南岭是重要的物种基因库和地理屏障, 加强对其哺乳类和鸟类的保护以维系区域生态功能的意义重大。本研究发现南岭哺乳类和鸟类的丰富度分布热点不同, 因此, 应在不同地区对特定的类群进行重点保护, 针对性地加强对其中哺乳类或鸟类活动的监测, 更加精准、系统地评估全球气候变暖对于哺乳类或鸟类生存的影响, 从而及时发现对哺乳类和鸟类生存不利的自然或人为因素, 并通过合理规划建设生态扩散廊道等方式减弱或消除其威胁, 保障物种丰富度各个层面的交流, 以维系该局域较高的哺乳类和鸟类物种丰富度, 并最终达到增强整个南岭地区生态功能的目的。
附录 Supplementary Material
参考文献
Using citizen science data to model the distributions of common songbirds of Turkey under different global climatic change scenarios
Geological and climatic influences on mountain biodiversity
DOI:10.1038/s41561-018-0236-z
[本文引用: 1]

Mountains are key features of the Earth's surface and host a substantial proportion of the world's species. However, the links between the evolution and distribution of biodiversity and the formation of mountains remain poorly understood. Here, we integrate multiple datasets to assess the relationships between species richness in mountains, geology and climate at global and regional scales. Specifically, we analyse how erosion, relief, soil and climate relate to the geographical distribution of terrestrial tetrapods, which include amphibians, birds and mammals. We find that centres of species richness correlate with areas of high temperatures, annual rainfall and topographic relief, supporting previous studies. We unveil additional links between mountain-building processes and biodiversity: species richness correlates with erosion rates and heterogeneity of soil types, with a varying response across continents. These additional links are prominent but under-explored, and probably relate to the interplay between surface uplift, climate change and atmospheric circulation through time. They are also influenced by the location and orientation of mountain ranges in relation to air circulation patterns, and how species diversification, dispersal and refugia respond to climate change. A better understanding of biosphere-lithosphere interactions is needed to understand the patterns and evolution of mountain biodiversity across space and time.
Biodiversity and topographic complexity: Modern and geohistorical perspectives
Physiological correlates of geographic range in animals
DOI:10.1146/ecolsys.2011.42.issue-1 URL [本文引用: 1]
Climatic history and dispersal ability explain the relative importance of turnover and nestedness components of beta diversity
DOI:10.1111/geb.2012.21.issue-2 URL [本文引用: 1]
Collinearity: A review of methods to deal with it and a simulation study evaluating their performance
DOI:10.1111/ecog.2013.36.issue-1 URL [本文引用: 1]
Characteristics and trends of climate change of Chinese subtropical Nanling Mountain
中国亚热带南岭山地气候变化特点及趋势
Communal nesting in asocial abert’s squirrels: The role of social thermoregulation and breeding strategy
DOI:10.1111/eth.2007.113.issue-2 URL [本文引用: 1]
A global perspective on tropical montane rivers
The role of mountain ranges in the diversification of birds
DOI:10.1146/ecolsys.2012.43.issue-1 URL [本文引用: 1]
Amphibian speciation rates support a general role of mountains as biodiversity pumps
DOI:10.1086/715500
PMID:34403310
[本文引用: 1]
Continental mountain areas cover 80% of global terrestrial diversity. One prominent hypothesis to explain this pattern proposes that high mountain diversities could be explained by higher diversification rates in regions of high topographic complexity (HTC). While high speciation in mountains has been detected for particular clades and regions, the global extent to which lineages experience faster speciation in mountains remains unknown. Here we addressed this issue using amphibians as a model system (>7,000 species), and we found that families showing high speciation rates contain a high proportion of species distributed in mountains. Moreover, we found that lineages inhabiting areas of HTC speciate faster than lineages occupying areas that are topographically less complex. When comparing across regions, we identified the same pattern in five biogeographical realms where higher speciation rates are associated with higher levels of complex topography. Low-magnitude differences in speciation rates between some low and high complex topographies suggest that high mountain diversity is also affected by low extinction and/or high colonization rates. Nevertheless, our results bolster the importance of mountains as engines of speciation at different geographical scales and highlight their importance for the conservation of global biodiversity.
The relationship between bird distribution patterns and environmental factors in an ecotone area of northeast Brazil
DOI:10.1016/j.jaridenv.2017.01.004 URL [本文引用: 1]
Drought causes and countermeasures of China southwest karst area from 2009 to 2010—A case in karst area of Guangxi
2009-2010年中国西南岩溶区旱情分析与减灾对策——以广西岩溶区为例
为研究中国西南岩溶区抗旱减灾对策,分析了2009年秋至2010年春广西岩溶区的旱情:与常年同期相比,大部分地区降水量偏少20% ~70%;气温除11月和4月偏低外,其余7个月偏高0.7~2.8℃;一些地方持续8个月无有效降雨;岩溶区农作物受旱面积约为100万hm2,约占岩溶区耕地面积的70%,干旱分布与岩溶发育区具有高度的一致性。极端气候是诱发西南岩溶区大旱灾的主要原因,岩溶区水文地质和地貌条件是旱情加剧的独特因素,人为因素造成的工程性缺水和环境恶化也是主要因素之一。提出了加强旱情监测、预报、预警和应急能力,加大水利工程建设力度,加强水利工程后期管理以及维修养护,重视岩溶生态恢复、重建和保护等抗旱减灾对策。
Physiological vagility affects population genetic structure and dispersal and enables migratory capacity in vertebrates
Energy, water, and broad-scale geographic patterns of species richness
DOI:10.1890/03-8006 URL [本文引用: 1]
Quantifying the effects of climate and anthropogenic change on regional species loss in China
The usefulness of elevation as a predictor variable in species distribution modelling
DOI:10.1016/j.ecolmodel.2012.07.028 URL [本文引用: 1]
What drives at-risk species richness? Environmental factors are more influential than anthropogenic factors or biological traits
Precipitation drives species accumulation whereas temperature drives species decline in Himalayan vertebrates
DOI:10.1111/jbi.v49.12 URL [本文引用: 1]
Analysis of bird diversity in Chebaling Nature Reserve
广东车八岭自然保护区鸟类多样性分析
When should species richness be energy limited, and how would we know?
DOI:10.1111/ele.12240
PMID:24393362
[本文引用: 1]
Energetic constraints are fundamental to ecology and evolution, and empirical relationships between species richness and estimates of available energy (i.e. resources) have led some to suggest that richness is energetically constrained. However, the mechanism linking energy with richness is rarely specified and predictions of secondary patterns consistent with energy-constrained richness are lacking. Here, we lay out the necessary and sufficient assumptions of a causal relationship linking energy gradients to richness gradients. We then describe an eco-evolutionary simulation model that combines spatially explicit diversification with trait evolution, resource availability and assemblage-level carrying capacities. Our model identified patterns in richness and phylogenetic structure expected when a spatial gradient in energy availability determines the number of individuals supported in a given area. A comparison to patterns under alternative scenarios, in which fundamental assumptions behind energetic explanations were violated, revealed patterns that are useful for evaluating the importance of energetic constraints in empirical systems. We use a data set on rockfish (genus Sebastes) from the northeastern Pacific to show how empirical data can be coupled with model predictions to evaluate the role of energetic constraints in generating observed richness gradients. © 2014 John Wiley & Sons Ltd/CNRS.
Seasonality in spatial distribution: Climate and land use have contrasting effects on the species richness of breeding and wintering birds
DOI:10.1002/ece3.2019.9.issue-13 URL [本文引用: 1]
Biodiversity in ecosystems:Some principles
In: Tasks for Vegetation Science (ed. Kratochwil A), pp. 5-38. Springer Netherlands, Dordrecht.
Cold adaptive thermogenesis in small mammals from different geographical zones of China
Plant diversity and its conservation values of Guangdong Nanling National Park
广东南岭国家公园植物多样性及其保护价值
The scale of community structure: Habitat variation and avian guilds in tropical forest understory
DOI:10.1890/0012-9615(2002)072[0019:TSOCSH]2.0.CO;2 URL [本文引用: 1]
Preliminary investigation on mammal in Huangniushi NR in Lianping County
广东连平黄牛石保护区兽类初步调查
β-diversity of bird assemblages in the Nanling Mountain ranges
DOI:10.13284/j.cnki.rddl.003047
[本文引用: 1]
How to divide the world into biogeographical regions is a long concerned issue and recently intrigues the research interest because rich data about phylogeny and traits of taxa has been gained nowadays. β-diversity patterns was applied, based on 46 places located in 3 subregions (one belongs to South China and two belong to Central China) of Chinese animal geography, to re-analyze the robustness of setting Nanling Mountain Ranges as part of the boundaries between Central and South China. The results revealed that: 1) the taxonomic and phylogenetic dissimilarities among the three subregions were small and dissimilarities related to nestedness and species turnover had varied contributions to the distances between assemblages of three subregions; 2) resident birds, summer breeders and wintering birds affected the β diversity patterns differently; the dissimilarities were strong when analyzing the assemblages of resident birds and they became weaker when counting the summer and winter visitors; 3) in the eastern-western direction remarkable phylogenetic distance happened between places far from each other; in the northern-southern direction great phylogenetic distance happened between far and neighboring locations. The results above indicated that the phylogenetic structures of bird assemblages changed beyond expectations from south to north side of the Nanling Mountains, which suggests that it is reasonable to treat the Nanling Mountain ranges as part of the border between Central and South China of animal geography.
南岭山脉及周边鸟类β多样性分析
Building mountain biodiversity: Geological and evolutionary processes
DOI:10.1126/science.aax0151
URL
[本文引用: 1]
Mountain regions are unusually biodiverse, with rich aggregations of small-ranged species that form centers of endemism. Mountains play an array of roles for Earth’s biodiversity and affect neighboring lowlands through biotic interchange, changes in regional climate, and nutrient runoff. The high biodiversity of certain mountains reflects the interplay of multiple evolutionary mechanisms: enhanced speciation rates with distinct opportunities for coexistence and persistence of lineages, shaped by long-term climatic changes interacting with topographically dynamic landscapes. High diversity in most tropical mountains is tightly linked to bedrock geology—notably, areas comprising mafic and ultramafic lithologies, rock types rich in magnesium and poor in phosphate that present special requirements for plant physiology. Mountain biodiversity bears the signature of deep-time evolutionary and ecological processes, a history well worth preserving.
Humboldt’s enigma: What causes global patterns of mountain biodiversity?
DOI:10.1126/science.aax0149
URL
[本文引用: 1]
Mountains contribute disproportionately to the terrestrial biodiversity of Earth, especially in the tropics, where they host hotspots of extraordinary and puzzling richness. With about 25% of all land area, mountain regions are home to more than 85% of the world’s species of amphibians, birds, and mammals, many entirely restricted to mountains. Biodiversity varies markedly among these regions. Together with the extreme species richness of some tropical mountains, this variation has proven challenging to explain under traditional climatic hypotheses. However, the complex climatic characteristics of rugged mountain regions differ fundamentally from those of lowland regions, likely playing a key role in generating and maintaining diversity. With ongoing global changes in climate and land use, the role of mountains as refugia for biodiversity may well come under threat.
Modeling the ecology and evolution of biodiversity: Biogeographical cradles, museums, and graves
Differential effects of environmental heterogeneity on global mammal species richness
DOI:10.1111/geb.2015.24.issue-9 URL [本文引用: 1]
Terminology and quantification of environmental heterogeneity in species-richness research
DOI:10.1111/brv.2015.90.issue-3 URL [本文引用: 1]
Heat stress on reproductive function and fertility in mammals
DOI:10.1007/s12522-011-0105-6
PMID:29699104
[本文引用: 1]
In most mammalian species including cattle, heat stress has deleterious effects on nutritional, physiological and reproductive functions. Exposure of animals to a hot environment causes an increase in body temperature in mammals, including domestic animals. High ambient temperature also causes a decrease in the length and intensity of estrus by disturbing ovarian function as well as decreasing pregnancy rate after artificial insemination. Therefore, it is important to understand the effects of heat stress on reproductive function in order to improve the production of domestic animals. Heat stress decreases appetite, weight gain, and milk yield in dairy cattle. It also adversely affects the reproductive performance of both sexes. In males, it reduces spermatogenic activity, while in females it adversely impacts oogenesis, oocyte maturation, fertilization development and implantation rate. Detection and evaluation of the deteriorating effects of heat stress on reproductive organs and cells can help to design measures to prevent them and improve reproductive functions. In this review, we discuss the impacts of heat stress on reproductive functions.
Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures
DOI:10.1046/j.0305-0270.2003.00994.x
URL
[本文引用: 1]
Aim In a selected literature survey we reviewed studies on the habitat heterogeneity–animal species diversity relationship and evaluated whether there are uncertainties and biases in its empirical support.
Phylogeography of Eomecon chionantha in subtropical China: The dual roles of the Nanling Mountains as a glacial refugium and a dispersal corridor
DOI:10.1186/s12862-017-1093-x [本文引用: 1]
Predicting range shifts of northern bird species: Influence of modelling technique and topography
DOI:10.1016/j.actao.2010.01.006 URL [本文引用: 1]
Heterothermy, body size, and locomotion as ecological predictors of migration in mammals
DOI:10.1111/mam.v52.1 URL [本文引用: 1]
The essence of testing structural equation models using popular fit indexes
结构方程模型常用拟合指数检验的实质
Factors responsible for forest and water bird distributions in rivers and lakes along an urban gradient in Beijing
DOI:10.1016/j.scitotenv.2020.139308 URL [本文引用: 1]
Distribution pattern of plant community species diversity along altitudinal gradients in Nanling Mountains, Guangdong
广东南岭植物群落物种多样性沿海拔梯度分布格局
Application of a species dispersal model to guide the establishment of a functional ecological corridor for Platalea minor in the coastal areas of Guangdong
利用物种扩散模型构建广东沿海地区黑脸琵鹭的功能性生态廊道
Effects of ice and snow disasters on mammals, reptiles and amphibians in Nanling, Guangdong Province
In: Symposium of the 5th Zoological Seminar of Guangdong, Hunan, Jiangxi, and Hubei Provinces. Guangdong Zoological Society, Guangzhou. (in Chinese with English abstract)
低温冰雪灾害对广东南岭灾区兽类和两爬动物生态影响
见: 第五届广东、湖南、江西、湖北四省动物学学术研讨会论文摘要汇编. 广东省动物学会, 广州.]
Spatiotemporal dynamics of NDVI and land use in China based on remote sensing images
Planning and construction strategy in priority area of biodiversity conservation in Nanling Mountain of Guangdong
广东南岭生物多样性保护优先区域规划建设策略
Analysis of temporal and spatial variation of rainfall in 1968-2015 in Nanling
南岭山地1968到2015年降雨的时空变化特征研究
Temporal patterns of three sympatric pheasant species in the Nanling Mountains: N-mixture modeling applied to detect abundance
DOI:10.1186/s40657-019-0181-6
[本文引用: 1]
The reliability of long-term population estimates is crucial for conservation and management purposes. Most previous studies assume that count indices are proportionally related to abundance; however, this assumption may not hold when detection varies spatially and temporally. We examined seasonal variations in abundance of three bird species (Cabot’s Tragopan Tragopan caboti, Silver Pheasant Lophura nycthemera, and White-necklaced Partridge Arborophila gingica) along an elevational gradient, using N-mixture models that take into account imperfect detection in our bird data.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}