



中国亚热带地区是中国经济最发达、人口最密集的地区, 拥有世界上面积最大的常绿阔叶林, 具有较高的生物多样性和生态服务功能(宋永昌, 2013; Fang et al, 2014; Xiang et al, 2016)。数千年来, 受人类经济活动与全球气候变化的深刻影响, 我国亚热带常绿阔叶林在纬度上不断发生变迁(宋永昌, 2013), 尤其是在20世纪中叶到21世纪初发生了大幅度向北推移(缪启龙等, 2009)。据政府间气候变化专门委员会(Intergovernmental Panel on Climate Change, IPCC)第五次评估报告, 未来全球气候将持续变暖, 全球平均温度将在21世纪末上升0.3-4.8℃ (Allen et al, 2007), 这将在不同程度上影响中国气候的变化和极端天气的产生(黄晓莹等, 2008; 赵宗慈等, 2008), 从而影响我国亚热带常绿阔叶林分布区及地上生物量的变化(Dai et al, 2016)。由此可见, 亚热带常绿阔叶林亟需保护(宋永昌, 2013), 建立自然保护区便是对其进行保护的有效手段, 但目前我国亚热带常绿阔叶林被保护的比例仅12.3% (陈雅涵等, 2009), 且自然保护区覆盖率偏低并呈片断化分布, 存在一定的保护空缺, 保护区布局有待优化(马克平, 2001; Tang et al, 2006; Lu et al, 2018; Cai et al, 2021)。因此, 在全球气候变暖等背景下, 有必要对现有的自然保护区进行优化, 尤其是提高亚热带常绿阔叶林的保护比例。
在亚热带常绿阔叶林群落中, 优势类群个体数量多、盖度大、生物量高、体积较大、生存能力强, 对群落结构的形成和群落环境的稳定有重要作用(王春晶, 2017)。然而, 大幅度的气候变化与极端气候可能会对优势类群产生消极影响(Parmesan & Yohe, 2003; Gebrekirstos et al, 2008), 再加上过度开发利用优势树种是世界许多地区普遍存在的现象(Muñoz et al, 2015, 这都将会导致天然林的稳定性降低, 直接威胁森林植物多样性(Canadell & Raupach,2008; Prieto-Torres et al, 2016)。因此, 在全球气候变化情景下, 为了更好地保护亚热带常绿阔叶林, 维持其生态系统稳定, 需要着重保护群落优势种(王春晶, 2017)。樟属(Cinnamomum)隶属于樟科, 为常绿乔木或灌木, 我国约有49种, 广泛分布于我国南方, 北达陕西及甘肃南部, 是集经济、工业和生态等价值于一身的重要类群(李锡文, 1982; 李捷和李锡文, 2004; Li et al, 2008)。作为被子植物的基部类群之一, 樟属在早始新世伴随着常绿阔叶林起源于劳亚古陆, 在新生代最温暖的时候扩张到北半球的高纬度地区, 在晚始新世后由于气候变冷向南退缩, 并因白令陆桥的消失而形成跨太平洋的间断分布, 是我国现代常绿阔叶林的重要组成部分(宋永昌, 2013; Huang et al, 2016)。樟属植物作为优势类群在我国典型常绿阔叶林乔木层中占有重要地位, 其属的区系重要值为56.52, 在常绿阔叶林所有物种属中位居第四(王希华, 2006); 且广泛分布于我国亚热带常绿阔叶林中, 如樟(C. camphora)、香桂(C. subavenium)在北亚热带及中亚热带是优势种, 云南樟(C. glanduliferum)、阴香(C. burmannii)则在中亚热带至南亚热带是优势种或者常见种, 而天竺桂(C. japonicum)常见于东部亚热带地区(倪建和宋永昌, 1997; Tang, 2015)。大部分樟属物种喜暖喜湿, 但也有部分物种适应了低温寒冷的气候, 例如樟、猴樟(C. bodinieri)和银木(C. septentrionale)等能耐-10℃以下的绝对最低温(张旻桓等, 2011; 程许娜, 2012 ( 程许娜 (2012) 猴樟对低温胁迫的生理响应及其耐寒性分析. 硕士学位论文, 河南农业大学, 郑州.); 王朴等, 2012), 这为樟属植物的分布区扩张与应对极端气候提供了可能。
以多样性格局数据为基础的物种分布区是该物种在一个区域内能够分布的最大范围, 并与其生存息息相关(Jenkins et al, 2013; Di Marco & Santini, 2015)。物种丰富度指一个区域内的总物种数, 常作为确定自然保护区和评估保护优先等级的标准(如Han et al, 2019; D'Amen & Azzurro, 2020), 而物种丰富度热点区域代表物种多样性最丰富的区域, 是在有限的人力、物力和财力下实现生物多样性最优保护的一种途径(Myers,1990; Brummitt & Lughadha, 2003)。因此, 对物种分布区和物种丰富度热点区域的研究有助于了解物种和类群迁移规律, 进而为生物类群保育策略的合理制定提供科学依据(李利平等, 2019)。物种分布模型(species distribution modelling, SDM)利用物种的分布数据, 并结合其生境下的环境变量推测物种的基础生态位(fundamental niche), 再结合不同历史时期的环境数据模拟不同时期物种的潜在分布区(Araújo & Peterson,2012)。在众多SDM中, 最大熵(maximum entropy, MaxEnt)模型具有建模快、使用广、精度高等优点(Merow et al, 2013), 即使在样本量较少的情况下也具有较高的精度和稳定性(Pearson et al, 2007), 相比于其他SDM模型能够更好地预测物种的潜在分布(Phillips et al, 2006)。
目前关于气候变化对中国亚热带常绿阔叶林优势类群影响的研究还较少, 大部分的研究涉及单个物种, 而对植物物种多样性和丰富度方面的研究较为匮乏, 且不同植物对气候变化有着不同的响应趋势(吕佳佳和吴建国, 2009; 郑维艳和曹坤芳, 2020)。因此, 需要综合研究多个物种的变化趋势来探讨气候变化对亚热带常绿阔叶林的影响。鉴于樟属植物作为优势类群在亚热带常绿阔叶林森林群落中占据着不可忽视的重要地位, 本研究将结合全球气候变化情景下的气候数据和樟属的地理分布数据, 利用MaxEnt模型模拟中国樟属植物在间冰期、冰期、当前及未来的潜在分布区, 探讨其在不同气候背景下的分布区和物种丰富度热点区域的变迁历史, 评估我国现有自然保护区对樟属植物的保护状况。期望本研究能有助于了解气候变化对亚热带常绿阔叶林优势类群的影响, 为我国亚热带常绿阔叶林自然保护地的规划和布局提供科学依据。
1 材料与方法
1.1 物种分布数据的收集
在全球生物多样性信息网络(GBIF,
1.2 环境数据
从古气候网站(
气候数据均采用2.5′的分辨率, 用ArcGIS 10.2软件按照研究区域进行裁剪, 并将裁剪后的气候数据转换为ASCII格式。中国地图来源于自然资源部标准地图服务(
为了避免环境因子之间的高相关性而导致模拟结果的过度拟合, 影响MaxEnt模型评估环境变量的贡献率(Luo et al, 2017), 利用R软件的cor函数对不同历史时期的19个气候变量进行Pearson相关性分析, 保留r < |0.7|中贡献率较小的气候因子(详见附录2)。
1.3 MaxEnt模型参数设置与可信度检验
将樟属的物种分布数据(.CSV格式)和5个时期的气候数据(.ASCII格式)导入MaxEnt3.4.1软件中进行物种生态位模拟。樟属物种可用坐标点的范围为15-317个不等, 均符合MaxEnt模型对最少数据的要求(Pearson et al, 2007)。对于坐标点少于25个的物种, 采用Jackknife方法进行模拟评估, 具体是在建立一个物种的模型时, 将其中一个物种的坐标移除, 并基于剩下的n-1个坐标建立模型, 由此每个物种可以建立n个模型, 选择最优模型来进行MaxEnt生态位模拟(Pearson et al, 2007); 对于可用坐标点大于25个的物种, 设置物种分布数据的75%作为训练集, 25%作为测试集。运算迭代次数设置为20次, 其余均采用默认值。以受试者工作特征曲线(receiver operator characteristic, ROC)下的面积(areas under curves, AUC)作为评估模拟结果的可靠性。AUC值的范围为[0, 1], 越接近1表明模拟的可靠性越高(Jiang et al, 2016): 当AUC值为[0.9, 1]时模拟结果非常准确; [0.8, 0.9)时模拟结果准确; [0.7, 0.8)时模拟结果一般; 小于0.7时结果不可信(Elith et al, 2006)。基于MaxEnt 3.4.1软件自动生成的气候变量贡献率、SPSS中的主成分分析(principal component analysis, PCA)及Pearson相关性分析, 综合分析限制樟属植物地理分布的主导气候变量。
1.4 物种丰富度热点区域预测及保护评估
将MaxEnt模型模拟结果导入ArcGIS 10.2软件中转化为栅格图层进行可视化处理, 采用10%训练存在阈值作为物种适生区临界值, 即高于该值为适生区, 低于该值为非适生区(Radosavljevic & Anderson, 2014)。通过图层叠加和栅格计算得到每一栅格内物种存在的数量, 即物种丰富度。将物种丰富度大于1的区域作为樟属分布区。根据前人的研究, 将物种丰富度前5%、10%、25%的区域作为物种丰富度热点区域(Xu et al, 2017; Yu et al, 2017b), 由于本研究的樟属物种有限(47种), 最终选择物种丰富度前25%的地区作为樟属的物种丰富度热点区域。根据中国森林覆盖率情况, 将某一市级行政区域内物种丰富度热点区域面积占该区域总面积23%以上的行政区域列为保护评估对象(王兮之, 2019)。利用中国国家级、省级、市县级自然保护区底图数据(Zhang et al, 2015)与评估对象相叠加, 并结合中华人民共和国生态环境部发布的《2017年全国自然保护区名录》(
2 结果
MaxEnt模型模拟结果显示, 中国樟属47种植物模拟曲线的AUC值均大于0.9, 表明MaxEnt模型预测结果可靠(附录1)。
2.1 当前樟属潜在分布区空间格局
模型模拟结果表明, 当前樟属物种潜在分布区域总面积为251.61万km2, 北达陕西南部和甘肃南部, 向南分布于南方各省; 樟属物种丰富度热点区域总面积为16.55万km2, 主要分布在广东、广西、贵州南部和四川东南部, 云南南部、海南南部也有少量分布(图1D)。大部分樟属物种的适宜生境在长江以南区域, 仅有少数物种的适宜生境可达长江以北的陕西、甘肃南部等地, 例如油樟(C. longepaniculatum)、阔叶樟(C. platyphyllum)、川桂(C. wilsonii)和银木; 部分物种的适宜分布区非常狭窄, 例如滇南桂(C. austroyunnanense)、爪哇肉桂(C. javanicum)、细毛樟(C. tenuipile)和毛叶樟仅分布在云南南部, 兰屿肉桂(C. kotoense)、土肉桂(C. osmophloeum)、菲律宾樟树(C. philippinense)和网脉桂(C.reticulatum)仅分布在台湾。
图1
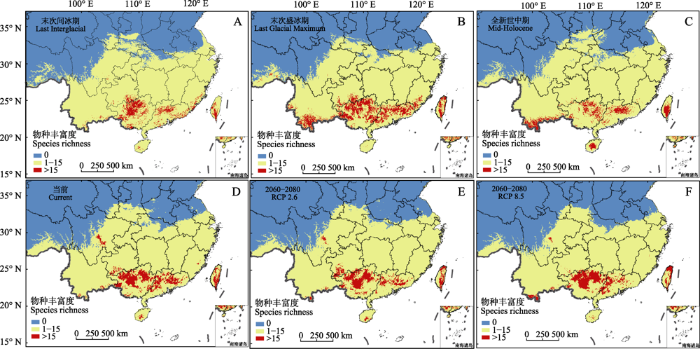
图1
中国樟属植物物种丰富度分布图。A-F依次为末次间冰期(LIG)、末次盛冰期(LGM)、全新世中期(MH)、当前(Current)以及未来2060-2080年RCP2.6和RCP8.5情景下樟属植物物种丰富度。
Fig. 1
Distribution of species richness of Cinnamomum in China. A-F shows the species richness of Cinnamomum under the scenarios of the Last Interglacial (LIG), Last Glacial Maximum (LGM), Mid-Holocene (MH), current, RCP2.6 and RCP8.5 in future 2060-2080, respectively.
模型模拟结果还表明, 大部分樟属物种的分布同时受到多个环境因子影响, 其中最干季降水量、最冷季降水量、年平均降水量和温度季节变化对大多数樟属物种的分布有显著影响。少数物种分布的主导环境因子(累积贡献率大于90%)仅为单气候因子, 例如毛叶樟、细毛樟主要受等温性影响, 而网脉桂主要受年温度变化范围的影响(附录1)。樟属物种丰富度热点区域内环境因子的前3个主成分方差解释率为88.966% (PCA1: 48.277%, PCA2: 31.961%, PCA3: 8.758%), 足够代表所有成分的特征(Cattell, 1966)。在方差解释最多的PCA1中, 载荷值的绝对值大于0.8的气候因子分别是年平均降水量、最干月降水量、最干季降水量和最冷季降水量, 这表明它们对樟属物种丰富度具有决定性作用。对樟属物种丰富度与这4个环境因子的相关性分析显示它们与樟属的物种丰富度均呈正相关关系(附录3)。
2.2 过去和未来樟属潜在分布空间格局
LIG、LGM和MH时期, 樟属植物潜在分布区面积分别为243.04万km2、249.46万km2和250.96万km2, 其中LGM比LIG增加2.6%, 扩增的区域主要在大别山、巫山、大巴山和喜马拉雅山脉东段, 但在关中平原、湖北中部和江苏沿海等低海拔地区有小部分收缩; MH比LGM增加0.6%, 扩增的区域主要在秦岭山脉周围及江苏、山东沿海; 相比于当前的分布范围, 3个历史时期樟属植物的潜在分布区面积分别减少3.4%、0.85%和0.26%。在未来RCP2.6和RCP8.5情景下, 樟属植物潜在分布区面积分别为253.33万km2和245.54万km2, 其中RCP2.6情景比当前潜在分布区面积增加0.7%, 扩增的区域主要在湖北东北部; RCP8.5情景比当前和RCP2.6情景分别减少2.4%和3.1%, 均沿着亚热带北部边缘轻度收缩。樟属植物潜在分布面积按大小排序为: 2080 (RCP2.6) > 当前 > MH > LGM > 2080 (RCP8.5) > LIG, 虽总体上看略有扩张的趋势, 但变化幅度不大( 图1)。
樟属物种丰富度热点区域在5个时期均主要分布在华南和西南地区, LIG、LGM和MH面积分别为11.56万km2、22.68万km2和12.05万km2, LGM的面积最大、分布范围也最广, 比LIG、MH和当前分别增加96%、88%和37%; 未来RCP2.6和RCP8.5情景下分别为15.17万km2和14.91万km2, 分别比当前减少8.4%和10%。可见, 在未来全球气候变暖增湿的两种情景条件下, 樟属物种丰富度热点区域均呈收缩趋势, 并且随着温室气体的持续排放, 热点区域逐渐缩减至华南地区。物种丰富度热点区域面积按大小排序为: LGM > 当前 > 2080 (RCP2.6) > 2080 (RCP8.5) > MH > LIG, 总体略有扩张的趋势, 且在5个时期均有较大变化( 图1)。
2.3 自然保护区保护评价
图2
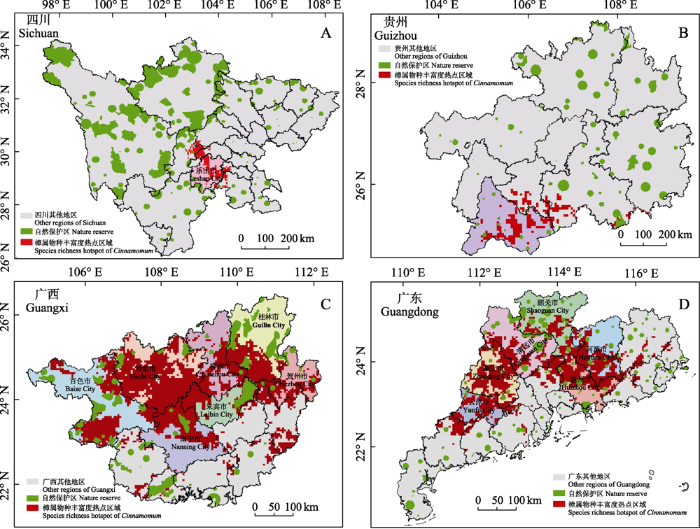
(1)四川东南部的樟属物种丰富度热点区域面积占该省市级行政区域面积23%以上的主要为乐山市, 樟属物种丰富度为22种。该市虽然建设有黑竹沟和马边大风顶2个国家级自然保护区, 但均未能对樟属物种丰富度热点区域进行保护。
(2)贵州南部的樟属物种丰富度热点区域面积占该省市级行政区域面积23%以上的主要为黔西南布依族苗族自治州, 樟属物种丰富度为19种。该地区设立了35个面积小、分散零落的市县级自然保护区, 仅有仙鹤坪和龙头大山市级自然保护区、赖子山县级自然保护区保护了该区域仅3.11%的樟属物种丰富度热点区域。
(3)广西的樟属物种丰富度热点区域面积占该省市级行政区域面积23%以上的主要有百色市、河池市、柳州市、桂林市、南宁市、来宾市和贺州市, 樟属物种丰富度分别为24种、23种、20种、18种、20种、18种和18种, 其中百色市建立了岑王老山国家级自然保护区、大王岭省级自然保护区、老虎跳省级自然保护区、百东河市级自然保护区等19个自然保护区; 桂林市建有花坪国家级自然保护区、架桥岭省级自然保护区、寿城省级自然保护区等12个自然保护区; 南宁市建有大明山国家级自然保护区、龙虎山省级自然保护区等6个自然保护区, 来宾市建有大瑶山国家级自然保护区、金秀老山省级自然保护区; 贺州市建有七冲国家级自然保护区、姑婆山省级自然保护区、滑水冲省级自然保护区、西岭山省级自然保护区、大桂山省级自然保护区, 各区域内的热点区域都得到了较好保护。河池市的樟属物种丰富度热点区域面积最大, 占该市面积的71.13%, 但该市的木论国家级自然保护区、龙滩省级自然保护区、九万山国家级自然保护区仅对该市7%的樟属物种丰富度热点区域进行了保护。柳州市建有九万山国家级自然保护区、元宝山国家级自然保护区、拉沟鸟类县级自然保护区, 但仅有拉沟鸟类县级自然保护区对该市8%的樟属物种丰富度热点区域进行了保护。
(4)广东的樟属物种丰富度热点区域面积占该省市级行政区域面积23%以上的主要有云浮市、肇庆市、清远市、韶关市、河源市和惠州市, 樟属物种丰富度分别为20种、22种、19种、18种、20种、21种。肇庆市建有鼎湖山国家级自然保护区、黑石顶省级自然保护区、深坑市级自然保护区、罗壳山市级自然保护区等24个自然保护区; 清远市建有笔架山省级自然保护区、石门台国家级自然保护区等26个自然保护区; 韶关市建有南岭国家级自然保护区、车八岭国家级自然保护区、乳源大峡谷省级自然保护区等36个自然保护区; 河源市建有新港省级自然保护区、康禾省级自然保护区等50个自然保护区; 惠州市建有象头山国家级自然保护区、南昆山省级自然保护区、罗浮山省级自然保护区、惠东白马山县级自然保护区等29个自然保护区, 各自对樟属物种丰富度热点区域进行了较好保护。云浮市虽然建有大云雾山市级自然保护区、罗定龙湾市级自然保护区、郁南望君山县级自然保护区等11个自然保护区, 但仅对该市6.6%的樟属物种丰富度热点区域进行了保护。
3 讨论
3.1 樟属植物潜在空间分布格局
本研究结果表明, 樟属植物广泛分布于长江以南区域, 向北可达陕西、甘肃南部等地, 其物种丰富度热点区域主要分布在广东、广西、云南南部、四川东南部和海南等我国大陆北热带及亚热带地区的中南部(图1D), 这与前人的研究结果基本吻合(李锡文, 1982; 查凤书等, 2008; 张规富和何帅艳, 2014; 郑维艳等, 2018)。中国亚热带常绿阔叶林分布在23°-33° N、98°-123° E之间, 大致在长江以南各省(宋永昌,2013), 与樟属植物的分布区范围基本一致。我国亚热带地区主要受到东亚季风的影响, 其中西部地区主要受印度洋西南季风的影响, 而东部地区主要受太平洋季风的影响, 季风带来的大量降水造就了我国亚热带丰富的生物多样性(Qiu et al, 2011; 陈灵芝等, 2015)。MaxEnt模型的拟合结果和主成分分析结果表明, 年平均降水量、最干季降水量、最冷季降水量等水分因子影响了大多数樟属植物的分布(附录1), 同时也在塑造樟属物种丰富度空间分布格局上有重要的作用, 说明水分因子是影响樟属植物分布的主要气候因子, 且随着水分的增多, 樟属植物分布区扩大、物种丰富度增加(附录3)。
根据化石孢粉学证据, 在末次盛冰期时, 我国常绿阔叶林向南退缩至华南地区, 冰期后又向北扩张至现在的纬度(Ni et al, 2010)。也有研究表明, 在未来气候条件下, 亚热带常绿阔叶林植物将会北扩东进, 推测我国亚热带常绿阔叶林将向北方移动(Ni et al, 2000; 吴显坤等, 2016; 胡文强等, 2017; Lim et al, 2018)。但本研究结果表明, 樟属植物的潜在分布区在各个时期的变化并不明显, 整体变化范围仅0.6%-3.4%, 只在亚热带常绿阔叶林边缘的各大山脉和平原出现局部的扩张与收缩。我们推测有以下几个原因: 第一, 亚热带常绿阔叶林植物的谱系地理研究表明, 在第四纪盛冰期部分植物有着多个隐形避难所(24°-33° N), 即多个避难所模式(Shiet al, 2014; Ma et al, 2015; Zhu et al, 2020), 它们主要通过在局部地区高低海拔间进行迁移来度过冰期与间冰期的交替(Qiu et al, 2011; Dao et al, 2013); 同时也有部分植物呈现出南方大型避难所(~24° N)的模式(Denk & Grimm,2009; Gong et al, 2016; Ye et al, 2019)。通过模型模拟我们发现, 樟属植物在面对第四纪间冰期与冰期的交替时同时存在两种避难所模式。分布在北亚热带地区较低海拔的樟属物种向附近具有复杂地形和微环境的各大山脉迁移, 如银木由关中平原向秦岭和大巴山山脉迁移、野黄桂(C. jensenianum)由四川盆地向巫山山脉迁移、猴樟由长江中下游平原向巫山和雪峰山山脉迁移; 而肉桂(C. cassia)、平托桂(C. tsoi)退到24° N以南的大型避难所中。第二, 樟属类群是热带起源、亚热带辐射分布(Huanget al, 2016), 进化出对气候有不同喜好的物种: 如喜温喜暖的毛桂(C. appelianum)、钝叶桂(C. bejolghota)、猴樟和红辣槁树(C. kwangtungense)的潜在分布区在冰期气候变冷时收缩, 冰期后至未来气候变暖又北扩东进; 喜冷好寒的银叶桂(C. mairei)、屏边桂(C. pingbienense)、狭叶桂(C. heyneanum)和米槁(C. migao)的潜在分布区则在冰期扩大, 冰期后收缩; 而横跨南北广泛分布的物种则具有较宽的生态位, 对环境的适应能力较强, 如樟、黄樟(C. parthenoxylon)、少花桂(C. pauciflorum)和香桂的潜在分布区在冰期与间冰期交替、冷暖气候变化情景下并无太大变化。同时, 樟属植物面对气候变化而表现出来的多避难所模式共存, 以及不同物种之间对气候的喜好程度, 也是模型模拟结果中末次盛冰期的樟属物种分布区面积稍高于末次间冰期, 并在陕西秦岭山脉以北的关中平原、湖北中部、江苏东部等高纬度低海拔地区出现局部收缩, 在秦岭、大别山、大巴山、巫山等山脉附近出现扩张的可能原因之一。
樟属物种丰富度热点区域在5个时期都主要分布于华南和西南(24° N以南), 并且不同时期变化比较明显, 变化范围为8.4%-96%。值得注意的是末次盛冰期时樟属物种丰富度热点区域面积在5个时期中最大, 这可能有以下几点原因: 第一, 我国华南大部分地区作为亚热带常绿阔叶林冰期时的避难所(Myerset al, 2000; 应俊生, 2001), 大多数分布在此地区的樟属物种受末次盛冰期的影响很小, 其分布区较其他时期无明显变化, 例如阴香、黄樟等; 第二, 东亚气候模拟结果发现, 末次盛冰期时中国大陆地表气温降低2-9℃, 但在我国南方地区却出现了局部的弱升温(Pinot et al, 1999), 分布在华南地区喜热的樟属物种分布区出现了小范围扩张, 例如八角樟(C. illicioides)、卵叶桂(C. rigidissimum)和滇南桂等; 第三, 末次盛冰期时某些物种向南或低海拔迁移到大型避难所中生存, 冰期后又向北或高海拔适宜生境地区迁移(Hewitt, 2000; Tiffney & Manchester, 2001), 例如银叶桂、屏边桂等。
虽然随着未来(2080年)温室气体的持续排放, 中国气候逐渐变暖变湿, 平均温度会增加3.5℃, 降水量增加7.5%, 温度和降水都较当前有小幅度提升(江志红等, 2008), 但华南地区将温度升高、干旱加剧(赵宗慈等, 2008; Yin et al, 2018)。在这样的气候变化情景下, 分布在华南地区以北的亚热带常绿阔叶林物种将出现潜在分布区增加的趋势, 如樟科的毛豹皮樟(Litsea coreana)、壳斗科的短尾柯(Lithocarpus brevicaudatus); 而分布在华南地区的亚热带常绿阔叶林物种则出现潜在分布区减少的趋势, 例如樟科的华南木姜子(Litsea greenmaniana)、壳斗科的木姜叶柯(Lithocarpus litseifolius)、烟斗柯(L. corneus) (郑维艳, 2019; 郑维艳和曹坤芳, 2020)。通过对樟属植物的分布区进行模拟发现, 在未来温室气体排放情景下, 樟属植物中同时存在潜在分布区增加和减少的物种; 其中分布在较北地区的樟属物种, 例如云南樟、天竺桂和阔叶樟呈现潜在分布区增加的趋势, 分布在华南地区的樟属物种, 例如红辣槁树、软皮桂(C. liangii)、沉水樟(C. micranthum)、米槁等的潜在分布区则呈现逐渐缩减、破碎化的趋势。由于樟属的物种分布中心位于我国热带北部和亚热带南部等华南地区(图1; 郑维艳, 2019(② 郑维艳 (2019) 中国大陆北热带及亚热带地区樟科、壳斗科物种多样性格局及潜在分布预测分析. 硕士学位论文, 广西大学, 南宁.)), 且通过MaxEnt模拟得出的环境因子贡献率和主成分分析结果可知, 相较于温度因子, 樟属植物受降水的影响更大(附录1和附录3)。因此, 在未来华南地区温度升高、干旱加剧的情景下, 模拟结果得出樟属植物潜在分布区减少的物种多于增加的物种, 导致未来两种情景下位于华南地区的樟属物种丰富度热点区域面积均小于当前, 并且相较于RCP2.6情景, RCP8.5情景下樟属物种丰富度热点区域向华南地区收缩的趋势更加明显。这一结果意味着温室气体的持续排放将在一定程度上迫使我国华南地区的亚热带常绿阔叶林面临不断缩减的风险。
3.2 对自然保护地的建议
根据5个时期的模型模拟结果可知, 樟属物种的潜在分布区面积在各个时期并无较大变化, 但物种丰富度热点区域在各个时期有明显不同的分布格局, 因此针对樟属植物的物种丰富度热点区域进行保护评估与规划是关键所在。本研究结果表明, 中国樟属植物的物种丰富度热点区域主要分布在四川东南部、贵州南部、广西和广东, 且仅有少部分热点区域分布在自然保护区内, 还存在大量的保护空缺, 而且这些保护区大多数非国家级, 在人力、财力、物力和科研方面均弱于国家级自然保护区(冯斌等, 2020)。
广西河池市植被类型主要为中亚热带常绿阔叶林和南亚热带季风常绿阔叶林, 其森林覆盖率为65.7%, 呈现中部地区向东西逐渐增高(李玉清, 2017 (③ 李玉清 (2017) 广西河池市森林风景资源评价研究. 硕士学位论文, 中南林业科技大学, 长沙.); 黄肖寒等, 2018); 该市建有2个国家级自然保护区, 保护了该区域内7%的樟属物种丰富度热点区域, 但由于河池市的樟属物种丰富度热点区域面积最大(占该市面积的71.13%), 并且樟属物种组成与其他地区有较大差异, 因此, 建议在其巴马瑶族自治县和都安瑶族自治县增加保护区的规划。柳州市是广西中部地区重要的工业城市, 工业化和人类活动对植被的破坏非常严重, 森林植被亟需保护(文和群等, 1992)。受区域地貌特征的影响, 柳州市城镇体系呈现为“南重北轻, 中心偏南”的格局(
从现有保护区的整体分布来看, 我国南方的保护区总体上呈现片断化不连续格局, 并未建立较为连续的自然保护区, 今后应在多样性高的区域优先进行自然保护区网络建设, 提高相邻自然保护区的连通性(Pouzols & Moilanen, 2014; Tarabon et al, 2021)。根据《生物多样性公约》《2020年后全球生物多样性框架》初稿的要求, 到2030年自然保护区和其他有效区域保护措施的面积要占到陆地国土面积的30% (
3.3 研究局限性及展望
尽管温度和降水是影响植物分布最具决定性的环境因子, 且具有大范围、长期与稳定的观测数据, 进而被广泛研究(Gallagher, 2016; Yu et al, 2017a); 但地形、土壤、植被覆盖因子和人口密度等对植物的分布和生态位模拟的结果都有一定的影响(Cantónet al, 2004; John et al, 2007; Huang et al, 2020)。另外, 我国于2020年在第75届联合国大会一般性辩论上提出CO2排放力争于2030年前达到峰值, 2060年前实现碳中和(王灿和张雅欣, 2020), 这对全球气候变化的改善起到推动作用, 能够有效地缓解全球升温趋势, 使得全球温度比预期降低0.2-0.3℃左右(
因此, 本研究还缺乏环境因子数据的完整性和确定性, 以及研究对象的综合性。今后可从以下几个方面进行完善: (1)在生态位模拟中结合除19个气候因子之外的其他可获得的环境因子, 诸如土壤、地形以及生物因子; (2)依据世界各国的碳达峰和碳中和目标, 期待更多的大气环流模拟研究, 并以最新的大气环流模型来对物种的潜在分布区进行模拟和探讨; (3)应尽可能结合多类群如壳斗科、山茶科、木兰科、樟科等亚热带常绿阔叶林中的优势类群, 来进行气候变化对亚热带常绿阔叶林的影响及保护评估研究。
附录 Supplementary Material
附录1 47种樟属植物及当前气候条件下排名前3的生物气候因子贡献率
Appendix 1 The contribution of the top 3 bioclimatic variables of the 47 species in Cinnamomum
附录2 19个环境变量及其描述
Appendix 2 The 19 bioclimatic variables and their descriptions
附录3 3个主成分的方差解释率、13个环境因子的负载值及Pearson相关性系数
Appendix 3 The variance percentage of 3 principal components, load values of 13 environmental factors and Pearson correlation index
附录4 樟属物种丰富度热点区域的物种组成
Appendix 4 Species composition of species richness hotspots of Cinnamomum
参考文献
Climate Change 2013: The physical science basis. An overview of the Working Group 1 contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC)
Uses and misuses of bioclimatic envelope modeling
DOI:10.1890/11-1930.1 URL [本文引用: 1]
Biodiversity: Where's hot and where's not
DOI:10.1046/j.1523-1739.2003.02344.x URL [本文引用: 1]
Geographical patterns in phylogenetic diversity of Chinese woody plants and its application for conservation planning
DOI:10.1111/ddi.v27.1 URL [本文引用: 2]
Managing forests for climate change mitigation
DOI:10.1126/science.1155458 URL [本文引用: 1]
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain
DOI:10.1016/S0341-8162(03)00108-5 URL [本文引用: 1]
The scree test for the number of factors
DOI:10.1207/s15327906mbr0102_10 URL [本文引用: 1]
Constructing ecological security patterns in Yunfu City based on the framework of importance-sensitivity-connectivity
基于“重要性-敏感性-连通性”框架的云浮市生态安全格局构建
DOI:10.11821/dlyj201703006
[本文引用: 1]

生态安全格局构建是保障城市生态安全,实现城市可持续性发展的重要途径,也是景观生态学研究的热点领域之一。生态源地识别与空间阻力面构建一直是生态安全格局构建中的技术难点。以广东省云浮市为例,从生态系统服务重要性、生态敏感性与景观连通性三个方面识别生态源地,利用DMSP/OLS夜间灯光数据修正基本生态阻力面,并运用最小累积阻力模型判定生态廊道,从而综合构建云浮市生态安全格局。研究表明:云浮市生态源地占全市总面积的36.47%,主要分布在西部与南部的山林地。云浮市生态廊道总长度为508.87 km,其中景观廊道总长度为315.58 km,组团廊道总长度为193.29 km,呈环状辐射分布于植被覆盖相对较好的山区地带。全市16个自然保护区基本都位于生态源地范围内。构建的“重要性—敏感性—连通性”框架可以为区域生态安全格局构建提供新思路,从而有效指引相关空间规划。
Distribution of nature reserves and status of biodiversity protection in China
DOI:10.3724/SP.J.1003.2009.08324 URL [本文引用: 1]
中国自然保护区分布现状及合理布局的探讨
DOI:10.3724/SP.J.1003.2009.08324
[本文引用: 1]
为了保护中国丰富的生物多样性, 我国已经建立了大量的自然保护区。评价这些保护区的布局对于生物多样性的有效保护无疑是十分重要的。本文收集了截至2007年底我国建立的2,047个保护区的有关资料, 利用地理信息系统技术, 分析了这些保护区的分布现状和生物多样性的保护状况, 包括保护的植被类型、野生保护物种以及热点地区。结果表明: 我国自然保护区的覆盖面积达到145.7万km2, 占中国陆地面积的15.2%, 超过世界平均水平(13.4%); 在我国47种自然植被类型中, 有21种植被类型的被保护面积比例低于10%, 说明这些类型可能没有得到充分的保护。应用Dobson筛除算法对216个保护区中的保护物种进行筛除分析, 发现仅西双版纳、武夷山、长白山、高黎贡山、祁连山5个保护区即包含了381个保护物种(约占总数783种的50%); 前21个保护区可包含占总数75%的保护物种(590种)。根据不同方案划分的生物多样性热点保护地区仍存在一些保护空缺地, 如新疆北部、四川与长江以南地区, 因此, 我国的保护区布局有待进一步改进。
Predicting the responses of forest distribution and aboveground biomass to climate change under RCP scenarios in Southern China
DOI:10.1111/gcb.2016.22.issue-11 URL [本文引用: 1]
Lessepsian fish invasion in Mediterranean marine protected areas: A risk assessment under climate change scenarios
DOI:10.1093/icesjms/fsz207 URL [本文引用: 1]
A new material of Lindera (Lauraceae) of the Late Pliocene from Tengchong, Yunnan and the genus' biogeography significance
DOI:10.1111/1755-6724.12081 URL [本文引用: 1]
The biogeographic history of beech trees
DOI:10.1016/j.revpalbo.2009.08.007 URL [本文引用: 1]
Human pressures predict species' geographic range size better than biological traits
DOI:10.1111/gcb.12834 URL [本文引用: 1]
Novel methods improve prediction of species' distributions from occurrence data
DOI:10.1111/j.2006.0906-7590.04596.x URL [本文引用: 1]
Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth
DOI:10.1111/gcb.12512 URL [本文引用: 1]
Evaluation on nature reserve management effectiveness of mitigation and adaptation on climate change: A case study of 12 typical nature reserves in Guangxi
DOI:10.17520/biods.2019352 URL [本文引用: 1]
自然保护区减缓和适应气候变化的管理有效性评估: 以广西12个典型自然保护区为例
Correlates of range size variation in the Australian seed-plant flora
DOI:10.1111/jbi.12711 URL [本文引用: 1]
Climate-growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia
DOI:10.1007/s00468-008-0221-z URL [本文引用: 1]
From glacial refugia to wide distribution range: Demographic expansion of Loropetalum chinense (Hamamelidaceae) in Chinese subtropical evergreen broadleaved forest
DOI:10.1007/s13127-015-0252-4 URL [本文引用: 1]
Integrated modeling to identify priority areas for the conservation of the endangered plant species in headwater areas of Asia
DOI:10.1016/j.ecolind.2019.05.064 URL [本文引用: 1]
The genetic legacy of the Quaternary ice ages
Global climate has fluctuated greatly during the past three million years, leading to the recent major ice ages. An inescapable consequence for most living organisms is great changes in their distribution, which are expressed differently in boreal, temperate and tropical zones. Such range changes can be expected to have genetic consequences, and the advent of DNA technology provides most suitable markers to examine these. Several good data sets are now available, which provide tests of expectations, insights into species colonization and unexpected genetic subdivision and mixture of species. The genetic structure of human populations may be viewed in the same context. The present genetic structure of populations, species and communities has been mainly formed by Quaternary ice ages, and genetic, fossil and physical data combined can greatly help our understanding of how organisms were so affected.
Geographic distribution and potential distribution estimation of Machilus breviflora
短序润楠的地理分布及潜在分布区估计
Refugia and their identification methods: A review
生物避难所及其识别方法评述
Origins and evolution of cinnamon and camphor: A phylogenetic and historical biogeographical analysis of the Cinnamomum group (Lauraceae)
DOI:10.1016/j.ympev.2015.12.007 URL [本文引用: 2]
NDVI simulation of vegetation in Hechi based on climate information
基于气候信息的河池市植被NDVI模拟
Scenario analyses on the changes of future surface air temperature and precipitation in South China
华南地区未来地面温度和降水变化的情景分析
Mapping biodiversity conservation priorities for protected areas: A case study in Xishuangbanna Tropical Area, China
DOI:10.1016/j.biocon.2020.108741 URL [本文引用: 1]
Global patterns of terrestrial vertebrate diversity and conservation
Evolutionary history of subtropical evergreen broad-leaved forest in Yunnan Plateau and adjacent areas: An insight from Quercus schottkyana (Fagaceae)
Projection of climate change in China in the 21st century by IPCC-AR4 Models
IPCC-AR4模式对中国21世纪气候变化的情景预估
Soil nutrients influence spatial distributions of tropical tree species
Advances in Lauraceae systematic research on the world scale
世界樟科植物系统学研究进展
Species range size patterns and their significance on biodiversity conservation
DOI:10.1360/N052018-00230 URL [本文引用: 1]
物种分布区特征及其对生物多样性保育的意义
Lauraceae
Conservation Value Evaluation and Reasonable Distribution of National Nature Reserve for Forest
国家级森林类型自然保护区保护价值评价及合理布局研究
Assessing climate change impact on forest habitat suitability and diversity in the Korean Peninsula
DOI:10.3390/f9050259 URL [本文引用: 1]
Evolutionary history of the angiosperm flora of China
DOI:10.1038/nature25485
[本文引用: 1]
High species diversity may result from recent rapid speciation in a 'cradle' and/or the gradual accumulation and preservation of species over time in a 'museum'(1,2). China harbours nearly 10% of angiosperm species worldwide and has long been considered as both a museum, owing to the presence of many species with hypothesized ancient origins(3,4), and a cradle, as many lineages have originated as recent topographic changes and climatic shifts-such as the formation of the Qinghai-Tibetan Plateau and the development of the monsoon-provided new habitats that promoted remarkable radiation(5). However, no detailed phylogenetic study has addressed when and how the major components of the Chinese angiosperm flora assembled to form the present-day vegetation. Here we investigate the spatio-temporal divergence patterns of the Chinese flora using a dated phylogeny of 92% of the angiosperm genera for the region, a nearly complete species-level tree comprising 26,978 species and detailed spatial distribution data. We found that 66% of the angiosperm genera in China did not originate until early in the Miocene epoch (23 million years ago (Mya)). The flora of eastern China bears a signature of older divergence (mean divergence times of 22.04-25.39 Mya), phylogenetic overdispersion (spatial co-occurrence of distant relatives) and higher phylogenetic diversity. In western China, the flora shows more recent divergence (mean divergence times of 15.29-18.86 Mya), pronounced phylogenetic clustering (co-occurrence of close relatives) and lower phylogenetic diversity. Analyses of species-level phylogenetic diversity using simulated branch lengths yielded results similar to genus-level patterns. Our analyses indicate that eastern China represents a floristic museum, and western China an evolutionary cradle, for herbaceous genera; eastern China has served as both a museum and a cradle for woody genera. These results identify areas of high species richness and phylogenetic diversity, and provide a foundation on which to build conservation efforts in China.
Study on conservation and construction of biodiversity (plants) in Leshan City
乐山市生物多样性(植物)保护与建设的研究
Survey of Wuyou Mountain evergreen broad-leaved forest
乌尤山常绿阔叶林概况
Chasing ghosts: Allopolyploid origin of Oxyria sinensis (Polygonaceae) from its only diploid congener and an unknown ancestor
DOI:10.1111/mec.2017.26.issue-11 URL [本文引用: 1]
Advances in the effects of climate change on the distribution of plant species and vegetation in China
气候变化对植物及植被分布的影响研究进展
Hotspots assessment and conservation priorities identification of biodiversity in China should be emphasized
中国生物多样性热点地区(hotspot)评估与优先保护重点的确定应该重视
Phylogeography of Davidia involucrata (Davidiaceae) inferred from cpDNA haplotypes and nSSR data
A practical guide to MaxEnt for modeling species' distributions: What it does, and why inputs and settings matter
DOI:10.1111/ecog.2013.36.issue-10 URL [本文引用: 1]
Impact of climate warming on the northern boundary of sub-tropical zone of China
气候变暖对中国亚热带北界位置的影响
Effect of South American grazing camelids on soil fertility and vegetation at the Bolivian Andean grasslands
DOI:10.1016/j.agee.2015.04.005 URL [本文引用: 1]
The biodiversity challenge: Expanded hot-spots analysis
This paper aims to throw light on the mass extinction that is overtaking Earth's species. Using an analytic methodology developed for an earlier partial assessment, it focuses on a series of "hotspot" areas, these being areas that a) feature exceptional concentrations of species with high levels of endemism and b) face exceptional threats of destruction. The paper identifies another 8 such areas, 4 of them in tropical forests and 4 in Mediterranean-type zones. The analysis reveals that the 4 tropical-forest areas contain at least 2835 endemic plant species in 18,700 sq. km, or 1.1% of Earth's plant species in 0.013% of Earth's land surface; and that the 4 Mediterranean-type areas contain 12,720 endemic plant species in 435,700 sq. km, or 5.1% of Earth's plant species in 0.3% of the Earth's land surface. Taken together, these 8 hotspot areas contain 15,555 endemic plant species in 454,400 sq. km, or 6.2% of Earth's plant species in 0.3% of Earth's land surface. This is to be compared with the earlier hotspot analysis of 10 tropical-forest areas, with 34,400 endemic plant species in 292,00 sq. km, or 13.8% of Earth's plant species in 0.2% of Earth's land surface. Taking all 18 hot-spot areas together, the authors find they support 49,995 endemic plant species, or 20% of Earth's plant species, in 746,400 sq. km, or 0.5% of Earth's land surface. This means that one-fifth of Earth's plant species are confined to 0.5% of the Earth's land surface--and they occur in habitats that are mostly threatened with imminent destruction. By concentrating on these hotspot areas where needs are greatest and where the pay-off from safeguard measures would be greatest, conservationists can engage in a more systematized response to the challenge of large-scale impending extinctions.
Biodiversity hotspots for conservation priorities
Conservationists are far from able to assist all species under threat, if only for lack of funding. This places a premium on priorities: how can we support the most species at the least cost? One way is to identify 'biodiversity hotspots' where exceptional concentrations of endemic species are undergoing exceptional loss of habitat. As many as 44% of all species of vascular plants and 35% of all species in four vertebrate groups are confined to 25 hotspots comprising only 1.4% of the land surface of the Earth. This opens the way for a 'silver bullet' strategy on the part of conservation planners, focusing on these hotspots in proportion to their share of the world's species at risk.
The water-temperature distributional groups of dominants and companions of subtropical evergreen broadleaved forest in China
中国亚热带常绿阔叶林优势种及常见种的水热分布类群
利用目前国际上比较常用的植被—气候相关分析的气候指标,如Kira的温暖指数和寒冷指数,徐文铎的湿润指数,Penman的可能蒸散和干燥度指标,Thornthwaite的潜在可能蒸散和水分指数,Holdridge生命地带分类系统指标以及气温和降水等单一气候因子,综合对中国亚热带常绿阔叶林优势种及常见种进行TWINSPAN分类和DCA排序,可将植物种分为8个水热分布类群,较好地反映出优势种及常见种沿热量和水分梯度的分布格局。并总结了这8个水热分布类群的气候指标范围。这8个类群是:Ⅰ高温湿润型,Ⅱ高中温湿润型,Ⅲ低中温湿润型,Ⅳ高低温中湿型,Ⅴ低低温中湿型,Ⅵ低温半湿润型,Ⅶ高低温低湿型,Ⅷ低低温低湿型
Modelling the vegetation of China using the process-based equilibrium terrestrial biosphere model BIOME3
DOI:10.1046/j.1365-2699.2000.00206.x URL [本文引用: 1]
Palaeovegetation in China during the late Quaternary: Biome reconstructions based on a global scheme of plant functional types
DOI:10.1016/j.palaeo.2010.02.008 URL [本文引用: 1]
Beyond Kyoto: Forest management in a time of rapid climate change
DOI:10.1046/j.1523-1739.2001.015003578.x URL [本文引用: 1]
A globally coherent fingerprint of climate change impacts across natural systems
Causal attribution of recent biological trends to climate change is complicated because non-climatic influences dominate local, short-term biological changes. Any underlying signal from climate change is likely to be revealed by analyses that seek systematic trends across diverse species and geographic regions; however, debates within the Intergovernmental Panel on Climate Change (IPCC) reveal several definitions of a 'systematic trend'. Here, we explore these differences, apply diverse analyses to more than 1,700 species, and show that recent biological trends match climate change predictions. Global meta-analyses documented significant range shifts averaging 6.1 km per decade towards the poles (or metres per decade upward), and significant mean advancement of spring events by 2.3 days per decade. We define a diagnostic fingerprint of temporal and spatial 'sign-switching' responses uniquely predicted by twentieth century climate trends. Among appropriate long-term/large-scale/multi-species data sets, this diagnostic fingerprint was found for 279 species. This suite of analyses generates 'very high confidence' (as laid down by the IPCC) that climate change is already affecting living systems.
Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar
DOI:10.1111/j.1365-2699.2006.01594.x URL [本文引用: 3]
Maximum entropy modeling of species geographic distributions
DOI:10.1016/j.ecolmodel.2005.03.026 URL [本文引用: 1]
How to protect half of Earth to ensure it protects sufficient biodiversity
Tropical paleoclimates at the Last Glacial Maximum: Comparison of Paleoclimate Modeling Intercomparison Project (PMIP) simulations and paleodata
DOI:10.1007/s003820050318 URL [本文引用: 1]
A method for building corridors in spatial conservation prioritization
DOI:10.1007/s10980-014-0031-1 URL [本文引用: 1]
Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation
DOI:10.1111/gcb.13090
PMID:26367278
[本文引用: 1]
Assuming that co-distributed species are exposed to similar environmental conditions, ecological niche models (ENMs) of bird and plant species inhabiting tropical dry forests (TDFs) in Mexico were developed to evaluate future projections of their distribution for the years 2050 and 2070. We used ENM-based predictions and climatic data for two Global Climate Models, considering two Representative Concentration Pathway scenarios (RCP4.5/RCP8.5). We also evaluated the effects of habitat loss and the importance of the Mexican system of protected areas (PAs) on the projected models for a more detailed prediction of TDFs and to identify hot spots that require conservation actions. We identified four major distributional areas: the main one located along the Pacific Coast (from Sonora to Chiapas, including the Cape and Bajío regions, and the Balsas river basin), and three isolated areas: the Yucatán peninsula, central Veracruz, and southern Tamaulipas. When considering the effect of habitat loss, a significant reduction (~61%) of the TDFs predicted area occurred, whereas climate-change models suggested (in comparison with the present distribution model) an increase in area of 3.0-10.0% and 3.0-9.0% for 2050 and 2070, respectively. In future scenarios, TDFs will occupy areas above its current average elevational distribution that are outside of its present geographical range. Our findings show that TDFs may persist in Mexican territory until the middle of the XXI century; however, the challenges about long-term conservation are partially addressed (only 7% unaffected within the Mexican network of PAs) with the current Mexican PAs network. Based on our ENM approach, we suggest that a combination of models of species inhabiting present TDFs and taking into account change scenarios represent an invaluable tool to create new PAs and ecological corridors, as a response to the increasing levels of habitat destruction and the effects of climate change on this ecosystem. © 2015 John Wiley & Sons Ltd.
Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world's most diverse temperate flora
DOI:10.1016/j.ympev.2011.01.012 URL [本文引用: 2]
Making better Maxent models of species distributions: Complexity, overfitting and evaluation
DOI:10.1111/jbi.12227 URL [本文引用: 1]
RCP 8.5—A scenario of comparatively high greenhouse gas emissions
DOI:10.1007/s10584-011-0149-y URL [本文引用: 1]
Phylogeography of a widespread Asian subtropical tree: Genetic east-west differentiation and climate envelope modelling suggest multiple glacial refugia
DOI:10.1111/jbi.12322 URL [本文引用: 1]
Distribution patterns of the subtropical evergreen broad-leaved forests of southwestern China, as compared with those of the eastern Chinese subtropical regions
DOI:10.3989/collectbot.2015.v34 URL [本文引用: 1]
Biodiversity in China's mountains
DOI:10.1890/1540-9295(2006)004[0347:BICM]2.0.CO;2 URL [本文引用: 1]
Pooling biodiversity offsets to improve habitat connectivity and species conservation
DOI:10.1016/j.jenvman.2020.111425 URL [本文引用: 1]
The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the Northern Hemisphere Tertiary
Buffered tree population changes in a quaternary refugium: Evolutionary implications
RCP2.6: Exploring the possibility to keep global mean temperature increase below 2℃
Implementation pathway and policy system of carbon neutrality vision
碳中和愿景的实现路径与政策体系
Geographical Distribution Pattern and Spatial Conservation Prioritization for Wild Plants in China under Climate Change
气候变化背景下中国野生植物地理分布格局及优先保护
Introduction and cultivation techniques of silverwood
银木的引种及栽培技术
Phytogeography and Species Diversity of Typical Evergreen Broad-leaved in China
中国常绿阔叶林植物地理与物种多样性研究
China's forest coverage rate is 22.96%
中国森林覆盖率22.96%
A preliminary study on the vegetation in Longtan and Dule parks in Liuzhou City
柳州市龙潭、都乐公园植被的初步研究
Impact of climate change on potential distribution range and spatial pattern of Phoebe chekiangensis
气候变化对浙江楠潜在分布范围及空间格局的影响
Biogeographical diversification of mainland Asian Dendrobium (Orchidaceae) and its implications for the historical dynamics of evergreen broad-leaved forests
DOI:10.1111/jbi.12726 URL [本文引用: 1]
Hotspot analyses indicate significant conservation gaps for evergreen broadleaved woody plants in China
DOI:10.1038/s41598-017-02098-0 URL [本文引用: 1]
Differential Quaternary dynamics of evergreen broadleaved forests in subtropical China revealed by phylogeography of Lindera aggregata (Lauraceae)
DOI:10.1111/jbi.2019.46.issue-6 URL [本文引用: 1]
Climate change risk to forests in China associated with warming
DOI:10.1038/s41598-017-18798-6 URL [本文引用: 1]
Species diversity and distribution pattern of seed plants in China
中国种子植物物种多样性及其分布格局
Climatic niche breadth can explain variation in geographical range size of alpine and subalpine plants
DOI:10.1080/13658816.2016.1195502 URL [本文引用: 1]
Rhododendron diversity patterns and priority conservation areas in China
DOI:10.1111/ddi.12607 URL [本文引用: 1]
Spatial patterns of species diversity of Lauraceae plants in Yunnan, China
云南樟科植物多样性的空间分布格局
Assessment and prediction of precipitation and droughts/floods changes over the world and in China
全球和中国降水、旱涝变化的检测评估
Ornamental tree species resources of Lauraceae and their application in gardens
樟科植物观赏树种资源及其在园林中的应用
Camphortree and the morphological characteristics of cold north correlation
樟树北移耐寒性与形态特征的相关性研究
Distribution and conservation of threatened plants in China
DOI:10.1016/j.biocon.2015.10.019 URL [本文引用: 3]
Species diversity and biogeographical patterns of Lauraceae and Fagaceae in northern tropical and subtropical regions of China
中国大陆北热带及亚热带地区樟科、壳斗科物种多样性及其生物地理格局分析
Impact of future climate change on potential geographical distribution of four Litsea species in China
未来气候变化对四种木姜子地理分布的影响
Patterns of genotype variation and demographic history in Lindera glauca (Lauraceae), an apomict-containing dioecious forest tree
DOI:10.1111/jbi.v47.9 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}