



访花昆虫主要指频繁活动在显花植物上的昆虫, 涉及半翅目、膜翅目、双翅目、鳞翅目、直翅目、螳螂目等多个目(李孟楼, 2005; Sousa et al, 2021)。在自然界中, 植物的物种形成和生长发育高度依赖访花昆虫(Kehinde & Samways, 2014), 而访花昆虫作为传粉者或盗食者, 参与到植物的物种形成和进化过程(张旭凤和邵有全, 2014), 影响生态系统植物的物种组成、群落结构以及群落的稳定性(冯立超等, 2015)。在人类的粮油作物和蔬菜水果生产体系中, 蜜蜂等重要访花昆虫为农作物授粉而增加的经济效益比蜂产品的价值总和要高出几十倍甚至上百倍(张茂林等, 2011)。无论是自然界中还是人类的生产生活中, 访花昆虫均有着无可替代的关键性地位。
访花昆虫物种多样性既能体现访花昆虫之间以及访花昆虫与环境之间的复杂关系, 又能体现访花昆虫资源的丰富性(李慧蓉, 2004; 宋海天等, 2013)。它们对环境变化较为敏感, 生境、海拔、干扰强度等因子均会影响其多样性。因此, 研究访花昆虫的多样性, 评估访花昆虫多样性的影响因素, 对生态系统环境状况和特性具有指示作用, 能够为其物种共存机制以及生存环境变化的监测、评价提供重要参考(龙莎, 2008①(① 龙莎 (2008) 黑龙江省帽儿山地区访花昆虫种类及其多样性研究. 硕士学位论文, 东北林业大学, 哈尔滨); 杜秀娟等, 2021), 进一步研究访花昆虫所提供的生态服务价值提供基础资料(钦俊德和王琛柱, 2001; 韦小平等, 2020)。
燕山山脉作为京津冀地区的天然屏障和水源涵养地, 海拔梯度较大, 涵盖了森林、灌丛、草地、农田、湿地等多样的生境类型以及阔叶林、针叶林、混交林、灌木林、草丛、草原、沼泽等丰富的植被类型, 物种多样性丰富, 是我国生物多样性优先保护区域之一(
1 材料与方法
1.1 样线选择和调查方法
根据燕山地区的植被、地形地貌等特征, 调查地点选在燕山山脉覆盖的北京市怀柔区、密云区和平谷区, 河北省承德市兴隆县和唐山市遵化市以及天津市蓟州下营镇(附录1), 应用分层随机抽样法布设样线, 涵盖燕山地区所有海拔和生境类型。
用样线法和灯诱法于2019年、2020年的7-8月对燕山地区的访花昆虫进行了3次系统性调查。根据《县域昆虫多样性调查与评估技术规定》(
分子辅助鉴定采用昆虫纲通用的DNA barcoding方法(Hebert, 2003), 基于线粒体细胞色素C氧化酶亚基I (COI)的5'端部分序列作为标记, 提取物种基因组DNA, 利用昆虫通用引物对物种COⅠ基因进行扩增和测序, 将测序结果与NCBI数据库(
1.2 环境数据
记录调查地点的生态系统类型、人为干扰类型和强度、实时温度、湿度和海拔信息, 用于冗余分析(redundancy analysis)。从WorldClim (
1.3 访花昆虫多样性
Shannon-Wiener多样性指数:
式中, Pi为第i个物种的个体数占总个体数的比例, S为昆虫群落的所有物种数, A为群落内总个体数。
群落相似性系数采用Jaccard相似性系数(Jaccard, 1912)计算:
式中, a为一种生境的物种数, b为另一种生境的物种数, c为两种生境共有的物种数。
相似性系数介于0-1之间, 其值越接近1, 表明两个地区分类单元的相似性程度越高。相似性系数在1-0.75之间为极相似, 在0.75-0.50之间为中等相似, 在0.25-0.50之间为中等不相似, 在0.25以下为极不相似。
1.4 数据分析
数据经Microsoft Excel 2019整理。应用Past 4.07b进行多样性指数计算; 运用R4.1.1软件分析多样性指数与环境因子的相关性; 利用Canoco 5软件, 以环境因子作为解释变量, 以生物多样性指数作为响应变量, 数据不做转化, 生物多样性指数中心化, 进行冗余分析; 应用R4.1.1、Canoco 5和GraphPad Prism 8软件作图。
利用R软件的“Hmisc”和“corrplot”软件包分别计算出21个环境因子与研究区访花昆虫4个多样性指数之间的相关性系数, 选择合适的环境因子进行下一步分析; 利用R软件的“iNEXT”程序包估计访花昆虫群落不同生境和海拔各水平(以200 m为梯度进行分组)之间的多样性群落组成差异。基于实际采集的访花昆虫个体数和物种数关系进行稀疏外推分析, 参数q = 0时, 计算得到物种丰富度, 表示物种是否存在, 不考虑相对丰度, 值越大表明群落中物种数越多; q = 1时, 计算得到Shannon-Wiener多样性, 表示群落中常见物种的有效数量; q = 2时, 计算得到Simpson多样性, 表示群落中优势种的有效数量(Hsieh et al, 2016)。利用“ggpubr” “ggplot2” “geosphere”软件包的线性回归分析方法, 分别对挑选出来的环境因子与群落多样性的指数之间的关系进行拟合。
2 结果
2.1 燕山地区样地内访花昆虫群落组成结构
经鉴定和统计, 30个网格共采集访花昆虫1,306头, 隶属7目44科153种(附录2)。鳞翅目昆虫物种数最多, 有40种, 占总物种数的26.14%; 其中, 蛱蝶科16种, 灰蝶科8种, 弄蝶科、粉蝶科均有4种, 草螟科3种, 凤蝶科2种, 天蛾科、夜蛾科和尺蛾科均仅有1种, 东方菜粉蝶(Pieris canidia, 10头)和丝带凤蝶(Sericinus montelus, 12头)个体数量较多。半翅目昆虫的个体数最多, 有634头, 隶属于8科35种, 占总个体数的48.55%; 其中小长蝽个体数最多(343头), 盲蝽科10种, 蝽科9种, 缘蝽科7种, 长蝽科4种, 姬蝽科2种, 土蝽科、猎蝽科和地长蝽科都仅有1种。螳螂目昆虫的物种数最少, 仅2种, 有20头, 占总个体数的1.53% (表1)。
表1 燕山地区访花昆虫群落结构
Table 1
| 类群 Order | 科数 No. of families | 种数 No. of species | 个体数 No. of individuals |
|---|---|---|---|
| 螳螂目 Mantodea | 1 (2.27%) | 2 (1.32%) | 20 (1.53%) |
| 直翅目 Orthoptera | 3 (6.82%) | 4 (2.62%) | 51 (3.91%) |
| 半翅目 Hemiptera | 8 (18.18%) | 35 (22.88%) | 634 (48.55%) |
| 鞘翅目 Coleoptera | 8 (18.18%) | 33 (21.57%) | 341 (26.11%) |
| 双翅目 Diptera | 6 (13.65%) | 17 (11.11%) | 60 (4.59%) |
| 鳞翅目 Lepidoptera | 9 (20.45%) | 40 (26.14%) | 98 (7.5%) |
| 膜翅目 Hymenoptera | 9 (20.45%) | 22 (14.37%) | 102 (7.81%) |
| 合计 Total | 44 | 153 | 1,306 |
括号内是各数据占比情况
The percentage is shown in brackets.
2.2 燕山地区访花昆虫多样性
2.2.1 不同生境下访花昆虫多样性
表2 燕山地区不同生境下访花昆虫群落结构
Table 2
| 类群 Order | 湿地 Wetland | 草地 Grassland | 农田 Farmland | 灌丛 Shrub | 森林 Forest | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | |
| 螳螂目 Mantodea | 0 | 0 | 0 | 1 | 1 | 4 | 1 | 1 | 5 | 1 | 1 | 5 | 2 | 2 | 6 |
| 直翅目 Orthoptera | 0 | 0 | 0 | 2 | 2 | 9 | 2 | 2 | 6 | 3 | 4 | 29 | 2 | 2 | 7 |
| 半翅目 Hemiptera | 4 | 4 | 22 | 5 | 15 | 293 | 5 | 7 | 54 | 7 | 24 | 145 | 6 | 23 | 120 |
| 鞘翅目 Coleoptera | 3 | 3 | 15 | 6 | 15 | 139 | 3 | 5 | 15 | 6 | 21 | 106 | 7 | 17 | 66 |
| 双翅目 Diptera | 2 | 2 | 3 | 2 | 2 | 6 | 1 | 1 | 13 | 4 | 8 | 17 | 6 | 10 | 21 |
| 鳞翅目 Lepidoptera | 0 | 0 | 0 | 5 | 6 | 12 | 0 | 0 | 0 | 5 | 11 | 13 | 9 | 30 | 73 |
| 膜翅目 Hymenoptera | 1 | 1 | 1 | 5 | 6 | 10 | 3 | 3 | 12 | 6 | 12 | 49 | 5 | 8 | 30 |
| 合计 Total | 10 | 10 | 41 | 26 | 47 | 473 | 15 | 19 | 105 | 32 | 81 | 364 | 37 | 92 | 323 |
F, No. of families; S, No. of species; Ind., No. of individuals
图1
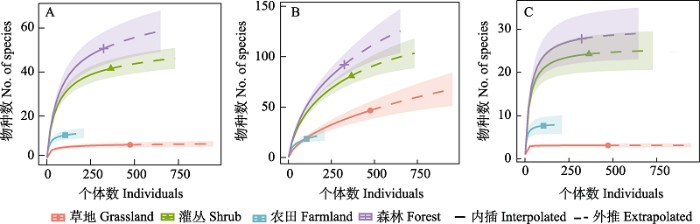
图1
燕山地区不同生境访花昆虫多样性。(A) Shannon-Wiener 多样性; (B) 物种丰富度; (C) Simpson 多样性。
Fig. 1
Diversity of flower-visiting insects under different habitats in the Yanshan area. (A) Shannon-Wiener diversity; (B) Species richnes; (C) Simpson diversity.
Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数均以灌丛最高(2.137、0.7763、0.8412), 而森林Shannon-Wiener多样性指数最低(2.022), 草地Pielou均匀度指数和Simpson优势度指数均最低(0.5629、0.7907); Margalef丰富度指数以草地为最高(4.043), 农田最低(2.837) (图2)。
图2
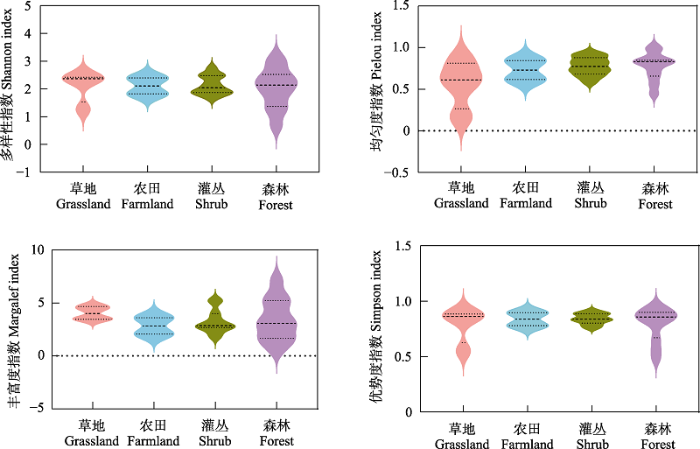
图2
燕山地区不同生境访花昆虫多样性差异
Fig. 2
Differences of flower-visiting insects diversity under different habitats in the Yanshan area
草地与农田、草地与灌丛、以及灌丛与森林的访花昆虫群落之间的相似性系数均在0.25-0.50之间, 为中等不相似, 其他生境访花昆虫群落之间的相似性系数均在0.25以下, 为极不相似。其中, 森林和灌丛的访花昆虫群落最相似, 在相似性系数为0.3008时聚为一类, 而在相似性系数为0.2692时, 农田和草地的访花昆虫群落聚为一类(图3)。
图3

图3
燕山地区不同生境访花昆虫群落相似性聚类
Fig. 3
Similarity cluster of flower-visiting insects communities under different habitats in the Yanshan area
2.2.2 燕山地区不同海拔访花昆虫多样性
表3 燕山地区不同海拔访花昆虫群落结构
Table 3
| 类群 Orders | 0-200 m | 200-400 m | 400-600 m | 600-800 m | 800-1000 m | 1000-1200 m | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | 科数 F | 种数 S | 个体数 Ind. | |
| 螳螂目 Mantodea | 1 | 1 | 9 | 2 | 2 | 2 | 1 | 1 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 直翅目 Orthoptera | 2 | 2 | 17 | 2 | 2 | 12 | 2 | 2 | 10 | 1 | 1 | 9 | 0 | 0 | 0 | 1 | 1 | 3 |
| 半翅目 Hemiptera | 5 | 13 | 296 | 6 | 14 | 44 | 8 | 24 | 166 | 7 | 20 | 57 | 3 | 4 | 7 | 6 | 11 | 64 |
| 鞘翅目 Coleoptera | 6 | 16 | 155 | 5 | 15 | 44 | 7 | 20 | 95 | 3 | 8 | 24 | 2 | 4 | 12 | 2 | 3 | 11 |
| 双翅目 Diptera | 2 | 3 | 9 | 3 | 3 | 11 | 6 | 9 | 26 | 3 | 3 | 3 | 1 | 1 | 2 | 3 | 4 | 9 |
| 鳞翅目 Lepidoptera | 5 | 6 | 12 | 6 | 9 | 12 | 8 | 19 | 39 | 7 | 19 | 35 | 0 | 0 | 0 | 0 | 0 | 0 |
| 膜翅目 Hymenoptera | 4 | 6 | 21 | 7 | 8 | 11 | 6 | 11 | 34 | 4 | 6 | 15 | 3 | 3 | 14 | 3 | 3 | 7 |
| 合计 Total | 25 | 47 | 519 | 31 | 53 | 136 | 38 | 86 | 379 | 25 | 57 | 143 | 9 | 12 | 35 | 15 | 22 | 94 |
F, No. of families; S, No. of species; Ind., No. of individuals
图4
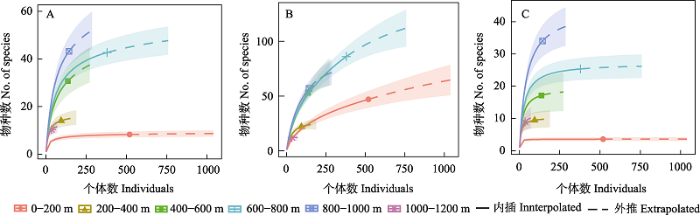
图4
燕山地区不同海拔访花昆虫多样性。(A) Shannon-Wiener 多样性; (B) 物种丰富度; (C) Simpson 多样性。
Fig. 4
Diversity of flower-visiting insects under different altitudes in Yanshan area. (A) Shannon-Wiener diversity; (B) Species richnes; (C) Simpson diversity.
在海拔范围200-400 m条件下, 访花昆虫的Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数均最高(2.267、4.019、0.8494); 在海拔800-1,000 m和1,000-1,200 m, 访花昆虫的Shannon-Wiener多样性指数和Margalef丰富度指数较低(均为1.776、1.921), 而Pielou均匀度指数较高(均为0.9115)。海拔400-600 m的Pielou均匀度指数最低(0.6734), 海拔0-200 m的Simpson优势度指数最低(0.7927) (图5)。
图5
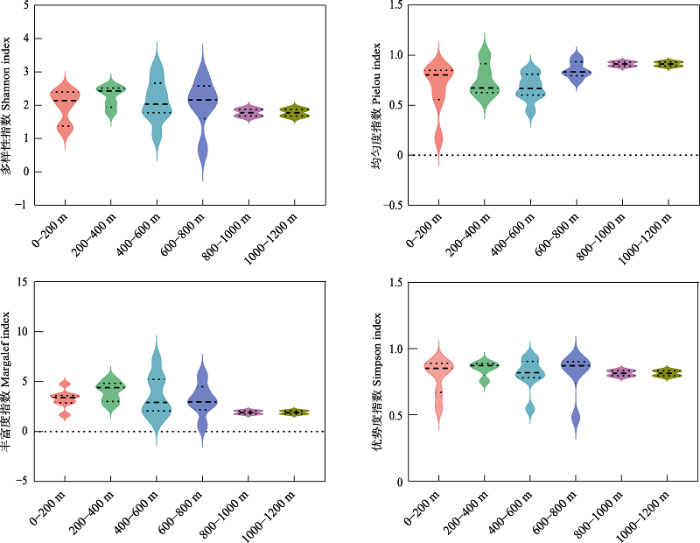
图5
燕山地区不同海拔条件访花昆虫多样性差异
Fig. 5
Differences of flower-visiting insects diversity under different altitudes in Yanshan area
低海拔0-200 m、200-400 m和400-600 m范围的访花昆虫群落之间, 以及高海拔800-1,000 m和1,000-1,200 m范围的访花昆虫群落的相似性系数均在0.25-0.50之间, 说明各海拔范围访花昆虫群落之间为中等不相似, 而其他海拔范围的访花昆虫群落之间的相似性系数均在0.25以下, 说明各海拔范围访花昆虫群落的相似性较低。其中200-400 m和400-600 m海拔范围内的访花昆虫群落最相似, 在相似性系数为0.3699时聚为一类, 而在相似性系数为0.3077时, 海拔800-1,000 m和1,000-1,200 m的访花昆虫群落聚为一类(图6)。
图6
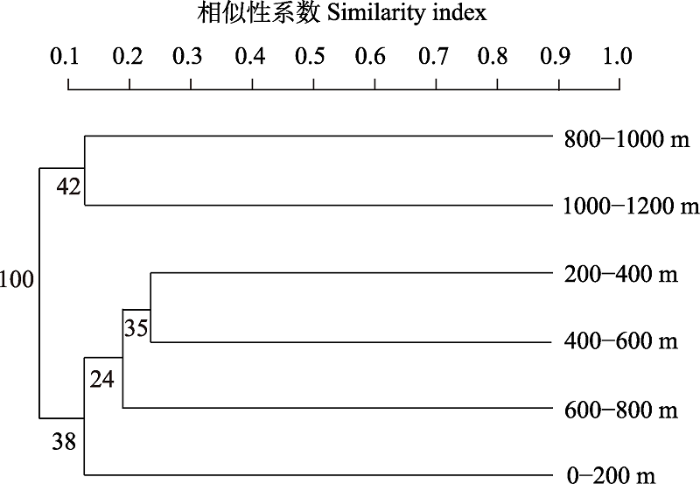
图6
燕山地区不同海拔条件访花昆虫群落相似性聚类
Fig. 6
Similarity cluster of flower-visiting insects communities under different altitudes in the Yanshan area
2.2.3 燕山地区访花昆虫多样性与环境因子的关系
冗余分析结果表明, Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数均与第1主成分负相关, 而Pielou均匀度指数与第1主成分正相关; Margalef丰富度指数与第1主成分相关性最高, 其次是Shannon-Wiener多样性指数。Shannon-Wiener多样性指数、Pielou均匀度指数以及Simpson优势度指数均与第2主成分负相关, 而Margalef丰富度指数与第2主成分正相关; Pielou均匀度指数与第2主成分相关性最高, 其次是Simpson优势度指数(图7)。
图7
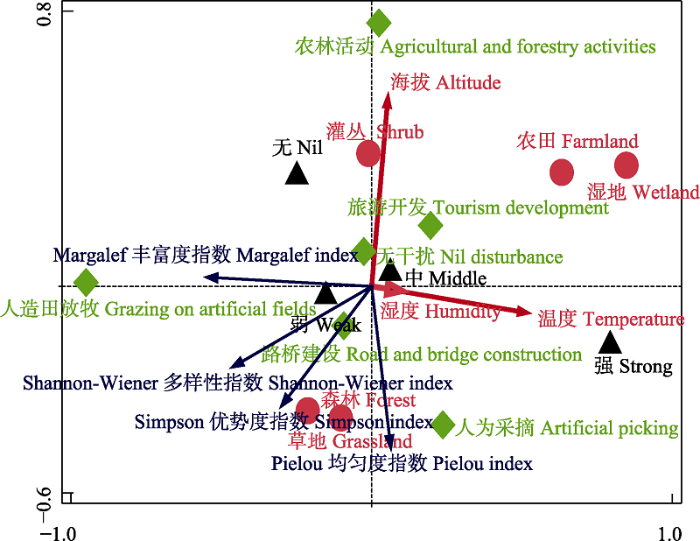
图7
燕山地区访花昆虫群落多样性与环境因子的RDA排序。  生境类型;
生境类型;  干扰程度;
干扰程度;  干扰类型。
干扰类型。
Fig. 7
RDA sequence diagram of community diversity of flower-visiting insects and five environmental factors in the Yanshan area.  Habitat type;
Habitat type;  Disturbance degree;
Disturbance degree;  Disturbance type.
Disturbance type.
RDA排序图显示, 海拔、湿度和温度3个环境因子均与第1主成分正相关; 其中, 温度与第1主成分相关性最高, 海拔和温度与第1主成分相关性极小。温度和湿度两个环境因子与第2主成分负相关, 而海拔与第2主成分正相关; 其中海拔与第2主成分相关性最高, 温度和湿度与第2主成分的相关性极小。Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数均与海拔因子负相关; Pielou均匀度指数与温度和湿度因子正相关; 而Shannon-Wiener多样性、Margalef丰富度指数和Simpson优势度却与温度和湿度因子负相关(图7)。
从RDA排序图中生境类型点的分布来看, 农田生境和湿地生境均位于第一象限, 位置相对较近, 表明这两类生境的访花昆虫群落有相似性。灌丛生境单独位于第二排序轴正半轴上, 表明与其他生境的访花昆虫群落多样性差异性较大。草地生境和森林生境位于第三象限, 且分布的位置接近, 说明它们的访花昆虫群落多样性相似度很高(图7)。
不同干扰程度和干扰类型在RDA图中的位置相对分散, 表明在不同干扰程度和类型下, 访花昆虫群落多样性差异较大。弱干扰和中度干扰在RDA排序图中位于坐标原点附近, 与各主成分相关度较小, 说明对访花昆虫各指数影响均较小, 无干扰程度下访花昆虫的群落多样性指数比弱干扰和中干扰的小一些, 强干扰对访花昆虫多样性指数影响均最大。无人为干扰对访花昆虫指数影响均最小, 其次为路桥建设和旅游开发, 农林活动和人为采摘对访花昆虫多样性指数影响均较大, 人造田放牧对Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数影响均较大(图7)。
双变量回归结果表明, 环境因子显著影响访花昆虫多样性, 但不同测度之间存在一定差异(图8)。Shannon-Wiener多样性和Margalef丰富度指数均随年降水量和最暖季降水量的增加而增大, 但年降水量和最暖季降水量对Margalef丰富度指数的解释度更高(R2 = 0.36和R2 = 0.323), 年平均气温和最冷季最低温对Margalef丰富度指数均产生负效应。Simpson优势度指数仅与坡度显著正相关(R2 = 0.142, P = 0.023), 而与其他环境因子无关。Pielou均匀度指数与所有环境因子均无统计学意义。
图8
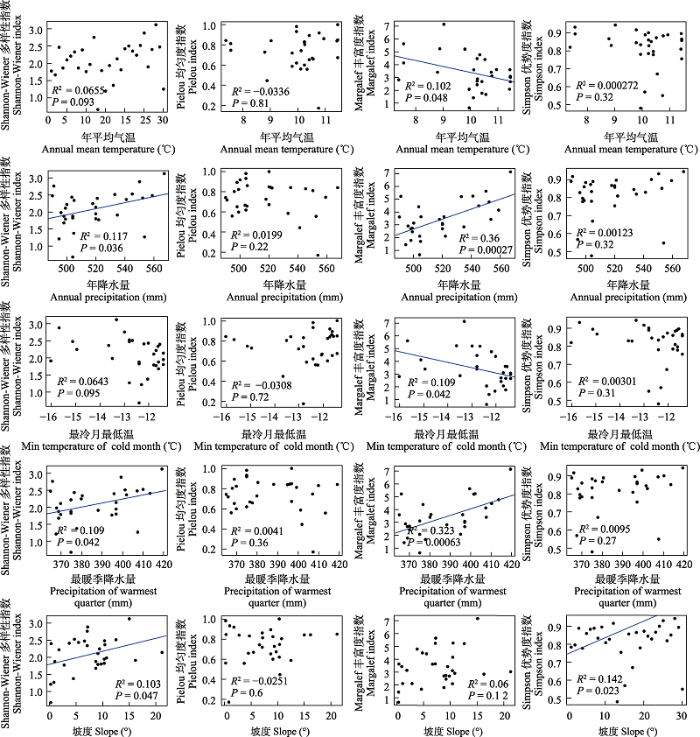
图8
访花昆虫多样性与5种环境因子的线性拟合
Fig. 8
Linear correlation fitting diagram between flower-visiting insects diversity and five environmental factor
3 讨论与结论
3.1 不同生境下访花昆虫多样性
昆虫与植物之间有着十分密切的关联(杜秀娟等, 2021), 而访花昆虫与蜜源植物之间存在很强的联系, 开花类植物的分布及数量往往对访花昆虫的分布、数量和活动的影响最具决定性, 访花昆虫的多样性相应地可以体现出开花类植物的多样性, 尤其是某特定地区的草本植物和灌木的多样性(任炳忠等, 2006; 尚利娜, 2010)。燕山地区不同生境下访花昆虫群落的组成有明显差异, 可能是不同生境中植被类型的差异所导致。由于每条样线生境下植被类型不完全一致, 访花昆虫通常在蜜源植物周围聚集分布, 对森林和灌丛生境下的多年生植物的花蜜资源利用具有明显的偏向性(李欣芸等, 2020), 灌丛访花昆虫的Shannon-Wiener多样性指数、Pielou均匀度指数以及Simpson优势度指数均最高, 说明灌丛为访花昆虫提供了较为稳定的生存环境。草地生境中访花昆虫Margalef丰富度指数最高, 但是Pielou均匀度和Simpson优势度指数最低, 且Shannon-Wiener多样性指数也较低, 说明草地的访花昆虫群落更易受到各种因子的干扰, 导致群落多样性低, 该生境中开花类草本植物多样性较高, 草本植物物种丰富度的维持对保护该生境中的访花昆虫多样性具重要意义(任炳忠等, 2012)。对于蜜源植物和草本植物的丰富度之间的相关性, 以及对访花昆虫多样性的作用仍有待进一步深入研究。
3.2 燕山地区不同海拔访花昆虫多样性
海拔梯度是山地生境差异的主导因子之一, 影响着其他环境因子, 进而影响着该地区昆虫群落结构及物种多样性(杨益春等, 2017)。访花昆虫群落结构随海拔梯度和燕山地区植被类型呈现一定的相关性(杜秀娟等, 2009; 张杰等, 2017)。访花昆虫在200-400 m海拔范围内的Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数均为最高, 说明在7-8月低海拔梯度范围的环境条件适宜许多访花昆虫的生存。处于不同高海拔梯度的昆虫物种构成差异性较大, 每个海拔梯度范围都存在各自特有的昆虫种类, 相互重叠种类较少①(① 吴艳光 (2006) 长白山地区访花昆虫多样性及访花行为的研究. 硕士学位论文, 东北师范大学, 长春.)。这种差异可能是由生境、温度、湿度及其他环境因子的不同而造成的。低海拔地区由于气候温和湿润、植被类型丰富, 昆虫群落会较为复杂; 而较高海拔地区因为温度较低、气候寒冷、植被类型呈现相对单一, 访花昆虫群落会比较简单(尚利娜等, 2012)。低海拔0-200 m、200-400 m和400-600 m范围的访花昆虫群落之间, 以及高海拔800-1,000 m和1,000-1,200 m范围的访花昆虫群落相似性系数在0.25-0.50, 为中等不相似, 表明相邻海拔的昆虫群落有着较高的相似性。可能是因为相邻海拔群落存在重叠的蜜源植物, 同种蜜源植物的开花节律基本相似, 以及泌蜜时间比较集中(刘林德等, 2002), 也可能是因为不同的昆虫飞行能力存在一定差异, 一些昆虫飞行能力较差, 很难飞入高海拔范围(刘莎等, 2018), 导致访花昆虫在不同海拔范围内有一定程度的相似性和差异性。
3.3 环境因子对燕山地区访花昆虫多样性的影响
在自然界中, 多个环境因子之间可能存有叠加和交互作用, 共同影响访花昆虫的群落结构与多样性。本研究基于实地考察对该地区干扰的类型和程度进行了划分。发现弱干扰和中干扰程度生境中的访花昆虫的种类和数量要明显高于无干扰和强干扰程度生境, 不同类型的人为干扰对访花昆虫多样性的影响均大于无干扰情况。很多山地由于人为干扰过度破坏了原有的植被, 访花昆虫生存环境遭到破坏, 导致其寄主植物的数量减少, 从而严重威胁到当地访花昆虫的多样性, 而访花昆虫多样性的下降会进一步影响相关植物的多样性, 导致物种多样性进一步下降。有研究表明, 适当的人为干扰会营造有利于访花昆虫生存的环境条件, 增加访花昆虫寄主植物的数量, 有利于提高访花昆虫多样性(王美娜等, 2018; 郑晓旭等, 2021)。
访花昆虫生存的环境温度和湿度是影响其多样性的重要因素。在RDA和双变量回归分析中, Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数会随海拔升高而减小, Pielou均匀度指数随温度和湿度的升高而增大, 而Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数却随着温度和湿度的升高而降低。相关研究表明: 在一定湿度范围内, 一些昆虫存活率随湿度的增加而变大(Wardhaugh, 1980)。Margalef丰富度指数随年平均气温和最冷季最低温升高而降低。低温不仅对昆虫的生长发育存在影响, 而且对昆虫的生殖能力和生存能力有很大影响, 随外界温度降低和暴露时间延长, 成虫抗寒能力降低, 成活率逐渐降低(Nedvěd, 1998; 冯宇倩, 2017; 宋玉锋, 2017②(② 宋玉锋 (2017) 绿豆象成虫低温耐受性及其生理生化机制研究. 硕士学位论文, 西南大学, 重庆.))。
3.4 燕山地区访花昆虫多样性保护建议
通过系统全面地研究燕山地区访花昆虫多样性, 发现调查的网格均受到不同程度和不同类型的干扰, 对访花昆虫多样性造成一定程度的影响。人造田放牧、农林活动以及人为采摘对访花昆虫影响较大, 旅游开发和路桥建设影响相对较小, 这些人为干扰破坏植被, 造成植被类型单一, 访花昆虫栖息地受到不同程度的破坏, 其生长发育和繁殖受到较大威胁, 导致访花昆虫物种多样性减少(Uhl & Brühl, 2019; 周立垚等, 2020)。我们建议: (1)增加保护区和国家公园的设立, 加强保护区内的管理; (2)政府相关部门加强监管力度, 减少开发与建设; (3)当地植保部门加强宣传, 提高当地村民保护生态环境意识, 并普及保护生物多样性和合理施用农药的相关知识。
附录 Supplementary Material
附录1 访花昆虫调查样线基本信息
Appendix 1 Information of transects for flower-visiting insects survey in the Yanshan area
附录2 燕山地区访花昆虫名录
Appendix 2 Checklist of flower-visiting insects in the Yanshan area
参考文献
Diversity of flower-visiting insects from various habitats in Shanxi Province
山西省几种生境访显花昆虫多样性分析
Differences of flower-visiting hoverfly (Diptera: Syrphidae) communities in habitats with various degrees of disturbance and altitude in Changbai Mountain, N. E. China
长白山北坡不同海拔及干扰程度下访花食蚜蝇群落的差异
Diversity and behavior of flower visitors insects of Malus komarovii in the southwest slope of Changbai Mountain
长白山西南坡山楂海棠访花昆虫多样性及其行为研究
Cold Hardiess and Adaptive Mechanism of the Anoplophora glabripennis Larva
光肩星天牛幼虫的耐寒性及其适应机制
iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers)
DOI:10.1111/mee3.2016.7.issue-12 URL [本文引用: 1]
The distribution of the flora in the alpine zone
DOI:10.1111/nph.1912.11.issue-2 URL [本文引用: 1]
Effects of vineyard management on biotic homogenization of insect-flower interaction networks in the Cape Floristic Region biodiversity hotspot
DOI:10.1007/s10841-014-9659-z URL [本文引用: 1]
Review on study of biodiversity and ecosystem functioning
生物多样性和生态系统功能研究综述
On fauna of butterflies in Chongqing and their geographic division
重庆蝶类区系与地理区划的探讨
Diversity of butterflies community and its environmental factors in Helan Mountain Nature Reserve, Ningxia
宁夏贺兰山自然保护区蝴蝶群落多样性及其环境影响因子
The relations among the nectar secretive rhythms, nectar compositions and diversities of floral visitors for both Eleutherococcus senticosus and E. sessiliflorus
刺五加、短梗五加的花蜜分泌节律、花蜜成分及访花者多样性的比较研究
Research advances on flight capacity of insect
昆虫飞行能力研究进展
Plant community diversity in Dongling Mountain, Beijing, China. II. Species richness, evenness and species diversities
北京东灵山地区植物群落多样性的研究. Ⅱ. 丰富度、均匀度和物种多样性指数
Modelling the relationship between cold injury and accumulated degree days in terrestrial arthropods
The relation of interaction between insects and plants to evolution
论昆虫与植物的相互作用和进化的关系
Relationship between floral scent of the major nectar source plants and their flower visitors in Changbai Mountain region
长白山地区优势蜜源植物花的气味与访花昆虫种类的关系
Research on pollinators in north slope of Changbai Mountain Ⅲ): Diversity of pollinators
长白山北坡访花昆虫研究(Ⅲ): 访花昆虫多样性
Coadaptation of Flowers and Their Visitors in Changbai Mountain Region
长白山地区访花昆虫与蜜源植物的协同适应
Diversity and spatio-temporal dynamics of flower visitors in the northern slope of Changbai Mountain
长白山北坡访花昆虫的多样性及时空动态
Flower-visiting insect diversity of the alien plant Erigeron annuus (Asteraceae) in Nanjing, southeastern China and an analysis of factors influencing their foraging preference
南京地区外来植物一年蓬上访花昆虫的多样性及其访花选择性的影响因素分析
Edible fruit plant species in the Amazon forest rely mostly on bees and beetles as pollinators
DOI:10.1093/jee/toaa284 URL [本文引用: 1]
The impact of pesticides on flower-visiting insects: A review with regard to European risk assessment
DOI:10.1002/etc.v38.11 URL [本文引用: 1]
Effects of woodland types with different levels of human disturbance on pollinators: A case study in Gongyi, Henan, China
不同人为干扰下林地类型对传粉昆虫的影响--以河南省巩义市为例
The effects of temperature and moisture on the inception of diapause in eggs of the Australian plague locust, Chortoicetes terminifera Walker (Orthoptera: Acrididae)
DOI:10.1111/aec.1980.5.issue-2 URL [本文引用: 1]
Pollinator diversity of Camellia oleifera forest and foraging behavior of dominant species under different habitat of Guizhou
贵州不同生境油茶林传粉昆虫的多样性及其优势种的访花行为
Effects of topographic factors on the distribution pattern of carabid species diversity in the Helan Mountains, northwestern China
地形对贺兰山步甲群落物种多样性分布格局的影响
Altitudinal variations of the reproductive strategies of Primula florindae
巨伞钟报春繁殖策略随海拔梯度的变异
The status and trend of insect diversity conservation
昆虫多样性的保护现状与趋势
Species and diversity analysis of flower-visiting insects of Asian sacred Lotus in Jiangxi, Hunan and Hubei provinces
Investigation and evaluation of insect diversity in Xingshan County, Three Gorges Reservoir Area, Hubei Province, Central China
湖北三峡库区兴山县昆虫多样性调查与评估
Effects of anthropogenic disturbance on species diversity and niche of dominant group in pollinators community
人为干扰对传粉昆虫群落物种多样性及其优势类群生态位的影响

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}