


蛾类种类繁多(Ober & Hayes, 2009), 对微环境敏感(Summerville, 2011), 在食物链中具有承上启下的地位(Holmes et al, 1979; Diaz et al, 2020), 经常作为监测森林生态效益的指示类群(Kitching et al, 2000)。当前人工林中的蛾类多样性研究大多基于整体环境(郭欣乐等, 2022)或单一林分(任雪毓等, 2022)。人工林内存在大量的边缘, 但目前针对其生物多样性特点的研究则较少。蛾类作为生物多样性研究的常见类群, 一定程度上可以反映人工林斑块边缘的生态效益。本研究以蛾类为研究对象, 基于4种常见斑块树种组合, 运用自制灯诱装置, 通过比较蛾类群落与多样性特征, 探究了人工林斑块边缘效应的生物多样性表现, 以期为人工林的管理与后续平原造林计划提供依据。
1 研究方法
1.1 研究区概况
本研究地点位于北京市通州区潞城集体林场(39°48′-39°51′ N, 116°47′-116°53′ E), 地处北京市东南部。林场树种组成以华北地区常见造林树种为主, 包括国槐(Styphnolobium japonicum)、毛白杨(Populus tomentosa)、栾树(Koelreuteria paniculata)、元宝枫(Acer truncatum)、桧柏(Juniperus chinensis)、旱柳(Salix matsudana)、杜仲(Eucommia ulmoides)等, 不同纯林斑块呈现不规则镶嵌状排列。
1.2 样地设置
选取该地区常见的4类树种组合进行试验研究: 毛白杨林-国槐林、国槐林-栾树林、旱柳林-杜仲林、国槐林-桧柏林, 各树种斑块宽度均大于70 m,边缘长度在100 m以上。样地概况见表1。
表1 人工林斑块蛾类调查样地概况(平均值 ± SD)
Table 1
| 平均树高 Average tree height (m) | 平均胸径 Average diameter at breast height (cm) | 株行距 Distance between trees (m) | 林下植被高度 Height of understory vegetation (m) | 林下植被盖度 Coverage of understory vegetation (%) | |
|---|---|---|---|---|---|
| 毛白杨林 Populus tomentosa forest (mby) | 13.53 ± 1.05 | 20.00 ± 1.63 | 4 × 4 | 0.18 ± 0.11 | 3.00 ± 1.41b |
| 毛白杨林-国槐林1边缘 Edge of Populus tomentosa forest & Styphnolobium japonicum forest 1 (Eyh) | - | - | - | 0.28 ± 0.10 | 53.89 ± 20.65a |
| 国槐林1 Styphnolobium japonicum forest 1 (gh1) | 9.17 ± 0.62 | 32.67 ± 2.05 | 4 × 4 | 0.31 ± 0.24 | 48.00 ± 21.02ab |
| 栾树林 Koelreuteria paniculata forest (ls) | 6.83 ± 0.85 | 17.67 ± 2.05 | 3.5 × 3.5 | 0.18 ± 0.19 | 20.67 ± 24.99b |
| 栾树林-国槐林2边缘 Edge of Koelreuteria paniculata forest & Styphnolobium japonicum forest 2 (Elh) | - | - | - | 0.28 ± 0.35 | 50.33 ± 27.19a |
| 国槐林2 Styphnolobium japonicum forest 2 (gh2) | 8.67 ± 0.47 | 21.33 ± 2.62 | 4 × 4.5 | 0.16 ± 0.09 | 59.22 ± 32.26a |
| 桧柏林-国槐林2边缘 Edge of Juniperus chinensis forest & Styphnolobium japonicum forest 2 (Ebh) | - | - | - | 0.35 ± 0.20 | 48.78 ± 29.18a |
| 桧柏林 Juniperus chinensis forest (gb) | 3.17 ± 0.24 | 13.00 ± 2.16 | 3 × 3 | 0.28 ± 0.13 | 64.44 ± 27.33a |
| 旱柳林 Salix matsudana forest (hl) | 12.00 ± 0.82 | 22.67 ± 2.05 | 4 × 4 | 0.35 ± 0.15 | 61.11 ± 12.86a |
| 旱柳林-杜仲林边缘 Edge of Salix matsudana forest & Eucommia ulmoides forest (Ehd) | - | - | - | 0.19 ± 0.07 | 51.78 ± 24.89a |
| 杜仲林 Eucommia ulmoides forest (dz) | 8.33 ± 0.47 | 25.00 ± 4.08 | 3.5 × 3.5 | 0.18 ± 0.06 | 24.22 ± 21.26b |
国槐林1和国槐林2为分开的两块不同样地, 间距300 m; 不同字母表示不同样地间差异显著。
Styphnolobium japonicum forest 1 and Styphnolobium japonicum forest 2 are two different plots; Different letters represent significant differences.
1.3 样本采集
图1
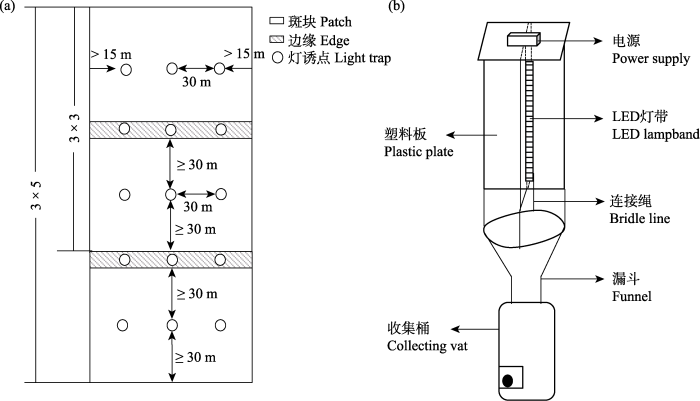
图1
人工林斑块蛾类调查灯诱点位(a)及灯诱装置(b)设置图
Fig. 1
Schematic diagram of the sample setting (a) and light trap (b) of the plantation patches moth survey
图2

图2
毛白杨林-国槐林1样地情况(a)及灯诱装置实物图(b, c)
Fig. 2
Schematic representation of Populus tomentosa forest & Styphnolobium japonicum forest 1 and its edge (a) and diagram of light trap (b, c)
本研究在2022年6-8月进行, 每月调查1次, 分别在每个树种组合的纯林斑块内及斑块边缘设置3个灯诱点。每组样地类型灯诱两晚, 以保障采集数量。针对样地具体情况, 采取3 × 3或3 × 5的排列方式, 以间距30 m设置灯诱样点(图1a)。灯诱时间为每晚的19:30-22:30, 采集到的蛾类样品于第2天进行分类整理。林下植被特征因子调查以灯诱点为中心设置1 m × 1 m的样方进行, 主要调查植被的平均盖度(Vc)和平均高度(Vh), 并测量环境温度、湿度。
1.4 数据分析
(1)采集数量: 统计不同斑块蛾类的个体数N和物种数S。(2)物种组成: Venn图可表示不同斑块的蛾类物种组成; 使用非度量多维尺度(NMDS)排序, 选择Bray-Curtis距离可视化物种组成, 通过相似性分析(Anosim)比较组内及组间的群落物种组成差异。(3)物种多样性: 优势度使用Simpson指数测度: C = ∑Pi2 (i = 1, 2, 3, ……, S); 多样性使用Shannon-Wiener多样性指数测度: H′ = −∑PilnPi (i = 1, 2, 3, ……, S); 丰富度使用Margalef丰富度指数测度: D = (S - 1)/lnN; 均匀度使用Pielou均匀度指数测度: J = H′/lnS; 其中Pi = ni/N, ni为物种i的个体数。(4)林下植被特征因子关联性分析: 使用Pearson相关性分析检验植被特征因子与蛾类特征数据的相互关系。(5)边缘效应强度: 使用边缘效应指数(王伯荪和彭少麟, 1986)计算边缘效应强度: IH′ = mH边′/ ∑H边i′, IC = mC边/∑C边i, H边′为边缘群落的蛾类多样性指数, H边i′为各个群落的蛾类多样性指数, C边为边缘群落的蛾类优势度指数, C边i为各个群落的蛾类优势度指数, m为组成边缘的群落数。IH′值越大则边缘效应强度越强, 当IH′值大于1时边缘效应呈正效应, 小于1则呈负效应; IC值越低则边缘效应强度越高。
采用单因素方差分析(one-way ANOVA)比较不同纯林斑块及其边缘人工林内的昆虫群落的个体数量及多样性指数, 显著性水平为0.05。
所有数据分析在R 4.2.1、SPSS 26.0以及PAST (Hammer et al, 2001)软件中完成, 使用R 4.2.1和Origin 2023软件完成作图。
2 结果
2.1 蛾类物种数和个体数量
本研究共采集到蛾类4,428只, 分属于24科144种, 其中有77.1%的个体鉴定到种, 其余鉴定到科。物种数上, 夜蛾科、螟蛾科为优势类群, 共占总物种数的25.7%; 个体数量上, 夜蛾科为优势类群, 占总个体数的49.1%, 其次为尺蛾科, 占14.8% (表2)。
表2 人工林斑块及边缘蛾类个体数量
Table 2
| 科 Family | mby | Eyh | gh1 | ls | Elh | gh2 | Ebh | gb | hl | Ehd | dz |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 夜蛾科 Noctuidae | 181 | 109 | 122 | 298 | 239 | 117 | 99 | 203 | 240 | 361 | 204 |
| 尺蛾科 Geometridae | 68 | 52 | 122 | 79 | 87 | 85 | 71 | 23 | 13 | 30 | 23 |
| 螟蛾科 Pyralidae | 21 | 30 | 59 | 100 | 106 | 72 | 38 | 40 | 33 | 62 | 67 |
| 菜蛾科 Plutellidae | 12 | 6 | 15 | 23 | 26 | 18 | 26 | 47 | 9 | 18 | 15 |
| 卷蛾科 Tortricidae | 4 | 21 | 8 | 17 | 24 | 30 | 16 | 23 | 22 | 22 | 16 |
| 织蛾科 Oecophoridae | 1 | 3 | 5 | 14 | 11 | 16 | 15 | 17 | 21 | 23 | 10 |
| 麦蛾科 Gelechiidae | 7 | 5 | 10 | 18 | 13 | 2 | 5 | 6 | 28 | 13 | 10 |
| 草螟科 Crambidae | 2 | 13 | 10 | 11 | 15 | 9 | 8 | 4 | 6 | 24 | 13 |
| 舟蛾科 Notodontidae | 6 | 5 | 2 | 1 | 1 | 4 | 2 | - | 3 | 14 | 6 |
| 瘤蛾科 Nolidae | - | - | 1 | - | 1 | - | 1 | - | 7 | 18 | 12 |
| 其他 Others | 1 | 7 | 3 | 17 | 14 | 13 | 7 | 16 | 4 | 12 | 10 |
| 合计 Total | 303 | 251 | 357 | 578 | 537 | 366 | 288 | 379 | 386 | 597 | 386 |
缩写含义见
The abbreviations see
物种数上, 毛白杨林-国槐林1边缘、栾树林-国槐林2边缘、桧柏林-国槐林2边缘、旱柳林-杜仲林边缘的物种数均位于两种纯林斑块之间或略高于斑块内, 仅旱柳林-杜仲林边缘显著高于旱柳林斑块; 在个体数量上, 旱柳林-杜仲林边缘高于纯林斑块内且差异显著, 其余边缘均位于纯林斑块之间或略低于斑块内(图3)。在4种边缘类型中, 个体数量及物种数均表现为旱柳林-杜仲林边缘 > 栾树林-国槐林2边缘 > 桧柏林-国槐林2边缘 > 毛白杨林-国槐林1边缘, 旱柳林-杜仲林边缘的个体总数(199 ± 30)及物种数(45 ± 4)均大于其余边缘, 与桧柏林-国槐林2边缘和毛白杨林-国槐林1边缘存在显著差异(P < 0.05)。
图3
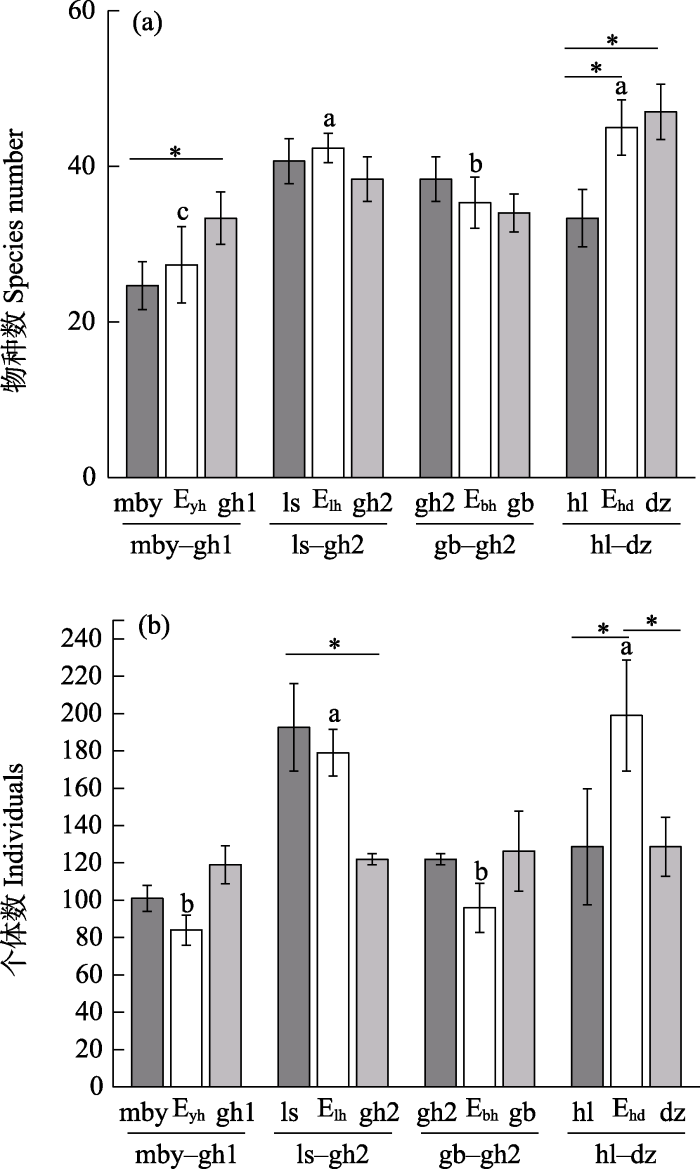
图3
人工林斑块及边缘蛾类物种数(a)和个体数(b) (平均值 ± SD)。*代表斑块与边缘间差异显著; 不同小写字母表示不同边缘间差异显著。缩写含义见
Fig. 3
Moth species number (a) and individuals (b) of plantation patches and edges (mean ± SD). * represents significant difference, different lowercase letters represent significant differences between different edges. The abbreviations see
2.2 蛾类物种组成及群落结构
桧柏林-国槐林2斑块及其边缘中共采集蛾类96种, 有14种仅在边缘出现; 桧柏林-国槐林2边缘与国槐林和桧柏林共有的种类分别占相应斑块的61.5%和72.2%。栾树林-国槐林2斑块及其边缘中共采集蛾类101种, 有16种仅在边缘出现; 栾树林-国槐林2边缘与栾树林和国槐林共有的种类分别占相应斑块的73.1%和67.7%。毛白杨林-国槐林1斑块及其边缘共采集蛾类74种, 有13种仅在边缘出现, 毛白杨林-国槐林1边缘与毛白杨林和国槐林共有的种类分别占相应斑块的75.0%和68.6%。旱柳林-杜仲林斑块及其边缘共采集蛾类109种, 其中有16种仅在边缘出现; 旱柳林-杜仲林边缘与旱柳林和杜仲林共有的种类分别占相应斑块的77.6%和64.9% (图4)。
图4
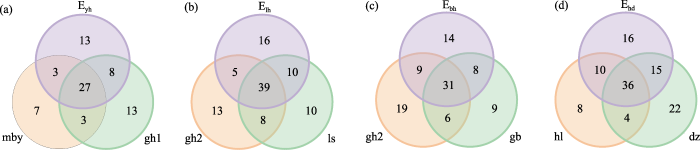
图4
人工林斑块及边缘蛾类物种数的Venn图。缩写含义见
Fig. 4
Venn diagram of moth species numbers in plantation patches and edges. The abbreviations see
图5
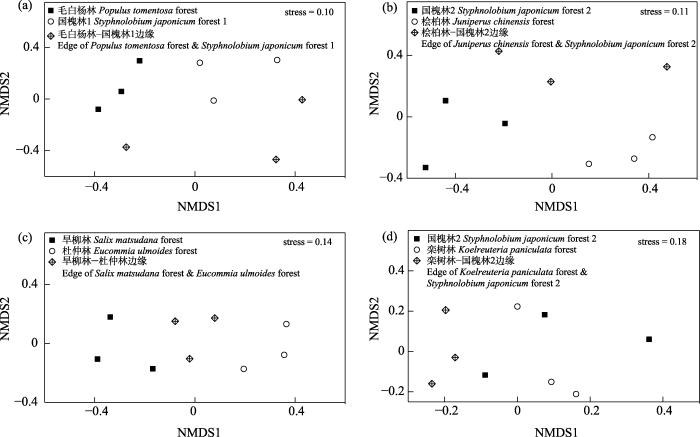
图5
人工林斑块及边缘的蛾类群落非度量多维尺度(NMDS)排序图
Fig. 5
Ordination plot of non-metric multidimensional scaling (NMDS) of moth communities in plantation patches and edge
图6
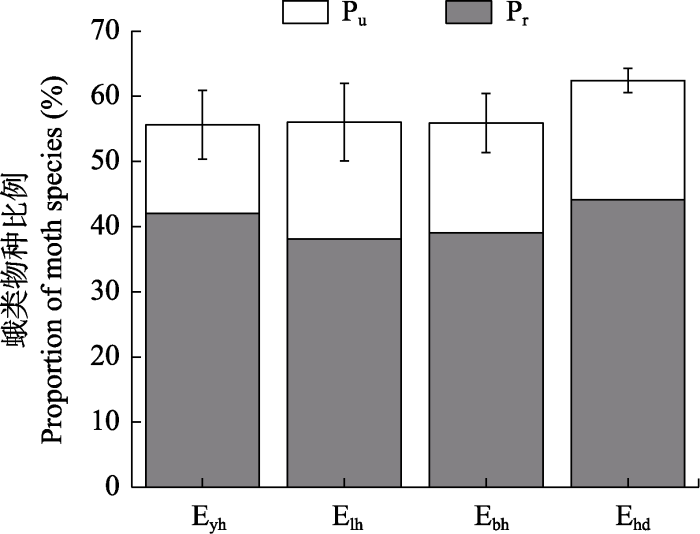
图6
人工林边缘类型蛾类物种占其所属样地物种数比例 (平均值 ± SD)。Pu代表独有种比例; Pr为剩余物种比例。缩写含义见
Fig. 6
Proportion of moth species of plantation edge types in the number of species in the plot (mean ± SD). Pu represents the proportion of unique species; Pr represents the proportion of the remaining species. The abbreviations see
2.3 蛾类多样性指数
4种边缘类型的Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数均位于两种纯林斑块之间或略高于斑块内; Simpson指数除毛白杨林-国槐林1边缘均位于两种纯林斑块之间; 桧柏林-国槐林2边缘及毛白杨林-国槐林1边缘的Pielou均匀度指数均高于斑块内。旱柳林-杜仲林边缘的Simpson指数显著高于杜仲林斑块; Shannon-Wiener多样性指数、Margalef丰富度指数显著高于旱柳林斑块; Pielou均匀度指数显著低于杜仲林斑块(图7)。
图7
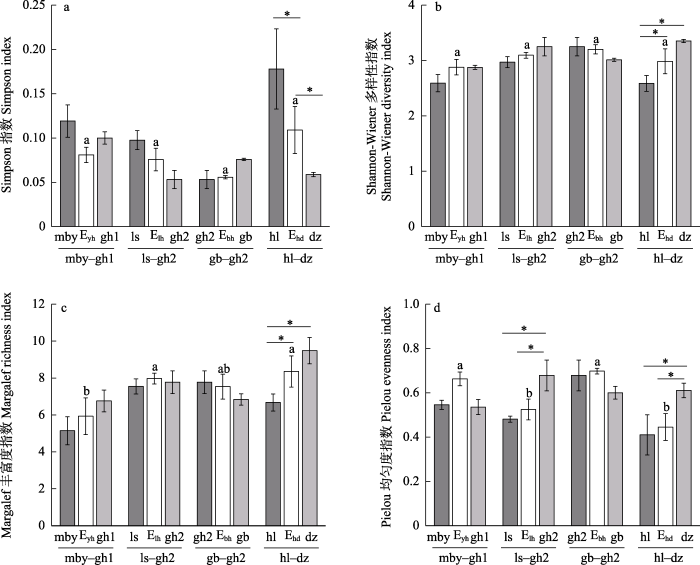
图7
人工林斑块及边缘物种多样性指数(平均值 ± SD)。*代表斑块与边缘间差异显著; 不同小写字母表示不同边缘间差异显著。缩写含义见
Fig. 7
The species diversity index of plantation patches and edges (mean ± SD). * represents significant difference, different lowercase letters represent significant differences between different edges. The abbreviations see
4种边缘类型中, 旱柳林-杜仲林边缘的Margalef丰富度指数、Simpson指数最高, 但Pielou均匀度指数和Shannon-Wiener多样性指数最低, 表明旱柳林-杜仲林边缘的蛾类群落物种丰富度较大但优势类群较为集中, 从而该群落呈现出了多样性降低的特点。旱柳林-杜仲林边缘与其余边缘相比存在优势物种较多的现象, 本研究发现该物种为委夜蛾(Athetis furvula) (附录1)。
2.4 蛾类群落组成与植被特征因子关联性
图8
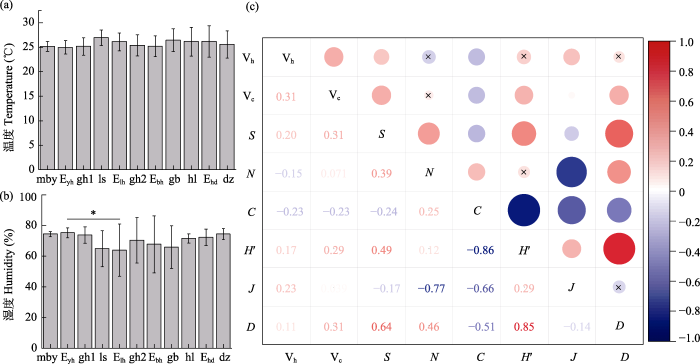
图8
蛾类特征指数与环境因子相关性分析。(a)温度(平均值 ± SD); (b)湿度(平均值 ± SD); (c)蛾类特征指数与植被特征因子相关性分析图, 圆圈大小代表相关性强度, “×”表示不存在显著相关性。Vh: 林下植被平均高度; Vc: 林下植被平均盖度; S: 物种数; N: 个体数; C: Simpson指数; H′: Shannon-Wiener多样性指数; J: Pielou均匀度指数; D: Margalef丰富度指数。其余缩写含义见
Fig. 8
Correlation analysis between moth characteristic index and environmental factors. (a) Temperature (mean ± SD); (b) Humidity (mean ± SD); (c) The plot of correlation analysis between moth characteristic index and vegetation characteristic factors. The circle size represents the correlation strength, “×” represents no significant correlation. Vh, Height of understory vegetation; Vc, Coverage of understory vegetation; S, Species numbers; N, Individuals; C, Simpson index; H′, Shannon-Wiener diversity index; J, Pielou evenness index; D, Margalef richness index. Other abbreviations see
蛾类Simpson指数与植被的平均高度和平均盖度呈显著负相关关系; 蛾类物种数与植被的平均高度和平均盖度呈显著正相关关系。除此之外, 蛾类Pielou均匀度指数与植被的平均高度呈显著正相关关系; Shannon-Wiener多样性指数、Margalef丰富度指数与植被的平均盖度呈显著正相关关系(图8c)。
2.5 边缘效应强度
不同边缘的边缘效应指数IH′值均大于1, 表明所选取的4种边缘类型均存在正向的边缘效应, 即边缘的物种丰富度和多样性均大于相邻斑块。边缘效应指数IC值的比较结果为旱柳林-杜仲林边缘 > 毛白杨林-国槐林1边缘 > 栾树林-国槐林2边缘 > 桧柏林-国槐林2边缘, 表明旱柳林-杜仲林边缘效应强度最低(表3)。
表3 不同人工林边缘类型的边缘效应指数
Table 3
| 边缘效应强度 Edge effect intensity | 毛白杨林-国槐林1边缘 Eyh | 栾树林-国槐林2边缘 Elh | 桧柏林-国槐林2边缘 Ebh | 旱柳林-杜仲林边缘 Ehd |
|---|---|---|---|---|
| IH’ | 1.102593793 | 1.078513732 | 1.105394079 | 1.080308277 |
| Ic | 0.862558656 | 0.854916806 | 0.771192529 | 0.915185808 |
3 讨论
边缘处于两种斑块的过渡地带, 起到连通不同林分中生物的作用(Niemelä et al, 2007)。森林中存在部分主要植物物种, 它们的出现会导致蛾类数量成倍的增加(Narango et al, 2020), 边缘会为这些植物提供生存地点, 这可能与边缘存在一定的光照有关, 该现象同样发生在纯林的林窗中(陈博等, 2021)。当前多数关于人工林多样性的研究发现, 边缘植被多样性相对于林内更高(李新荣和Павлов, 1999; 于顺利等, 2000; 高俊峰和张芸香, 2005; 王德艺等, 2016)。边缘更加开阔多样的环境增加了植食性昆虫对植物的利用率, 从而影响到昆虫群落(Altamirano et al, 2016), 例如访花昆虫(Korpela et al, 2015)以及刺吸式昆虫(de Araujo & do Espirito-Santo Filho, 2012)的增加。Pinksen等(2021)的研究发现, 在砍伐后的林地边缘蛾类的丰富度并没有显著增加, 但出现了一定的独有种。本研究中采集到的大部分蛾类物种为人工林斑块与人工林斑块边缘共有, 但边缘也存在一定的独有种, 其个体数量、物种数量有时甚至大于斑块内, 人工林斑块内的物种组成与边缘存在差异。边缘独有种占其斑块物种数的比例为16.7% ± 1.9%, 且4种边缘类型之间差异不显著, 表明不同的边缘类型虽独有种不同, 但仍为其所属样地提供了新的多样性基础, 在原有的斑块组成上, 边缘的出现提高了其所属样地的生物多样性。
本研究中, 不同类型的人工林斑块边缘的蛾类群落存在不同的特点。旱柳林-杜仲林边缘的蛾类物种数、个体数量均大于纯林斑块, 且在4种边缘类型中最高; 旱柳林-杜仲林边缘蛾类物种多样性指数和物种组成与纯林斑块内存在显著性差异, 表明旱柳林-杜仲林边缘与其他3类边缘相比与纯林斑块的蛾类群落差异更大。4种边缘类型中, 旱柳林-杜仲林边缘的Pielou均匀度指数最低但Simpson指数最高, 与其他边缘相比其优势种更为集中, 这表明旱柳林-杜仲林边缘造成了部分优势物种的增加从而导致其生物多样性降低。此外, 旱柳林-杜仲林边缘的边缘效应强度最低, 进一步表明其边缘的存在导致的生物多样性提升最低。人工林边缘的出现对昆虫多样性效益的提升及生态系统的稳定均起到了一定的正向作用, 但同时也会带来优势物种暴发的风险。
影响蛾类物种组成的因素包括: 寄主植物的密度(Luque et al, 2007)、冠层、林龄(Ober & Hayes, 2009)、植被密度(Highland et al, 2013)等。边缘环境与林分内部存在一定的差异, 例如温度和光照的增加(Pinzon et al, 2021)等, 且边缘的空旷性会导致蛾类更易被捕食者发现, 这可能会给部分蛾类带来被捕食的危机(Zuk & Kolluru, 1998), 从而进一步影响蛾类群落分布。在本研究中, 边缘的温度虽未与斑块内呈现明显的相关性, 但大部分边缘温度仍处于相邻的纯林斑块之间, 边缘在人工林生态系统中起到了一定的过渡作用。植被平均盖度在不同斑块内差异显著, 且植被盖度与蛾类丰富度存在显著的正相关关系, 相似的结论在甲虫类昆虫中也有体现(Magura, 2002; 刘继亮等, 2021)。本研究发现部分边缘的植被平均高度高于林地内部, 表现出明显的边缘差异, 且蛾类Simpson指数与植被高度呈显著的负相关关系, Pielou均匀度指数与植被高度存在显著的正相关关系, 且旱柳林-杜仲林边缘的植被平均高度最低。部分研究发现某些寡食性蛾类幼虫的生长主要依赖寄主植物, 与多食性蛾类相比, 它们更喜欢高度较低的植被(Pöyry et al, 2006)。因此, 在人工林林分管理上, 适当减少林地杂草割除, 保留一定的高植被, 可防止寡食性蛾类的大幅度增加, 这在一定程度上会提升其带来的生物多样性效益, 更有利于当地生态系统的稳定。
边缘效应一直是生态学讨论研究的问题, 本研究结果表明, 斑块状人工林边缘的出现一定程度上提高了当地的蛾类物种多样性, 但不同树种组合斑块的边缘效应所呈现的生物多样性特点不同, 可能存在优势物种暴发的风险。在昆虫数量减少的大环境下(Hallmann et al, 2017), 如何恢复和提高人工林昆虫群落的多样性和稳定性成为一个值得考虑的问题。因此, 选择适宜的块状组合林分, 减少植被割除, 保证一定的植被高度及盖度, 避免单一物种的大量出现, 从而进一步提高人工林生态系统的稳定性是以后森林管理中需要考虑的。
附录 Supplementary Material
附录1 人工林斑块及边缘蛾类物种原始采集记录表
Appendix 1 Table for original collection records of moth species in plantation patches and edges
参考文献
Galling insects in a fragmented forest: Incidence of habitat loss, edge effects and plant availability
DOI:10.1007/s10841-016-9845-2 URL [本文引用: 1]
Old forest edges may promote the distribution of forest species in carabid assemblages (Coleoptera: Carabidae) in Croatian forests
DOI:10.14411/eje.2014.090 URL [本文引用: 1]
Comparison of moth communities following clear-cutting and wildfire disturbance in the southern boreal forest
DOI:10.1016/j.foreco.2011.08.021 URL [本文引用: 1]
Taxonomic and phylogenetic diversity of plants in a Castanopsis kawakamii natural forest
DOI:10.17520/biods.2020399
[本文引用: 1]

<p id="p00005"><strong>Aims:</strong> Environmental heterogeneity of forest gaps leads to variation in taxonomic and phylogenetic diversity of trees in these areas. Studying tree diversity in different sizes of forest gap communities can help to reveal the mechanisms that drive the formation and maintenance of biodiversity. This study took <i>Castanopsis kawakamii</i>gaps as the research object, and aimed to reveal the relationship between the taxonomic and phylogenetic diversity of plants and its environmental influence factors. </p> <p id="p00010"><strong>Methods:</strong> We examined different sizes of forest gaps in a <i>Castanopsis kawakamii</i> natural forest as to study the taxonomic and phylogenetic diversity of plants, and used a generalized linear model (GLM) to explore the environmental factors driving the community assembly. </p> <p id="p00015"><strong>Results:</strong> We found that the plant species and plant abundance in large gaps (> 200 m<sup>2</sup>) were higher than those of medium gaps ([50 m<sup>2</sup>, 100 m<sup>2</sup>)), small gaps ([30 m<sup>2</sup>, 50 m<sup>2</sup>)) and non-gaps (100 m<sup>2</sup>). The phylogenetic community structure of the large gaps tends to diverge, while that of the medium gaps, small gaps and non-gaps were affected by the combined effect of habitat filtering and competitive exclusion. The phylogenetic community diversity index (PD) was significantly positively correlated with species richness (SR), Margalef index and Shannon-Wiener index, which is related to the higher species composition of sparse species than dense species in forest gaps. Overall, forest gap size had a significantly positive effect on species diversity, and the soil total nitrogen content had a significantly positive effect on community phylogenetic diversity and phylogenetic structure. </p> <p id="p00020"><strong>Conclusion:</strong> The formation of forest gaps increase the taxonomic and phylogenetic diversity of trees in natural forests, with gap size and soil total nitrogen jointly driving tree diversity in these natural forest gaps. </p>
格氏栲天然林林窗植物物种多样性与系统发育多样性
DOI:10.17520/biods.2020399
[本文引用: 1]
林窗环境异质性导致群落物种多样性与系统发育多样性(phylogenetic diversity, PD)存在差异, 研究不同大小的林窗中群落的物种多样性与系统发育多样性有助于揭示林下生物多样性的形成及维持机制。本文以格氏栲(Castanopsis kawakamii)天然林为研究对象, 通过Pearson相关性分析与广义线性模型探讨了林窗内物种多样性与系统发育多样性间的相互关系及其环境影响因素。结果表明: (1)大林窗(面积 > 200 m<sup>2</sup>)植物种类及多度均高于中林窗(50 m<sup>2</sup> ≤ 面积 < 100 m<sup>2</sup>)、小林窗(30 m<sup>2</sup> ≤ 面积 < 50 m<sup>2</sup>)和非林窗(面积 = 100 m<sup>2</sup>)。大林窗群落系统发育结构趋于发散, 中、小林窗和非林窗群落系统发育结构受到生境过滤和竞争排斥综合作用。(2)群落系统发育多样性指数与物种丰富度(species richness, SR)、Margalef丰富度指数和Shannon-Wiener指数均呈显著正相关, 这与林窗内稀有种种类组成多于优势种有关。(3)林窗面积对物种多样性存在显著正效应; 土壤全氮含量对系统发育多样性和系统发育结构存在显著正效应。林窗形成提高了格氏栲天然林群落物种多样性和系统发育多样性, 林窗面积与土壤全氮共同驱动了格氏栲天然林林窗物种多样性和系统发育多样性的变化。
Edge effect benefits galling insects in the Brazilian Amazon
DOI:10.1007/s10531-012-0333-z URL [本文引用: 1]
The moth specialist spider Cyrtarachne akirai uses prey scales to increase adhesion
DOI:10.1098/rsif.2019.0792
URL
[本文引用: 1]
\n Contaminants decrease adhesive strength by interfering with substrate contact. Spider webs adhering to moths present an ideal model to investigate how natural adhesives overcome contamination because moths' sacrificial layer of scales rubs off on sticky silk, facilitating escape. However, Cyrtarachninae spiders have evolved gluey capture threads that adhere well to moths.\n Cyrtarachne\n capture threads contain large glue droplets oversaturated with water, readily flowing but also prone to drying out. Here, we compare the spreading and adhesion of\n Cyrtarachne akirai\n glue on intact mothwings, denuded cuticle and glass to the glue of a common orb-weaving spider,\n Larinioides cornutus,\n to understand how\n C. akirai\n glue overcomes dirty surfaces. Videos show that\n C. akirai\n 's glue spreading accelerates along the underlying moth cuticle after the glue seeps beneath the moth scales—not seen on denuded cuticle or hydrophilic glass.\n Larinioides cornutus\n glue droplets failed to penetrate the moth scales, their force of adhesion thus limited by the strength of attachment of scales to the cuticle. The large size and low viscosity of\n C. akirai\n glue droplets function together to use the three-dimensional topography of the moth's scales against itself via capillary forces. Infrared spectroscopy shows\n C. akirai\n glue droplets readily lose free-flowing water. We hypothesize that this loss of water leads to increased viscosity during spreading, increasing cohesive forces during pull-off. This glue's two-phase behaviour shows how natural selection can leverage a defensive specialization of prey against themselves and highlights a new design principle for synthetic adhesives for adhering to troublesome surfaces.\n
Composition and elevation of spruce forests affect susceptibility to bark beetle attacks: Implications for forest management
DOI:10.3390/f5010088 URL [本文引用: 1]
Effectiveness evaluation based on the afforestine construction in Beijing Plain
北京平原百万亩造林工程建设效果评价研究
Species diversity in overlapped zones of typical secondary forests in Guandishan Mountains
关帝山次生林区典型森林交错带物种多样性研究
Characteristics of moth community in different types of urban forest plantations in Beijing
北京不同类型人工林中蛾类群落特征
PAST: Paleontological statistics software package for education and data analysis
Determinants of moth diversity and community in a temperate mountain landscape: Vegetation, topography, and seasonality
Bird predation on forest insects: An exclosure experiment
Exclusion experiments show that birds significantly reduce densities of larval Lepidoptera on forest understory vegetation. When insect densities are already low, bird predation may act both as a population regulator and as a strong agent of natural selection.
Moth assemblages as indicators of environmental quality in remnants of upland Australian rain forest
DOI:10.1046/j.1365-2664.2000.00490.x URL [本文引用: 1]
Logging in boreal field-forest ecotones promotes flower-visiting insect diversity and modifies insect community composition
DOI:10.1111/icad.2015.8.issue-2 URL [本文引用: 1]
Impacts of plain afforestation on forest landscape patterns in Beijing
平原造林对北京森林景观格局的影响
A study on composition structure and species diversity in ecotone of coniferous and broad-leaved forest in Russia plain
俄罗斯平原针-阔林过渡带森林群落组成结构与物种多样性的研究
Ground beetle diversity and their value as bioindicators for desertification in a natural desert of the middle of the Hexi Corridor, Northwest China
河西走廊天然固沙植被区地表甲虫多样性及其对沙漠化的指示作用
Analysis of forestry impacts and biodiversity in two Pyrenean forests through a comparison of moth communities (Lepidoptera, Heterocera)
DOI:10.1111/ins.2007.14.issue-4 URL [本文引用: 1]
Carabids and forest edge: Spatial pattern and edge effect
DOI:10.1016/S0378-1127(00)00654-X URL [本文引用: 1]
Populations and assemblages living on the edge: Dung beetles responses to forests-pasture ecotones
DOI:10.7717/peerj.6148
URL
[本文引用: 1]
Edge effects alter insect biodiversity in several ways. However, we still have a limited understanding on simultaneous responses of ecological populations and assemblages to ecotones, especially in human modified landscapes. We analyze edge effects on dung beetle populations and assemblages between livestock pastures and native temperate forests (Juniperusand pine-oak forests (POFs)) to describe how species abundances and assemblage parameters respond to edge effects through gradients in forest-pasture ecotones. InJuniperusforest 13 species avoided the ecotones: six species showed greater abundance in forest interior and seven in pasturelands, while the other two species had a neutral response to the edge. In a different way, in POF we found five species avoiding the edge (four with greater abundance in pastures and only one in forest), two species had a neutral response, and two showed a unimodal pattern of abundance near to the edge. At the assemblage level edge effects are masked, as species richness, diversity, functional richness, functional evenness, and compositional incidence dissimilarity did not vary along forest-pasture ecotones. However, total abundance and functional divergence showed higher values in pastures in one of the two sampling localities. Also, assemblage similarity based on species’ abundance showed a peak near to the edge in POF. We propose that conservation efforts in human-managed landscapes should focus on mitigating current and delayed edge effects. Ecotone management will be crucial in livestock dominated landscapes to conserve regional biodiversity and the environmental services carried out by dung beetles.
Macro-moth families differ in their attraction to light: Implications for light-trap monitoring programmes
DOI:10.1111/icad.2014.7.issue-5 URL [本文引用: 1]
Few keystone plant genera support the majority of Lepidoptera species
DOI:10.1038/s41467-020-19565-4
PMID:33188194
[本文引用: 1]
Functional food webs are essential for the successful conservation of ecological communities, and in terrestrial systems, food webs are built on a foundation of coevolved interactions between plants and their consumers. Here, we collate published data on host plant ranges and associated host plant-Lepidoptera interactions from across the contiguous United States and demonstrate that among ecosystems, distributions of plant-herbivore interactions are consistently skewed, with a small percentage of plant genera supporting the majority of Lepidoptera. Plant identities critical for retaining interaction diversity are similar and independent of geography. Given the importance of Lepidoptera to food webs and ecosystem function, efficient and effective restoration of degraded landscapes depends on the inclusion of such 'keystone' plants.
The effects of forestry on carabid beetles (Coleoptera: Carabidae) in boreal forests
DOI:10.1007/s10841-006-9014-0 URL [本文引用: 1]
Determinants of nocturnal Lepidopteran diversity and community structure in a conifer-dominated forest
DOI:10.1007/s10531-009-9732-1 URL [本文引用: 2]
Different responses of plants and herbivore insects to a gradient of vegetation height: An indicator of the vertebrate grazing intensity and successional age
DOI:10.1111/j.2006.0030-1299.15126.x URL [本文引用: 1]
Living on the edge: Effects of clear-cut created ecotones on nocturnal macromoth assemblages in the eastern boreal forest, Canada
DOI:10.1016/j.foreco.2021.119309 URL [本文引用: 2]
Short-term effects of wildfire in boreal peatlands: Does fire mitigate the linear footprint of oil and gas exploration?
Analysis of moth diversity in oak forests, Quercus variabilis and Q. mongolica in northern China
北方蒙古栎和栓皮栎林蛾类多样性比较分析
Managing the forest for more than the trees: Effects of experimental timber harvest on forest Lepidoptera
DOI:10.1890/10-0715.1 URL [本文引用: 1]
Edge effect and its impacts on forest ecosystem: A review
Edge effect is an important concept in ecology and biological conservation, playing an important role in the study of ecological processes such as energy and material flow at ecosystem scale and landscape scale. This paper expatiated the connotation, features, quantitative evaluation (basis of quantitative analysis, strength, impact zone, and models, <em>etc</em>.), and applied aspects of edge effect, summarized the impacts of edge effect on forest ecosystem, analyzed the deficiencies in the study of edge effect, and prospected related research directions, aimed to provide references for forest and protected area management.
边缘效应及其对森林生态系统影响的研究进展
Attraction to light—From how far do moths (Lepidoptera) return to weak artificial sources of light?
DOI:10.14411/eje.2012.010 URL [本文引用: 1]
Analysis on the forest communities of Dinghushan Guangdong. X. Communities edge effect
鼎湖山森林群落分析. X. 边缘效应
Examination of edge effects in a Cryptomeria fortune plantation in Zhougong Mountain, western Sichuan
DOI:10.17520/biods.2015349
[本文引用: 1]
To investigate edge effects on community structure, species composition and diversity in an artificial forest, research was conducted on Zhougong Mountain, a forest park in western Sichuan, and a large fragmented plantation of Cryptomeria fortunei was selected as the sample plot. The average DBH (diameter at breast height), average height, average density, richness index (D) and Shannon-Wiener index (H) were used to evaluate edge effects. Based on a field survey, 5 transects (width = 10 m) were established from the edge to interior forest (gradient 1 to gradient 5), and 4 small plots (10 m × 10 m) were chosen in each transect using a random sampling method. Results showed that 111 species, belonging to 96 genera and 54 families were recorded in 20 small plots with a total area of 2,000 m2. The following results were also found in this investigation: (1) The number of species decreased away from the edge to forest interior. (2) In terms of community structure, a decrease of average DBH was found in the tree layer with the increase of edge gradients. Average density followed the opposite pattern and no significant differences were found in average height. In the shrub layer, average density decreased with the increase of edge gradients and no significant differences were found in average height. Average density and height both decreased from the edge to interior forest in the herb layer. (3) Based on the analysis of species diversity, richness index (D) and Shannon-Wiener index (H) of the plantation, decreased values were generally found with an increase of edge gradients, and were much more obvious in the shrub and herb layer. Moreover, the number of common species and Sørensen’s similarity coefficient between edge gradients in the forest interior and edge 1 both showed a decreasing trend from the edge to interior forest. (4) Based on a comprehensive analysis, a clear change of community was found between edge 2 and edge 3, indicating that edge effects can reach 20 m into the fragmented patch of the Cryptomeria fortunei plantation.
川西周公山柳杉人工林群落的边缘效应
DOI:10.17520/biods.2015349
[本文引用: 1]
为研究人工林群落的边缘效应特征, 本文以川西周公山森林公园的柳杉(Cryptomeria fortunei)人工林破碎化大斑块为对象, 以植株平均胸径、平均高度、平均密度、丰富度指数(D)和Shannon-Wiener指数(H)来综合衡量边缘效应深度。在实地踏查的基础上, 从林缘向林内(梯度1至梯度5)设置5条样带(宽度为10 m), 在每条样带中设置4个10 m × 10 m的小样方进行调查。结果表明: (1)从物种组成上看, 在总面积为2,000 m<sup>2</sup>的20个小样方中共记录到111个物种, 隶属于54科96属, 物种数从林缘至林内递减。(2)从群落结构上看, 乔木层的平均胸径从林缘至林内呈减小趋势, 平均密度则相反, 平均高度无显著变化; 灌木层的平均密度从林缘向林内减小, 平均高度无显著变化; 草本层的平均密度和平均高度均呈减小趋势。(3)从物种多样性上看, 总体上各层次的丰富度指数和Shannon-Wiener指数均从林缘向林内呈减小趋势, 其中灌木层和草本层的变化趋势最明显; 同时, 林内各梯度与梯度1 (林缘)的共有种和相似性系数从林缘向林内递减。(4)分析各项群落特征发现, 平均高度、平均密度和相似性系数的数值在梯度2向梯度3过渡时的起伏变化最明显, 推断本研究中柳杉人工林斑块的边缘深度可达林内20 m。
Biological and ecological bases of using insect as a bio-indicator to assess forest health
以昆虫作为指示生物评估森林健康的生物学与生态学基础
The technical and performance characteristics of a low-cost, simply constructed, black light moth trap
Diversity of darkling beetle community in the artificial cultivation Caragana intermedia shrub-desert grassland ecotone in Yanchi County, Ningxia, China
人工柠条-荒漠草地交错带拟步甲昆虫群落多样性
Analyse on soil improvement of different types of water and soil conservation forests mixed with Pinus tabulaeformis in semi-arid area of western Liaoning Province
辽西半干旱区不同类型油松水土保持林土壤改良效应分析
A study on the ecotones between Quercus mongolica community and other communities
蒙古栎群落交错带(ecotone)的研究
Exploitation of sexual signals by predators and parasitoids
DOI:10.1086/420412 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}