Ex-situ collections and their potential for the restoration of extinct plants
1
2020
... 作为最古老的保护策略之一, 迁地保护能够为珍稀濒危植物在其自然栖息地以外的特定区域提供生长和繁殖所需的条件.随着生境丧失、人类采伐和气候变化影响的加剧, 迁地保护的需求不断增长(Abeli et al, 2020).迁地保护与就地保护相辅相成, 通过在人为干预和精细管护下保护、研究、评价和利用植物多样性, 不仅是收集和保存珍稀濒危植物种质资源的重要方式, 也是植物回归引种及野生种群恢复重建的材料保障(Westwood et al, 2020). ...
Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology
1
2012
... 研究极小种群野生植物的种群衰退原因, 预测多种胁迫下其种群的生存动态至关重要(Guisan et al, 2013).生境质量的时空变化动态和种群的生活史特征决定了种群在持续恶化生境中的维持时间(Alexander et al, 2012).种群动态是种群统计速率与个体间交互作用的具体体现(Hart et al, 2016), 在理论模型中考虑景观结构与种群动态之间复杂的相互作用, 对于准确预测极小种群野生植物的种群轨迹十分必要.由于极小种群物种样本数量极少, 获取的个体信息难以完整反映种群的特征; 且个体之间存在强烈的相关性, 单个个体的消亡可能对整个种群的延续造成严重影响, 不符合经典的种群统计学参数估计对大样本、正态性、样本间相互独立的前提要求.因此, 陈冬东和李镇清(2020)提出, 构建极小种群物种的种群动态模型时有必要采用基于小样本的非统计分析方法, 如灰色系统理论、贝叶斯方法以及自助法(bootstrap), 来提高种群统计学参数的估计精度, 同时在模型中考虑种群的适应力也有助于更加精准地模拟环境变化下的种群动态与灭绝概率, 为极小种群野生植物保护提供更适宜的理论指导. ...
1
2014
... 很多珍稀濒危植物具有繁殖瓶颈, 繁殖过程表现出高度的脆弱性和敏感性(Baskin & Baskin, 2014).成功繁殖对于维持种群数量, 尤其是濒临灭绝物种的种群数量至关重要, 因此繁殖瓶颈的突破是极小种群野生植物解濒研究的重中之重, 发展规模化扩繁技术是其保育体系的重要环节(臧润国等, 2016).对极小种群野生植物进行高效的人工繁殖, 能够扩大种群数量并将扩繁的苗木应用于迁地保护、野外回归甚至满足商品市场的需求, 有利于其种质资源的保护和可持续利用(邓莎等, 2020). ...
Ranking threats to biodiversity and why it doesn’t matter
1
2022
... 目前人类正处于历史上前所未有的生物多样性危机之中.许多证据表明, 由于栖息地破坏、过度开发、生物入侵、气候变化和污染等的威胁(Mouillot et al, 2013), 当前的物种灭绝速度远远超过了估计的背景灭绝速度(De Vos et al, 2015).世界自然保护联盟(International Union for Conservation of Nature, IUCN)评估了142,517个物种的濒危状况, 其中40,084个被认为是受威胁种, 包括极危(Critically Endangered)、濒危(Endangered)和易危(Vulnerable)等级, 占评估物种总数的28% (IUCN, 2022).但是, 与前几次物种大灭绝不同的是, 人类既是此次物种灭绝的责任人和受害者, 也有能力成为拯救者(Bellard et al, 2022). ...
Overcoming limitations of modelling rare species by using ensembles of small models
1
2015
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
Evolutionary history of mycorrhizal symbioses and global host plant diversity
1
2018
... 生态群落中不同物种之间发生着多样化的相互作用, 种间互作是实现生态系统功能和服务的基础, 对于物种的分布、定居和长期存活至关重要.菌根共生是大多数陆生植物的共同特征, 据统计约有86%的维管植物都与菌根真菌共生, 其中兰科植物约占全部菌根植物的10% (Brundrett & Tedersoo, 2018), 菌根真菌对这些植物完成完整的生活史并成功存活必不可少.研究发现, 白旗兜兰(Paphiopedilum spicerianum)可以与多种真菌共生, 这些真菌显著提高了其成年个体忍耐干旱和环境变化的能力(Han et al, 2016).物种间的竞争强度影响着植物的存活和自然更新, 并推动着群落的演替, 植物在群落中的竞争地位是指导其保护与恢复工作的重要依据.孙哲明等(2022)通过分析7种极小种群野生植物在群落中的优势度及种内、种间竞争状况, 提出了对应的就地保护和恢复措施. ...
极小种群野生植物海南假韶子群落土壤特性研究
1
2018
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
极小种群野生植物海南假韶子群落土壤特性研究
1
2018
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
Predicting climate change impacts on the rare and endangered Horsfieldia tetratepala in China
1
2022
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
Directions in conservation biology
1
1994
... 栖息地的退化和破碎化已经使得许多物种的种群规模减少到临界水平.理论预测和实证证据均表明, 小种群的灭绝风险高于大种群(Caughley, 1994).只有了解“小种群”的形成原因, 才能制定相应的解决方案, 保持种群生存能力、防止种群灭绝(Volis & Deng, 2019).极小种群野生植物一般是由自身和外部因素共同造成的, 自身因素包括遗传多样性低、近交衰退、繁殖障碍、种子萌发率低、适应性差等, 外部因素包括地质历史事件、冰期作用、自然灾害、病虫害、气候变化、人类利用、生境退化和破碎化等(Chen et al, 2014).极小种群野生植物濒危机制的揭示是种群得以保护和恢复的重要基础.每一种极小种群野生植物都有其独特的生理生态特性和致危机理, 需要采取不同的保育措施. ...
极小种群野生植物生存力分析: 方法、问题与展望
1
2020
... 研究极小种群野生植物的种群衰退原因, 预测多种胁迫下其种群的生存动态至关重要(Guisan et al, 2013).生境质量的时空变化动态和种群的生活史特征决定了种群在持续恶化生境中的维持时间(Alexander et al, 2012).种群动态是种群统计速率与个体间交互作用的具体体现(Hart et al, 2016), 在理论模型中考虑景观结构与种群动态之间复杂的相互作用, 对于准确预测极小种群野生植物的种群轨迹十分必要.由于极小种群物种样本数量极少, 获取的个体信息难以完整反映种群的特征; 且个体之间存在强烈的相关性, 单个个体的消亡可能对整个种群的延续造成严重影响, 不符合经典的种群统计学参数估计对大样本、正态性、样本间相互独立的前提要求.因此, 陈冬东和李镇清(2020)提出, 构建极小种群物种的种群动态模型时有必要采用基于小样本的非统计分析方法, 如灰色系统理论、贝叶斯方法以及自助法(bootstrap), 来提高种群统计学参数的估计精度, 同时在模型中考虑种群的适应力也有助于更加精准地模拟环境变化下的种群动态与灭绝概率, 为极小种群野生植物保护提供更适宜的理论指导. ...
极小种群野生植物生存力分析: 方法、问题与展望
1
2020
... 研究极小种群野生植物的种群衰退原因, 预测多种胁迫下其种群的生存动态至关重要(Guisan et al, 2013).生境质量的时空变化动态和种群的生活史特征决定了种群在持续恶化生境中的维持时间(Alexander et al, 2012).种群动态是种群统计速率与个体间交互作用的具体体现(Hart et al, 2016), 在理论模型中考虑景观结构与种群动态之间复杂的相互作用, 对于准确预测极小种群野生植物的种群轨迹十分必要.由于极小种群物种样本数量极少, 获取的个体信息难以完整反映种群的特征; 且个体之间存在强烈的相关性, 单个个体的消亡可能对整个种群的延续造成严重影响, 不符合经典的种群统计学参数估计对大样本、正态性、样本间相互独立的前提要求.因此, 陈冬东和李镇清(2020)提出, 构建极小种群物种的种群动态模型时有必要采用基于小样本的非统计分析方法, 如灰色系统理论、贝叶斯方法以及自助法(bootstrap), 来提高种群统计学参数的估计精度, 同时在模型中考虑种群的适应力也有助于更加精准地模拟环境变化下的种群动态与灭绝概率, 为极小种群野生植物保护提供更适宜的理论指导. ...
Habitat heterogeneity mediates effects of individual variation on spatial species coexistence
2
2020
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
... 鉴于极小种群物种种群数量小的特点, 基于大样本方法发展的植物种群理论并不完全适用, 例如使用生态位模型或物种分布模型预测物种对全球变化的响应, 通常需要20个或更多的观测样本(van Proosdij et al, 2016).根据极小种群野生植物的分布、种群、群落、生境等调查、监测和实验数据, 发展具有针对性的假说和预测模型, 解释其小种群形成机制和恢复机制, 是亟待突破的瓶颈(Chen et al, 2020).其扩繁、回归、栖息地恢复等保护方法也可能与其他物种存在巨大差异, 很少有可以效仿的成功案例.因此, 相关保护方案必须考虑极小种群野生植物的特性, 研发基于小样本的保护实践体系(臧润国等, 2016). ...
外源激素对海南特有极小种群经济植物文昌锥生根的影响
1
2021
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
外源激素对海南特有极小种群经济植物文昌锥生根的影响
1
2021
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
基于功能性状的水杉原生母树种群生境适应策略
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
基于功能性状的水杉原生母树种群生境适应策略
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
基于MaxEnt模型的东北红豆杉潜在适生区预测
1
2022
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
基于MaxEnt模型的东北红豆杉潜在适生区预测
1
2022
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
Factors affecting the distribution pattern of Wild Plants with Extremely Small Populations in Hainan Island, China
2
2014
... 栖息地的退化和破碎化已经使得许多物种的种群规模减少到临界水平.理论预测和实证证据均表明, 小种群的灭绝风险高于大种群(Caughley, 1994).只有了解“小种群”的形成原因, 才能制定相应的解决方案, 保持种群生存能力、防止种群灭绝(Volis & Deng, 2019).极小种群野生植物一般是由自身和外部因素共同造成的, 自身因素包括遗传多样性低、近交衰退、繁殖障碍、种子萌发率低、适应性差等, 外部因素包括地质历史事件、冰期作用、自然灾害、病虫害、气候变化、人类利用、生境退化和破碎化等(Chen et al, 2014).极小种群野生植物濒危机制的揭示是种群得以保护和恢复的重要基础.每一种极小种群野生植物都有其独特的生理生态特性和致危机理, 需要采取不同的保育措施. ...
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
Importance of Plants with Extremely Small Populations (PSESPs) in endemic-rich areas, elements often forgotten in conservation strategies
1
2021
... 英文文献中, 有4篇文章第一单位为国外科研机构.其中, Wade等(2016)利用英国邱园种子信息数据库(http://data.kew.org/sid/)收集了中国120种极小种群野生植物的繁殖生物学信息, Solórzano等(2016)研究了墨西哥极小种群野生仙人掌白露 (Mammillaria albiflora)的种内遗传变异及优先保护种群, Cogoni等(2021)监测了意大利撒丁岛极小种群蕨类瓶尔小草(Ophioglossum vulgatum)的分布、种群大小、生境和就地保护效果.著名植物学家Peter Crane对中国极小种群野生植物保护计划的实施给予了高度评价, 认为该保护计划打破了理论和实践的壁垒, 能长远保护灭绝风险极高的植物(Crane, 2020). ...
Conserving our global botanical heritage: The PSESP plant conservation program
4
2020
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 随着中国国际地位的提升, 中国的环保行动也正变得越来越有影响力, 中国的自然保护事业将造福世界, 这一点毋庸置疑(Grumbine & Xu, 2011).在此背景下, 世界植物保护行动可以通过以中国极小种群野生植物保护为代表的项目, 关注稀有物种的长远未来.通过确保这些极小种群野生植物的零灭绝, 中国彰显出对全球生物多样性负责任的大国形象(Crane, 2020), 也为其他国家野生植物保护提供宝贵经验.为更好地指导极小种群野生植物保护, 本文系统总结了极小种群野生植物保护的相关理论研究和保护实践进展, 并探讨了未来的工作重点. ...
... 英文文献中, 有4篇文章第一单位为国外科研机构.其中, Wade等(2016)利用英国邱园种子信息数据库(http://data.kew.org/sid/)收集了中国120种极小种群野生植物的繁殖生物学信息, Solórzano等(2016)研究了墨西哥极小种群野生仙人掌白露 (Mammillaria albiflora)的种内遗传变异及优先保护种群, Cogoni等(2021)监测了意大利撒丁岛极小种群蕨类瓶尔小草(Ophioglossum vulgatum)的分布、种群大小、生境和就地保护效果.著名植物学家Peter Crane对中国极小种群野生植物保护计划的实施给予了高度评价, 认为该保护计划打破了理论和实践的壁垒, 能长远保护灭绝风险极高的植物(Crane, 2020). ...
... 极小种群野生植物概念的提出及相关保护规划的实施, 给濒危物种保护工作带来了全新的保护思路.这一崭新的保护理念在中国得到了各级政府部门和公众的广泛认可, 国家及各省(市、区)均颁布了极小种群野生植物的保护规划和行动措施, 各行政部门还制定了新的政策和法律法规来保护这些亟需拯救的物种(孙卫邦和徐永福, 2022).极小种群野生植物保护项目将高质量的理论研究与实际的保护行动相结合, 取得了较为成功的保护成果(Crane, 2020). ...
1
2013
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
1
2013
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
1
2012
... 鉴于全球物种和自然栖息地的加速消失, 目前对破碎化栖息地的被动保护是不够的(Heywood, 2017).野外回归是野生植物种群重建的重要途径, 作为就地保护和迁地保护之间的桥梁以及迁地保护的最终目标,其保护效果超过了单一的物种保护措施(Ren et al, 2020).在人类世, 保护的未来在于栖息地的恢复和大规模的植物回归(Volis, 2018a).作为克服生境丧失、破碎化和繁殖隔离等问题的方法, 回归已成为完整的物种恢复计划的一个重要组成部分(Dalrymple et al, 2012). ...
Estimating the normal background rate of species extinction
1
2015
... 目前人类正处于历史上前所未有的生物多样性危机之中.许多证据表明, 由于栖息地破坏、过度开发、生物入侵、气候变化和污染等的威胁(Mouillot et al, 2013), 当前的物种灭绝速度远远超过了估计的背景灭绝速度(De Vos et al, 2015).世界自然保护联盟(International Union for Conservation of Nature, IUCN)评估了142,517个物种的濒危状况, 其中40,084个被认为是受威胁种, 包括极危(Critically Endangered)、濒危(Endangered)和易危(Vulnerable)等级, 占评估物种总数的28% (IUCN, 2022).但是, 与前几次物种大灭绝不同的是, 人类既是此次物种灭绝的责任人和受害者, 也有能力成为拯救者(Bellard et al, 2022). ...
14种中国典型极小种群野生植物繁育特性和人工繁殖研究进展
3
2020
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... 很多珍稀濒危植物具有繁殖瓶颈, 繁殖过程表现出高度的脆弱性和敏感性(Baskin & Baskin, 2014).成功繁殖对于维持种群数量, 尤其是濒临灭绝物种的种群数量至关重要, 因此繁殖瓶颈的突破是极小种群野生植物解濒研究的重中之重, 发展规模化扩繁技术是其保育体系的重要环节(臧润国等, 2016).对极小种群野生植物进行高效的人工繁殖, 能够扩大种群数量并将扩繁的苗木应用于迁地保护、野外回归甚至满足商品市场的需求, 有利于其种质资源的保护和可持续利用(邓莎等, 2020). ...
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
14种中国典型极小种群野生植物繁育特性和人工繁殖研究进展
3
2020
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... 很多珍稀濒危植物具有繁殖瓶颈, 繁殖过程表现出高度的脆弱性和敏感性(Baskin & Baskin, 2014).成功繁殖对于维持种群数量, 尤其是濒临灭绝物种的种群数量至关重要, 因此繁殖瓶颈的突破是极小种群野生植物解濒研究的重中之重, 发展规模化扩繁技术是其保育体系的重要环节(臧润国等, 2016).对极小种群野生植物进行高效的人工繁殖, 能够扩大种群数量并将扩繁的苗木应用于迁地保护、野外回归甚至满足商品市场的需求, 有利于其种质资源的保护和可持续利用(邓莎等, 2020). ...
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
1
2022
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
1
2022
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
Extinction debt of high-mountain plants under twenty-first-century climate change
1
2012
... 栖息地的退化和破碎化已使许多物种的种群规模减少到临界水平.生境破坏与未来灭绝之间关系——即灭绝债(extinction debt) (在栖息地被破坏后暂时存在但注定要走向灭绝的物种数量)——的一个重要启示是, 即使生境破碎化已经停止, 它对生物多样性的影响仍将继续(Tilman et al, 1994;Dullinger et al, 2012).以小而孤立的种群为代表的许多植物物种, 长时间存活(例如大乔木)掩盖了它们真正的濒危和保护状况.因此, 种群及群落结构、动态、影响因素的分析是保护生物学研究的重点(Volis, 2018b).物种生态位、种群结构、年龄及性别组成和更新状况研究揭示了其生态需求、濒危原因、种群的生存能力和发展态势, 有助于理解种群动态和制定保护策略(Sá et al, 2018).《中华人民共和国野生植物保护条例》规定“野生植物行政主管部门应当定期组织国家重点保护野生植物和地方重点保护野生植物资源调查, 建立资源档案”.但是, 目前在保护生物学中物种的空间分布、遗传变异、保护优先性评估等相关研究占主导地位, 却忽略了种群结构统计和物种生态位分析(Volis & Deng, 2020). ...
Consequences of ex situ cultivation of plants: Genetic diversity, fitness and adaptation of the monocarpic Cynoglossum officinale L. in botanic gardens
1
2011
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
The commonness of rarity: Global and future distribution of rarity across land plants
1
2019
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
Species delimitation of the Cycas segmentifida complex (Cycadaceae) resolved by phylogenetic and distance analyses of molecular data
1
2016
... 极小种群野生植物保护名录是编制保护行动的依据, 准确的物种界定是开展种群遗传学、保护生物学、生物地理学和进化生物学等保护研究的前提(Feng et al, 2021).极小种群野生植物由于野外调查到的种群规模通常较小, 难以对其形态特征和生活史过程进行全面考察, 加上生境异质性导致不同种群存在较大的形态特征差异, 以及物种间可能存在的自然杂交现象(Tao et al, 2021), 导致名录中的部分物种在分类上存在一定困难和争议, 如桦木科的盐桦与其近缘种形态差异非常小, 其种间界定具有不确定性.针对部分物种种间界定尚未澄清的问题, 建议采用形态和分子相结合的方法, 加强分类与系统发育的研究(Feng et al, 2016).同时利用更为精细的野生种群现状及其生境条件调查数据, 删去那些经过深入的野外调查发现了新的分布点或种群、种群数量超过预期的物种, 或已经脱离了灭绝风险的物种, 对极小种群野生植物名录定期更新和完善. ...
Species delimitation with distinct methods based on molecular data to elucidate species boundaries in the Cycas taiwaniana complex (Cycadaceae)
2
2021
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
... 极小种群野生植物保护名录是编制保护行动的依据, 准确的物种界定是开展种群遗传学、保护生物学、生物地理学和进化生物学等保护研究的前提(Feng et al, 2021).极小种群野生植物由于野外调查到的种群规模通常较小, 难以对其形态特征和生活史过程进行全面考察, 加上生境异质性导致不同种群存在较大的形态特征差异, 以及物种间可能存在的自然杂交现象(Tao et al, 2021), 导致名录中的部分物种在分类上存在一定困难和争议, 如桦木科的盐桦与其近缘种形态差异非常小, 其种间界定具有不确定性.针对部分物种种间界定尚未澄清的问题, 建议采用形态和分子相结合的方法, 加强分类与系统发育的研究(Feng et al, 2016).同时利用更为精细的野生种群现状及其生境条件调查数据, 删去那些经过深入的野外调查发现了新的分布点或种群、种群数量超过预期的物种, 或已经脱离了灭绝风险的物种, 对极小种群野生植物名录定期更新和完善. ...
极小种群浙江安息香的物候观测
1
2021
... 监测是掌握种群及其生境状况并实时管控的关键手段.我国的植物监测目前主要集中在群落或植被水平, 针对植物种群的连续监测相对较少.开展种群动态和繁殖物候的长期监测有助于确定物种的受威胁因素, 制定种质资源收集时间.谭绍斌等(2014)建议在保护小区或保护点内, 利用集成了传感器、远程遥测监控和数据判读技术的物联网技术对极小种群物种开展连续和实时监测.傅国林等(2021)对浙江安息香(Styrax zhejiangensis)开展了开花、结实和种子等不同生活史阶段的物候观测, 揭示了影响其种子萌发的生物学特性. ...
极小种群浙江安息香的物候观测
1
2021
... 监测是掌握种群及其生境状况并实时管控的关键手段.我国的植物监测目前主要集中在群落或植被水平, 针对植物种群的连续监测相对较少.开展种群动态和繁殖物候的长期监测有助于确定物种的受威胁因素, 制定种质资源收集时间.谭绍斌等(2014)建议在保护小区或保护点内, 利用集成了传感器、远程遥测监控和数据判读技术的物联网技术对极小种群物种开展连续和实时监测.傅国林等(2021)对浙江安息香(Styrax zhejiangensis)开展了开花、结实和种子等不同生活史阶段的物候观测, 揭示了影响其种子萌发的生物学特性. ...
中国自然保护地整合优化关键问题
1
2021
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
中国自然保护地整合优化关键问题
1
2021
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
Genomics in conservation: case studies and bridging the gap between data and application
1
2016
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
Creating a conservation with Chinese characteristics
3
2011
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... 随着中国国际地位的提升, 中国的环保行动也正变得越来越有影响力, 中国的自然保护事业将造福世界, 这一点毋庸置疑(Grumbine & Xu, 2011).在此背景下, 世界植物保护行动可以通过以中国极小种群野生植物保护为代表的项目, 关注稀有物种的长远未来.通过确保这些极小种群野生植物的零灭绝, 中国彰显出对全球生物多样性负责任的大国形象(Crane, 2020), 也为其他国家野生植物保护提供宝贵经验.为更好地指导极小种群野生植物保护, 本文系统总结了极小种群野生植物保护的相关理论研究和保护实践进展, 并探讨了未来的工作重点. ...
... 生物多样性的消失是一个不可逆转的过程, 极小种群物种的丧失将影响其生态功能的发挥, 甚至破坏整个自然生态系统.极小种群野生植物拯救保护工程的实施以及相关的研究与实践, 帮助我们确保这些物种的零灭绝, 使我们有机会能够欣赏、研究、了解和利用它们.自2017年以来, 生物多样性保护已逐渐在中国社会主流化.2021年在中国昆明召开了联合国《生物多样性公约》第十五次缔约方大会第一阶段会议, 中国启动了国家公园和国家植物园的建设, 这些举措揭开了中国植物多样性保护的新篇章.中国具有独特的环境价值观和相关政策, 以西方模式为基础、由国际非政府组织支持的保护项目并不完全适合中国, 实施“中国特色的环境保护”更有利于保护我们国家的物种、生态系统、景观和文化多样性(Grumbine & Xu, 2011).在政府主导下, 通过完善法律法规和体制机制, 加强保育研究和实践体系建设, 推动公众参与, 深化国际合作, 为实施保护创造更有力的社会支持, 极小种群野生植物也许能够拥有一个光明的未来(Yang et al, 2020). ...
Predicting species distributions for conservation decisions
1
2013
... 研究极小种群野生植物的种群衰退原因, 预测多种胁迫下其种群的生存动态至关重要(Guisan et al, 2013).生境质量的时空变化动态和种群的生活史特征决定了种群在持续恶化生境中的维持时间(Alexander et al, 2012).种群动态是种群统计速率与个体间交互作用的具体体现(Hart et al, 2016), 在理论模型中考虑景观结构与种群动态之间复杂的相互作用, 对于准确预测极小种群野生植物的种群轨迹十分必要.由于极小种群物种样本数量极少, 获取的个体信息难以完整反映种群的特征; 且个体之间存在强烈的相关性, 单个个体的消亡可能对整个种群的延续造成严重影响, 不符合经典的种群统计学参数估计对大样本、正态性、样本间相互独立的前提要求.因此, 陈冬东和李镇清(2020)提出, 构建极小种群物种的种群动态模型时有必要采用基于小样本的非统计分析方法, 如灰色系统理论、贝叶斯方法以及自助法(bootstrap), 来提高种群统计学参数的估计精度, 同时在模型中考虑种群的适应力也有助于更加精准地模拟环境变化下的种群动态与灭绝概率, 为极小种群野生植物保护提供更适宜的理论指导. ...
Seasonal dynamics of mycorrhizal fungi in Paphiopedilum spicerianum (Rchb. F) Pfitzer—A critically endangered orchid from China
1
2016
... 生态群落中不同物种之间发生着多样化的相互作用, 种间互作是实现生态系统功能和服务的基础, 对于物种的分布、定居和长期存活至关重要.菌根共生是大多数陆生植物的共同特征, 据统计约有86%的维管植物都与菌根真菌共生, 其中兰科植物约占全部菌根植物的10% (Brundrett & Tedersoo, 2018), 菌根真菌对这些植物完成完整的生活史并成功存活必不可少.研究发现, 白旗兜兰(Paphiopedilum spicerianum)可以与多种真菌共生, 这些真菌显著提高了其成年个体忍耐干旱和环境变化的能力(Han et al, 2016).物种间的竞争强度影响着植物的存活和自然更新, 并推动着群落的演替, 植物在群落中的竞争地位是指导其保护与恢复工作的重要依据.孙哲明等(2022)通过分析7种极小种群野生植物在群落中的优势度及种内、种间竞争状况, 提出了对应的就地保护和恢复措施. ...
How variation between individuals affects species coexistence
1
2016
... 研究极小种群野生植物的种群衰退原因, 预测多种胁迫下其种群的生存动态至关重要(Guisan et al, 2013).生境质量的时空变化动态和种群的生活史特征决定了种群在持续恶化生境中的维持时间(Alexander et al, 2012).种群动态是种群统计速率与个体间交互作用的具体体现(Hart et al, 2016), 在理论模型中考虑景观结构与种群动态之间复杂的相互作用, 对于准确预测极小种群野生植物的种群轨迹十分必要.由于极小种群物种样本数量极少, 获取的个体信息难以完整反映种群的特征; 且个体之间存在强烈的相关性, 单个个体的消亡可能对整个种群的延续造成严重影响, 不符合经典的种群统计学参数估计对大样本、正态性、样本间相互独立的前提要求.因此, 陈冬东和李镇清(2020)提出, 构建极小种群物种的种群动态模型时有必要采用基于小样本的非统计分析方法, 如灰色系统理论、贝叶斯方法以及自助法(bootstrap), 来提高种群统计学参数的估计精度, 同时在模型中考虑种群的适应力也有助于更加精准地模拟环境变化下的种群动态与灭绝概率, 为极小种群野生植物保护提供更适宜的理论指导. ...
Fifty years of botanical gardens in China
1
2002
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
云南省极小种群野生植物保护研究现状——基于遗传多样性分析
1
2016
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
云南省极小种群野生植物保护研究现状——基于遗传多样性分析
1
2016
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
Plant conservation in the Anthropocene— Challenges and future prospects
1
2017
... 鉴于全球物种和自然栖息地的加速消失, 目前对破碎化栖息地的被动保护是不够的(Heywood, 2017).野外回归是野生植物种群重建的重要途径, 作为就地保护和迁地保护之间的桥梁以及迁地保护的最终目标,其保护效果超过了单一的物种保护措施(Ren et al, 2020).在人类世, 保护的未来在于栖息地的恢复和大规模的植物回归(Volis, 2018a).作为克服生境丧失、破碎化和繁殖隔离等问题的方法, 回归已成为完整的物种恢复计划的一个重要组成部分(Dalrymple et al, 2012). ...
Auxiliary seed treatment is necessary to increase recruitment of a critically endangered species
1
2022
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
1
2014
... 我国地域辽阔、地形复杂、气候多样, 优越的自然条件使得我国成为世界上生物多样性最丰富的国家之一.最新发布的《中国生物物种名录2022版》(Catalogue of Life China, http://www.sp2000.org.cn/)显示, 中国有维管植物35,214种, 约占世界维管植物多样性的10%.作为地球上种子植物的起源和分化中心之一(Lu et al, 2018), 中国拥有丰富的特有物种, 占中国种子植物物种总数的52.1% (Huang et al, 2014).由于悠久的历史、剧烈增长的人口和高速发展的经济, 中国濒危植物比例也较高, 中国高等植物红色名录评估确定了40种灭绝、野外灭绝和地区灭绝物种, 3,879种受威胁物种, 2,818种近危物种, 占被评估高等植物的18.83% (覃海宁等, 2017). ...
1
2014
... 我国地域辽阔、地形复杂、气候多样, 优越的自然条件使得我国成为世界上生物多样性最丰富的国家之一.最新发布的《中国生物物种名录2022版》(Catalogue of Life China, http://www.sp2000.org.cn/)显示, 中国有维管植物35,214种, 约占世界维管植物多样性的10%.作为地球上种子植物的起源和分化中心之一(Lu et al, 2018), 中国拥有丰富的特有物种, 占中国种子植物物种总数的52.1% (Huang et al, 2014).由于悠久的历史、剧烈增长的人口和高速发展的经济, 中国濒危植物比例也较高, 中国高等植物红色名录评估确定了40种灭绝、野外灭绝和地区灭绝物种, 3,879种受威胁物种, 2,818种近危物种, 占被评估高等植物的18.83% (覃海宁等, 2017). ...
极小种群濒危植物盐桦迁地保护研究
1
2020
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
极小种群濒危植物盐桦迁地保护研究
1
2020
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
1
2022
... 目前人类正处于历史上前所未有的生物多样性危机之中.许多证据表明, 由于栖息地破坏、过度开发、生物入侵、气候变化和污染等的威胁(Mouillot et al, 2013), 当前的物种灭绝速度远远超过了估计的背景灭绝速度(De Vos et al, 2015).世界自然保护联盟(International Union for Conservation of Nature, IUCN)评估了142,517个物种的濒危状况, 其中40,084个被认为是受威胁种, 包括极危(Critically Endangered)、濒危(Endangered)和易危(Vulnerable)等级, 占评估物种总数的28% (IUCN, 2022).但是, 与前几次物种大灭绝不同的是, 人类既是此次物种灭绝的责任人和受害者, 也有能力成为拯救者(Bellard et al, 2022). ...
极小种群海南风吹楠生态学特性及濒危成因分析
1
2018
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
极小种群海南风吹楠生态学特性及濒危成因分析
1
2018
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
极小种群野生植物崖柏的生殖物候、传粉及胚胎发育研究
1
2020
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
极小种群野生植物崖柏的生殖物候、传粉及胚胎发育研究
1
2020
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
云南蓝果树两性花的花药发育机制
1
2019
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
云南蓝果树两性花的花药发育机制
1
2019
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
Inbreeding effects in wild populations
1
2002
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
Population structure and regeneration dynamics of Firmiana major, a dominant but endangered tree species
1
2020
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
The science and economics of ex situ plant conservation
1
2009
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
1
2020
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
1
2020
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
Current conservation status and reproductive biology of the giant tree Rhododendron in China
1
2018
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
Seed dormancy, germination and storage behavior of Magnolia sinica, a plant species with extremely small populations of Magnoliaceae
1
2022
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
自毒和森林凋落物化感作用对极小种群野生植物大树杜鹃种子萌发的影响
1
2017
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
自毒和森林凋落物化感作用对极小种群野生植物大树杜鹃种子萌发的影响
1
2017
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
Translocation of threatened plants as a conservation measure in China
2
2015
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
... 回归的成功标准分为短期和长期两类, 前者包括个体的成活、种群的建立和扩散; 后者包括回归种群的自我维持和在生态系统中发挥稳定功能等, 也是回归成功的最终标准(周翔和高江云, 2011), 但这些标准对评价长寿命物种的回归成功与否可操作性不强.目前关于回归的科学监测数据非常缺乏.Liu等(2015)评估了中国154个物种的222个野外回归案例, 发现仅有不到50%的案例有个体是否存活的记录, 仅有30%的案例有关于个体是否开花或结果的记录.对建立的回归种群开展持续监测, 并与野生种群进行种群动态、繁殖能力、群落多样性等的比较, 是评价回归地点选择适宜性、物种回归实践是否成功的可靠途径.利用高通量测序技术比较野生和回归华盖木种群根际和土壤微生物, 发现二者的微生物群落组成相似, 但类群的相对多度差异显著(Shen et al, 2020; Su et al, 2021). ...
Effect of artificially accelerated aging on the vigor of Metasequoia glyptostroboides seeds
1
2020
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
极小种群野生植物海南假韶子繁育技术初步研究
1
2019
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
极小种群野生植物海南假韶子繁育技术初步研究
1
2019
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
濒危植物黄梅秤锤树野生与迁地保护种群的果实性状比较
1
2018
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
濒危植物黄梅秤锤树野生与迁地保护种群的果实性状比较
1
2018
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
The population status and threats of Taxus cuspidata, a plant species with extremely small populations in China
1
2021
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
Evolutionary history of the angiosperm flora of China
1
2018
... 我国地域辽阔、地形复杂、气候多样, 优越的自然条件使得我国成为世界上生物多样性最丰富的国家之一.最新发布的《中国生物物种名录2022版》(Catalogue of Life China, http://www.sp2000.org.cn/)显示, 中国有维管植物35,214种, 约占世界维管植物多样性的10%.作为地球上种子植物的起源和分化中心之一(Lu et al, 2018), 中国拥有丰富的特有物种, 占中国种子植物物种总数的52.1% (Huang et al, 2014).由于悠久的历史、剧烈增长的人口和高速发展的经济, 中国濒危植物比例也较高, 中国高等植物红色名录评估确定了40种灭绝、野外灭绝和地区灭绝物种, 3,879种受威胁物种, 2,818种近危物种, 占被评估高等植物的18.83% (覃海宁等, 2017). ...
五针白皮松的生态条件及其潜在分布区的分析
1
1994
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
五针白皮松的生态条件及其潜在分布区的分析
1
1994
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
极小种群野生植物坡垒的生境特征及其对幼苗多度的影响
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
极小种群野生植物坡垒的生境特征及其对幼苗多度的影响
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
Conserving plant species with extremely small populations (PSESP) in China
1
2013
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
Demographic history and identification of threats revealed by population genomic analysis provide insights into conservation for an endangered maple
1
2022
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
极小种群野生植物抢救性保护面临的机遇与挑战
1
2015
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
极小种群野生植物抢救性保护面临的机遇与挑战
1
2015
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
An Anthropocene map of genetic diversity
1
2016
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
Rare species support vulnerable functions in high-diversity ecosystems
1
2013
... 目前人类正处于历史上前所未有的生物多样性危机之中.许多证据表明, 由于栖息地破坏、过度开发、生物入侵、气候变化和污染等的威胁(Mouillot et al, 2013), 当前的物种灭绝速度远远超过了估计的背景灭绝速度(De Vos et al, 2015).世界自然保护联盟(International Union for Conservation of Nature, IUCN)评估了142,517个物种的濒危状况, 其中40,084个被认为是受威胁种, 包括极危(Critically Endangered)、濒危(Endangered)和易危(Vulnerable)等级, 占评估物种总数的28% (IUCN, 2022).但是, 与前几次物种大灭绝不同的是, 人类既是此次物种灭绝的责任人和受害者, 也有能力成为拯救者(Bellard et al, 2022). ...
化感作用的研究意义及发展前景
1
2001
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
化感作用的研究意义及发展前景
1
2001
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
Pollination ecology of Bretschneidera sinensis (Hemsley), a rare and endangered tree in China
1
2012
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
中国高等植物受威胁物种名录
1
2017
... 我国地域辽阔、地形复杂、气候多样, 优越的自然条件使得我国成为世界上生物多样性最丰富的国家之一.最新发布的《中国生物物种名录2022版》(Catalogue of Life China, http://www.sp2000.org.cn/)显示, 中国有维管植物35,214种, 约占世界维管植物多样性的10%.作为地球上种子植物的起源和分化中心之一(Lu et al, 2018), 中国拥有丰富的特有物种, 占中国种子植物物种总数的52.1% (Huang et al, 2014).由于悠久的历史、剧烈增长的人口和高速发展的经济, 中国濒危植物比例也较高, 中国高等植物红色名录评估确定了40种灭绝、野外灭绝和地区灭绝物种, 3,879种受威胁物种, 2,818种近危物种, 占被评估高等植物的18.83% (覃海宁等, 2017). ...
中国高等植物受威胁物种名录
1
2017
... 我国地域辽阔、地形复杂、气候多样, 优越的自然条件使得我国成为世界上生物多样性最丰富的国家之一.最新发布的《中国生物物种名录2022版》(Catalogue of Life China, http://www.sp2000.org.cn/)显示, 中国有维管植物35,214种, 约占世界维管植物多样性的10%.作为地球上种子植物的起源和分化中心之一(Lu et al, 2018), 中国拥有丰富的特有物种, 占中国种子植物物种总数的52.1% (Huang et al, 2014).由于悠久的历史、剧烈增长的人口和高速发展的经济, 中国濒危植物比例也较高, 中国高等植物红色名录评估确定了40种灭绝、野外灭绝和地区灭绝物种, 3,879种受威胁物种, 2,818种近危物种, 占被评估高等植物的18.83% (覃海宁等, 2017). ...
中国高等植物濒危状况评估
1
2017
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
中国高等植物濒危状况评估
1
2017
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
Planning priority conservation areas under climate change for six plant species with extremely small populations in China
1
2018
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
Relation of minimum viable population size to biology, time frame, and objective
2
2014
... 为全面掌握我国野生植物资源状况, 国家林业局于1997-2003年组织开展了第一次全国重点保护野生植物资源调查, 共调查了189种国家重点保护野生植物的资源数量、分布和受威胁状况等.调查结果显示, 百山祖冷杉(Abies beshanzuensis)、银杉(Cathaya argyrophylla)、华盖木(Pachylarnax sinica)、落叶木莲(Manglietia decidua)等55种野生植物的野外种群低于稳定存活界限的最小存活种群(minimum viable population, MVP, 指特定物种以一定概率存活一定时间所需的最少个体数量) (Reed & McCoy, 2014), 随时面临着野外灭绝的危险.其中, 稳定存活界限是指保证种群在特定时间内能稳定健康生存所需的最小阈值, 低于这个阈值种群会逐渐趋向灭绝.基于此, 国家林业局提出了极小种群野生植物(Wild Plants with Extremely Small Populations, WPESP; 也有学者翻译为Plant Species with Extremely Small Populations, PSESP)的概念: 具体是指分布地域狭窄, 长期受到外界因素胁迫干扰, 呈现出种群退化和个体数量持续减少, 种群和个体数量都极少, 已经低于稳定存活界限的最小可存活种群, 而随时濒临灭绝的野生植物(国家林业局, 2010)①(①国家林业局 (2010) 全国极小种群野生植物拯救保护工程规划(2010-2015). 非正式出版材料.).极小种群植物保育强调的对象是野生植物种群, 对于栽培种群或人工繁育数量大的物种, 如水杉(Metasequoia glyptostroboides)、东北红豆杉(Taxus cuspidata)等更要注意区分野生种群和人工种群, 否则容易在保育实践中引起困惑(臧润国, 2020).因此, 本文中极小种群野生植物统一使用WPESP的译法.《全国极小种群野生植物拯救保护工程规划(2011-2015年)》中选择首批保护极小种群野生植物的原则主要包括3个方面: 一是野外数量极小、极度濒危、随时有灭绝危险的野生植物; 二是生境要求独特、生态幅狭窄的野生植物; 三是潜在基因价值不清楚、其灭绝将引起基因流失、生物多样性降低、社会经济价值损失巨大的种群数量相对较小的野生植物.依据这些原则, 最终确定的首批120种重点保护极小种群野生植物包含36种国家I级保护植物, 26种国家II级保护植物, 59种省级重点保护植物. ...
... 极小种群野生植物概念中引入了最小存活种群(MVP)这一指标, 即特定物种以一定概率存活一定时间所需的最少个体数(Reed & McCoy, 2014).MVP值可以根据物种的生物学特性以及设定的存活概率和时间, 通过种群生存力分析(population viability analysis, PVA)求得(Rosenfeld, 2014).然而我国野生植物资源监测体系尚不完善, 难以及时掌握极小种群野生植物资源的动态变化.国内对特定野生植物生存力分析的研究也较少, 当前对于绝大多濒危物种只有种群分布和种群大小的粗略数据(图2), 很少有种群数量结构信息, 种群可存活力的工作更少(Volis, 2018b).MVP对于极小种群野生植物的物种选列和保护实践具有重要意义, 因此应该基于对种群数量和繁殖物候的长期观测, 加强对极小种群野生植物种群结构的研究和预测, 根据不同物种的生物学特性和明确的保护目标(存活概率和存活时间)来确定种群可存活力. ...
1
2020
... 鉴于全球物种和自然栖息地的加速消失, 目前对破碎化栖息地的被动保护是不够的(Heywood, 2017).野外回归是野生植物种群重建的重要途径, 作为就地保护和迁地保护之间的桥梁以及迁地保护的最终目标,其保护效果超过了单一的物种保护措施(Ren et al, 2020).在人类世, 保护的未来在于栖息地的恢复和大规模的植物回归(Volis, 2018a).作为克服生境丧失、破碎化和繁殖隔离等问题的方法, 回归已成为完整的物种恢复计划的一个重要组成部分(Dalrymple et al, 2012). ...
Advances in the reintroduction of rare and endangered wild plant species
1
2014
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
Wild plant species with extremely small populations require conservation and reintroduction in China
2
2012
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 回归已在越来越多的珍稀濒危植物保护实践中得到应用.云南省目前建立了“极小种群野生植物回归自然试验示范研究基地”, 成功开展了华盖木、毛果木莲、西畴青冈、漾濞槭等极小种群物种的野外回归工作(孙卫邦等, 2019).“典型极小种群野生植物保护与恢复技术研究”项目组建立了回归示范区6处, 回归了东北红豆杉、崖柏、坡垒、盐桦、瑶山苣苔(Dayaoshania cotinifolia)、海伦兜兰个体超过3万株.国家苏铁种质资源保护中心于2008年在广西黄连山自然保护区开展德保苏铁(Cycas debaoensis)回归试验, 回归5年后的调查表明, 德保苏铁回归种群生长良好, 17.6%的个体长出了大、小孢子叶球, 利用回归居群产生的种子培育苗木约5,300株(王运华等, 2018).截至2019年底, 中国有300个植物回归项目, 涉及206个物种, 其中中国特有物种112种.中国科学院华南植物园负责完成的“华南珍稀濒危植物的野外回归研究与应用”项目建立了“选取适当的珍稀植物进行基础研究和繁殖技术攻关, 再进行野外回归和市场化生产, 实现其有效保护, 同时通过区域生态规划及国家战略咨询, 推动整个国家珍稀濒危植物回归工作”的模式, 初步实现了珍稀濒危植物的产业化, 产生了良好的生态、社会和经济效益(Ren et al, 2012). ...
50/500 or 100/1000? Reconciling short- and long-term recovery targets and MVPs
1
2014
... 极小种群野生植物概念中引入了最小存活种群(MVP)这一指标, 即特定物种以一定概率存活一定时间所需的最少个体数(Reed & McCoy, 2014).MVP值可以根据物种的生物学特性以及设定的存活概率和时间, 通过种群生存力分析(population viability analysis, PVA)求得(Rosenfeld, 2014).然而我国野生植物资源监测体系尚不完善, 难以及时掌握极小种群野生植物资源的动态变化.国内对特定野生植物生存力分析的研究也较少, 当前对于绝大多濒危物种只有种群分布和种群大小的粗略数据(图2), 很少有种群数量结构信息, 种群可存活力的工作更少(Volis, 2018b).MVP对于极小种群野生植物的物种选列和保护实践具有重要意义, 因此应该基于对种群数量和繁殖物候的长期观测, 加强对极小种群野生植物种群结构的研究和预测, 根据不同物种的生物学特性和明确的保护目标(存活概率和存活时间)来确定种群可存活力. ...
Effects of ecological and anthropogenic factors on population demography of the harvested Butia capitata Palm in the Brazilian Cerrado
1
2020
... 栖息地的退化和破碎化已使许多物种的种群规模减少到临界水平.生境破坏与未来灭绝之间关系——即灭绝债(extinction debt) (在栖息地被破坏后暂时存在但注定要走向灭绝的物种数量)——的一个重要启示是, 即使生境破碎化已经停止, 它对生物多样性的影响仍将继续(Tilman et al, 1994;Dullinger et al, 2012).以小而孤立的种群为代表的许多植物物种, 长时间存活(例如大乔木)掩盖了它们真正的濒危和保护状况.因此, 种群及群落结构、动态、影响因素的分析是保护生物学研究的重点(Volis, 2018b).物种生态位、种群结构、年龄及性别组成和更新状况研究揭示了其生态需求、濒危原因、种群的生存能力和发展态势, 有助于理解种群动态和制定保护策略(Sá et al, 2018).《中华人民共和国野生植物保护条例》规定“野生植物行政主管部门应当定期组织国家重点保护野生植物和地方重点保护野生植物资源调查, 建立资源档案”.但是, 目前在保护生物学中物种的空间分布、遗传变异、保护优先性评估等相关研究占主导地位, 却忽略了种群结构统计和物种生态位分析(Volis & Deng, 2020). ...
Comparative analysis of fungal diversity in rhizospheric soil from wild and reintroduced Magnolia sinica estimated via high-throughput sequencing
1
2020
... 回归的成功标准分为短期和长期两类, 前者包括个体的成活、种群的建立和扩散; 后者包括回归种群的自我维持和在生态系统中发挥稳定功能等, 也是回归成功的最终标准(周翔和高江云, 2011), 但这些标准对评价长寿命物种的回归成功与否可操作性不强.目前关于回归的科学监测数据非常缺乏.Liu等(2015)评估了中国154个物种的222个野外回归案例, 发现仅有不到50%的案例有个体是否存活的记录, 仅有30%的案例有关于个体是否开花或结果的记录.对建立的回归种群开展持续监测, 并与野生种群进行种群动态、繁殖能力、群落多样性等的比较, 是评价回归地点选择适宜性、物种回归实践是否成功的可靠途径.利用高通量测序技术比较野生和回归华盖木种群根际和土壤微生物, 发现二者的微生物群落组成相似, 但类群的相对多度差异显著(Shen et al, 2020; Su et al, 2021). ...
Genetics and conservation of plant species of extremely narrow geographic range
1
2016
... 英文文献中, 有4篇文章第一单位为国外科研机构.其中, Wade等(2016)利用英国邱园种子信息数据库(http://data.kew.org/sid/)收集了中国120种极小种群野生植物的繁殖生物学信息, Solórzano等(2016)研究了墨西哥极小种群野生仙人掌白露 (Mammillaria albiflora)的种内遗传变异及优先保护种群, Cogoni等(2021)监测了意大利撒丁岛极小种群蕨类瓶尔小草(Ophioglossum vulgatum)的分布、种群大小、生境和就地保护效果.著名植物学家Peter Crane对中国极小种群野生植物保护计划的实施给予了高度评价, 认为该保护计划打破了理论和实践的壁垒, 能长远保护灭绝风险极高的植物(Crane, 2020). ...
西藏极小种群野生植物密叶红豆杉种群的性比及雌雄空间格局
1
2020
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
西藏极小种群野生植物密叶红豆杉种群的性比及雌雄空间格局
1
2020
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
Comparison of the bulk and rhizosphere soil prokaryotic communities between wild and reintroduced Manglietiastrum sinicum plants, a threatened species with extremely small populations
1
2021
... 回归的成功标准分为短期和长期两类, 前者包括个体的成活、种群的建立和扩散; 后者包括回归种群的自我维持和在生态系统中发挥稳定功能等, 也是回归成功的最终标准(周翔和高江云, 2011), 但这些标准对评价长寿命物种的回归成功与否可操作性不强.目前关于回归的科学监测数据非常缺乏.Liu等(2015)评估了中国154个物种的222个野外回归案例, 发现仅有不到50%的案例有个体是否存活的记录, 仅有30%的案例有关于个体是否开花或结果的记录.对建立的回归种群开展持续监测, 并与野生种群进行种群动态、繁殖能力、群落多样性等的比较, 是评价回归地点选择适宜性、物种回归实践是否成功的可靠途径.利用高通量测序技术比较野生和回归华盖木种群根际和土壤微生物, 发现二者的微生物群落组成相似, 但类群的相对多度差异显著(Shen et al, 2020; Su et al, 2021). ...
通过遗传多样性探讨极小种群野生植物的致濒机理及保护策略: 以裸子植物为例
1
2020
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
通过遗传多样性探讨极小种群野生植物的致濒机理及保护策略: 以裸子植物为例
1
2020
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
1
2013
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
1
2013
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
Special issue for plant species with extremely small populations
1
2016
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
论极小种群野生植物的研究及科学保护
2
2015
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
论极小种群野生植物的研究及科学保护
2
2015
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
极小种群野生植物保护研究进展与未来工作的思考
3
2021
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
... 目前针对列入《全国极小种群野生植物拯救保护工程规划》的120个物种开展的研究越来越多, 但关于单个物种的系统性研究有待增强(孙卫邦等, 2021).特定物种的系统性研究应包含以下内容: 通过极小种群野生植物的保护基因组学和生理生态适应性研究揭示其种群进化历史和濒危原因, 基于对物种生境条件、种群结构、群落组成的深入了解开展就地保护以及确定迁地地点, 在分析种群遗传结构和遗传多样性的基础上开展种质资源保护以及确定迁地保护和扩繁材料的取样数量, 通过物种繁殖生物学特性研究制定科学的有性和无性繁殖技术, 将大规模扩繁的苗木用于迁地保护和野外回归, 对迁地和回归种群开展长期监测与管护, 评估与野生种群的性状表现和适应性差异, 对保育效果进行评估并适当调整保护策略.物种的保育是一个长期的过程, 各个环节互相支撑, 离不开对特定物种的系统研究以及专项资金的长期稳定投入. ...
极小种群野生植物保护研究进展与未来工作的思考
3
2021
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
... 目前针对列入《全国极小种群野生植物拯救保护工程规划》的120个物种开展的研究越来越多, 但关于单个物种的系统性研究有待增强(孙卫邦等, 2021).特定物种的系统性研究应包含以下内容: 通过极小种群野生植物的保护基因组学和生理生态适应性研究揭示其种群进化历史和濒危原因, 基于对物种生境条件、种群结构、群落组成的深入了解开展就地保护以及确定迁地地点, 在分析种群遗传结构和遗传多样性的基础上开展种质资源保护以及确定迁地保护和扩繁材料的取样数量, 通过物种繁殖生物学特性研究制定科学的有性和无性繁殖技术, 将大规模扩繁的苗木用于迁地保护和野外回归, 对迁地和回归种群开展长期监测与管护, 评估与野生种群的性状表现和适应性差异, 对保育效果进行评估并适当调整保护策略.物种的保育是一个长期的过程, 各个环节互相支撑, 离不开对特定物种的系统研究以及专项资金的长期稳定投入. ...
极小种群野生植物中国的保护行动
2
2022
... 2017年, 国家林业局评估了120种极小种群野生植物的保护成效, 发现尽管大多数物种都实施了全面的调查, 但包括海南石豆兰(Bulbophyllum hainanense)、海南鹤顶兰(Phaius hainanensis)和海南大苞兰(Sunipia hainanensis)在内的有些类群却没有找到; 有37个物种分布在自然保护区, 60个物种通过委托护林员或当地管理部门得到保护, 为26个物种建立了就地保护小区; 对80个物种进行了迁地保护试验示范, 56个物种开展了人工繁殖, 26个物种开展了种群增强与回归试验, 对一半物种进行了种质资源保存(孙卫邦和徐永福, 2022).国家林业和草原局于2012-2018年组织开展的第二次全国重点保护野生植物资源调查发现, 与第一次调查有可比性的54种极小种群野生植物中, 有36种的野外种群数量稳中有升, 占67% (http://www.forestry.gov.cn/main/586/20220210/091653623218205.html).可见, 尽管取得了一些可喜的成果, 但拯救规划的目标仅部分实现, 资金不足、缺乏保护生物学的科学指导和专业实践人员等原因影响了拯救规划工程的实施效果(Yang et al, 2020).极小种群野生植物保育实践的论文主要涉及就地保护、迁地保护与种质资源保存、野外回归、人工繁育、标准化体系等5个方面(图3). ...
... 极小种群野生植物概念的提出及相关保护规划的实施, 给濒危物种保护工作带来了全新的保护思路.这一崭新的保护理念在中国得到了各级政府部门和公众的广泛认可, 国家及各省(市、区)均颁布了极小种群野生植物的保护规划和行动措施, 各行政部门还制定了新的政策和法律法规来保护这些亟需拯救的物种(孙卫邦和徐永福, 2022).极小种群野生植物保护项目将高质量的理论研究与实际的保护行动相结合, 取得了较为成功的保护成果(Crane, 2020). ...
极小种群野生植物中国的保护行动
2
2022
... 2017年, 国家林业局评估了120种极小种群野生植物的保护成效, 发现尽管大多数物种都实施了全面的调查, 但包括海南石豆兰(Bulbophyllum hainanense)、海南鹤顶兰(Phaius hainanensis)和海南大苞兰(Sunipia hainanensis)在内的有些类群却没有找到; 有37个物种分布在自然保护区, 60个物种通过委托护林员或当地管理部门得到保护, 为26个物种建立了就地保护小区; 对80个物种进行了迁地保护试验示范, 56个物种开展了人工繁殖, 26个物种开展了种群增强与回归试验, 对一半物种进行了种质资源保存(孙卫邦和徐永福, 2022).国家林业和草原局于2012-2018年组织开展的第二次全国重点保护野生植物资源调查发现, 与第一次调查有可比性的54种极小种群野生植物中, 有36种的野外种群数量稳中有升, 占67% (http://www.forestry.gov.cn/main/586/20220210/091653623218205.html).可见, 尽管取得了一些可喜的成果, 但拯救规划的目标仅部分实现, 资金不足、缺乏保护生物学的科学指导和专业实践人员等原因影响了拯救规划工程的实施效果(Yang et al, 2020).极小种群野生植物保育实践的论文主要涉及就地保护、迁地保护与种质资源保存、野外回归、人工繁育、标准化体系等5个方面(图3). ...
... 极小种群野生植物概念的提出及相关保护规划的实施, 给濒危物种保护工作带来了全新的保护思路.这一崭新的保护理念在中国得到了各级政府部门和公众的广泛认可, 国家及各省(市、区)均颁布了极小种群野生植物的保护规划和行动措施, 各行政部门还制定了新的政策和法律法规来保护这些亟需拯救的物种(孙卫邦和徐永福, 2022).极小种群野生植物保护项目将高质量的理论研究与实际的保护行动相结合, 取得了较为成功的保护成果(Crane, 2020). ...
7
2019
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
... 对于极小种群野生植物来说, 通过迁地保护收集和保存种质资源, 为就地保护和野外回归提供了至关重要的备份.云南省已对61种极小种群野生植物开展了迁地保护; 昆明植物园极小种群野生植物专类园迁地保存了44种极小种群植物物种, 部分物种已开花结实; 中国西南野生生物种质资源库保存了极小种群野生植物种子20种94份、DNA材料28种156份(孙卫邦等, 2019).华东师范大学收集了包括水杉、东北红豆杉、天目铁木等9种极小种群物种在内的种质资源1,500份, 并在湖北利川建立了种质资源圃.十三五重点研发计划“典型极小种群野生植物保护与恢复技术研究”在不同气候区建立了8处迁地保护示范基地, 迁地保存了14种极小种群物种的27,215株个体(臧润国和黄继红, 2020). ...
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
... 回归已在越来越多的珍稀濒危植物保护实践中得到应用.云南省目前建立了“极小种群野生植物回归自然试验示范研究基地”, 成功开展了华盖木、毛果木莲、西畴青冈、漾濞槭等极小种群物种的野外回归工作(孙卫邦等, 2019).“典型极小种群野生植物保护与恢复技术研究”项目组建立了回归示范区6处, 回归了东北红豆杉、崖柏、坡垒、盐桦、瑶山苣苔(Dayaoshania cotinifolia)、海伦兜兰个体超过3万株.国家苏铁种质资源保护中心于2008年在广西黄连山自然保护区开展德保苏铁(Cycas debaoensis)回归试验, 回归5年后的调查表明, 德保苏铁回归种群生长良好, 17.6%的个体长出了大、小孢子叶球, 利用回归居群产生的种子培育苗木约5,300株(王运华等, 2018).截至2019年底, 中国有300个植物回归项目, 涉及206个物种, 其中中国特有物种112种.中国科学院华南植物园负责完成的“华南珍稀濒危植物的野外回归研究与应用”项目建立了“选取适当的珍稀植物进行基础研究和繁殖技术攻关, 再进行野外回归和市场化生产, 实现其有效保护, 同时通过区域生态规划及国家战略咨询, 推动整个国家珍稀濒危植物回归工作”的模式, 初步实现了珍稀濒危植物的产业化, 产生了良好的生态、社会和经济效益(Ren et al, 2012). ...
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
... 随着经济发展、资源开发规模的日益扩大, 珍稀濒危植物及其生境遭受的干扰和破坏难以遏止.尽管目前已经成功挽救了部分物种(孙卫邦等, 2019), 但目前有关极小种群野生植物濒危原因和解濒技术的研究还较为缺乏, 不能满足全国极小种群野生植物拯救保护工程有效实施的科技需求(臧润国等, 2020).基于国内外最先进的保护理论和方法, 在开展大量技术实践示范基础上, 建设极小种群野生植物保护和恢复的标准化技术体系, 建立相应的评估管理规范, 能够填补在各保育环节存在的技术空白, 有助于开展并推广积极有效的保护和恢复措施. ...
7
2019
... 此外, 《云南省极小种群野生植物保护实践与探索》(孙卫邦, 2013)和《云南省极小种群野生植物研究与保护》(孙卫邦等, 2019)两本专著对极小种群野生植物的概念、特点、价值及保护意义进行了梳理和论述, 总结了云南省在极小种群野生植物保护与研究方面的成果, 并对苏铁属(Cycas spp.)植物及华盖木等7个物种以典型保护实践案例的形式进行了系统研究和保护措施的分析和综述.《珍稀濒危植物种群与保护生物学研究》基于珍稀濒危植物(尤其是极小种群野生植物)保护与恢复的基本科学理论, 全面总结了珍稀濒危植物种群和保护生物学近年来的研究成果和发展态势, 并系统介绍了包括9种极小种群野生植物在内的23种珍稀濒危植物的保护生物学实践实例(董鸣和臧润国, 2021).这些专著也能够为其他极小种群野生植物的保护提供重要参考. ...
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
... 对于极小种群野生植物来说, 通过迁地保护收集和保存种质资源, 为就地保护和野外回归提供了至关重要的备份.云南省已对61种极小种群野生植物开展了迁地保护; 昆明植物园极小种群野生植物专类园迁地保存了44种极小种群植物物种, 部分物种已开花结实; 中国西南野生生物种质资源库保存了极小种群野生植物种子20种94份、DNA材料28种156份(孙卫邦等, 2019).华东师范大学收集了包括水杉、东北红豆杉、天目铁木等9种极小种群物种在内的种质资源1,500份, 并在湖北利川建立了种质资源圃.十三五重点研发计划“典型极小种群野生植物保护与恢复技术研究”在不同气候区建立了8处迁地保护示范基地, 迁地保存了14种极小种群物种的27,215株个体(臧润国和黄继红, 2020). ...
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
... 回归已在越来越多的珍稀濒危植物保护实践中得到应用.云南省目前建立了“极小种群野生植物回归自然试验示范研究基地”, 成功开展了华盖木、毛果木莲、西畴青冈、漾濞槭等极小种群物种的野外回归工作(孙卫邦等, 2019).“典型极小种群野生植物保护与恢复技术研究”项目组建立了回归示范区6处, 回归了东北红豆杉、崖柏、坡垒、盐桦、瑶山苣苔(Dayaoshania cotinifolia)、海伦兜兰个体超过3万株.国家苏铁种质资源保护中心于2008年在广西黄连山自然保护区开展德保苏铁(Cycas debaoensis)回归试验, 回归5年后的调查表明, 德保苏铁回归种群生长良好, 17.6%的个体长出了大、小孢子叶球, 利用回归居群产生的种子培育苗木约5,300株(王运华等, 2018).截至2019年底, 中国有300个植物回归项目, 涉及206个物种, 其中中国特有物种112种.中国科学院华南植物园负责完成的“华南珍稀濒危植物的野外回归研究与应用”项目建立了“选取适当的珍稀植物进行基础研究和繁殖技术攻关, 再进行野外回归和市场化生产, 实现其有效保护, 同时通过区域生态规划及国家战略咨询, 推动整个国家珍稀濒危植物回归工作”的模式, 初步实现了珍稀濒危植物的产业化, 产生了良好的生态、社会和经济效益(Ren et al, 2012). ...
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
... 随着经济发展、资源开发规模的日益扩大, 珍稀濒危植物及其生境遭受的干扰和破坏难以遏止.尽管目前已经成功挽救了部分物种(孙卫邦等, 2019), 但目前有关极小种群野生植物濒危原因和解濒技术的研究还较为缺乏, 不能满足全国极小种群野生植物拯救保护工程有效实施的科技需求(臧润国等, 2020).基于国内外最先进的保护理论和方法, 在开展大量技术实践示范基础上, 建设极小种群野生植物保护和恢复的标准化技术体系, 建立相应的评估管理规范, 能够填补在各保育环节存在的技术空白, 有助于开展并推广积极有效的保护和恢复措施. ...
湖北省极小种群野生植物在原生群落中的竞争地位及保护建议
1
2022
... 生态群落中不同物种之间发生着多样化的相互作用, 种间互作是实现生态系统功能和服务的基础, 对于物种的分布、定居和长期存活至关重要.菌根共生是大多数陆生植物的共同特征, 据统计约有86%的维管植物都与菌根真菌共生, 其中兰科植物约占全部菌根植物的10% (Brundrett & Tedersoo, 2018), 菌根真菌对这些植物完成完整的生活史并成功存活必不可少.研究发现, 白旗兜兰(Paphiopedilum spicerianum)可以与多种真菌共生, 这些真菌显著提高了其成年个体忍耐干旱和环境变化的能力(Han et al, 2016).物种间的竞争强度影响着植物的存活和自然更新, 并推动着群落的演替, 植物在群落中的竞争地位是指导其保护与恢复工作的重要依据.孙哲明等(2022)通过分析7种极小种群野生植物在群落中的优势度及种内、种间竞争状况, 提出了对应的就地保护和恢复措施. ...
湖北省极小种群野生植物在原生群落中的竞争地位及保护建议
1
2022
... 生态群落中不同物种之间发生着多样化的相互作用, 种间互作是实现生态系统功能和服务的基础, 对于物种的分布、定居和长期存活至关重要.菌根共生是大多数陆生植物的共同特征, 据统计约有86%的维管植物都与菌根真菌共生, 其中兰科植物约占全部菌根植物的10% (Brundrett & Tedersoo, 2018), 菌根真菌对这些植物完成完整的生活史并成功存活必不可少.研究发现, 白旗兜兰(Paphiopedilum spicerianum)可以与多种真菌共生, 这些真菌显著提高了其成年个体忍耐干旱和环境变化的能力(Han et al, 2016).物种间的竞争强度影响着植物的存活和自然更新, 并推动着群落的演替, 植物在群落中的竞争地位是指导其保护与恢复工作的重要依据.孙哲明等(2022)通过分析7种极小种群野生植物在群落中的优势度及种内、种间竞争状况, 提出了对应的就地保护和恢复措施. ...
物联网在云南极小种群野生植物保护中的应用研究
1
2014
... 监测是掌握种群及其生境状况并实时管控的关键手段.我国的植物监测目前主要集中在群落或植被水平, 针对植物种群的连续监测相对较少.开展种群动态和繁殖物候的长期监测有助于确定物种的受威胁因素, 制定种质资源收集时间.谭绍斌等(2014)建议在保护小区或保护点内, 利用集成了传感器、远程遥测监控和数据判读技术的物联网技术对极小种群物种开展连续和实时监测.傅国林等(2021)对浙江安息香(Styrax zhejiangensis)开展了开花、结实和种子等不同生活史阶段的物候观测, 揭示了影响其种子萌发的生物学特性. ...
物联网在云南极小种群野生植物保护中的应用研究
1
2014
... 监测是掌握种群及其生境状况并实时管控的关键手段.我国的植物监测目前主要集中在群落或植被水平, 针对植物种群的连续监测相对较少.开展种群动态和繁殖物候的长期监测有助于确定物种的受威胁因素, 制定种质资源收集时间.谭绍斌等(2014)建议在保护小区或保护点内, 利用集成了传感器、远程遥测监控和数据判读技术的物联网技术对极小种群物种开展连续和实时监测.傅国林等(2021)对浙江安息香(Styrax zhejiangensis)开展了开花、结实和种子等不同生活史阶段的物候观测, 揭示了影响其种子萌发的生物学特性. ...
极小种群野生植物海伦兜兰的地理分布及生境调查
1
2022
... 部分极小种群野生植物由于被发现的时间短, 加之分布范围狭窄、资源稀少, 对其种群结构、群落组成及生境条件的描述较为粗略, 准确性和科学性也有待考证.对其地理分布和主要生态特征的详细调查能够为进一步的引种栽培和种质资源保护提供科学依据, 是开展有效保护的首要条件.唐凤鸾等(2022)通过对海伦兜兰(Paphiopedilum helenae)的系统调查, 共发现野生居群5个、植株162株, 较文献报道的35个植株显著增加.调查同时发现海伦兜兰生境完整、受干扰程度较低、适应性强, 自然更新困难是其致濒的主要原因, 对开展针对性保育具有重要意义.对野外仅记载5株的漾濞槭(A. yangbiense)充分调查后发现了12个种群共577株个体, 为评估该物种的受威胁状况和优化保护方案提供了科学数据(Tao et al, 2020). ...
极小种群野生植物海伦兜兰的地理分布及生境调查
1
2022
... 部分极小种群野生植物由于被发现的时间短, 加之分布范围狭窄、资源稀少, 对其种群结构、群落组成及生境条件的描述较为粗略, 准确性和科学性也有待考证.对其地理分布和主要生态特征的详细调查能够为进一步的引种栽培和种质资源保护提供科学依据, 是开展有效保护的首要条件.唐凤鸾等(2022)通过对海伦兜兰(Paphiopedilum helenae)的系统调查, 共发现野生居群5个、植株162株, 较文献报道的35个植株显著增加.调查同时发现海伦兜兰生境完整、受干扰程度较低、适应性强, 自然更新困难是其致濒的主要原因, 对开展针对性保育具有重要意义.对野外仅记载5株的漾濞槭(A. yangbiense)充分调查后发现了12个种群共577株个体, 为评估该物种的受威胁状况和优化保护方案提供了科学数据(Tao et al, 2020). ...
In?situ and ex situ pollination biology of the four threatened plant species and the significance for conservation
1
2020
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
A tree species with an extremely small population: Recategorizing the critically endangered Acer yangbiense
1
2020
... 部分极小种群野生植物由于被发现的时间短, 加之分布范围狭窄、资源稀少, 对其种群结构、群落组成及生境条件的描述较为粗略, 准确性和科学性也有待考证.对其地理分布和主要生态特征的详细调查能够为进一步的引种栽培和种质资源保护提供科学依据, 是开展有效保护的首要条件.唐凤鸾等(2022)通过对海伦兜兰(Paphiopedilum helenae)的系统调查, 共发现野生居群5个、植株162株, 较文献报道的35个植株显著增加.调查同时发现海伦兜兰生境完整、受干扰程度较低、适应性强, 自然更新困难是其致濒的主要原因, 对开展针对性保育具有重要意义.对野外仅记载5株的漾濞槭(A. yangbiense)充分调查后发现了12个种群共577株个体, 为评估该物种的受威胁状况和优化保护方案提供了科学数据(Tao et al, 2020). ...
Genome-wide evidence for complex hybridization and demographic history in a group of Cycas from China
1
2021
... 极小种群野生植物保护名录是编制保护行动的依据, 准确的物种界定是开展种群遗传学、保护生物学、生物地理学和进化生物学等保护研究的前提(Feng et al, 2021).极小种群野生植物由于野外调查到的种群规模通常较小, 难以对其形态特征和生活史过程进行全面考察, 加上生境异质性导致不同种群存在较大的形态特征差异, 以及物种间可能存在的自然杂交现象(Tao et al, 2021), 导致名录中的部分物种在分类上存在一定困难和争议, 如桦木科的盐桦与其近缘种形态差异非常小, 其种间界定具有不确定性.针对部分物种种间界定尚未澄清的问题, 建议采用形态和分子相结合的方法, 加强分类与系统发育的研究(Feng et al, 2016).同时利用更为精细的野生种群现状及其生境条件调查数据, 删去那些经过深入的野外调查发现了新的分布点或种群、种群数量超过预期的物种, 或已经脱离了灭绝风险的物种, 对极小种群野生植物名录定期更新和完善. ...
Habitat destruction and the extinction debt
1
1994
... 栖息地的退化和破碎化已使许多物种的种群规模减少到临界水平.生境破坏与未来灭绝之间关系——即灭绝债(extinction debt) (在栖息地被破坏后暂时存在但注定要走向灭绝的物种数量)——的一个重要启示是, 即使生境破碎化已经停止, 它对生物多样性的影响仍将继续(Tilman et al, 1994;Dullinger et al, 2012).以小而孤立的种群为代表的许多植物物种, 长时间存活(例如大乔木)掩盖了它们真正的濒危和保护状况.因此, 种群及群落结构、动态、影响因素的分析是保护生物学研究的重点(Volis, 2018b).物种生态位、种群结构、年龄及性别组成和更新状况研究揭示了其生态需求、濒危原因、种群的生存能力和发展态势, 有助于理解种群动态和制定保护策略(Sá et al, 2018).《中华人民共和国野生植物保护条例》规定“野生植物行政主管部门应当定期组织国家重点保护野生植物和地方重点保护野生植物资源调查, 建立资源档案”.但是, 目前在保护生物学中物种的空间分布、遗传变异、保护优先性评估等相关研究占主导地位, 却忽略了种群结构统计和物种生态位分析(Volis & Deng, 2020). ...
Minimum required number of specimen records to develop accurate species distribution models
1
2016
... 鉴于极小种群物种种群数量小的特点, 基于大样本方法发展的植物种群理论并不完全适用, 例如使用生态位模型或物种分布模型预测物种对全球变化的响应, 通常需要20个或更多的观测样本(van Proosdij et al, 2016).根据极小种群野生植物的分布、种群、群落、生境等调查、监测和实验数据, 发展具有针对性的假说和预测模型, 解释其小种群形成机制和恢复机制, 是亟待突破的瓶颈(Chen et al, 2020).其扩繁、回归、栖息地恢复等保护方法也可能与其他物种存在巨大差异, 很少有可以效仿的成功案例.因此, 相关保护方案必须考虑极小种群野生植物的特性, 研发基于小样本的保护实践体系(臧润国等, 2016). ...
How to conserve threatened Chinese plant species with extremely small populations?
2
2016
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 繁殖是植物最为关键的生活史阶段之一, 也是种群更新与维持的重要环节.极小种群野生植物自身生殖繁育力的衰退是导致其濒临灭绝的重要原因之一.开展繁殖生物学研究, 深入了解物种的个体成熟年龄、结果(种)时间动态、繁殖结构、授粉方式和主要传粉者、种子扩散、种子休眠情况和萌发条件等, 可为研发极小种群野生植物的人工扩繁技术奠定理论基础(Volis, 2016).对于存在繁殖困难的种群可以借助实验来控制其发芽和生长条件, 揭示种群更新失败的原因, 并最终通过减少竞争物种、引入保育植物、增加传粉昆虫等相应的方式来促进自然更新.由于大部分极小种群野生植物生长在偏远的地方, 开展长期的观测和实验研究较为困难, 目前对于极小种群野生植物繁殖生物学的相关研究还较为欠缺.Wade等(2016)通过收集120种极小种群野生植物的繁殖生物学信息, 发现仅有28个物种可以获得萌发信息, 10个物种可以获得储存特性信息. ...
Plant conservation in the Anthropocene: Definitely not win-win but maybe not lose-lose?
1
2018a
... 鉴于全球物种和自然栖息地的加速消失, 目前对破碎化栖息地的被动保护是不够的(Heywood, 2017).野外回归是野生植物种群重建的重要途径, 作为就地保护和迁地保护之间的桥梁以及迁地保护的最终目标,其保护效果超过了单一的物种保护措施(Ren et al, 2020).在人类世, 保护的未来在于栖息地的恢复和大规模的植物回归(Volis, 2018a).作为克服生境丧失、破碎化和繁殖隔离等问题的方法, 回归已成为完整的物种恢复计划的一个重要组成部分(Dalrymple et al, 2012). ...
Securing a future for China’s plant biodiversity through an integrated conservation approach
2
2018b
... 栖息地的退化和破碎化已使许多物种的种群规模减少到临界水平.生境破坏与未来灭绝之间关系——即灭绝债(extinction debt) (在栖息地被破坏后暂时存在但注定要走向灭绝的物种数量)——的一个重要启示是, 即使生境破碎化已经停止, 它对生物多样性的影响仍将继续(Tilman et al, 1994;Dullinger et al, 2012).以小而孤立的种群为代表的许多植物物种, 长时间存活(例如大乔木)掩盖了它们真正的濒危和保护状况.因此, 种群及群落结构、动态、影响因素的分析是保护生物学研究的重点(Volis, 2018b).物种生态位、种群结构、年龄及性别组成和更新状况研究揭示了其生态需求、濒危原因、种群的生存能力和发展态势, 有助于理解种群动态和制定保护策略(Sá et al, 2018).《中华人民共和国野生植物保护条例》规定“野生植物行政主管部门应当定期组织国家重点保护野生植物和地方重点保护野生植物资源调查, 建立资源档案”.但是, 目前在保护生物学中物种的空间分布、遗传变异、保护优先性评估等相关研究占主导地位, 却忽略了种群结构统计和物种生态位分析(Volis & Deng, 2020). ...
... 极小种群野生植物概念中引入了最小存活种群(MVP)这一指标, 即特定物种以一定概率存活一定时间所需的最少个体数(Reed & McCoy, 2014).MVP值可以根据物种的生物学特性以及设定的存活概率和时间, 通过种群生存力分析(population viability analysis, PVA)求得(Rosenfeld, 2014).然而我国野生植物资源监测体系尚不完善, 难以及时掌握极小种群野生植物资源的动态变化.国内对特定野生植物生存力分析的研究也较少, 当前对于绝大多濒危物种只有种群分布和种群大小的粗略数据(图2), 很少有种群数量结构信息, 种群可存活力的工作更少(Volis, 2018b).MVP对于极小种群野生植物的物种选列和保护实践具有重要意义, 因此应该基于对种群数量和繁殖物候的长期观测, 加强对极小种群野生植物种群结构的研究和预测, 根据不同物种的生物学特性和明确的保护目标(存活概率和存活时间)来确定种群可存活力. ...
Importance of a single population demographic census as a first step of threatened species conservation planning
2
2020
... 栖息地的退化和破碎化已使许多物种的种群规模减少到临界水平.生境破坏与未来灭绝之间关系——即灭绝债(extinction debt) (在栖息地被破坏后暂时存在但注定要走向灭绝的物种数量)——的一个重要启示是, 即使生境破碎化已经停止, 它对生物多样性的影响仍将继续(Tilman et al, 1994;Dullinger et al, 2012).以小而孤立的种群为代表的许多植物物种, 长时间存活(例如大乔木)掩盖了它们真正的濒危和保护状况.因此, 种群及群落结构、动态、影响因素的分析是保护生物学研究的重点(Volis, 2018b).物种生态位、种群结构、年龄及性别组成和更新状况研究揭示了其生态需求、濒危原因、种群的生存能力和发展态势, 有助于理解种群动态和制定保护策略(Sá et al, 2018).《中华人民共和国野生植物保护条例》规定“野生植物行政主管部门应当定期组织国家重点保护野生植物和地方重点保护野生植物资源调查, 建立资源档案”.但是, 目前在保护生物学中物种的空间分布、遗传变异、保护优先性评估等相关研究占主导地位, 却忽略了种群结构统计和物种生态位分析(Volis & Deng, 2020). ...
... 栖息地的退化和破碎化已经使得许多物种的种群规模减少到临界水平.理论预测和实证证据均表明, 小种群的灭绝风险高于大种群(Caughley, 1994).只有了解“小种群”的形成原因, 才能制定相应的解决方案, 保持种群生存能力、防止种群灭绝(Volis & Deng, 2019).极小种群野生植物一般是由自身和外部因素共同造成的, 自身因素包括遗传多样性低、近交衰退、繁殖障碍、种子萌发率低、适应性差等, 外部因素包括地质历史事件、冰期作用、自然灾害、病虫害、气候变化、人类利用、生境退化和破碎化等(Chen et al, 2014).极小种群野生植物濒危机制的揭示是种群得以保护和恢复的重要基础.每一种极小种群野生植物都有其独特的生理生态特性和致危机理, 需要采取不同的保育措施. ...
Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective
2
2016
... 繁殖是植物最为关键的生活史阶段之一, 也是种群更新与维持的重要环节.极小种群野生植物自身生殖繁育力的衰退是导致其濒临灭绝的重要原因之一.开展繁殖生物学研究, 深入了解物种的个体成熟年龄、结果(种)时间动态、繁殖结构、授粉方式和主要传粉者、种子扩散、种子休眠情况和萌发条件等, 可为研发极小种群野生植物的人工扩繁技术奠定理论基础(Volis, 2016).对于存在繁殖困难的种群可以借助实验来控制其发芽和生长条件, 揭示种群更新失败的原因, 并最终通过减少竞争物种、引入保育植物、增加传粉昆虫等相应的方式来促进自然更新.由于大部分极小种群野生植物生长在偏远的地方, 开展长期的观测和实验研究较为困难, 目前对于极小种群野生植物繁殖生物学的相关研究还较为欠缺.Wade等(2016)通过收集120种极小种群野生植物的繁殖生物学信息, 发现仅有28个物种可以获得萌发信息, 10个物种可以获得储存特性信息. ...
... 英文文献中, 有4篇文章第一单位为国外科研机构.其中, Wade等(2016)利用英国邱园种子信息数据库(http://data.kew.org/sid/)收集了中国120种极小种群野生植物的繁殖生物学信息, Solórzano等(2016)研究了墨西哥极小种群野生仙人掌白露 (Mammillaria albiflora)的种内遗传变异及优先保护种群, Cogoni等(2021)监测了意大利撒丁岛极小种群蕨类瓶尔小草(Ophioglossum vulgatum)的分布、种群大小、生境和就地保护效果.著名植物学家Peter Crane对中国极小种群野生植物保护计划的实施给予了高度评价, 认为该保护计划打破了理论和实践的壁垒, 能长远保护灭绝风险极高的植物(Crane, 2020). ...
Floral characteristics and pollination ecology of Manglietia ventii (Magnoliaceae), a plant species with extremely small populations (PSESP) endemic to South Yunnan of China
3
2017
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
The spatial distribution of threats to plant species with extremely small populations
0
2017
广西重点物种就地保护现状分析
1
2011
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
广西重点物种就地保护现状分析
1
2011
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
微生境对黄梅秤锤树野生种群叶片功能性状的影响
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
微生境对黄梅秤锤树野生种群叶片功能性状的影响
1
2020
... 研究不同生态因素如气候、土壤理化性质、微生境特征等对物种表型性状、幼苗更新等的影响, 有助于理解极小种群野生植物的生理生态适应性, 对预测极小种群野生植物的种群变化趋势和环境变化响应至关重要.尹雯丽(2020)①(①尹雯丽 (2020) 极小种群野生植物旱地油杉抗旱机制研究. 硕士学位论文, 云南大学, 昆明.)利用解剖结构、生理指标和转录组分析探究了旱地油杉(Keteleeria xerophila)的干旱调控机制, 发现其在干旱胁迫下具有较强的水分协调能力及保水能力和渗透调节能力, 并揭示了旱地油杉的抗旱基因.通过分析土壤理化性质、土壤微生物和土壤酶活性, 发现海南假韶子(Paranephelium hainanensis)不同群落对土壤因子的响应较为显著, 为探究其对异质性土壤的适应性奠定了基础(蔡开朗等, 2018).路兴慧等(2020)分析了生境特征对坡垒种群更新幼苗多度的影响, 揭示了其幼苗至幼树阶段种群的增补限制和关键限制因子.植物的功能性状变异和表型可塑性是其适应异质生境的主要机制, 对植物生长和分布有重要影响.陈俊等(2020)分析了水杉原生母树种群叶片功能性状对树木形态、地形因子及人为干扰的响应机制.王世彤等(2020)分析了黄梅秤锤树野生种群的叶片功能性状和生态化学计量特征对不同微生境的响应及其适应策略.还有一些研究基于MaxEnt模型分析了极小种群野生植物在不同情景模式下的潜在分布区(Cai et al, 2022; 陈舒豪和程广有, 2022).Qu等(2018)采用生态位模型研究了我国6个极小种群野生植物在当前和未来的栖息地分布, 并确定了这些物种的保护优先区.这些研究有助于应对气候变化对极小种群野生植物带来的威胁, 确保保护地对其分布范围的覆盖能力. ...
中国生物多样性就地保护成效与展望
1
2021
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
中国生物多样性就地保护成效与展望
1
2021
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
云南省自然保护区保护空缺分析
1
2018
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
云南省自然保护区保护空缺分析
1
2018
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
德保苏铁回归种群繁殖特征的初步研究
1
2018
... 回归已在越来越多的珍稀濒危植物保护实践中得到应用.云南省目前建立了“极小种群野生植物回归自然试验示范研究基地”, 成功开展了华盖木、毛果木莲、西畴青冈、漾濞槭等极小种群物种的野外回归工作(孙卫邦等, 2019).“典型极小种群野生植物保护与恢复技术研究”项目组建立了回归示范区6处, 回归了东北红豆杉、崖柏、坡垒、盐桦、瑶山苣苔(Dayaoshania cotinifolia)、海伦兜兰个体超过3万株.国家苏铁种质资源保护中心于2008年在广西黄连山自然保护区开展德保苏铁(Cycas debaoensis)回归试验, 回归5年后的调查表明, 德保苏铁回归种群生长良好, 17.6%的个体长出了大、小孢子叶球, 利用回归居群产生的种子培育苗木约5,300株(王运华等, 2018).截至2019年底, 中国有300个植物回归项目, 涉及206个物种, 其中中国特有物种112种.中国科学院华南植物园负责完成的“华南珍稀濒危植物的野外回归研究与应用”项目建立了“选取适当的珍稀植物进行基础研究和繁殖技术攻关, 再进行野外回归和市场化生产, 实现其有效保护, 同时通过区域生态规划及国家战略咨询, 推动整个国家珍稀濒危植物回归工作”的模式, 初步实现了珍稀濒危植物的产业化, 产生了良好的生态、社会和经济效益(Ren et al, 2012). ...
德保苏铁回归种群繁殖特征的初步研究
1
2018
... 回归已在越来越多的珍稀濒危植物保护实践中得到应用.云南省目前建立了“极小种群野生植物回归自然试验示范研究基地”, 成功开展了华盖木、毛果木莲、西畴青冈、漾濞槭等极小种群物种的野外回归工作(孙卫邦等, 2019).“典型极小种群野生植物保护与恢复技术研究”项目组建立了回归示范区6处, 回归了东北红豆杉、崖柏、坡垒、盐桦、瑶山苣苔(Dayaoshania cotinifolia)、海伦兜兰个体超过3万株.国家苏铁种质资源保护中心于2008年在广西黄连山自然保护区开展德保苏铁(Cycas debaoensis)回归试验, 回归5年后的调查表明, 德保苏铁回归种群生长良好, 17.6%的个体长出了大、小孢子叶球, 利用回归居群产生的种子培育苗木约5,300株(王运华等, 2018).截至2019年底, 中国有300个植物回归项目, 涉及206个物种, 其中中国特有物种112种.中国科学院华南植物园负责完成的“华南珍稀濒危植物的野外回归研究与应用”项目建立了“选取适当的珍稀植物进行基础研究和繁殖技术攻关, 再进行野外回归和市场化生产, 实现其有效保护, 同时通过区域生态规划及国家战略咨询, 推动整个国家珍稀濒危植物回归工作”的模式, 初步实现了珍稀濒危植物的产业化, 产生了良好的生态、社会和经济效益(Ren et al, 2012). ...
极小种群植物猪血木的种群现状及保护对策
1
2020
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
极小种群植物猪血木的种群现状及保护对策
1
2020
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
Meta-analysis of genetic representativeness of plant populations under ex situ conservation in contrast to wild source populations
1
2021
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
1
2021
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
1
2021
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
Botanic garden solutions to the plant extinction crisis
1
2020
... 作为最古老的保护策略之一, 迁地保护能够为珍稀濒危植物在其自然栖息地以外的特定区域提供生长和繁殖所需的条件.随着生境丧失、人类采伐和气候变化影响的加剧, 迁地保护的需求不断增长(Abeli et al, 2020).迁地保护与就地保护相辅相成, 通过在人为干预和精细管护下保护、研究、评价和利用植物多样性, 不仅是收集和保存珍稀濒危植物种质资源的重要方式, 也是植物回归引种及野生种群恢复重建的材料保障(Westwood et al, 2020). ...
Perspectives of genomics for genetic conservation of livestock
1
2010
... 物种的遗传多样性大小是决定其环境适应性的重要因素.小种群容易发生遗传漂变, 使遗传多样性迅速减少, 甚至导致种群间遗传分化迅速增加.遗传多样性的丧失会降低种群的延续性和物种的进化潜力, 高度的遗传分化可能导致远交衰退(Yang et al, 2022).探究极小种群野生植物的遗传组成、阐明其进化历史和种群动态、揭示其响应环境变化的遗传机制, 对理解其致濒机理并采取相应的保护措施极为重要, 也是保护生物学研究的重要内容.DNA测序技术的进步极大地促进了保护遗传学在生物多样性保护实践中的应用(Windig & Engelsma, 2010).尤其是全基因组测序成本的降低, 使得通过分析物种的种群历史动态和适应性演化来揭示极小种群野生植物的灭绝风险成为研究热点. ...
Limits to the adaptive potential of small populations
1
2006
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
Seed banking not an option for many threatened plants
1
2018
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
中国苏铁属植物资源和保护
4
2022
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
中国苏铁属植物资源和保护
4
2022
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
Conservation and fruit biology of Sichou oak (Quercus sichourensis, Fagaceae)—A critically endangered species in China
1
2016
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
东北红豆杉种群的回归成效及影响因素
1
2020
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
东北红豆杉种群的回归成效及影响因素
1
2020
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
国家重点保护植物“迁地”与“近地”保护有效性的比较研究
1
2012
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
国家重点保护植物“迁地”与“近地”保护有效性的比较研究
1
2012
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
极小种群野生植物的近地保护
2
2014
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
... ).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
极小种群野生植物的近地保护
2
2014
... 云南林业厅基于保护实践, 提出了极小种群野生植物的“近地保护” (near situ conservation)方法, 即在物种现有分布区/点附近选择气候、生境和群落相似的自然或半自然地段建立人工保护点(许再富和郭辉军, 2014).由于该方法也是将保护对象转移到其自然栖息地以外并进行人工管护, 本文将其归为迁地保护的一种形式.云南省对云南蓝果树、华盖木、西畴青冈、巧家五针松、滇桐(Craigia yunnanensis)等9种极小种群野生植物开展了近地保护试验, 发现近地保护种群的成活率和生长情况良好(孙卫邦等, 2019).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
... ).近地保护方法已被认为是我国极小种群野生植物主要拯救性保护措施之一(许再富和郭辉军, 2014).中国科学院西双版纳热带植物园近地保护了38种国家重点保护植物, 其中适应性良好的占总数的92%, 已开花结果的占84% (许再富等, 2012), 证明近地保护是一种有效而资源投入较小的方法, 这一结果也强调了植物园应注重当地区系成分植物的引种、栽培和保护. ...
我国近30年来植物迁地保护及其研究的综述
2
2008
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
... ).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
我国近30年来植物迁地保护及其研究的综述
2
2008
... 植物迁地保护是一个长期的过程.迁地保护种群是否能够在自然条件下完成从“种子到种子”的生活史全过程, 是迁地效果评价的重要标准之一(He, 2002).深圳仙湖植物园于2002年建立了“国家苏铁种质资源保护中心”, 目前已收集、保存全球苏铁类植物2科10属240余种, 迁地保护的部分物种已经可以完成生活周期(席辉辉等, 2022).将仅分布于我国新疆阿勒泰地区的盐桦(Betula halophila)迁地保护到山东烟台, 发现其在烟台地区适应性良好, 第3年即能开花结果(黄睿智等, 2020).基于11个植物园的植物迁地保护的生长适应与开花结果统计, 发现植物生长适应良好的占48%, 中等的占40%, 差的占12%; 已开花的种类占38%, 结果的种类仅占24% (许再富等, 2008).迁地保护的目标是建立一个能够应对和适应环境变化并且完成自我维持和更新的种群, 但迁地保护的个体数有限导致的遗传漂变、自交衰退等问题, 可能使迁地种群无法长期延续(Enßlin et al, 2011).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
... ).我国植物园目前保存的珍稀濒危植物, 大多数种子都来自个别自然种群和少数母株, 而且其中40%-60%的物种是在植物园间相互引种的, 此外其中45%的物种仅保存了1-5株个体(许再富等, 2008).通过比较迁地种群和野生种群的生活史特征和遗传多样性差异, 来评价迁地保护的有效性和合理性十分必要.比较黄梅秤锤树迁地保护种群与野生种群的果实性状, 发现迁地保护种群与野生种群果实重量差异不显著, 果实形态性状的种内变异程度不低于野生种群, 表明该物种的迁地保护基本成功(刘梦婷等, 2018).荟萃分析表明, 由于不完善的取样策略以及迁地保护过程中的遗传侵蚀, 大部分迁地保护种群不能有效涵盖其野生种群的遗传变异.当迁地保护样本数不少于30或50时, 迁地保护种群与野生种群的遗传多样性才没有表现出显著差异(Wei & Jiang, 2021).因此, 未来迁地实践中必须采取全面采样的策略, 对于已有的迁地种群进行周期性的遗传多样性检测和适应性评价, 必要时补充新的个体. ...
Genomic data reveals population genetic and demographic history of Magnolia fistulosa (Magnoliaceae), a Plant Species With Extremely Small Populations in Yunnan Province, China
2
2022
... 物种的遗传多样性大小是决定其环境适应性的重要因素.小种群容易发生遗传漂变, 使遗传多样性迅速减少, 甚至导致种群间遗传分化迅速增加.遗传多样性的丧失会降低种群的延续性和物种的进化潜力, 高度的遗传分化可能导致远交衰退(Yang et al, 2022).探究极小种群野生植物的遗传组成、阐明其进化历史和种群动态、揭示其响应环境变化的遗传机制, 对理解其致濒机理并采取相应的保护措施极为重要, 也是保护生物学研究的重要内容.DNA测序技术的进步极大地促进了保护遗传学在生物多样性保护实践中的应用(Windig & Engelsma, 2010).尤其是全基因组测序成本的降低, 使得通过分析物种的种群历史动态和适应性演化来揭示极小种群野生植物的灭绝风险成为研究热点. ...
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
China’s conservation program on Plant Species with Extremely Small Populations (PSESP): Progress and perspectives
2
2020
... 2017年, 国家林业局评估了120种极小种群野生植物的保护成效, 发现尽管大多数物种都实施了全面的调查, 但包括海南石豆兰(Bulbophyllum hainanense)、海南鹤顶兰(Phaius hainanensis)和海南大苞兰(Sunipia hainanensis)在内的有些类群却没有找到; 有37个物种分布在自然保护区, 60个物种通过委托护林员或当地管理部门得到保护, 为26个物种建立了就地保护小区; 对80个物种进行了迁地保护试验示范, 56个物种开展了人工繁殖, 26个物种开展了种群增强与回归试验, 对一半物种进行了种质资源保存(孙卫邦和徐永福, 2022).国家林业和草原局于2012-2018年组织开展的第二次全国重点保护野生植物资源调查发现, 与第一次调查有可比性的54种极小种群野生植物中, 有36种的野外种群数量稳中有升, 占67% (http://www.forestry.gov.cn/main/586/20220210/091653623218205.html).可见, 尽管取得了一些可喜的成果, 但拯救规划的目标仅部分实现, 资金不足、缺乏保护生物学的科学指导和专业实践人员等原因影响了拯救规划工程的实施效果(Yang et al, 2020).极小种群野生植物保育实践的论文主要涉及就地保护、迁地保护与种质资源保存、野外回归、人工繁育、标准化体系等5个方面(图3). ...
... 生物多样性的消失是一个不可逆转的过程, 极小种群物种的丧失将影响其生态功能的发挥, 甚至破坏整个自然生态系统.极小种群野生植物拯救保护工程的实施以及相关的研究与实践, 帮助我们确保这些物种的零灭绝, 使我们有机会能够欣赏、研究、了解和利用它们.自2017年以来, 生物多样性保护已逐渐在中国社会主流化.2021年在中国昆明召开了联合国《生物多样性公约》第十五次缔约方大会第一阶段会议, 中国启动了国家公园和国家植物园的建设, 这些举措揭开了中国植物多样性保护的新篇章.中国具有独特的环境价值观和相关政策, 以西方模式为基础、由国际非政府组织支持的保护项目并不完全适合中国, 实施“中国特色的环境保护”更有利于保护我们国家的物种、生态系统、景观和文化多样性(Grumbine & Xu, 2011).在政府主导下, 通过完善法律法规和体制机制, 加强保育研究和实践体系建设, 推动公众参与, 深化国际合作, 为实施保护创造更有力的社会支持, 极小种群野生植物也许能够拥有一个光明的未来(Yang et al, 2020). ...
云南蓝果树保护小区——中国首个极小种群野生植物保护小区建设实践
1
2016
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
云南蓝果树保护小区——中国首个极小种群野生植物保护小区建设实践
1
2016
... 由于极小种群野生植物种群生境面积小、破碎化程度高, 在很多情况下被受到严重破坏的环境所包围, 或者位于人口稠密、人类活动频繁的地区, 建立大面积的自然保护区是不现实的, 一个有效的解决办法是建立保护小区或微保护区.作为由县级及以下人民政府设立的面积较小的保护区(点), 自然保护小区面积一般不超过1,000 ha, 但对于保护国家或地方重点保护的分布范围较为狭窄的野生动植物群落及栖息地具有重要意义(崔国发, 2013).深圳市梅林水库建立了仙湖苏铁(C. fairylakea)保护小区(Feng et al, 2021).云南西双版纳分布的142种兰科植物中有44种位于青石寨自然保护小区内, 有42种位于银厂自然保护小区内(Liu et al, 2015).基于极小种群野生植物云南蓝果树的保护小区建设实践, 杨文忠等(2016)探讨了保护小区规划的原则、内容、程序以及管理机构设置和运行机制, 为制定极小种群野生植物保护小区建设方案提供了指导和参考. ...
2
2018
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
2
2018
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
极小种群野生植物的概念及其对我国野生植物保护的影响
5
2015
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... ).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
极小种群野生植物的概念及其对我国野生植物保护的影响
5
2015
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... ).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
Genomic effects of population collapse in a critically endangered ironwood tree Ostrya rehderiana
0
2018
中国纳入一级保护的极小种群野生植物濒危机制
2
2021
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
中国纳入一级保护的极小种群野生植物濒危机制
2
2021
... 自实施极小种群野生植物保护拯救工程以来, 很多省份(北京、新疆、黑龙江、广西、海南、四川、安徽、云南)、地区(西藏林芝市、广东河源市、云南文山州)和保护区(江西官山国家级自然保护区、海南霸王岭国家级自然保护区、浙江天目山国家级自然保护区、广东连山县大旭山市级自然保护区)等基于资源调查数据统计了当地的极小种群野生植物资源及保护现状.部分研究针对特定极小种群野生植物开展了其生存状况、种群动态、群落结构和生境特征调查, 共涉及到包括梓叶槭(Acer catalpifolium)、崖柏(Thuja sutchuenensis)、黄梅秤锤树(Sinojackia huangmeiensis)、水杉、东北红豆杉、坡垒(Hopea hainanensis)等在内的61个物种, 这些种群和群落调查数据为探索其濒危机制, 开展就地和迁地保护、生境修复等保护方案提供了理论依据.姚志等(2021)分析了中国28种极小种群野生植物的种群特征, 发现13个物种遗传多样性低, 11个呈现衰退型种群结构, 11个呈现狭域、聚集型分布.幼苗阶段是影响种群定居和更新的关键时期, 能够决定未来的物种组成和群落结构.猪血木(Euryodendron excelsum)野生种群的年龄结构和动态变化表明, 该物种幼树数量丰富, 但由于向小树发展的过程受阻, 种群无法及时更新补充(魏雪莹等, 2020).云南梧桐(Firmiana major)在云南境内的所有分布点都位于自然保护地之外, 包括利用其树皮制造绳索和放牧等人为干扰导致了该物种数量的急剧下降(Li et al, 2020).尽管目前中国有8个与东北红豆杉有关的自然保护区, 为了追求经济利益, 伐木、砍枝、采种等人类活动依然导致了其种群大小和栖息地面积显著减少(Long et al, 2021). ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
峨眉拟单性木兰的开花生物学特性与繁育系统
1
2019
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
峨眉拟单性木兰的开花生物学特性与繁育系统
1
2019
... 仅检索到9篇英文文献开展了极小种群野生植物的繁殖生物学研究, 涉及到11个物种.Li等(2018)观测了大树杜鹃(Rhododendron protistum var. giganteum)的开花时间、繁殖结构、颜色、气味、可能传粉者等, 发现该物种野外种群的传粉限制并不严重.传粉实验结果表明, 毛果木莲(Manglietia ventii)对传粉者有依赖性, 但由于种群数量少、花粉落置量(pollen deposition)不足导致自然种群结实率低, 保护该物种需要通过增加可育个体密度来打破传粉限制(Wang et al, 2017).Lin等(2022)研究了华盖木的种子休眠、萌发需求和储存条件, 发现其种子耐干燥, 能在传统条件和超低温贮藏条件下保存几个月.而西畴青冈(Cyclobalanopsis sichourensis)则是果实对干燥环境的敏感性限制了其更新(Xia et al., 2016).旱地木槿(Hibiscus aridicola)、白魔芋(Amorphophallus albus)、细花百部(Stemona parviflora)和百部(S. japonica)的传粉媒介在不同地点或时间存在缺失, 影响了其种群的成功繁殖概率以及长期存在(Tang et al, 2020).检索到15篇开展了繁殖生物学研究的中文文献, 研究内容包括云南蓝果树(Nyssa yunnanensis)的花药发育过程和败育原因(康洪梅等, 2019)、峨眉拟单性木兰(Parakmeria omeiensis)的开花生物学特性与繁育系统(余道平等, 2019)、崖柏的生殖物候及胚胎发育(金江群等, 2019)等.这些研究能够使保护工作者更加了解极小种群野生植物生存繁衍所需要的环境条件, 为物种繁殖技术的发展提供了关键信息, 也为在生境破碎化、气候变化、传粉媒介改变等背景下重建种群提供有效的指导意见(孙卫邦等, 2019). ...
5
2016
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
... 很多珍稀濒危植物具有繁殖瓶颈, 繁殖过程表现出高度的脆弱性和敏感性(Baskin & Baskin, 2014).成功繁殖对于维持种群数量, 尤其是濒临灭绝物种的种群数量至关重要, 因此繁殖瓶颈的突破是极小种群野生植物解濒研究的重中之重, 发展规模化扩繁技术是其保育体系的重要环节(臧润国等, 2016).对极小种群野生植物进行高效的人工繁殖, 能够扩大种群数量并将扩繁的苗木应用于迁地保护、野外回归甚至满足商品市场的需求, 有利于其种质资源的保护和可持续利用(邓莎等, 2020). ...
... 鉴于极小种群物种种群数量小的特点, 基于大样本方法发展的植物种群理论并不完全适用, 例如使用生态位模型或物种分布模型预测物种对全球变化的响应, 通常需要20个或更多的观测样本(van Proosdij et al, 2016).根据极小种群野生植物的分布、种群、群落、生境等调查、监测和实验数据, 发展具有针对性的假说和预测模型, 解释其小种群形成机制和恢复机制, 是亟待突破的瓶颈(Chen et al, 2020).其扩繁、回归、栖息地恢复等保护方法也可能与其他物种存在巨大差异, 很少有可以效仿的成功案例.因此, 相关保护方案必须考虑极小种群野生植物的特性, 研发基于小样本的保护实践体系(臧润国等, 2016). ...
5
2016
... 极小种群野生植物概念的提出及其拯救保护工程的实施对我国野生植物保护理念的转变影响深远.由于我国需要保护的野生植物种类众多, 此前保护管理部门只能针对珍稀濒危植物进行宏观的保护管理和规划, 以法律法规、行政手段和宣传教育等为主要策略, 而极小种群野生植物更加强调“基于种群管理的物种保护”理念, 运用植物种群生态学原理和方法针对明确的目标物种开展相应的保护实践(杨文忠等, 2015).此外, 科学研究与管理实施之间的脱节一直被认为是影响中国生物多样性保护的系统障碍之一(Grumbine & Xu, 2011).极小种群野生植物保护过程中特别强调科学研究要与拯救保护实践相接轨, 能够促进植物地理学、种群生态学、生殖生物学和保护生物学等相关学科的基础理论和应用技术相结合, 在实现学科发展的同时提升我国野生植物保护管理水平(杨文忠等, 2015; 臧润国等, 2016).这一概念的提出标志着中国的植物保护事业进入了一个政策与保护更为匹配的全新时代(Ma et al, 2013). ...
... 极小种群野生植物由于种群数量小、面临胁迫大及繁殖困难等固有特性, 决定了其保护生物学研究具有如下特点: (1)极小种群物种种群数量极少, 濒临高度灭绝风险, 极易因偶然事件的发生而永久消失, 因此对其研究具有紧迫性(孙卫邦和韩春艳, 2015).(2)由于野外种群数量极小, 实施保护时强调在传统保护措施的基础上引入种群管理的理念和方法, 因此研究中需要运用种群生态学的原理和方法, 基于种群调查的基础数据对种群数量、规模、结构和动态等进行调节与管理(杨文忠等, 2015).(3)植物种群理论大多是基于大样本方法发展起来的, 而极小种群物种种群数量极少, 很多遗传学、生态学研究缺乏统计意义, 样本量太少导致模拟时出现模型过度拟合(Breiner et al, 2015), 因此研发保育方案需要重点考虑基于小样本的方法和理论体系(臧润国等, 2016).(4)繁殖能力低下是大多数极小种群物种的共同特征, 因此其繁殖生物学特性以及限制繁殖的内在和外在因素是研究的重要方面(邓莎等, 2020). ...
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
... 很多珍稀濒危植物具有繁殖瓶颈, 繁殖过程表现出高度的脆弱性和敏感性(Baskin & Baskin, 2014).成功繁殖对于维持种群数量, 尤其是濒临灭绝物种的种群数量至关重要, 因此繁殖瓶颈的突破是极小种群野生植物解濒研究的重中之重, 发展规模化扩繁技术是其保育体系的重要环节(臧润国等, 2016).对极小种群野生植物进行高效的人工繁殖, 能够扩大种群数量并将扩繁的苗木应用于迁地保护、野外回归甚至满足商品市场的需求, 有利于其种质资源的保护和可持续利用(邓莎等, 2020). ...
... 鉴于极小种群物种种群数量小的特点, 基于大样本方法发展的植物种群理论并不完全适用, 例如使用生态位模型或物种分布模型预测物种对全球变化的响应, 通常需要20个或更多的观测样本(van Proosdij et al, 2016).根据极小种群野生植物的分布、种群、群落、生境等调查、监测和实验数据, 发展具有针对性的假说和预测模型, 解释其小种群形成机制和恢复机制, 是亟待突破的瓶颈(Chen et al, 2020).其扩繁、回归、栖息地恢复等保护方法也可能与其他物种存在巨大差异, 很少有可以效仿的成功案例.因此, 相关保护方案必须考虑极小种群野生植物的特性, 研发基于小样本的保护实践体系(臧润国等, 2016). ...
典型极小种群野生植物保护与恢复技术研究
0
2016
典型极小种群野生植物保护与恢复技术研究
0
2016
2
2019
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
... )、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
2
2019
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
... )、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
5
2020
... 为全面掌握我国野生植物资源状况, 国家林业局于1997-2003年组织开展了第一次全国重点保护野生植物资源调查, 共调查了189种国家重点保护野生植物的资源数量、分布和受威胁状况等.调查结果显示, 百山祖冷杉(Abies beshanzuensis)、银杉(Cathaya argyrophylla)、华盖木(Pachylarnax sinica)、落叶木莲(Manglietia decidua)等55种野生植物的野外种群低于稳定存活界限的最小存活种群(minimum viable population, MVP, 指特定物种以一定概率存活一定时间所需的最少个体数量) (Reed & McCoy, 2014), 随时面临着野外灭绝的危险.其中, 稳定存活界限是指保证种群在特定时间内能稳定健康生存所需的最小阈值, 低于这个阈值种群会逐渐趋向灭绝.基于此, 国家林业局提出了极小种群野生植物(Wild Plants with Extremely Small Populations, WPESP; 也有学者翻译为Plant Species with Extremely Small Populations, PSESP)的概念: 具体是指分布地域狭窄, 长期受到外界因素胁迫干扰, 呈现出种群退化和个体数量持续减少, 种群和个体数量都极少, 已经低于稳定存活界限的最小可存活种群, 而随时濒临灭绝的野生植物(国家林业局, 2010)①(①国家林业局 (2010) 全国极小种群野生植物拯救保护工程规划(2010-2015). 非正式出版材料.).极小种群植物保育强调的对象是野生植物种群, 对于栽培种群或人工繁育数量大的物种, 如水杉(Metasequoia glyptostroboides)、东北红豆杉(Taxus cuspidata)等更要注意区分野生种群和人工种群, 否则容易在保育实践中引起困惑(臧润国, 2020).因此, 本文中极小种群野生植物统一使用WPESP的译法.《全国极小种群野生植物拯救保护工程规划(2011-2015年)》中选择首批保护极小种群野生植物的原则主要包括3个方面: 一是野外数量极小、极度濒危、随时有灭绝危险的野生植物; 二是生境要求独特、生态幅狭窄的野生植物; 三是潜在基因价值不清楚、其灭绝将引起基因流失、生物多样性降低、社会经济价值损失巨大的种群数量相对较小的野生植物.依据这些原则, 最终确定的首批120种重点保护极小种群野生植物包含36种国家I级保护植物, 26种国家II级保护植物, 59种省级重点保护植物. ...
... 对于极小种群野生植物来说, 通过迁地保护收集和保存种质资源, 为就地保护和野外回归提供了至关重要的备份.云南省已对61种极小种群野生植物开展了迁地保护; 昆明植物园极小种群野生植物专类园迁地保存了44种极小种群植物物种, 部分物种已开花结实; 中国西南野生生物种质资源库保存了极小种群野生植物种子20种94份、DNA材料28种156份(孙卫邦等, 2019).华东师范大学收集了包括水杉、东北红豆杉、天目铁木等9种极小种群物种在内的种质资源1,500份, 并在湖北利川建立了种质资源圃.十三五重点研发计划“典型极小种群野生植物保护与恢复技术研究”在不同气候区建立了8处迁地保护示范基地, 迁地保存了14种极小种群物种的27,215株个体(臧润国和黄继红, 2020). ...
... 随着经济发展、资源开发规模的日益扩大, 珍稀濒危植物及其生境遭受的干扰和破坏难以遏止.尽管目前已经成功挽救了部分物种(孙卫邦等, 2019), 但目前有关极小种群野生植物濒危原因和解濒技术的研究还较为缺乏, 不能满足全国极小种群野生植物拯救保护工程有效实施的科技需求(臧润国等, 2020).基于国内外最先进的保护理论和方法, 在开展大量技术实践示范基础上, 建设极小种群野生植物保护和恢复的标准化技术体系, 建立相应的评估管理规范, 能够填补在各保育环节存在的技术空白, 有助于开展并推广积极有效的保护和恢复措施. ...
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
... )、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
5
2020
... 为全面掌握我国野生植物资源状况, 国家林业局于1997-2003年组织开展了第一次全国重点保护野生植物资源调查, 共调查了189种国家重点保护野生植物的资源数量、分布和受威胁状况等.调查结果显示, 百山祖冷杉(Abies beshanzuensis)、银杉(Cathaya argyrophylla)、华盖木(Pachylarnax sinica)、落叶木莲(Manglietia decidua)等55种野生植物的野外种群低于稳定存活界限的最小存活种群(minimum viable population, MVP, 指特定物种以一定概率存活一定时间所需的最少个体数量) (Reed & McCoy, 2014), 随时面临着野外灭绝的危险.其中, 稳定存活界限是指保证种群在特定时间内能稳定健康生存所需的最小阈值, 低于这个阈值种群会逐渐趋向灭绝.基于此, 国家林业局提出了极小种群野生植物(Wild Plants with Extremely Small Populations, WPESP; 也有学者翻译为Plant Species with Extremely Small Populations, PSESP)的概念: 具体是指分布地域狭窄, 长期受到外界因素胁迫干扰, 呈现出种群退化和个体数量持续减少, 种群和个体数量都极少, 已经低于稳定存活界限的最小可存活种群, 而随时濒临灭绝的野生植物(国家林业局, 2010)①(①国家林业局 (2010) 全国极小种群野生植物拯救保护工程规划(2010-2015). 非正式出版材料.).极小种群植物保育强调的对象是野生植物种群, 对于栽培种群或人工繁育数量大的物种, 如水杉(Metasequoia glyptostroboides)、东北红豆杉(Taxus cuspidata)等更要注意区分野生种群和人工种群, 否则容易在保育实践中引起困惑(臧润国, 2020).因此, 本文中极小种群野生植物统一使用WPESP的译法.《全国极小种群野生植物拯救保护工程规划(2011-2015年)》中选择首批保护极小种群野生植物的原则主要包括3个方面: 一是野外数量极小、极度濒危、随时有灭绝危险的野生植物; 二是生境要求独特、生态幅狭窄的野生植物; 三是潜在基因价值不清楚、其灭绝将引起基因流失、生物多样性降低、社会经济价值损失巨大的种群数量相对较小的野生植物.依据这些原则, 最终确定的首批120种重点保护极小种群野生植物包含36种国家I级保护植物, 26种国家II级保护植物, 59种省级重点保护植物. ...
... 对于极小种群野生植物来说, 通过迁地保护收集和保存种质资源, 为就地保护和野外回归提供了至关重要的备份.云南省已对61种极小种群野生植物开展了迁地保护; 昆明植物园极小种群野生植物专类园迁地保存了44种极小种群植物物种, 部分物种已开花结实; 中国西南野生生物种质资源库保存了极小种群野生植物种子20种94份、DNA材料28种156份(孙卫邦等, 2019).华东师范大学收集了包括水杉、东北红豆杉、天目铁木等9种极小种群物种在内的种质资源1,500份, 并在湖北利川建立了种质资源圃.十三五重点研发计划“典型极小种群野生植物保护与恢复技术研究”在不同气候区建立了8处迁地保护示范基地, 迁地保存了14种极小种群物种的27,215株个体(臧润国和黄继红, 2020). ...
... 随着经济发展、资源开发规模的日益扩大, 珍稀濒危植物及其生境遭受的干扰和破坏难以遏止.尽管目前已经成功挽救了部分物种(孙卫邦等, 2019), 但目前有关极小种群野生植物濒危原因和解濒技术的研究还较为缺乏, 不能满足全国极小种群野生植物拯救保护工程有效实施的科技需求(臧润国等, 2020).基于国内外最先进的保护理论和方法, 在开展大量技术实践示范基础上, 建设极小种群野生植物保护和恢复的标准化技术体系, 建立相应的评估管理规范, 能够填补在各保育环节存在的技术空白, 有助于开展并推广积极有效的保护和恢复措施. ...
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
... )、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
1
2019
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
1
2019
... 因此, 除了以上4个方面, 我们还梳理了涉及极小种群野生植物的相关标准, 这些标准基本归属林业部门主管.目前已建立如下8个相关的行业标准: 《极小种群野生植物保护与扩繁技术规范(LY/T 2652-2016)》(臧润国等, 2015)、《极小种群野生植物保护原则与方法(LY/T 2938-2018)》(杨文忠等, 2018)、《极小种群野生植物保护技术标准综合体第1部分 就地保护及生境修复技术规程(LY/T 3086.1-2019)》(臧润国等, 2019)、《极小种群野生植物保护技术标准综合体(第2部分) 迁地保护技术规程(LY/T 3086.2-2019)》(臧润国等, 2019)、《极小种群野生植物野外回归技术规范(LY/T 3185-2020)》(李俊清等, 2020)、《极小种群野生植物种质资源保存技术规程(LY/T 3187-2020)》(臧润国等, 2020)、《极小种群野生植物苗木繁育技术规程(LY/T 3186-2020)》(臧润国等, 2020)、《极小种群野生植物水松保护与回归技术规程(LY/T 3259-2021)》(文亚峰等, 2021).这些标准针对极小种群野生植物形成了一套全链条式的技术集成和示范体系, 能够为极小种群野生植物拯救保护工程提供重要的技术支撑, 同时能够带动相关生态建设和产业的规范化发展, 产生良好的生态和社会效益. ...
1
2020
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
1
2020
... Plant Diversity和《生物多样性》对于推动极小种群野生植物研究和实践起到了非常积极的作用.2015年6月《生物多样性》发表了3篇文章, 其中杨文忠等(2015)梳理了极小种群野生植物概念的提出背景、过程、理解途径及其对我国野生植物保护领域的重要意义, 孙卫邦和韩春艳(2015)对极小种群野生植物科学保护的多个研究方向进行了展望, 马永鹏和孙卫邦(2015)指出了极小种群野生植物抢救性保护面临的机遇与挑战.2016年10月Plant Diversity出版了极小种群野生植物保护研究专辑, 发表了种子及孢子的保存和萌发、遗传多样性与遗传结构、传粉生物学和种子传播、繁殖生物学、保护遗传学、野外回归种群适应性相关的7篇极小种群野生植物研究文章, 探讨了部分物种的濒危原因并提出了相应的保护建议(Sun, 2016).2020年3月《生物多样性》出版了“极小种群野生植物保育专辑”, 基于14个典型物种的案例研究分析了极小种群野生植物的种群及群落动态、生境适应策略、遗传多样性及核心种质资源、繁殖特性和人工繁殖方法等内容, 并呼吁开展更多针对极小种群野生植物种群数量小、面临胁迫大及繁殖困难等固有特点的保护理论和技术研究(臧润国, 2020). ...
Climate change- induced water stress suppresses the regeneration of the critically endangered forest tree Nyssa yunnanensis
1
2017
... 当前, 不断加剧的人为干扰和气候变暖增强了上述自然过程的发生, 对种群恢复产生负面影响, 进一步导致物种灭绝速度加快(Miraldo et al, 2016).中国高等植物濒危状况评估结果表明, 高等植物最主要的致危因子是生境退化和丧失, 基础建设、经济作物种植以及生态旅游等均对野生植物的生存环境产生了巨大的压力; 人为过度采挖位列植物致危因子的第二位(覃海宁和赵莉娜, 2017).20世纪70年代以来, 随着我国园林景观行业的发展、园艺博览会等的宣传以及野生植物线上交易的兴起, 原生的苏铁植物和兰科植物等被大量采挖贩卖.Wang等(2017)绘制了极小种群野生植物的威胁因子分布图, 发现它们主要受到过度利用、生境破碎化、种群太小的威胁.对海南岛极小种群野生植物分布格局的研究也发现首要威胁因素是人为干扰(Chen et al, 2014).气候变暖通过改变极小种群野生植物的物候节律, 缩减了其气候适宜分布区, 从而增加了其灭绝风险(Chen et al, 2020).野外和室内试验结果表明, 云南蓝果树对干旱胁迫的抗逆性和适应性差, 气候变化引起的水分亏缺加重严重影响了其自然更新, 栖息地恢复和小气候改善等有效的保护措施有助于该物种打破更新限制(Zhang et al, 2017). ...
Autotoxicity as a cause for natural regeneration failure in Nyssa yunnanensis and its implications for conservation
1
2015
... 目前从机理上揭示珍稀濒危植物濒危机制或致濒因子的研究还较少.利用遗传多样性手段如保护基因组学分析濒危机制是最有效和最直接的方法, 能够为种群规模的进化历史和物种未来的恢复潜力提供重要见解(Garner et al, 2016).研究发现, 天目铁木(Ostrya rehderiana)种群数量锐减是历史气候变化和人为干扰共同作用的结果.末次冰盛期天目铁木种群数量下降到阈值, 进入灭绝漩涡, 极小的种群规模、低水平的遗传多样性削弱了其适应能力, 全新世期间人类砍伐和耕种又进一步直接减小了其种群规模(Yang et al, 2018).种群结实率和种子萌发率低、幼苗适应性和抗逆性差难以发育成幼树导致了海南风吹楠的濒危(蒋迎红, 2018).自毒现象(autotoxicity)是植物化感作用的一种重要形式, 通过释放次生代谢物对自身或种内其他植物产生危害, 是植物适应种内竞争的结果, 在植物自然更新、种群结构调节、环境胁迫抵御等关键生态过程中均发挥着重要作用(彭少麟和邵华, 2001).云南蓝果树通过根、茎、叶、蒴果产生的自毒作用显著抑制了其种子萌发和幼苗生长, 应通过清理凋落物、添加活性炭以及迁地保护等方式改善种群自然更新(Zhang et al, 2015).大树杜鹃叶片也具有较强的自毒作用, 可能是导致其种群数量稀少和自然更新障碍的因素之一(刘芳黎等, 2017). ...
光强和土壤含水量对云南蓝果树幼苗生长及光合特征的影响
1
2018
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
光强和土壤含水量对云南蓝果树幼苗生长及光合特征的影响
1
2018
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
裂叶沙参分布区域和生物学生态学习性的调查
1
1998a
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
裂叶沙参分布区域和生物学生态学习性的调查
1
1998a
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
裂叶沙参分布区域和生物学生态学习性的调查
1
1998b
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
裂叶沙参分布区域和生物学生态学习性的调查
1
1998b
... 尽管极小种群野生植物这一概念提出较晚, 但20世纪90年代前后部分学者开展的研究已经隐含了这一概念.例如, 陆素娟和邓莉兰(1994)发现云南昭通市巧家县特有的五针白皮松(Pinus squamata)仅存20株个体, 更新不良, 并通过分析其地质历史和生态需求探讨了它的潜在分布区.张文辉和祖元刚(1998a)调查濒危植物裂叶沙参(Adenophora lobophylla)时发现该物种的天然分布区局限在四川西北部金川县的5个乡, 分布面积不超过5,000 km2, 且该物种年龄结构老化, 种群呈衰退趋势, 并分析了该物种的生境条件和致危因素(张文辉和祖元刚, 1998b).尽管这些研究并没有正式提出极小种群野生植物的概念, 但强调了这些物种狭窄的分布区、特殊的生物生态学特性、极小的种群数量和不断加剧的威胁因子, 与极小种群概念和保护对象的选择原则不谋而合. ...
Genetic diversity and structure of Rhododendron meddianum, a plant species with extremely small populations
1
2021
... 检索到的中文文献中, 有22篇文章基于微卫星标记、叶绿体基因组等方法, 对包括白花兜兰(P. emersonii)、海南风吹楠(Horsfieldia hainanensis)、喙核桃(Annamocarya sinensis)等在内的21个物种进行了遗传多样性和遗传结构分析.另外, 贺水莲等(2016)综述了云南省极小种群野生植物基于分子标记的遗传多样性研究现状, 同时展望了遗传多样性分析在极小种群野生植物保护中的应用前景.苏金源等(2020)综述了极小种群裸子植物的遗传多样性研究案例, 探讨了濒危裸子植物的种群维持机制、致濒因素和保护方案, 表明通过遗传多样性研究揭示极小种群野生植物致濒机理对于高效保护的重要性.77篇英文文献中, 有43篇开展了种群遗传变异研究, 涉及到38个物种, 其中50%以上运用了全基因组技术.对漾濞槭10个种群105个个体的全基因组重测序结果表明, 该物种遗传多样性较低, 种群间基因流动频繁, 重复的瓶颈事件、人类活动导致的生境破碎化以及自然生境中成熟个体的性别偏倚现象共同造成了漾濞槭的濒危状态(Ma et al, 2022).Zhang等(2021)通过采集红萼杜鹃(Rhododendron meddianum) 3个野生种群的45个个体, 发现该物种具有较高的遗传多样性, 其最有效的保护方式是就地保护, 由于不同种群间具有较高的遗传分化, 种质资源采集时各个种群需分别采样以避免远交衰退.Yang等(2022)利用基因组学的手段分析了显脉木兰(Magnolia fistulosa)的种群遗传多样性和种群大小波动, 发现该物种具有较高的遗传多样性和种群分化水平, 末次冰盛期后其有效种群规模恢复到较高的水平, 人为干扰是造成该物种目前种群规模较小和分布受限的主要因素.以上研究表明, 极小种群野生植物通常具有高度近交、遗传多样性低和遗传漂变频率高的特点, 亟需通过实施遗传拯救(genetic rescue)增加遗传多样性, 提高适合度, 增强适应能力, 以维持物种的长期存活(孙卫邦等, 2021). ...
不同郁闭度对野外回归的梓叶槭幼树形态和生理特征的影响
1
2020
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
不同郁闭度对野外回归的梓叶槭幼树形态和生理特征的影响
1
2020
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
中国极小种群野生植物的保护现状评估
2
2018
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
中国极小种群野生植物的保护现状评估
2
2018
... 极小种群野生植物概念的提出对促进我国生物多样性保护和生态文明建设具有重要意义.首先, 极小种群野生植物大多数为我国特有植物, 具有重要的生态、经济和文化价值(Ren et al, 2012).由于分布狭窄、种群数量小、面临胁迫大, 极小种群野生植物是最易丧失的生物资源之一, 如果保护不够及时, 其独特基因、生物价值和生态功能将随着物种的灭绝而消失, 给自然生态系统和人类社会带来难以估量的损失.因此, 对其开展抢救性保护有助于避免物种灭绝, 维护生态平衡, 对于我国乃至世界的生物多样性保护都具有极为重要的意义(张则瑾等, 2018).第二, 自然界的大多数物种都是稀有的, 具有较小的种群数量和狭窄的分布范围.Enquist等(2019)基于2亿条标本采集、野外调查等数据记录的详细评估发现, 约36.5%的陆生植物是“极其罕见的”.这一结果表明, 地球上很大比例的植物面临着日益加剧的灭绝风险, 了解这些稀有物种的种群形成过程和保育方法对于保护生物学以及应对全球变化的适应潜力至关重要, 而极小种群野生植物的研究和保护对于这些稀有物种具有重要的引领作用.第三, 很多极小种群野生植物起源较早, 是现存种子植物中较原始的类群, 对研究种子植物的起源与演化具有重要价值, 蕴藏的遗传信息可以为研究古植物区系、古地理和古气候变迁提供珍贵的素材, 研究其适应环境的机制也有助于解释物种的长期生存及对环境变迁的响应(Crane, 2020; 席辉辉等, 2022).第四, 极小种群野生植物的概念易于理解和接受, 能够唤起公众对生物多样性的保护意识, 随着各种宣教活动的开展, 目前社会各界对极小种群野生植物拯救保护重要性的认识大为提高, 并积极参与到保护行动中来, 有助于减缓过度采集这些珍稀濒危物种带来的严重威胁.第五, 与现有其他同类概念相比, 极小种群野生植物的概念与物种选择标准具有独特性.我国曾多次发布和更新珍稀濒危保护植物和国家重点保护野生植物名录, 这些名录难以从字面理解其确切含义, 选列和评价物种的依据也多为定性标准.而极小种群野生植物物种选列的指标中提出了关于个体数和分布点的明确量化评定指标, 减少了专家的主观判断, 增强了物种选列的客观性(杨文忠等, 2015).对比IUCN物种红色名录, 尽管其濒危等级和标准历经了多次修订, 评估标准也越来越细化和量化, 但IUCN红色名录假定种群表现出正常的包含所有生活史周期的种群结构, 并不适用于极小种群物种(Volis, 2016).大部分极小种群物种由于繁殖或更新困难, 表现出种群性别结构或年龄结构的偏倚(宋垚彬等, 2020).极小种群野生植物保护强调物种种群统计速率和种群动态的长期观察, 可以作为物种濒危程度的可靠评估指标. ...
... 就地保护对于维持生物的繁衍、适应与进化, 维系生态系统服务和功能等均具有重要的作用(王伟和李俊生, 2021), 是生物多样性保护中最有效的措施之一.由于极小种群野生植物对生境的特殊要求, 维护其自然生境是保护的重要方式之一.国家公园、自然保护区、自然公园和生态保护红线各有侧重、相互弥补, 形成了中国的自然保护地体系(高吉喜等, 2021), 将我国最重要、最具代表性的生态系统纳入了最严格的保护体系,对于保护极小种群野生植物、维护国家和区域生态安全、建设生态文明发挥着重要作用.中国有苏铁属植物约20种, 均被列为国家一级重点保护野生植物, 其中50%被列入了极小种群野生植物名录.四川攀枝花市建立了以攀枝花苏铁(C. panzhihuaensis)为主要保护对象的国家级自然保护区, 云南普渡河的攀枝花苏铁分布区被纳入云南轿子雪山国家级自然保护区(席辉辉等, 2022).野生东北红豆杉及其赖以生存的针阔混交林被纳入黑龙江穆棱东北红豆杉国家级自然保护区进行保护.张则瑾等(2018)整理了120种极小种群野生植物的高精度分布图, 发现国家级自然保护区对极小种群野生植物分布区的平均覆盖率为21.5%, 省级自然保护区的平均覆盖率为10.9%, 有14%的物种未受到任何国家级或省级自然保护区覆盖.云南省有21种国家重点保护野生植物及极小种群野生植物分布在59个国家级、省级自然保护区之外(王勇等, 2018).广西已建立了各类自然保护区78个, 覆盖了82%的重点保护野生植物, 但部分物种的大多数分布点还位于保护区之外, 已被保护的部分种群所在生境破碎化严重或生境质量较差, 无法阻止种群数量的持续下降(王双玲等, 2011). ...
Seed germination of Huagaimu, a critically endangered plant endemic to southeastern Yunnan, China
1
2009
... 很多物种在末次冰盛期分布范围急剧缩小, 以小而孤立的种群分布在避难所(Willi et al, 2006).由于种群规模小导致了很多问题, 如遗传变异的丧失、有害等位基因的积累、近交衰退、对随机事件的敏感性、Allee效应导致的繁殖失败等, 都增加了物种的灭绝风险(Keller & Waller, 2002).例如, 苏铁属植物雌雄异株, 由于居群较小, 成熟的开花植株较少, 花粉传播距离有限, 形成了传粉限制, 导致结实率较低, 自我更新困难(席辉辉等, 2022).繁殖能力低下是大多数极小种群物种的共同特征, 也是导致它们濒危的一个重要因素(臧润国等, 2016).例如种子休眠难以打破(如华盖木, Zheng & Sun, 2009)、种子活力和种子萌发差(如水杉, Liu et al, 2020)、花粉和种子传播效率低(如伯乐树Bretschneidera sinensis, Qiao et al, 2012)、种子产量低(如毛果木莲, Wang et al, 2017)等.中国28种纳入一级保护的极小种群野生植物的濒危原因和受威胁因素研究表明, 导致它们濒危的内在原因主要是繁殖力低(21种)和竞争能力弱(16种), 受威胁因素主要包括过度采挖等人类活动导致的种群数量减少(15种)和生境破坏(25种)以及气候变化等(姚志等, 2021). ...
极小种群植物博罗红豆的光合生理特征日变化研究
1
2021
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
极小种群植物博罗红豆的光合生理特征日变化研究
1
2021
... 植物生理生态学研究生态因子与植物生理特性之间的关系, 能够从生理机制上探讨植物对不同环境条件的响应和适应能力.基于极小种群野生植物的调查、观测与实验数据, 研究不同生态因子对各个生活史阶段的作用, 分析极小种群野生植物对生境变化的响应和适应能力, 有助于揭示其致危的生理生态机制并采取有针对性的解濒措施(孙卫邦等, 2021).目前, 极小种群野生植物生理生态学的相关研究还比较薄弱.大部分研究集中在分析极小种群野生植物光合生理特性, 如研究光照强度、土壤含水量和水分胁迫等对物种光合特性的影响(张珊珊等, 2018; 钟象景等, 2021), 为选择合适的迁地和回归地点及制定科学有效的管护措施提供依据. ...
珍稀濒危植物的回归: 理论和实践
2
2011
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
... 回归的成功标准分为短期和长期两类, 前者包括个体的成活、种群的建立和扩散; 后者包括回归种群的自我维持和在生态系统中发挥稳定功能等, 也是回归成功的最终标准(周翔和高江云, 2011), 但这些标准对评价长寿命物种的回归成功与否可操作性不强.目前关于回归的科学监测数据非常缺乏.Liu等(2015)评估了中国154个物种的222个野外回归案例, 发现仅有不到50%的案例有个体是否存活的记录, 仅有30%的案例有关于个体是否开花或结果的记录.对建立的回归种群开展持续监测, 并与野生种群进行种群动态、繁殖能力、群落多样性等的比较, 是评价回归地点选择适宜性、物种回归实践是否成功的可靠途径.利用高通量测序技术比较野生和回归华盖木种群根际和土壤微生物, 发现二者的微生物群落组成相似, 但类群的相对多度差异显著(Shen et al, 2020; Su et al, 2021). ...
珍稀濒危植物的回归: 理论和实践
2
2011
... 在自然生态系统中, 植物与其他生物之间的关系极为复杂, 一个新物种很难在一个稳定的群落中建立, 即使这个物种曾经是该群落的组成成分(Ren et al, 2014), 珍稀濒危植物又往往生存能力和适应性较差, 因此珍稀濒危植物的回归是一项高风险和高花费的工程(周翔和高江云, 2011).回归生境的适宜性对物种生长具有重要影响, 研究植物的生理生态特征对不同野外回归生境的适应性可以为评估极小种群物种回归生境的适宜性提供科学依据.不同光照条件对梓叶槭幼树形态和生理特征的影响研究表明, 林缘适宜的光照条件更适合梓叶槭野外回归(张宇阳等, 2020).人工管护可以帮助植物克服定居限制而实现成功回归.选择不同苗龄的东北红豆杉幼苗在不同生境开展野外回归试验, 发现长时间低温胁迫影响了1-2年生幼苗的生长状况, 而4-5年生幼苗因受动物啃食而回归效果不良, 表明在回归试验点采用幼苗保护装置的必要性(徐超等, 2020). ...
... 回归的成功标准分为短期和长期两类, 前者包括个体的成活、种群的建立和扩散; 后者包括回归种群的自我维持和在生态系统中发挥稳定功能等, 也是回归成功的最终标准(周翔和高江云, 2011), 但这些标准对评价长寿命物种的回归成功与否可操作性不强.目前关于回归的科学监测数据非常缺乏.Liu等(2015)评估了中国154个物种的222个野外回归案例, 发现仅有不到50%的案例有个体是否存活的记录, 仅有30%的案例有关于个体是否开花或结果的记录.对建立的回归种群开展持续监测, 并与野生种群进行种群动态、繁殖能力、群落多样性等的比较, 是评价回归地点选择适宜性、物种回归实践是否成功的可靠途径.利用高通量测序技术比较野生和回归华盖木种群根际和土壤微生物, 发现二者的微生物群落组成相似, 但类群的相对多度差异显著(Shen et al, 2020; Su et al, 2021). ...
极小种群野生植物丹霞梧桐繁育技术
1
2021
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...
极小种群野生植物丹霞梧桐繁育技术
1
2021
... 采用种子繁殖育苗是保持物种遗传多样性的有效方法, 扦插、嫁接和组织培养等无性繁殖技术则可用于对有性繁殖能力低下的种类进行快速扩繁.国家重点研发计划项目“典型极小种群野生植物保护与恢复技术研究”对14种典型极小种群野生植物的繁殖特性和人工繁殖方法进行了研究, 并利用播种技术繁殖了其中12个物种的230,000株种苗, 利用扦插、嫁接和组织培养等无性繁殖技术扩繁了其中10个物种的60,365株苗木(邓莎等, 2020).打破种子的休眠, 去除其机械或化学阻碍, 探索种子储存和萌发所需的光照、温度、湿度等条件, 是开展人工繁育的必要手段.百山祖冷杉种子野外萌发困难, 实验证明20℃/25℃的温度条件、14 d的低温分层处理、接种外生菌根等种子辅助处理手段可以提高其种子萌发率(Hu et al, 2022).丹霞梧桐(Firmiana danxiaensis)属于硬实种子, 播种前种子适当破皮处理可有效提高其萌发率(周小芬等, 2021).还有些研究应用不同基质、不同种类和不同浓度的生长调节剂确定了物种扦插生根的最佳处理组合(刘俊等, 2019; 陈加利等, 2021).至少36%的极度濒危植物物种、27%的濒危物种和35%的易危物种的种子是顽拗型(recalcitrant)种子(Wyse et al, 2018), 如广西青梅(Vatica guangxiensis)、狭叶坡垒(H. chinensis)等极小种群物种(孙卫邦等, 2019), 这些种子无法在种子库标准的干燥/低温环境中存活; 此外, 虽然一些正常型(orthodox)种子耐低温和干燥, 但其种子活力很难超过10年(Li & Pritchard, 2009), 这些物种采种后应及时播种育苗或者优先采用超低温技术进行保存. ...




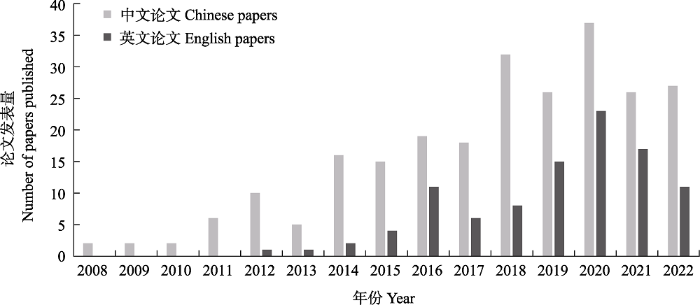
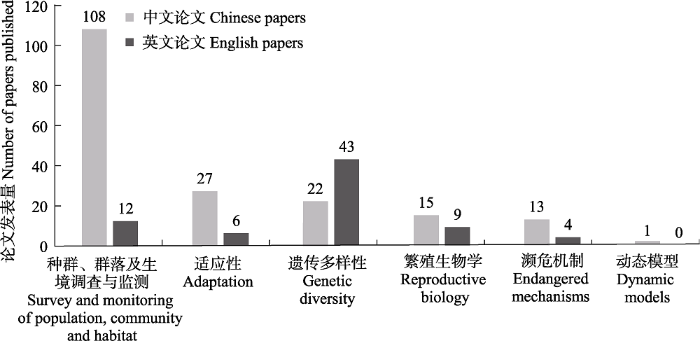
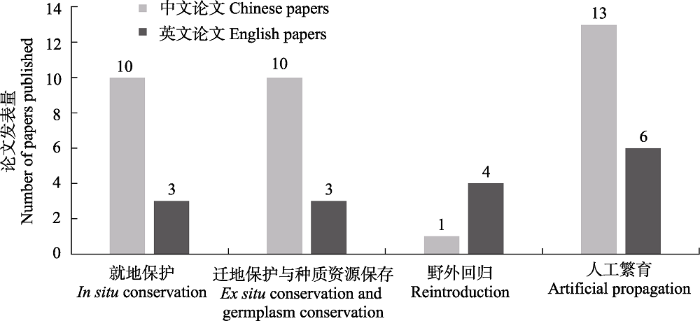


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}