



原生动物是浮游动物的主要类群之一, 其作为微食物网中的重要营养级, 担负着维持生态平衡、水环境健康、能量转运枢纽以及生物地球化学循环等重要功能, 是水生态系统中不可或缺的一部分(许木启等, 1998; 沈韫芬, 1999)。与其他高等生物相比, 原生动物受环境影响更强, 从而能够更加灵敏迅速地反映出水环境的变化, 常作为水环境评价的指示物种(沈韫芬, 1999)。近年来, 随着分子技术的快速发展, 高通量测序技术越来越多地被运用到不同环境和生态系统原生动物群落多样性的研究中。例如, Li等(2021)利用高通量测序技术研究了南海北部沿河口至盆地样带活动原生动物群落的生物地理分布模式和群落构建机制; Jiang等(2021)对南海4个河口纤毛类原生动物群落的多样性和季节性动态展开了研究, 表明了栖息地和环境条件的变化比季节变化对原生动物群落组成的影响更大; Xu等(2021)利用高通量测序技术揭示原生动物对酸性矿山废水侵蚀土壤的生态响应机制。相较于传统形态学, 利用高通量测序技术研究原生动物能够使得样品分析更为迅速、准确, 进而能更详尽、细致、全面地揭示原生动物群落结构的组成和多样性变化。
青藏高原作为“地球第三极”, 有着独特的自然条件和丰富的生物资源(蒋志刚, 2018)。青藏高原地区原生动物研究虽然在近年来受到了国内外学者的广泛关注, 但主要是基于传统形态学的研究方法探讨原生动物的多样性(龚迎春等, 2012; 杜红春等, 2022)。目前将高通量测序技术运用于青藏高原地区原生动物, 尤其是雅鲁藏布江原生动物群落多样性的研究还未有较为全面的相关报道(马宝珊等, 2015; 梅建西, 2020①(①梅建西 (2020) 雅鲁藏布江周丛原生动物群落结构特征及其与环境因子的关系. 硕士学位论文, 西南大学, 重庆.))。本研究于2021年7月(丰水期)采样,然后通过18S rDNA高通量测序技术对雅鲁藏布江中游28个干、支流样点的原生动物群落多样性及其与水环境之间的关系进行了分析, 以期揭示高原河流生态系统中原生动物群落多样性的动态特征以及群落变化的驱动因素, 为进一步研究雅鲁藏布江流域以及为青藏高原水环境的监测和保护提供基础数据和决策参考。
1 材料与方法
1.1 研究区概况
雅鲁藏布江发源于西藏西南部的杰马央宗冰川, 在中国境内, 河流全长2,057 km, 流域范围在28°00'-31°16′ N和82°00'-97°07′ E之间。根据流域自然地形地貌、河谷特征, 雅鲁藏布江可划分为上、中、下游3个河段。其中, 从里孜‒派镇为雅鲁藏布江中游段(关志华和陈传友, 1984; 马腾霄等, 2022)。雅鲁藏布江整个中游河段为宽谷和窄谷相间的高山河谷地貌, 尤其是曲水县‒派镇段, 河岸地貌多为典型的高山峡谷。雅鲁藏布江中游河道落差大, 处于温带半湿润半干旱高原季风气候区, 气候温暖兼凉性, 年均降水量约为300-600 mm, 径流总体上以降水、融雪和地下水补给组成, 故而蕴含了丰富的水能资源并形成了从荒漠到高山灌丛再到森林过渡的复杂生境和植被类型(王蕊等, 2015)。同时, 雅鲁藏布江中游也是全河段人口最密集区域, 是西藏自治区开发较早、生产发达、经济繁荣的区域之一。独特的气候特征、地形地貌以及频繁的人类活动造就了雅鲁藏布江中游复杂多样的生态系统和丰富的生物资源, 使其成为研究雅鲁藏布江整个流域乃至青藏高原甚至亚洲生物多样性的重要基础区域(李正飞等, 2022)。
1.2 采样点布设与样品采集
根据雅鲁藏布江自然地貌、具体生境特征, 在雅鲁藏布江中游(曲水县‒派镇)共设置28个样点, 根据实际环境状况以及考虑到样品采集过程的安全性, 所有样点均设置于靠近河岸处, 且所有样点均位于雅鲁藏布江干流及其支流上。样点布设涉及雅鲁藏布江一级支流7条(雅砻河、杰当浦、加雄河、丝波绒曲、金东曲、里龙扑曲以及多空浦曲)、二级支流1条(雅堆曲), 其中干流11个样点、支流17个样点(图1)。在每个样点表层(大约20 cm深)采集9 L水样, 水样采集后立即通过0.22 μm的聚碳酸酯膜进行过滤, 最后得到84份水体样本(28个地点 × 3个重复), 随即将聚碳酸酯膜用无菌冻管装封后放置于液氮中保存, 储存在‒80℃条件下, 用于DNA提取。
图1

图1
雅鲁藏布江中游样点分布示意图
Fig. 1
Distribution of sample sites in the middle reaches of the Yarlung Zangbo River
1.3 水体理化因子
利用HI-98195便携式多参数水质测定仪测定水体pH值、水温(water temperature, WT)、电导率(electrical conductivity, EC); 利用HI-98193便携式溶解氧测定仪测定溶解氧(dissolved oxygen, DO); 利用HI-98703高精度数据型浊度测定仪测定水体浊度(turbidity, TUR); 利用FP-111直读式流速仪测定水体流速(water flow speed, WS)。利用全球定位系统(Explorist500, 美国麦哲伦公司)记录样点经度、纬度和海拔(altitude, ALT)高度, 上述理化指标均在现场测得, 并且进行3次重复。参照水和废水监测分析方法(第四版) (国家环境保护总局和水和废水监测分析方法编委会, 2002)和地表水环境质量标准(GB3838-2002)于室内完成总氮(total nitrogen, TN)、总磷(total phosphorus, TP)、铵态氮(ammonia nitrogen, NH4+-N)和化学需氧量(chemical oxygen demand, COD)的测定。
1.4 DNA提取和高通量测序
1.5 数据处理与分析
使用QIIME2软件中的DADA2插件(Bokulich et al, 2018; Bolyen, 2019)获得扩增子序列变异(amplicon sequence variants, ASVs), 并使用SILVA数据库(138版) (Quast et al, 2013)进行注释。非原生动物的ASVs和低丰度的ASVs (< 10个reads)被删除, 随后将ASV表进行标准化处理用于后续分析。α多样性指数(包括丰富度指数、Shannon多样性指数、Pielou均匀度指数和Simpson多样性指数)使用R中的vegan包进行计算; β多样性及其组分分解利用R中的adespatial包根据Jaccard相异指数进行计算; 主坐标分析(principal coordinate analysis, PCoA)和相似性分析(analysis of similarities, ANOSIM)利用R语言中的vegan包根据Bray-Curtis距离进行分析; 距离衰减曲线被用来评估空间因素对生物群落结构的影响, 群落Bray-Curtis相似性随地理距离变化的衰减特征利用R软件中的vegan包和geosphere包进行分析; 理化因子与群落的相关性热图利用R软件中的pheatmap包进行可视化; 采样点地图在ArcMap 10.6.1中绘制。
2 结果
2.1 原生动物群落结构及丰度
本研究共检获原生动物11门34纲58目84科204属686个ASVs。在所有样点中, 优势类群(相对丰度> 1%)共有7个, 分别为丝足虫门(35.93%)、纤毛虫门(19.70%)、双鞭毛虫门(16.06%)、顶复门(9.38%)、变形虫门(8.13%)、眼虫门(6.71%)和中阳虫门(3.35%)。其他类群(Myxogastria、Gracilipodida、Heterolobosea和Picozoa)占比较少, 共占0.74%。在河段上, 干流平均相对丰度排名前3的类群依次为丝足虫门、双鞭毛虫门、纤毛虫门, 占比分别为29.76%、22.67%和20.50%。支流平均相对丰度前3的类群依次为丝足虫门、纤毛虫门、双鞭毛虫门, 占比分别为39.05%、19.30%和12.73% (图2a)。在所有样点中, 丰度在样点L23有最高值(1.45 × 104 ind./L), 在样点L6有最低值(4.27 × 103 ind./L) (图2b)。在河段上, 干流丰度小于支流, 但两者间无显著性差异(Wilcoxon test, P = 0.208)。根据干、支流样点间丰度方差(σ)可知, 支流样点间丰度波动大于干流(图2c)。
图2
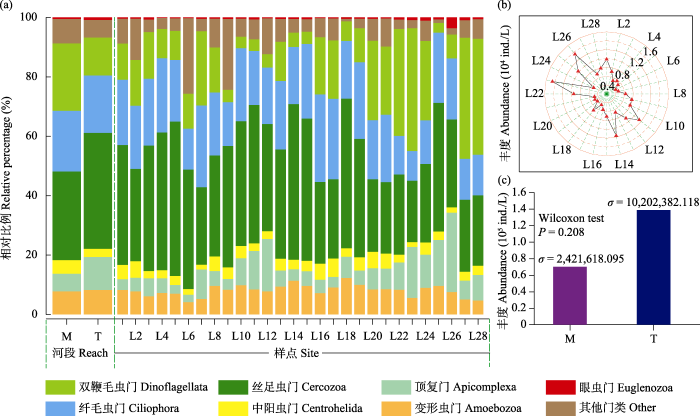
图2
研究区域各样点及河段门级水平上原生动物群落组成(a)、各样点丰度(b)和各河段丰度(c)。M: 干流; T: 支流; σ: 方差。
Fig. 2
Protozoan community composition at the phylum level at each site and reach in the study area (a), abundance at each site (b) and abundance at each reach (c). M, Mainstream; T, Tributary; σ, Variance.
2.2 原生动物群落多样性
总体上干流Simpson多样性指数 (均值 = 0.98)高于支流(0.96), 在河段间具有显著性差异(P = 0.022); 干流Shannon多样性指数(5.27)高于支流(4.95), 在河段间该指数没有显著性差异(P = 0.329); 干流Pielou均匀度指数(0.82)高于支流(0.77), 在河段间不具有显著性差异(P = 0.066); 干流物种丰富度(614.82)低于支流(635.88), 在河段间没有显著性差异(P = 0.611) (图3)。
图3
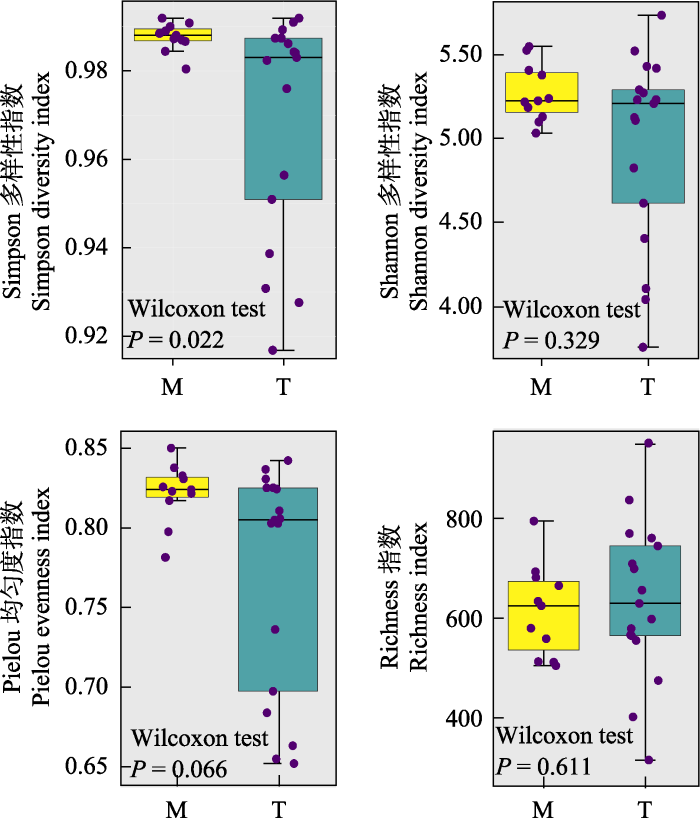
图3
研究区域原生动物群落α多样性。M: 干流; T: 支流。
Fig. 3
Protozoan community α diversity in the study area. M, Mainstream; T, Tributary.
本研究对雅鲁藏布江中游丰水期原生动物群落进行β多样性分析, 结果表明, 在支流、干流和全河段3个空间尺度下, 原生动物群落总体β多样性分别为0.34、0.31、0.34, 总体上差异不大。周转组分在所有空间尺度下对群落间物种组成差异(β多样性)的贡献均占主导地位, 分别各占其总体β多样性的76.5%、87.1%和82.1%; 嵌套组分的贡献均很小, 分别各占23.5%、12.9%和17.9% (图4a)。对各空间尺度下配对样点之间的原生动物群落进行β多样性组分分解, 结果表明, 在全河段中, 共有378组配对样点, 配对样点之间的β多样性(均值 = 0.69)主要是由周转组分(均值 = 0.64)贡献, 占比高达92.8%, 嵌套组分(均值 = 0.05)对β多样性的贡献仅为7.2%; 在支流河段中, 共有136组配对样点, 配对样点之间的β多样性(均值 = 0.68)主要由周转组分(均值 = 0.62)贡献, 占比高达91.2%, 嵌套组分(均值 = 0.06)对β多样性的贡献仅为8.8%; 在干流河段中, 共有55组配对样点, 配对样点之间的β多样性(均值 = 0.62)同样主要由周转组分(均值 = 0.59)贡献, 占比高达95.2%, 嵌套组分(均值 = 0.03)对β多样性的贡献仅为4.8% (图4b)。雅鲁藏布江中游原生动物群落β多样性分析结果说明, 在研究期间, 无论是在整体空间尺度上, 还是在配对样点间, 原生动物群落物种组成差异主要是由物种替换(即周转)引起的, 并未形成明显的嵌套格局。
图4
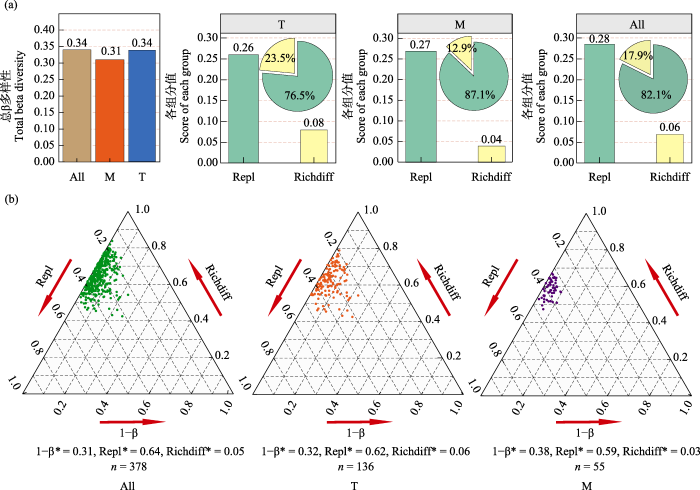
图4
研究区域原生动物群落β多样性分析。总β多样性及各组分占比(a), 群落β多样性组分分解(b)。All: 全河段; M: 干流; T: 支流; *: 平均值; n: 样点对; Repl和Richdiff表示β多样性的两种组分。
Fig. 4
β diversity analysis of protozoan communities in the study area. Total β diversity of the protozoan community and the proportion of each component (a). Decomposition of the β diversity component of the community (b). All, All river reaches; M, Mainstream; T, Tributary; *, Mean; n, Sample point pair; Repl and Richdiff denote the two components of β diversity.
2.3 群落PCoA和地理格局分析
对雅鲁藏布江中游丰水期的原生动物群落进行基于Bray-Curtis距离的主坐标分析(PCoA), 结果显示, 地理上的原生动物群落集群及其在95%的置信区间内没有重叠, 表明不同河段的原生动物群落能够被明确分开。主坐标分析的第一轴PCoA 1能解释雅鲁藏布江中上游丰水期原生动物群落25.25%的变化, 第二轴PCoA 2能解释原生动物群落14.43%的变化。相似性分析(ANOSIM)结果表明, 原生动物群落在干流、支流河段间存在显著差异(P < 0.01, R = 0.197), 且支流样点组内差异强于干流样点组内差异(图5a)。
图5
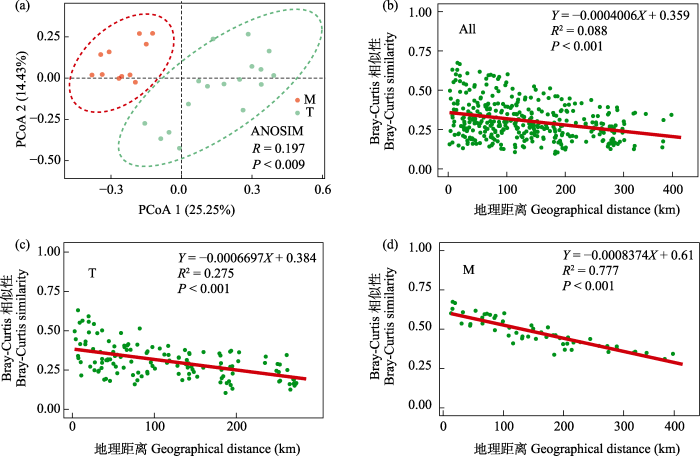
图5
研究区域原生动物群落PCoA分析(a)和全河段(b)、干流河段(c)、支流河段(d)地理格局分析。All: 全河段; M: 干流; T: 支流。
Fig. 5
Protozoan community PCoA analysis (a) and geographic pattern analysis for the all river section (b), mainstream section (c), and tributary section (d) in the study area. All, All river reaches; M, Mainstream; T, Tributary.
原生动物群落Bray-Curtis相似性随地理距离分析结果显示, 无论在整体流域上还是在各河段上, 距离衰减分析都显示出一个明显的负斜率曲线, 原生动物群落相似性随地理距离的增加均呈极显著的距离衰减效应(P < 0.01), 且衰减程度为干流河段最强(R2 = 0.777), 支流河段次之(R2 = 0.275), 流域整体河段水平上衰减程度最弱(R2 = 0.088)。群落相似性的地理距离衰减特征表明雅鲁藏布江中游丰水期原生动物群落的空间分布特征符合生物地理分布的一般模式(图5b, c, d)。
2.4 原生动物群落共现网络分析
筛选全河段丰度前300的ASV进行物种共存分析, 筛选各河段丰度前300的ASV进行群落共线网络分析。共存物种分析结果显示干流和支流两个河段间共存物种较多(ASVs = 286), 单独出现在支流的物种较少(ASVs = 15), 而干流没有单独出现的物种(图6a)。在共存物种中, 丝足虫门所占比例最高(39%), 黏菌门所占比例最低(1%); 在支流特有物种中, 纤毛虫门所占比例最高(60%), 眼虫门所占比例最低(7%) (图6b)。雅鲁藏布江中游不同河段空间尺度上原生动物群落共现网络分析结果显示, 在3个河段空间尺度上, 原生动物群落共现网络连接数、平均度、图密度、网络模块化系数为干流最高; 共现网络直径、平均聚类系数和平均路径长度为全河段最高(图6c, 表1)。
图6
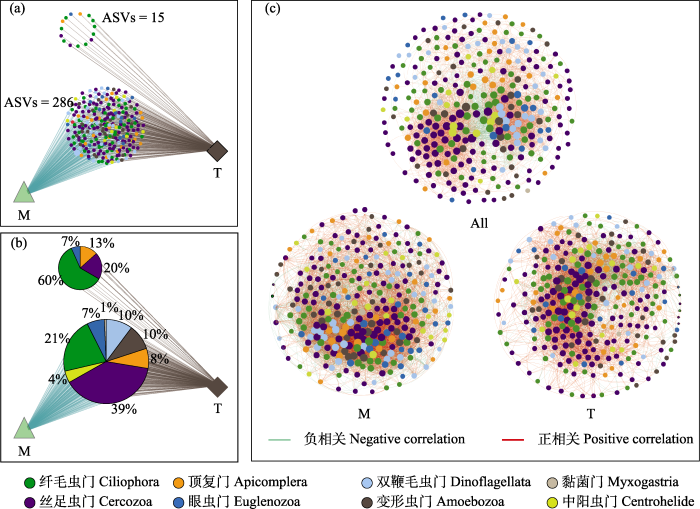
图6
研究区域原生动物群落共现模式分析。共有ASVs (a)及其所属类群(b), 群落共现网络(c)。All: 全河段; M: 干流; T: 支流。
Fig. 6
Protozoan community co-occurrence pattern analysis in the study area. Shared ASVs (a) and the taxa they belong to (b), community co-occurrence network (c). All, All river reaches; M, Mainstream; T, Tributary.
表1 原生动物群落共现网络的拓扑结构特征
Table 1
| 网络拓扑指标 Network topology indicators | 全河段 All river reaches (All) | 干流 Mainstream (M) | 支流 Tributary (T) |
|---|---|---|---|
| 连接数 Number of connections | 4,171 | 6,397 | 5,044 |
| 平均度 Average degree | 27.807 | 42.647 | 33.627 |
| 网络直径 Network diameter | 8 | 4 | 6 |
| 平均路径长度 Mean path length | 2.641 | 2.201 | 2.422 |
| 图密度 Density of figure | 0.093 | 0.143 | 0.112 |
| 模块化系数 Modularity coefficient | 0.815 | 1.410 | 0.909 |
| 平均聚类系数 Mean clustering coefficient | 0.614 | 0.511 | 0.557 |
| 正相关占比 Positive correlation (%) | 82.6 | 64.5 | 73.5 |
| 负相关占比 Negative correlation (%) | 17.4 | 35.5 | 26.5 |
2.5 原生动物群落与理化因子相关性
图7
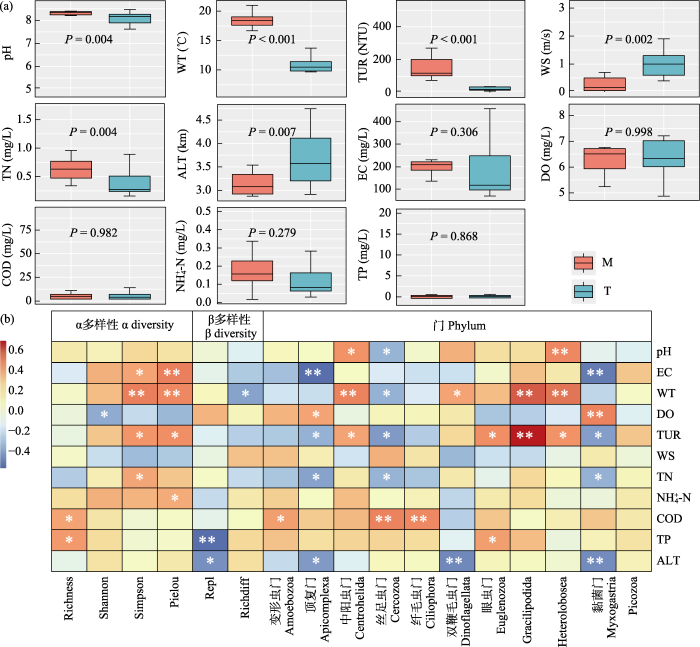
图7
研究区域原生动物群落与理化因子相关性分析。EC: 电导率; WT: 水温; DO: 溶解氧; TUR: 浊度; TN: 总氮; TP: 总磷; NH4+-N: 铵态氮; COD: 化学需氧量; WS: 水流速度; ALT: 海拔; M: 干流; T: 支流。
Fig. 7
Correlation analysis between protozoan communities and environmental factors in the study area. EC, Electrical conductivity; WT, Water temperature; DO, Dissolved oxygen; TUR, Turbidity; TN, Total nitrogen; TP, Total phosphorus; NH4+-N, Ammonia nitrogen; COD, Chemical oxygen demand; WS, Waterflow speed; ALT, Altitude; M, Mainstream; T, Tributary.
表2 雅鲁藏布江中游水体理化因子。EC: 电导率; WT: 水温; DO: 溶解氧; TUR: 浊度; TN: 总氮; TP: 总磷; NH4+-N: 铵态氮; COD: 化学需氧量; WS: 水流速度; ALT: 海拔; T: 支流; M: 干流。
Table 2
| 样点 Site | pH | EC (mg/L) | WT (℃) | DO (mg/L) | TUR (NTU) | TN (mg/L) | TP (mg/L) | NH4+-N (mg/L) | COD (mg/L) | WS (m/s) | ALT (km) | 河段 Reach |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 8.21 ± 0.03 | 230.67 ± 1.25 | 17.04 ± 0.11 | 5.53 ± 0.74 | 248.67 ± 17.91 | 0.74 | 0.04 | 0.02 | 20.00 | 0.00 | 3.54 | M |
| L2 | 8.66 ± 0.02 | 223.67 ± 0.47 | 21.02 ± 0.00 | 5.40 ± 0.02 | 530.00 ± 56.79 | 0.35 | 0.02 | 0.11 | 13.00 | 0.40 | 3.53 | M |
| L3 | 4.99 ± 0.12 | 457.67 ± 0.82 | 15.17 ± 0.06 | 5.78 ± 0.20 | 81.33 ± 2.73 | 0.57 | 0.14 | 0.14 | 98.00 | 0.70 | 3.71 | T |
| L4 | 6.76 ± 0.64 | 317.00 ± 0.82 | 12.44 ± 0.22 | 5.45 ± 0.08 | 14.07 ± 0.51 | 0.45 | 12.50 | 0.11 | 5.00 | 0.60 | 4.11 | T |
| L5 | 4.86 ± 0.12 | 248.00 ± 0.82 | 10.48 ± 0.06 | 4.87 ± 0.20 | 6.08 ± 1.09 | 0.89 | 0.08 | 0.28 | 4.00 | 1.10 | 4.51 | T |
| L6 | 7.62 ± 0.12 | 117.00 ± 0.00 | 6.58 ± 0.02 | 6.33 ± 0.24 | 10.93 ± 0.62 | 0.22 | 0.04 | 0.21 | 12.00 | 1.70 | 4.22 | T |
| L7 | 7.89 ± 0.04 | 148.67 ± 0.47 | 6.15 ± 0.04 | 6.08 ± 0.07 | 10.63 ± 0.87 | 0.24 | 0.01 | 0.06 | 2.00 | 0.40 | 4.74 | T |
| L8 | 8.23 ± 0.01 | 226.67 ± 0.47 | 19.34 ± 0.04 | 5.24 ± 0.10 | 269.67 ± 11.09 | 0.45 | 0.01 | 0.03 | 2.00 | 0.00 | 3.51 | M |
| L9 | 8.08 ± 0.07 | 366.00 ± 0.00 | 13.72 ± 0.00 | 6.01 ± 0.01 | 394.33 ± 4.11 | 0.56 | 0.03 | 0.15 | 6.00 | 0.60 | 3.57 | T |
| L10 | 8.32 ±0.02 | 321.33 ± 6.60 | 13.25 ± 0.01 | 5.26 ± 0.39 | 59.43 ± 3.72 | 0.27 | 0.01 | 0.06 | 2.00 | 1.30 | 3.88 | M |
| L11 | 8.22 ± 0.04 | 105.00 ± 0.00 | 11.20 ±0.02 | 6.47 ± 0.33 | 26.33 ± 0.93 | 0.36 | 0.01 | 0.15 | 2.00 | 0.40 | 3.51 | T |
| L12 | 8.27 ± 0.02 | 99.00 ± 0.82 | 9.87 ± 0.47 | 6.18 ± 0.20 | 9.97 ± 0.09 | 0.25 | 0.01 | 0.16 | 2.00 | 0.70 | 3.75 | T |
| L13 | 8.34 ± 0.04 | 219.00 ± 0.00 | 18.76 ± 0.00 | 8.26 ± 0.08 | 137.00 ± 3.56 | 0.80 | 0.01 | 0.10 | 2.00 | 1.90 | 3.17 | M |
| L14 | 8.16 ± 0.20 | 96.00 ± 0.00 | 10.01 ± 0.01 | 6.78 ± 0.11 | 5.85 ± 0.40 | 0.23 | 0.05 | 0.24 | 2.00 | 1.90 | 3.37 | T |
| L15 | 8.16 ± 0.03 | 108.33 ± 0.94 | 9.83 ± 0.07 | 6.23 ± 0.06 | 6.32 ± 0.52 | 0.16 | 0.01 | 0.32 | 5.00 | 0.50 | 4.14 | T |
| L16 | 8.40 ± 0.03 | 215.00 ± 0.00 | 19.35 ± 0.04 | 6.38 ± 0.09 | 118.33 ± 7.32 | 0.74 | 0.02 | 0.08 | 7.00 | 0.20 | 3.12 | M |
| L17 | 8.37 ± 0.06 | 208.00 ± 0.00 | 18.38 ± 0.00 | 6.76 ± 0.13 | 154.67 ± 3.68 | 0.96 | 0.04 | 0.02 | 2.00 | 0.60 | 3.68 | M |
| L18 | 8.05 ± 0.22 | 244.00 ± 5.19 | 10.83 ± 0.00 | 7.14 ± 0.08 | 37.97 ± 1.81 | 0.44 | 0.05 | 0.16 | 2.00 | 1.60 | 3.01 | T |
| L19 | 8.47 ± 0.02 | 181.00 ± 0.00 | 9.66 ± 0.05 | 7.02 ± 0.12 | 9.86 ± 0.60 | 0.61 | 0.01 | 0.06 | 4.00 | 1.00 | 3.48 | T |
| L20 | 8.38 ± 0.01 | 204.00 ± 0.00 | 18.25 ± 0.08 | 6.51 ± 0.04 | 113.33 ± 0.47 | 1.69 | 0.04 | 0.13 | 2.00 | 0.70 | 2.95 | M |
| L21 | 8.38 ± 0.03 | 202.00 ± 0.00 | 18.21 ± 0.02 | 6.34 ± 0.18 | 118.33 ± 4.50 | 0.50 | 0.04 | 0.22 | 2.00 | 0.10 | 2.93 | M |
| L22 | 8.33 ± 0.00 | 69.33 ± 0.94 | 11.41 ± 0.01 | 7.03 ± 0.02 | 10.61 ± 0.64 | 0.26 | 0.02 | 0.08 | 2.00 | 1.00 | 2.96 | T |
| L23 | 8.29 ± 0.06 | 75.00 ± 0.00 | 10.63 ± 0.00 | 6.71 ± 0.08 | 8.11 ± 0.34 | 0.24 | 0.01 | 0.05 | 14.00 | 3.10 | 3.20 | T |
| L24 | 8.27 ± 0.06 | 166.00 ± 0.00 | 16.68 ± 0.00 | 6.69 ± 0.05 | 93.30 ± 3.00 | 0.63 | 0.01 | 0.20 | 11.00 | 0.10 | 2.91 | M |
| L25 | 8.07 ± 0.10 | 71.33 ± 0.47 | 10.42 ± 0.03 | 7.21 ± 0.10 | 1.00 ± 0.10 | 0.51 | 20.90 | 0.08 | 94.00 | 1.20 | 2.91 | T |
| L26 | 8.21 ± 0.06 | 77.00 ± 0.00 | 9.73 ± 0.66 | 7.15 ± 0.06 | 1.00 ± 0.16 | 0.16 | 0.03 | 0.10 | 43.00 | 0.70 | 2.79 | T |
| L27 | 8.21 ± 0.01 | 142.00 ± 0.00 | 16.71 ± 0.09 | 6.74 ± 0.04 | 75.27 ± 1.20 | 0.34 | 0.04 | 0.24 | 6.00 | 0.10 | 2.89 | M |
| L28 | 8.24 ± 0.02 | 135.00 ± 0.00 | 18.54 ± 0.25 | 6.54 ± 0.21 | 78.90 ± 3.52 | 0.60 | 0.04 | 0.34 | 8.00 | 0.00 | 2.87 | M |
通过群落多样性参数以及门级类群与环境理化因子的Spearman相关分析, EC、WT、TUR、TN、NH4+-N、COD、TP与群落α多样性具有显著或极显著正相关性; WT和ALT与群落β多样性具有显著负相关性; 在所分析的理化因子中, 绝大多数与门级类群具有相关性, 如EC与黏菌门、顶复门呈极显著负相关; TUR与Gracilipodida类群呈极显著正相关; 与Heterolobosea类群、眼虫门和中阳虫门呈显著正相关; 与黏菌门、丝足虫门和顶复门呈显著负相关(图7b)。
为了进一步阐释原生动物群落结构与理化因子之间的关系, 探明理化因子对群落的交叉影响, 将原生动物群落与理化因子进行生物区系-环境(BIOENV)分析。结果显示在支流河段理化因子组合中, pH、WT、ALT组合的相关系数最高(0.4971); 其次为pH、WT、WS、ALT组合(0.4966), 影响支流原生动物群落的前5组理化因子主要发生在pH、WT、ALT、WS、EC和DO的组合上; 在干流河段理化因子组合中, EC的相关系数最高(0.7565); 其次为EC、ALT组合(0.7513), 影响干流原生动物群落的前5组理化因子主要发生在pH、EC、WT、TUR和ALT的组合上(表3)。
表3 理化因子与原生动物群落的BIOENV分析。EC: 电导率; WT: 水温; DO: 溶解氧; TUR: 浊度; WS: 水流速度; ALT: 海拔; T: 支流; M: 干流。
Table 3
| 分组 Group | 级别 Rank | 理化因子 Physicochemical factors | 相关系数 Correlation coefficient (ρ) | 显著性 Significance (P) |
|---|---|---|---|---|
| T | 1 | pH、WT、ALT | 0.4971 | 0.01 |
| 2 | pH、WT、WS、ALT | 0.4966 | 0.01 | |
| 3 | WT、ALT | 0.4820 | 0.01 | |
| 4 | pH、EC、WT、WS、ALT | 0.4760 | 0.01 | |
| 5 | pH、EC、WT、DO、WS、ALT | 0.4663 | 0.01 | |
| M | 1 | EC | 0.7565 | 0.01 |
| 2 | EC、ALT | 0.7513 | 0.01 | |
| 3 | EC、WT、ALT | 0.7369 | 0.01 | |
| 4 | pH、EC、WT、ALT | 0.6818 | 0.01 | |
| 5 | pH、EC、WT、TUR、ALT | 0.6818 | 0.01 |
3 讨论
3.1 原生动物群落多样性格局
本研究基于18S rDNA在雅鲁藏布江中游丰水期共鉴定到原生动物ASVs 5,964个, 共检获原生动物11门34纲58目84科204属686个ASVs, 远远超过基于传统形态学鉴定方法的物种数量(马宝珊等, 2015; 吴聪, 2022①(①吴聪 (2022) 雅鲁藏布江及其主要支流浮游动物与环境因子间的关系. 硕士学位论文, 武汉轻工大学, 武汉.))。传统形态学鉴定主要依靠显微镜进行物种鉴别, 往往需要原生动物活体状态下的特征, 因此常局限于能够培养或在样品采集后数日仍能保持生存和一定丰度的种类, 但极多的类群远远达不到这样的要求, 另一方面, 即使舍弃活体特征, 对固定后的样品进行种类鉴定和计数, 仍存在众多主观因素, 如有的物种由于体积极其微小难以识别、有的种类形态相似难以区分等, 整体上使得鉴定物种数少于环境样本实际的物种数(陈旭淼和徐奎栋, 2014)。18S rDNA高通量测序技术从DNA分子水平上对原生动物进行识别鉴定, 极大地避免了主观因素, 只要是存在于环境样本中的原生动物基本都能通过测序技术进行识别, 极大程度地保证了样本环境中物种数量的准确性。本文对雅鲁藏布江中上游原生动物群落物种组成进行分析, 结果显示无论在干流河段还是支流河段, 丝足虫、纤毛虫和双鞭毛虫均为优势类群。这与许木启和Petter Kasprzar (2001)对白洋淀、李凤超等(2006)对拒马河北京段、巴桑等(2017)对拉萨河原生动物群落特征研究结果相类似, 这可能是因为雅鲁藏布江中游水温普遍较低、丰水期水体扰动大, 水环境发生变化, 导致一些原生动物类群生态位降低, 使得丝足虫、纤毛虫和双鞭毛虫具有更低的生存压力(刘佳驹等, 2018; 张鹏等, 2022), 从而使其成为优势类群。
原生动物群落α多样性指数分析结果显示, 丰水期雅鲁藏布江中游原生动物多样性存在河段空间性差异, 原生动物多样性指数为干流 > 支流, 这与大多数河流原生动物多样性研究相一致(龚迎春等, 2012; 胡笑妍等, 2017; 张敏等, 2017)。这可能与本研究区域的流域范围大、覆盖的地形地貌复杂有关。雅鲁藏布江中游段支流较多, 各支流流域生境具有差异性, 造成了各支流的原生动物群落结构、物种组成不尽相同。另外, 原生动物多为附着性生物, 丰水期水体流量加大、流速快, 从而各支流原生动物随着水体汇入雅鲁藏布江干流, 使得雅鲁藏布江干流原生动物多样性整体升高。雅鲁藏布江支流原生动物物种数和丰度高于干流, 这与大多数水体原生动物的研究存在差异(杨威等, 2011; 白海锋, 2014②(②白海锋 (2014) 渭河流域浮游动物群落结构时空格局的研究. 硕士学位论文, 大连海洋大学, 辽宁大连.); 包宇飞, 2019)。一方面, 这可能与青藏高原独特的地理环境、原生动物长期的适应性生存有关。雅鲁藏布江流域的原生动物长期处于青藏高原的高寒生境, 长期的适应性进化使其在低温的水环境中生存能力、适应能力较强, 而雅鲁藏布江支流多发源于冰川融水, 多数属于冷水性河流(Harrison et al, 1992; 文安邦等, 2002), 水温普遍低于干流, 适应于大多数原生动物的生存和繁殖。另一方面, 雅鲁藏布江中游人口较为密集, 各支流周边多为牧地、农田, 丰水期农牧活动频繁(关志华和陈传友, 1984; Lennon et al, 2001), 人类活动对水体环境的干扰较大, 且支流水体流速相对较大, 使得由农牧活动形成的营养物质进入水体时不会造成持续的堆积, 支流营养物质增加较为适当, 有利于原生动物的生存。反观雅鲁藏布江干流河段, 在丰水期各大支流汇入雅鲁藏布江干流, 加上自然降水使得大量外来水体侵入干流, 河水体量大、水流速度相对缓慢, 使得水体对外来营养物质的稀释能力降低, 在一定程度上改变了干流原本的水体营养结构以及各种水环境特征, 从而使得一些原生动物不能适应于当前的水体环境, 造成部分原生动物物种的消亡或丰度的减少。
群落β多样性反映了不同地点间群落物种组成的变化, 这种变化可能是物种替换或周转(Repl), 以及物种损失或嵌套(Richdiff)的结果。关于群落β多样性, 如果物种组成变化是随机的并伴随自相关的生物过程, 且与物种的形成和扩散历史相关, 那么物种组成变化就主要体现在不同物种的替换, 即β多样性可由周转(即物种在空间上的更替)来解释(Legendre et al, 2005; 王魏根, 2018)。雅鲁藏布江中游原生动物群落β多样性分析结果显示, 周转在β多样性中占据主导地位, 这表明原生动物β多样性可能主要来源于物种随机分布和自相关的过程, 是由于物种在空间上或群落间发生更替导致的。这一结果与大多数水体微生物群落β多样性研究相似, 例如戴美霞等(2017)进行象山港浮游动物β多样性分析时发现总体以周转为主, 嵌套所占比例很低; 李明家等(2020)研究发现西藏横断山区溪流细菌β多样性组分中, 周转占据主导地位。在雅鲁藏布江中游原生动物群落分析中, 出现周转成分占比较高的现象, 这可能与原生动物的运动缓慢、扩散能力弱、其分布更多与物种形成历史、扩散程度以及流域生境和频繁的人为干扰有关。
3.2 群落动态变化及其相关性因素
关于地理因素对原生动物群落结构的影响。对雅鲁藏布江中游原生动物群落地理格局分析发现, 随着地理距离的增加, 原生动物群落的相似性下降, 表现出距离-衰减的生物地理分布格局, 尤其在干流河段衰减趋势最为强烈。这一现象可认为是扩散限制导致了雅鲁藏布江中游原生动物群落的距离-衰减关系。原生动物属于微生物的一大类, 具有体积微小和丰富度高的特征, 被认为具有较强的扩散能力, 但其主要依靠于载体进行被动扩散。雅鲁藏布江中游干流河道较宽、河体较深、河道曲折蜿蜒、水体缓冲力大、水流速度相较于支流低, 从而影响到以水体为载体进行扩散的原生动物的扩散能力以及影响局地生物群落的物种组成、多样性和群落功能。因此, 扩散限制成为塑造水体微生物群落生物地理分布格局的原因之一。这与国内外多数水环境微生物地理格局的研究相似, 如有研究表明(Fillinge et al, 2019; Logares et al, 2020), 扩散限制显著影响了热带、亚热带海洋表层以及德国地下含水层中微生物群落的构建; Lu等(2020)对长江上游真核浮游生物群落生物地理学研究表明, 地理距离对于构建真核浮游生物群落和确定河流网络中的空间差异很重要; Wu等(2017)对中国东海和南海表层水体中丰富微生物类群的研究发现, 扩散限制主导了其群落的构建; 高鹏飞等(2022)对中国湖泊细菌群落的生物地理分布格局及驱动机制研究中发现, 空间扩散限制是青藏高原湖泊浮游及沉积物细菌群落构建的主要驱动因素。
关于生物因子对原生动物群落结构的影响, 在共现网络中, 网络连接数可反映共现网络结构的复杂性; 平均度和网络直径可反映共现网络结构的紧密性; 图密度和平均路径长度可反映共现网络结构的稳定性; 边的正、负相关性分别反映物种之间的协同和竞争关系(Deng et al, 2012; Zhang et al, 2018)。本研究发现, 在全河段、干流河段和支流河段3个空间尺度上, 网络边之间均以正相关为主, 表明共现网络物种之间的互作关系以协同作用为主导。一般情况下, 同类群微生物往往存在拮抗作用, 但在极端生境中, 它们之间的协同作用却通常占据互作关系的主导地位, 这说明生态网络中不同微生物类群之间的互作关系是随具体生境情况发生变化的, 而不是固定不变的(Selbmann et al, 2013)。本研究原生动物共现网络分析结果与崔戈等(2023)对金沙江底栖真核微生物、孙胜浩等(2020)对澜沧江硅藻等一系列具有高海拔、低温特征的水环境微生物的研究具有相似之处。雅鲁藏布江位于青藏高原, 水环境具有高海拔、低温等极端生境特征。雅鲁藏布江原生动物群落为了适应环境特征, 导致种间互作关系以协同作用为主, 这可能是在极端生境中, 如果种间拮抗作用较强, 某些物种会由于竞争力弱而被淘汰, 从而使得群落物种减少、多样性降低。相反, 在极端环境下, 较强的种间协同作用更有利于群落内各物种的生存, 使得群落结构更加完整, 群落稳定性更高(Shu & Huang, 2022)。从网络的拓扑结构特征还可以反映出网络结构的连通性、复杂性和抗干扰能力。通常具有较高连通性、复杂性和稳定性的网络对于环境干扰的反应更为迅速(Zhou et al, 2010)。根据网络拓扑特征结果显示, 雅鲁藏布江中游干流河段的原生动物群落共现网络具有最大的连接数、平均度和图密度, 最短的网络直径和平均路径长度, 表明干流河段原生动物共现网络最为复杂、稳定, 干流河段原生动物群落抗干扰能力最强、对水环境变化最敏感。
关于理化因子对原生动物群落结构的影响, 本研究使用Spearman相关性分析和生物-环境分析(BIOENV)探究理化因子与原生动物群落结构的关系。Spearman相关分析结果显示, EC、WT、TUR、TN、NH4+-N、COD、TP是影响群落α多样性的主要理化因子。龚森森等(2021)对长湖浮游动物的研究发现, 总氮、总磷、硝态氮、磷酸盐、总悬浮物以及pH值是影响浮游动物群落结构的主要理化因子; 张鹏等(2022)对拉萨河原生动物的研究发现, 水温、溶解氧和总磷是影响群落结构的主要因子。这说明不同水体中影响浮游动物群落结构的理化因子不尽相同, 并且是多种理化因子共同作用的结果。群落α多样性主要由群落物种数和丰度决定, 在本研究中, EC、WT、TUR、TN、NH4+-N、COD、TP等理化因子是影响群落物种组成的关键因子, 其含量或水平的高低直接或间接地影响到原生动物的生存和繁殖。有研究表明, 电导率(EC)能够反映水体中各种离子的总含量①(①何琦 (2011) 增江流域和东江惠州段底栖硅藻多样性及分布特征. 硕士学位论文, 暨南大学, 广州.)。在丰水期, 通过雨水的冲刷作用以及河道水位的涨退, 河岸带植物、土壤有机质以及枯枝碎叶等随水体的涨退进入河流, 在水体微生物的分解作用下, 对水体中的离子总含量产生直接影响, 从而通过改变水体的营养结构水平而对原生动物群落结构产生影响(Zheng et al, 2018; 杨欣兰等, 2019)。水温(WT)对浮游动物的生存繁殖和物种密度等的影响极为显著。水温可以通过作用于浮游动物的生长代谢能力直接影响其生命周期, 也可以影响浮游植物的生长, 然后通过上行效应间接影响浮游动物的生长繁殖(Elliott et al, 2006; Kolmakov et al, 2008)。浊度(TUR)对水体原生动物的影响有两个方面, 其一为影响水体透光性进而影响到浮游植物的光合作用, 降低水体的初级生产力, 使浮游植物生物量下降, 然后依据食物网营养级关系, 使以浮游植物为饵料的浮游动物的生物量也相应地减少(周礼斌和陈非洲, 2015; 王丽等, 2018); 另一方面, 在适当的范围内, 浊度的增加有利于水环境中细菌的生存繁殖, 从而有利于以细菌作为食物来源的原生动物的生存繁殖(高原等, 2019)。雅鲁藏布江中游原生动物α多样性与TUR呈显著正相关, 表明当前雅鲁藏布江中游的浊度水平对浮游植物的光合作用强度影响较小, 且有利于细菌的繁殖, 从而对浮游动物的生长繁殖具有促进作用。总氮(TN)、铵态氮(NH4+-N)、总磷(TP)、化学需氧量(COD)等营养盐是浮游植物生存的物质基础。有研究表明(朱旭宇等, 2013; 许海等, 2019), 营养盐浓度适当的升高有利于浮游植物密度增加, 从而为原生动物等浮游动物提供丰富的食物来源。
水温(WT)和海拔(ALT)是影响雅鲁藏布江中游原生动物群落β多样性的主要理化因子, 且两者分别影响β多样性的不同组分, 这可能与雅鲁藏布江处于青藏高原这一特殊环境有关。原生动物经过了长期的进化, 适应了青藏高原低温的环境, 所以群落多样性在空间上与水温一般呈现出负相关关系, 这一结果与多数内陆河流的研究存在差异(季世琛等, 2018; 闵文武等, 2021), 但与其他高原水环境的研究具有相似之处(刘钢等, 2018; 杨欣兰等, 2019; 梅建西, 2020)。海拔通过影响太阳辐射强度和氧气含量对原生动物群落结构产生影响。青藏高原辐射强度大(谷星月等, 2018), 从而影响到原生动物的正常生理代谢活动。此外, 相较于其他水期, 雅鲁藏布江丰水期水体含氧量较低, 对原生动物也会产生一定的影响(李红敬等, 2010)。BIOENV分析再次表明, 理化因子对雅鲁藏布江中游原生动物群落的影响是多个因子共同作用的。支流河段主要受pH、WT、ALT共同影响, 而干流河段受EC的影响最大, 这可能与河段具体地形地貌以及生境特征有关。此外, 溶解氧(DO)、水流速度(WS)等理化因子也对群落结构具有一定的影响, DO是原生动物生存所必须的因素, 其含量高低直接影响原生动物的生长、代谢、繁殖等一系列生理活动; 在水环境中, 原生动物的迁徙、移动主要依靠水体流动, WS对原生动物在水体间的扩散具有决定性作用, 对群落间的相互交融至关重要(肖佰财等, 2012; 杨敏等, 2012; 吴利等, 2015)。
参考文献
Community characteristic of protozoan in middle and upper reaches of the Lhasa River during spring season
拉萨河中上游春季原生动物群落特征
The Study of Hydrochemical Characteristics and Carbon Cycles in the Yarlung Zangbo River Basin
雅鲁藏布江水文水化学特征及流域碳循环研究
Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin
DOI:10.1186/s40168-018-0470-z [本文引用: 1]
Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2
DOI:10.1038/s41587-019-0209-9 PMID:31341288 [本文引用: 1]
A review of beta diversity studies
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 1]

<p>Beta diversity is an important component of biological diversity, measuring compositional change in species assemblages across temporal and spatial scales. Beta diversity concerns not only a number of ecological and evolutionary issues, but can also guide the selection of protected areas and help to optimize conservation networks. It has thus become a hot topic in biodiversity research in recent years. Researchers have used various measures and analytical methods to investigate patterns of beta diversity and its underlying mechanisms for various taxa and in different regions. Here, we reviewed literature from the past decade pertaining to the following aspects of beta diversity: metrics, temporal and spatial patterns, determinants and applications in biodiversity conservation. Whittaker introduced the term beta diversity in 1960, but defined it vaguely. As the concept of beta diversity evolved, a high variety of measures were developed to quantify the concept. The comparison of results from different studies may be hindered by the variety of measures used to quantify beta diversity. Presently, the most popular methods for measuring beta diversity are similarity/ dissimilarity coefficients such as Jaccard index and Sørensen index. In the last few years, several methods to quantify beta diversity have emerged, some of which are worth noting. Beta diversity depends on temporal scale, spatial scale and taxonomic scale, and decreases with increasing analytical grain size. There is no consensus among scientists that beta diversity decreases with latitude, i.e. that it is higher in tropics and lower near the poles. Beta diversity is high within mountain ranges and at the interface of biogeographic realms; thus, larger/more reserves are needed in these regions to cover the entire gradient of species turnover. Studies on beta diversity across temporal scales have shown that climatic change has resulted in shifts in species composition through time, and that the migration of species between different continents/regions has led to biotic homogenization. Based on a thorough review of beta diversity literature, we think the following questions might be the focus of future research: (1) the influence of evolutionary history and biological characteristics of different taxonomic groups on their beta diversity; (2) the influence of temporal/spatial scales on beta diversity and its determinants; and (3) the effect of anthropogenic activities on beta diversity.</p>
Beta多样性研究进展
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 1]
Beta多样性度量时空尺度上物种组成的变化, 是生物多样性的重要组成部分, 与许多生态学和进化生物学问题密切相关, 并且其信息可用于保护区选址和布局规划, 因此在最近10年间成为生物多样性研究的热点问题之一。多年来, 学者们利用各种度量方式和分析方法, 在不同地理区域, 对许多生物类群beta多样性的时空格局和形成机制进行了大量研究。本文主要从beta多样性的度量方法、时空格局、形成机制及其在生物多样性保护中的应用等几个方面, 总结了最近10多年来相关研究的进展。Whittaker(1960)最初提出beta多样性概念时就缺乏严格的定义, 随着概念的不断演化, 度量方法也同样呈现出多样化, 而度量手段的多样化非常不利于不同研究之间的比较。目前应用最普遍的度量方法是采用相似性指数, 如Jaccard和Sørensen指数。最近几年, 新的度量方法还在不断出现, 其中一些方法非常值得注意。Beta多样性具有时空尺度和分类尺度依赖性, 一般随分析粒度(grain)的增加而降低。虽然有些研究表明beta多样性随纬度增加而降低, 但学者们并没有达成共识。山区和生物地理区的交界处beta多样性都比较高, 因而需要在这些地区增加保护区的面积或者数量以囊括物种变化梯度。对时间尺度上beta多样性的研究表明, 气候变化确实导致了物种组成在时间上的变化, 并且物种在不同大陆和地区间的迁移导致了生物同质化。扩散过程和生态位过程共同决定了beta多样性, 只是这两个过程的相对重要性依尺度、地理区域和物种类群的不同而有所差异。综上所述, 我们认为未来beta多样性研究的热点问题是:(1)不同生物类群的进化历史和生物学特征对beta多样性的影响; (2)不同的时空尺度对beta多样性及其维持机制的影响; (3)人类活动对beta多样性的影响。
Progress in the biodiversity of protozoa in the deep-sea environment
深海原生动物多样性研究进展
Biogeographic distribution patterns and ecological mechanisms of benthic eukaryotic microorganisms in Jinsha River
金沙江底栖真核微生物地理分布特征及生态学机制
Interpretation of environmental factors affecting zooplanktonic beta diversity and its components in Xiangshan Bay
象山港浮游动物β多样性及其成分变化的环境因子解释
Molecular ecological network analyses
DOI:10.1186/1471-2105-13-113
PMID:22646978
[本文引用: 1]
Background: Understanding the interaction among different species within a community and their responses to environmental changes is a central goal in ecology. However, defining the network structure in a microbial community is very challenging due to their extremely high diversity and as-yet uncultivated status. Although recent advance of metagenomic technologies, such as high throughout sequencing and functional gene arrays, provide revolutionary tools for analyzing microbial community structure, it is still difficult to examine network interactions in a microbial community based on high-throughput metagenomics data.;Results: Here, we describe a novel mathematical and bioinformatics framework to construct ecological association networks named molecular ecological networks (MENs) through Random Matrix Theory (RMT)-based methods. Compared to other network construction methods, this approach is remarkable in that the network is automatically defined and robust to noise, thus providing excellent solutions to several common issues associated with high-throughput metagenomics data. We applied it to determine the network structure of microbial communities subjected to long-term experimental warming based on pyrosequencing data of 16 S rRNA genes. We showed that the constructed MENs under both warming and unwarming conditions exhibited topological features of scale free, small world and modularity, which were consistent with previously described molecular ecological networks. Eigengene analysis indicated that the eigengenes represented the module profiles relatively well. In consistency with many other studies, several major environmental traits including temperature and soil pH were found to be important in determining network interactions in the microbial communities examined. To facilitate its application by the scientific community, all these methods and statistical tools have been integrated into a comprehensive Molecular Ecological Network Analysis Pipeline (MENAP), which is open-accessible now (http://ieg2.ou.edu/MENA).;Conclusions: The RMT-based molecular ecological network analysis provides powerful tools to elucidate network interactions in microbial communities and their responses to environmental changes, which are fundamentally important for research in microbial ecology and environmental microbiology.
Zooplankton community structure and relationship to environmental factors in the Tibetan reach of the Lancang River
澜沧江西藏段浮游动物群落结构及其与环境因子的关系
Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake
DOI:10.1007/s10750-005-1233-y URL [本文引用: 1]
Selection imposed by local environmental conditions drives differences in microbial community composition across geographically distinct groundwater aquifers
DOI:10.1093/femsec/fiz160
URL
[本文引用: 1]
Several studies have analyzed biogeographic distribution patterns of microbial communities across broad spatial scales. However, it is often unclear to what extent differences in community composition across different regions are caused by dispersal limitation or selection, and if selection is caused by local environmental conditions alone or additional broad-scale region-specific factors. This is especially true for groundwater environments, which have been understudied in this context relative to other non-subsurface habitats. Here, we analyzed microbial community composition based on exact 16S rRNA amplicon sequence variants (ASVs) from four geographically separated aquifers located in different regions along a latitudinal transect of ∼700 km across Germany. Using a combination of variation partitioning and ecological null models revealed that differences in microbial community composition were mainly the product of selection imposed by local environmental conditions and to a smaller but still significant extent dispersal limitation and drift across regions. Only ∼23% of the total variation in microbial community composition remained unexplained, possibly due to underestimated effects of dispersal limitation among local communities within regions and temporal drift. No evidence was found for selection due to region-specific factors independent of local environmental conditions.
Biogeographic patterns and assembly mechanisms of bacterial communities in lakes, China—Statistical analysis based on the published literature
中国湖泊细菌群落的生物地理分布格局及驱动机制——基于文献数据的统计分析
Dominant zooplankton species and their ecological niches under different hydrologic regimes in rivers of the Pearl River Delta
珠三角河网不同水文期浮游动物优势种及生态位
Structural characteristics and seasonal changes in zooplankton community in Changhu Lake
长湖浮游动物群落结构特征及其季节变化
Characteristics of zooplankton community structure in Niyang River basin of Tibet
西藏尼洋河流域浮游动物群落结构研究
Climatic characteristics of surface radiation flux over the Qinghai-Tibetan Plateau
DOI:10.7522/j.issn.1000-0534.2018.00051
[本文引用: 1]
In this paper, we used observational radiation data from the Amdo Center Station (The following is the Amdo Station, with 19 years of surface radiation observations), BJ Station (with 17 years of surface radiation observations), D105 Station (with 17 years of surface radiation observations) and NPAM (Also known as the MS3478 Site, with 17 years of surface radiation observations) Station on the Northern Qinghai-Tibetan Plateau in the GAME/Tibet[GEWEX (Global Energy and Water cycle Experiment) Asian Monsoon Experiment on the Qinghai-Tibetan Plateau] and the CAMP/Tibet[CEOP (Coordinated Enhanced Observing Period) Asia-Australia Monsoon Project (CAMP) on the Qinghai-Tibetan Plateau], and the Qomolangma Station for Atmosphere and Environment Observation and Research of the Chinese Academy of Sciences (The following is the Qomolangma Station, with 12 years of surface radiation observations) and Nam Co Station for Multisphere Observation and Research of the Chinese Academy of Sciences (The following is the Nam Co Station, with 12 years of surface radiation observations). There are about 10 to 20 years of observational radiation data, calculated the daily average value at 10:00 to 18:00 on sunny days, and counted monthly average value. Finally, use these average value to find out the annual average value of observational surface radiation fluxes (short wave downward radiation, short wave upward radiation, long wave downward radiation, long wave upward radiation, net radiation flux and albedo) from the above sites. Based on the analysis of the annual variations of short wave downward radiation, short wave upward radiation, long wave upward radiation, long wave downward radiation, net radiation and albedo of the typical underlying surface (plateau meadows with a height of 5 cm, plateau meadows with a height of 10 cm, sparse plateau meadows, plateau meadows with a height of 15 cm, gobi, and plateau meadows near the lake), the long-term climate change characteristics of surface radiation fluxes of the Qinghai-Tibetan Plateau were obtained. It was found that the short wave downward radiation flux at most sites on the Qinghai-Tibetan Plateau has a decreasing trend of varying degrees of annual change. Moreover, the long wave upward radiation flux by all the stations has an increasing trend of varying degrees, and the long wave downward radiation flux by all the stations has an increasing trend of varying degrees. The annual variation of the net radiation flux at most stations in the Qinghai-Tibetan Plateau region is basically consistent with the annual change in the short wave downward radiation. And the albedo at most sites on the Qinghai-Tibetan Plateau decreases year by year to varying degrees.
青藏高原地表辐射通量的气候特征分析
DOI:10.7522/j.issn.1000-0534.2018.00051
[本文引用: 1]
基于对"全球能量水循环亚洲季风青藏高原试验研究"(GAME/Tibet)和"全球协调加强观测计划(CEOP)之亚澳季风青藏高原试验"(CAMP/Tibet)设在藏北高原的安多站、BJ站、D105站和NPAM站以及中国科学院珠峰站和中国科学院纳木错站10~20年晴天日间的辐射观测资料求年均值,分析了高原草甸(草高为5 cm的高原草甸,10 cm的高原草甸和高原稀疏草甸,15 cm的高原草甸)、戈壁和临湖高原草甸这些典型下垫面观测站多年观测的短波向下辐射、短波向上辐射、长波向上辐射、长波向下辐射、净辐射通量和地表反照率的年际变化,得出了青藏高原地表辐射通量的气候特征,发现高原上大部分站点观测到的短波向下辐射有不同程度的减小的年变化趋势,基本所有站点观测的长波向上辐射有不同程度的逐年增加趋势,且高原上基本所有站点观测的长波向下辐射有不同程度的增加趋势,高原地区大部分站点的净辐射通量的年变化趋势基本与短波向下辐射的年变化相一致,青藏高原大部分站点的地表反照率在不同程度上逐年减小。
Beta-diversity on geographic gradients in Britain
DOI:10.2307/5518 URL [本文引用: 1]
Study on community structure of zooplankton and water quality health condition in Puyang River basin
浦阳江流域浮游动物群落结构及其评价水质健康状况的研究
Study on the community structure of zooplankton in Biliuhe Reservoir and basin
碧流河水库及其流域河流浮游动物的群落结构研究
Diversity and seasonality dynamics of ciliate communities in four estuaries of Shenzhen, China (South China Sea)
DOI:10.3390/jmse9030260
URL
[本文引用: 1]
Ciliates are fundamental components of microzooplankton, with important ecological roles. However, ciliate communities are particularly difficult to monitor using conventional morphological approaches. New molecular tools, such as DNA metabarcoding, can facilitate the study of these communities. This study used high-throughput sequencing to examine the diversity and seasonal dynamics of ciliate communities in four estuarine ecosystems in the South China Sea from June 2019 to March 2020. The amplification of the V4 region of 18S rDNA using ciliate-specific primers identified a total of 1645 amplicon sequence variants (ASVs), corresponding to 13 ciliate classes, 97 families, and 157 genera. The dominant species across all four sampling stations were spirotrichs (including choreotrichs, oligotrichs, and stichotrichs), oligohymenophorean scuticociliates, litostomateans Didinium, and prostomateans Cryptocaryon. Significant differences in ciliate diversity and community composition in the four stations were mainly due to differences in rare, rather than abundant, ASVs. Analysis of the ciliate communities and seasonal patterns in their composition revealed that variations in habitat and environmental conditions have a greater effect than seasonal changes on community composition.
Exploring the distribution patterns and conservation approaches of biodiversity on the Qinghai Tibetan Plateau
DOI:10.17520/biods.2018064 URL [本文引用: 1]
探索青藏高原生物多样性分布格局与保育途径
DOI:10.17520/biods.2018064 [本文引用: 1]
Estimation of periphytic microalgae gross primary production with DCMU-fluorescence method in Yenisei River (Siberia, Russia)
DOI:10.1007/s10811-007-9246-8 URL [本文引用: 1]
Analyzing beta diversity: Partitioning the spatial variation of community composition data
DOI:10.1890/05-0549 URL [本文引用: 1]
The geographical structure of British bird distributions: Diversity, spatial turnover and scale
DOI:10.1046/j.0021-8790.2001.00563.x URL [本文引用: 1]
The need for richness-independent measures of turnover when delineating biogeographical regions
DOI:10.1111/jbi.12266 URL [本文引用: 1]
Protozoan community character in relation to trophic level in the Beijing section of the Juma River
DOI:10.1360/biodiv.060004
[本文引用: 1]
To study the ecological status of the Juma River, we conducted a survey of the protozoa in the Beijing section of the river during March to November 2004. A total of 310 protozoa species, belonging to 157 genera, were observed, including 134 flagellate species, 64 sarcodina, and 112 ciliate. The most species-rich orders were Euglenoidida (39 species), Kinetoplastida (25 species), Volvocida (21 species), Chrysomonadida (21 species), Hypotrichida (18 species), Scuticociliatida (17 species), and Hymenostomatida (15 species). <em>Cryptomonas ovata</em> was the dominant species, whereas <em>Cinetochilum margaritaceum</em> was subdominant. Jac-card similarity indices among the protozoan communities from the nine sampling stations varied between 0.4569 and 0.5950. After comparing the characteristics of the protozoan communities and the func-tional-trophic groups with those of other waters, we found that the percentage of Phytomastigophora, identity of dominant taxa and the functional-trophic structure of the protozoan communities indicate an oligotrophic to mesotrophic state.
拒马河北京段原生动物群落特征及其对河流营养状况的指示
DOI:10.1360/biodiv.060004
[本文引用: 1]
原生动物是水生生态系统中的重要组成部分, 具有重要的生态功能。为揭示拒马河北京段的生态现状, 作者于2004年3–11月对拒马河北京段9个采样点的原生动物群落进行了调查。共鉴定出原生动物310种, 其中鞭毛虫134种(包括植鞭毛虫96种, 动鞭毛虫38种), 肉足类64种, 纤毛虫112种。原生动物种类数较多的目有:眼虫目(39种)、动基体目(25种)、团鞭毛虫目(21种)、金滴虫目(21种)、下毛目(18种)、盾纤目(17种)、膜口目(15种)。优势种为卵形隐藻(Cryptomonas ovata), 次优势种为珍珠映毛虫(Cinetochilum margaritaceum)。9个采样点原生动物群落的Jaccard相似性系数介于0.4569和0.5950之间。通过对原生动物群落结构及营养功能类群的分析并与其他水域的资料相比较, 认为目前拒马河北京段营养水平应介于贫营养和中营养之间。
Spatio-temporal characteristics of Yarlung Zangbo River in Tibet
西藏雅鲁藏布江水质时空特征分析
Beta diversity of stream bacteria in Hengduan Mountains: The effects of climatic and environmental variables
DOI:10.17520/biods.2019390 URL [本文引用: 1]
西藏横断山区溪流细菌beta多样性组分对气候和水体环境的响应
Biogeographical distribution and community assembly of active protistan assemblages along an estuary to a basin transect of the northern South China Sea
DOI:10.3390/microorganisms9020351
URL
[本文引用: 1]
Marine protists are essential for globally critical biological processes, including the biogeochemical cycles of matter and energy. However, compared with their prokaryotic counterpart, it remains largely unclear how environmental factors determine the diversity and distribution of the active protistan communities on the regional scale. In the present study, the biodiversity, community composition, and potential drivers of the total, abundant, and rare protistan groups were studied using high throughput sequencing on the V9 hyper-variable regions of the small subunit ribosomal RNA (SSU rRNA) along an estuary to basin transect in the northern South China Sea. Overall, Bacillariophyta and Cercozoa were abundant in the surface water; heterotrophic protists including Spirotrichea and marine stramenopiles 3 (MAST-3) were more abundant in the subsurface waters near the heavily urbanized Pearl River estuary; Chlorophyta and Pelagophyceae were abundant at the deep chlorophyll maximum depth, while Hacrobia, Radiolaria, and Excavata were the abundant groups in the deep water. Salinity, followed by water depth, temperature, and other biological factors, were the primary factors controlling the distinct vertical and horizontal distribution of the total and abundant protists. Rare taxa were driven by water depth, followed by temperature, salinity, and the concentrations of PO43−. The active protistan communities were mainly driven by dispersal limitation, followed by drift and other ecological processes.
Species diversity and driving factors of benthic macroinvertebrate assemblages in the middle and lower reaches of the Yarlung Zangbo River
DOI:10.17520/biods.2021431
[本文引用: 1]
<p id="p00005"><strong>Aims:</strong> The Yarlung Zangbo River basin supports rich and unique biological resources, which makes it a global biodiversity hotspot. However, surveys on benthic macroinvertebrates in this river basin are far from sufficient. To fill this gap, this study focused on the middle and lower reaches of the Yarlung Zangbo River, where macroinvertebrates were sampled from the main stream and tributaries in autumn (October 2015) and spring (March 2016). <br><strong>Methods:</strong> One-way ANOVA was used to examine the differences of abundance, biomass and ecological indices between the main stream and tributaries. Canonical analysis of principal coordinates (CAP) was adopted to test if community structures varied among different site groups. Canonical correspondence analysis (CCA) was Applied to identify the key environmental factors that significantly influence the community structure of macroinvertebrates during each season. <br><strong>Results:</strong> A total of 270 species were identified, belonging to 5 phyla 8 classes 20 orders and 92 families. The community included 246 aquatic insects, 14 oligochaetes, 4 mollusks and 6 others. The average density was 939.1 ind./m<sup>2</sup>, and the average biomass was 5.44 g/m<sup>2</sup>. 184 and 214 macroinvertebrate species were collected in spring and autumn, respectively. The dominant species were aquatic insects that preferred clean and cold water, including <i>Baetis </i>sp., <i>Baetiella </i>sp., <i>Simulium </i>sp., <i>Micropsetra</i> sp. and <i>Brachycentrus </i>sp. The community structure, density and diversity indices exhibited significant temporal and spatial variation, and the diversity in tributaries was significantly higher than that of the main stream. CCA analysis indicated that environmental factors including altitude, velocity, river width and substrate types were key factors structuring the benthic community in the Yarlung Zangbo River. <br><strong>Conclusion:</strong> The variation in community structure and diversity pattern were mainly derived from the variable climate types and geological barriers in the Grand Canyon area. This study can provide important basis and reference for macroinvertebrate diversity assessments and environmental monitoring in the Yarlung Zangbo River basin.</p>
雅鲁藏布江中下游底栖动物物种多样性及其影响因素
DOI:10.17520/biods.2021431
[本文引用: 1]
雅鲁藏布江流域维系着丰富而独特的生物资源, 是全球生物多样性研究的热点区域。然而, 该流域底栖动物多样性的调查却极不充分。本文于2015年10月和2016年3月对雅鲁藏布江干流(朗县至墨脱段)和主要支流的底栖动物进行了调查, 并采用单因素方差分析(one-way ANOVA)和典范对应分析(canonical correspondence analysis)等对群落多样性格局进行解析。共采集到底栖动物270种, 隶属于5门8纲20目92科, 包括昆虫纲246种, 寡毛纲14种, 腹足纲4种, 其他动物6种。春季和秋季分别采集到底栖动物184种和214种, 优势种均以喜清洁和冷水的水生昆虫为主, 包括四节蜉属一种(Baetis sp.)、花翅蜉属一种(Baetiella sp.)、蚋属一种(Simulium sp.)、小突摇蚊属一种(Micropsetra sp.)和短石蛾属一种(Brachycentrus sp.)等。全流域平均密度为939.1 ind./m<sup>2</sup>,sp.)等。平均生物量为5.44 g/m<sup>2</sup>。底栖动物的物种组成、密度和多样性在季节和区域之间存在一定差异, 支流的多样性显著高于干流。典范对应分析显示, 海拔、流速、河宽和底质类型等环境因子是影响雅鲁藏布江流域底栖动物群落结构的关键环境因素, 而大峡谷地区多变的气候类型和地理阻隔是造成群落变化的根本原因。本研究可为雅鲁藏布江流域底栖动物多样性评估和环境监测提供重要的基础和参考。
Characteristics of zooplankton community structure and water quality assessment in Keluke Lake in the Qinghai-Tibetan Plateau
青藏高原可鲁克湖浮游动物群落结构特征及水质评价
Spatiotemporal variations of hydrochemistry and its controlling factors in the Yarlung Tsangpo River
雅鲁藏布江流域水化学时空变化及其控制因素
Disentangling the mechanisms shaping the surface ocean microbiota
DOI:10.1186/s40168-020-00827-8
PMID:32312331
[本文引用: 1]
The ocean microbiota modulates global biogeochemical cycles and changes in its configuration may have large-scale consequences. Yet, the underlying ecological mechanisms structuring it are unclear. Here, we investigate how fundamental ecological mechanisms (selection, dispersal and ecological drift) shape the smallest members of the tropical and subtropical surface-ocean microbiota: prokaryotes and minute eukaryotes (picoeukaryotes). Furthermore, we investigate the agents exerting abiotic selection on this assemblage as well as the spatial patterns emerging from the action of ecological mechanisms. To explore this, we analysed the composition of surface-ocean prokaryotic and picoeukaryotic communities using DNA-sequence data (16S- and 18S-rRNA genes) collected during the circumglobal expeditions Malaspina-2010 and TARA-Oceans.We found that the two main components of the tropical and subtropical surface-ocean microbiota, prokaryotes and picoeukaryotes, appear to be structured by different ecological mechanisms. Picoeukaryotic communities were predominantly structured by dispersal-limitation, while prokaryotic counterparts appeared to be shaped by the combined action of dispersal-limitation, selection and drift. Temperature-driven selection appeared as a major factor, out of a few selected factors, influencing species co-occurrence networks in prokaryotes but not in picoeukaryotes, indicating that association patterns may contribute to understand ocean microbiota structure and response to selection. Other measured abiotic variables seemed to have limited selective effects on community structure in the tropical and subtropical ocean. Picoeukaryotes displayed a higher spatial differentiation between communities and a higher distance decay when compared to prokaryotes, consistent with a scenario of higher dispersal limitation in the former after considering environmental heterogeneity. Lastly, random dynamics or drift seemed to have a more important role in structuring prokaryotic communities than picoeukaryotic counterparts.The differential action of ecological mechanisms seems to cause contrasting biogeography, in the tropical and subtropical ocean, among the smallest surface plankton, prokaryotes and picoeukaryotes. This suggests that the idiosyncrasy of the main constituents of the ocean microbiota should be considered in order to understand its current and future configuration, which is especially relevant in a context of global change, where the reaction of surface ocean plankton to temperature increase is still unclear. Video Abstract.
Biogeography of eukaryotic plankton communities along the upper Yangtze River: The potential impact of cascade dams and reservoirs
DOI:10.1016/j.jhydrol.2020.125495 URL [本文引用: 1]
Resource status and seasonal variation of plankton in the Xaitongmoin reach of the Yarlung zangbo river
雅鲁藏布江谢通门江段浮游生物资源现状及其季节动态
Geomorphic parameters and their tectonic geomorphic significance in the middle reaches of Yarlung Zangbo River, China
雅鲁藏布江中游地貌参数特征及其构造地貌意义
Can additive beta diversity be reliably partitioned into nestedness and turnover components?
DOI:10.1111/geb.12921
[本文引用: 1]
Aims Quantifying beta-diversity (differences in the composition of communities) is central to many ecological studies. There are many beta-diversity metrics, falling mostly into two approaches: variance-based (e.g., the Sorensen index), or diversity partitioning (e.g., additive beta-diversity). The former cannot be used when species-sites matrices are unavailable (which is often the case in island biogeography in particular) and only species richness data are provided. Recently, efforts have been made to partition additive beta-diversity, a metric calculated using only alpha-diversity and gamma-diversity, into nestedness and turnover components (termed here "richness-only beta-diversity partitioning"). We set out to test whether this form of beta-diversity partitioning generates interpretable results, comparable with metrics based on species incidence beta-diversity partitioning. Location Global. Time period Present day. Major taxa studied Multiple taxa. Methods We first provide a brief review of beta-diversity partitioning methods, with a particular focus on the development of richness-only beta-diversity partitioning. Second, we use 254 empirical incidence matrices (provided with the paper) sourced from the literature to measure turnover and nestedness using incidence beta-diversity partitioning, comparing the resulting values with those calculated using richness-only beta-diversity. Results We provide an account of the emergence of beta-diversity partitioning, with particular reference to the analysis of richness-only datasets, and to the definition and usage of the relevant metrics. Analytically, we report weak correlations between turnover and nestedness calculated using the two different approaches. We show that this is because identical values of alpha-diversity and gamma-diversity can correspond to incidence matrices with a range of different structures. Main conclusions Our results demonstrate that the use of richness-only beta-diversity partitioning to measure turnover and nestedness is problematic and can produce patterns unrelated to conventional measures of turnover and nestedness. We therefore recommend that more accurate definitions are adopted for these terms in future studies.
The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions
DOI:10.1016/0378-1119(88)90066-2
PMID:3224833
[本文引用: 1]
Polymerase chain reaction conditions were established for the in vitro amplification of eukaryotic small subunit ribosomal (16S-like) rRNA genes. Coding regions from algae, fungi, and protozoa were amplified from nanogram quantities of genomic DNA or recombinant plasmids containing rDNA genes. Oligodeoxynucleotides that are complementary to conserved regions at the 5' and 3' termini of eukaryotic 16S-like rRNAs were used to prime DNA synthesis in repetitive cycles of denaturation, reannealing, and DNA synthesis. The fidelity of synthesis for the amplification products was evaluated by comparisons with sequences of previously reported rRNA genes or with primer extension analyses of rRNAs. Fewer than one error per 2000 positions were observed in the amplified rRNA coding region sequences. The primary structure of the 16S-like rRNA from the marine diatom, Skeletonema costatum, was inferred from the sequence of its in vitro amplified coding region.
Community structure of zooplankton in Chishui River in autumn and its relationship with environmental factors
赤水河秋季浮游动物群落结构及其与环境因子关系
The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools
Biodiversity, evolution and adaptation of fungi in extreme environments
DOI:10.1080/11263504.2012.753134 URL [本文引用: 1]
Microbial diversity in extreme environments
DOI:10.1038/s41579-021-00648-y [本文引用: 1]
Beta-diversity partitioning: Methods, applications and perspective
DOI:10.17520/biods.2017024 URL [本文引用: 1]
Beta多样性分解: 方法、应用与展望
DOI:10.17520/biods.2017024
[本文引用: 1]
Beta多样性是指不同群落间物种组成的差异, 由物种周转(或物种替换)和嵌套(或丰富度差异)这两种过程决定。Beta多样性分解是将这两种过程对总体beta多样性的作用进行拆分, 然后分别探讨这两种过程对群落间物种组成差异的影响。2010年之后, 人们提出了beta多样性分解的方法, 其中占据主导地位的是由Andrés Baselga于2010年提出的BAS法(总体beta多样性分解为物种周转和嵌套组分)和由János Podani和Dénes Schmera于2011年以及José C. Carvalho等于2012年提出的POD法(总体beta多样性分解为物种替换和丰富度差异组分)。这两种分解方法引起了持续的争论, 促进了该领域的快速发展。作者归纳分析了2010年后有关beta多样性分解的文献后发现, 使用BAS法的论文无论在发表量和引用次数上都多于POD法(75% vs. 20%)。Beta多样性分解的研究主要集中在欧洲(45%), 研究类群则以动物(64%)为主。本文在回顾beta多样性分解方法的提出及其发展过程的基础上, 从时空维度(纬度梯度、海拔梯度、生境片断化过程以及季节和年际动态)、多样性的不同方面(物种、功能和谱系多样性)和不同生物类群之间的比较等研究角度出发, 进一步阐述了beta多样性分解方法在探讨生物多样性分布格局以及形成机制中的应用。对于beta多样性分解的研究, 我们认为需要深入探讨的问题有: (1) beta多样性分解方法的比较分析和整合; (2)结合物种多度信息探讨beta多样性及其组分的分布格局; (3)对大尺度下beta多样性分解的结果进行普适性验证。
Biogeographic distribution patterns of Diatoms in Lancang River and their key drivers
澜沧江硅藻的地理分布模式与关键驱动因素
Benthic algal communities of Taizi River and its main tributaries in relation to environmental variables
太子河流域及其主要支流底栖藻类群落与环境因子的关系
Changes in climate and runoff in the middle course area of the Yarlung Zangbo River Basin
Based on monthly data for precipitation,air temperature and runoff in and around the middle course area of the Yarlung Zangbo River Basin from 1956 to 2010,changes and runoff responses were analyzed quantitatively using wavelet analysis,correlation analysis,and multiple linear regression. We found that warming is obvious in the study area,showing that the average rate of warming is 0.31°C/10a. Trends in precipitation and runoff are highly consistent,showing a fluctuation of increase-reduce-increase-reduce. The periodic change in interval runoff and precipitation maintains a high level of consistency and at two scales of 6.0~6.5a and 15.5~16.5a,however,temperature change periodicity is weak. Precipitation is the most dominant factor controlling changes in annual runoff and the correlation coefficient was 0.89 between precipitation and runoff;however,both air temperature and precipitation are impacting runoff processes within a year,and the runoff response shows hysteresis. Generally,runoff is the result of precipitation from June to November,and air temperature has a positive effect on runoff in other periods. The magnitude of precipitation determines runoff from April to September,and as a result the runoff coefficient is below 1.0. In contrast,the runoff coefficient is much larger than 1.0 during other months because river feeding takes place,rather than precipitation. With climate warming and wetting,the runoff coefficient is gradually decreasing,indicating that the influence of factors other than precipitation on runoff is increasing in the area.
雅鲁藏布江中游地区气候要素变化及径流的响应
基于1956-2010年雅鲁藏布江流域中游及周边17个气象站逐日降水、气温数据,以及拉孜和羊村水文站逐月天然径流资料,采用小波分析、相关分析及多元线性回归等方法,定量分析了径流与气候要素变化特征及其响应关系。研究表明:①研究区增温趋势明显,平均增温速率为0.31℃/10a,径流的变化趋势与降水高度一致,均表现出增加→减少→增加→减少的波动规律;②年径流量与降水量均存在6.0~6.5a和15.5~16.5a的变化周期,而气温变化周期性稍差;③降水是研究区年径流变化最主要的控制因素,年径流量与年降水量的相关系数达0.89;但气温和降水同时影响着径流年内分配过程,且径流响应具有一定滞后性;④4-9月降水量的丰枯决定着径流量变化,径流系数小于1.0;其它月份则以非降水补给为主,径流系数远大于1.0,年径流系数呈下降趋势,非降水补给因素对径流的影响在逐渐增强。
Analysis of the beta diversity of snails in the lakes of mid-lower reaches of the Yangtze River
长江中下游湖泊螺类beta多样性分析
Spatial patterns of tree species richness in two temperate forests
DOI:10.1111/jec.2011.99.issue-6 URL [本文引用: 1]
Current change on sedimentation and control its method in middle Yalungtsangpo River
雅鲁藏布江中游地区河流泥沙近期变化及防治对策
Characteristics of community structures of zooplankton in the mainstream of Huaihe River
DOI:10.18307/2015.0521 URL [本文引用: 1]
淮河干流浮游动物群落结构特征
Abundant and rare picoeukaryotic sub-communities present contras- ting patterns in the epipelagic waters of marginal seas in the northwestern Pacific Ocean
DOI:10.1111/emi.2017.19.issue-1 URL [本文引用: 1]
Meta-zooplankton community structure and its relationship with environmental factors in Wenruitang River
温瑞塘河后生浮游动物群落结构及其与环境因子的关系
Effect of concentrations of phosphorus and nitrogen on the dominance of cyanobacteria
DOI:10.18307/2019.0518 URL [本文引用: 1]
水体氮、磷营养盐水平对蓝藻优势形成的影响
Preliminary study of community diversity of Stechlin Lake in Berlin, Germany
德国柏林市Stechlin湖原生动物群落多样性特征的初步研究
Water quality in Beijing Tonghui River, using PFU protozoan communities as indicators
利用PFU原生动物群落多样性快速监测北京通惠河水质
Response of soil protozoa to acid mine drainage in a contaminated terrace
DOI:10.1016/j.jhazmat.2021.126790 URL [本文引用: 1]
A preliminary study on effect of artificial controlled flow velocity on phytoplankton in Xiangxi Bay
人工控制条件下水流速对香溪河库湾浮游植物影响的初步研究
The community structure of crustacean zooplankton in the lower reaches and its tributary of the Ganjiang River
赣江下游及其支流浮游甲壳动物的群落结构
Spatial and temporal variation in ciliate communities and relationships with environmental conditions in the middle and upper reaches of the Lhasa River
拉萨河中上游夏秋季纤毛虫群落时空变动及其与环境的关系
Co-occurrence patterns of soybean rhizosphere microbiome at a continental scale
DOI:10.1016/j.soilbio.2017.12.011 URL [本文引用: 1]
Research progress of beta-diversity measurement methods
Beta多样性度量方法研究进展
The community structure of protozoa and the evaluation of water quality in Taihu Lake basin in Zhejiang Province
浙江省太湖流域原生动物四季群落结构及水质评价
Spatio-temporal niche of dominant protozoa species in the midstream and downstream of Lhasa River, Tibet, China
西藏拉萨河中下游原生动物优势种时空生态位
Effects of resource availability and hydrological regime on autochthonous and allochthonous carbon in the food web of a large cross-border river (China)
DOI:10.1016/j.scitotenv.2017.08.266 URL [本文引用: 1]
Functional molecular ecological networks
Effects of sediment resuspention on predation of planktivorous fish on zooplankton
沉积物再悬浮对食浮游动物鱼类捕食浮游动物的影响
Effects of nitrogen and phosphorus ratios on phytoplankton community structure in winter
DOI:10.3724/SP.J.1145.2013.00293 URL [本文引用: 1]
氮磷比对冬季浮游植物群落结构的影响

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}