


植物功能性状是指能够响应生存环境的变化, 并对生态系统功能有一定影响的植物性状(Díaz & Cabido, 2001)。McGill等(2006)认为功能性状对生物体的行为表现影响很大, 并提出了利用功能性状重建群落生态学研究的新思路。植物功能性状的研究已经深入到生态学的许多领域, 其中最为关键的一个方面就是探索性状和环境之间的关系, 为研究生态系统功能的驱动因子奠定基础, 也为预测在全球变化前提下的生态系统响应提供方法和依据(Wright et al., 2005; Beaumont & Burns, 2009)。
国内外的生态学家对植物功能性状同环境之间的关系已进行了大量的研究。在全球尺度上, 生活在不同生态系统类型中的植物在功能性状上都存在着显著差异(Reich et al., 1999; Villar & Merino, 2001)。但Wright等(2004)依据全球175个地点的2,548种植物所建立的全球叶片经济型谱, 发现在全球尺度上, 维管植物叶片在化学成分、结构及生理过程等特性之间存在着紧密的联系, 尽管某些性状与气候因子间存在显著的相关关系, 但从全球整体格局来看, 气候对性状的调节作用并不十分明显。在区域尺度上, 植物功能性状与环境的关系表现得更为明显(Díaz et al., 1998)。如Craine和Lee (2003)发现新西兰岛上118种植物的叶厚度随海拔升高而增加。而在北极地区, 土壤养分含量对植物叶性状、繁殖性状等都有影响(Dormann & Woodin, 2002)。
同一种植物在不同地点, 其叶片的形态、解剖结构以及水分利用状况都可能发生变异(Gratani et al., 2003); 而不同种植物却可能因为相似的环境选择作用, 表现出相似的功能性状响应(Meinzer, 2003)。我国学者对植物功能性状和植物功能型方面的研究也取得了不少的进展。例如Han等(2005)发现中国753种陆生植物随着纬度增高、年均温降低, 叶片氮、磷含量均有增加, 但氮/磷比变化不显著。He等(2006)研究了青藏高原74种高寒植物的叶性状, 发现植物的功能性趋同的特点在极端环境下同样适用。Luo等(2005)分析了从热带/亚热带到温带/高山地区6个森林群落的性状数据, 发现温度和降水对林冠层平均叶寿命有显著影响。
冯云等(2008)通过DCCA排序, 发现海拔是影响东灵山辽东栎(Quercus wutaishanica)林物种分布的主要环境因子。刘志民等(2000)发现随着海拔上升, 青藏高原几种常见植物叶片的叶绿素含量明显增加。祁建等(2007)发现辽东栎的叶绿素含量、气孔密度等生理生态指标与海拔等地形因子具有较为显著的相关性, 叶形态性状也随海拔而变化。李芳兰等(2007)发现生长在岷江上游干旱河谷的四川黄栌(Cotinus szechuanensis)的叶片厚度随海拔上升而增大。植物功能性状的这些变化, 都是植物对环境异质性长期适应的结果。
关于植物功能性状的研究仍然有许多问题亟待解决。首先, 大尺度上的研究使得许多重要的环境因子可能被忽略。Wright等(2005)发现气候因子大约能够解释植物叶性状变异的18%; 而如果将研究尺度缩小, 生境或微地形变量或许能够解释大尺度研究中所无法解释的环境过滤效应(Kraft et al., 2008)。在野外条件下, 影响植物功能性状的主要环境因子, 如光照、温度、降水、养分等的差别, 集中体现在地形(如海拔、坡度、坡向等)和土壤状况的差别上(Díaz et al., 1998)。尤其在小尺度上, 微生境的差别能够直接影响光照、温度、湿度、土壤微生物等环境因子, 进而影响植物的生理生态过程、生长与繁殖, 乃至分布格局。
其次, 目前研究较多的主要是一些易于测量的形态性状, 如叶面积、叶长/宽比等。这些形态性状又称作软性状(soft traits), 它们在响应环境变化方面要比生理性状滞后, 使得环境因子对植物功能的作用不能得到最为灵敏的体现。而生理生态性状(如光合作用、呼吸作用、蒸腾作用、叶绿素荧光动力学等)在一些重要的生态过程方面起着更为重要的作用(孟婷婷等, 2007), 在反映小尺度的环境变化方面, 无疑要比形态性状敏感。由于测量生理性状的技术手段较为复杂, 测量周期长, 实验仪器昂贵, 以及在野外运输困难等原因, 其测定一直受到较大的限制。在世界上许多研究热点地区, 尤其是在森林动态监测样地的研究中, 有关群落组成的研究已经比较成熟了, 但生理性状与微环境方面的信息却相对缺乏(Reich et al., 2007)。因此, 研究小尺度范围内植物生理生态性状和微环境之间的关系, 对揭示微环境对植物功能的影响以及植物对环境因子的响应具有重要的意义。
浙江省古田山24 ha森林长期监测样地是中国森林生物多样性监测网络(CForBio)的重要组成部分, 样地内生境异质性大, 地形复杂, 背景数据调查较为详尽(赖江山, 2008), 为研究小尺度下环境因子对植物功能性状的影响提供了良好的条件。本研究旨在通过分析古田山样地内地形和土壤特性与植物的形态性状及生理性状的关系, 探讨了小尺度下微环境因子是如何影响植物功能性状的变化的, 并试图揭示影响古田山亚热带常绿阔叶林植物形态和生理性状的主要环境驱动因子。
1 方法
1.1 取样方法
研究地点位于古田山国家级自然保护区, 地处浙江省衢州市开化县(29°10′19.4′′-29°17′41.4′′ N, 118°03′49.7′′-118°11′12.2′′ E)。保护区内绝大多数土壤呈酸性, pH值在5.5-6.5之间。样地建设的方法参照CTFS(Center for Tropical Forest Science)样地建设方案进行(Condit, 1995)。整个样地东西长600 m、南北长400 m, 保存着典型的天然亚热带常绿阔叶林, 样地中木本植物(胸径≥1 cm)共159种(祝燕等, 2008)。样地内地形复杂多变, 地势陡峭, 最高海拔为714.9 m, 最低海拔为446.3 m, 平均海拔为580.6 m, 最大高差为268.6 m, 坡度范围为12.8°-62.0°, 平均坡度为37.5°。复杂的地形为本研究提供了理想的地理条件。
样地内功能性状数据的野外取样和采集工作在2008年和2009年夏天进行。在样地全部600个20 m×20 m样方中, 随机选取147个样方(图1), 共计115种被子植物, 其中包括77种乔木, 38种灌木, 1,059株个体, 每个物种个体数不少于3个。取样方案主要参照Cornelissen等(2003)编写的植物功能性状测量手册, 在没有明显遮荫的冠层外部, 选取发育良好、未分枝的当年枝和完全展开的当年叶测量。取样时记录每个个体的标牌号, 以便与样方中的环境数据相结合。地形数据和土壤养分数据来源于中国科学院植物研究所古田山24 ha样地背景调查数据库。
图1
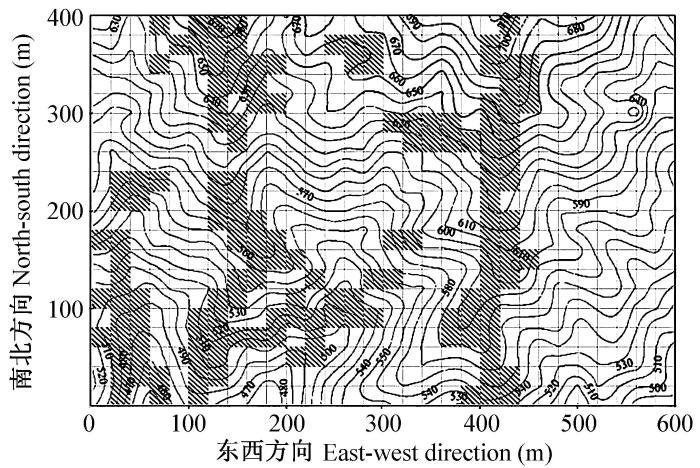
图1
古田山24-ha样地植物功能性状采样地点空间分布图。
图中曲线为等高线, 阴影部分为取样样方。
Fig. 1
Spatial distribution map of sampling quadrats in the Gutianshan 24-ha plot.
Curved lines represent contours, and shadows represent sampling quadrats.
1.2 功能性状的测定
1.2.1 形态性状的测定
选择比叶面积(SLA)、叶长/宽比(LL/LW)、叶厚度(T)和气孔密度(SD)等与植物养分、水分利用有关的4个主要的形态功能性状进行测定。
叶性状的测量方法依照文献Cornelissen等(2003): 在采摘下来的叶子中, 选取3-10片完整且发育良好的叶片, 去掉叶柄后, 使用EPSON V10扫描仪(爱普生公司, 日本)获取叶面积图像, 输入到winFOLIA软件(Hölscher et al., 2002), 测量叶面积、叶长度和叶宽度。随后将新鲜叶片放入60°C的烘箱中烘干72 h以上至恒重, 然后使用1/1000普通电子天平称量干重(精确到0.001 g)。
比叶面积即为叶面积与叶干重之比, 叶长/宽比为叶子长度和宽度之比。叶厚度使用游标卡尺测量(精确到0.01 mm), 每一个体选取3-5片发育良好的叶子同时测量, 测时避开叶脉, 并尽量选择叶子的中心部位进行测量。每个个体在不同部位测量5次, 最后取平均值作为该个体叶厚度。
气孔密度采用印迹法进行取样(Sachs & Novoplansky, 1993): 首先使用透明指甲油涂抹叶片下表皮(避开主脉和大的次级脉), 静置待指甲油干透后, 小心用镊子撕下后置于小封口袋中保存。样品带回实验室后使用Nikon Ri-1(尼康公司, 日本)电子显微镜观测, 在40倍物镜下测量气孔密度。
1.2.2 生理生态性状的测定
根据研究需要, 测定叶绿素含量(Chl)、叶绿素荧光(包含Fv/Fm和PIABS两个参数), 以及枝条比导率(Ksp)等有关植物光合作用和同化能力及枝条的水力传导效率的参数和指标。
叶绿素含量使用便携式叶绿素吸收仪SPAD-502 (Minolta Camera, 日本)测量, 该仪器测出来的数值是叶绿素的相对含量(无量纲)。每片叶子测量3-5个数值, 取均值。叶绿素荧光则使用Handy PEA 叶绿素荧光仪(Hansatech Instruments Ltd., 英国)来测量。
枝条比导率使用自制的水力结构设备, 使用改良的“冲洗法”进行测定(Zimmermann, 1978; Sperry et al., 1988; 徐新武等, 2009)。用橡胶管和密封白胶带将枝条样品固定在该装置上, 上端接10 kPa水压高度的去离子水冲洗液, 并采用真空加压泵(15G 0.4-8, 杭州羊岗泵业有限公司大溪分公司)加压100 kPa, 合计加压110 kPa。枝条下端接电子天平(Satorius, 德国, 精确到0.0001 g)。将待测枝条先在110 kPa的压力下使用去离子水冲洗3 min数次, 排出样品木质部导管中可能存在的空穴化气泡, 然后关闭真空加压泵, 使10 kPa的去离子水流经枝条。记录滴下的水重量, 并记录单位滴水时间, 枝条的水力导度Kh(kg·m·MPa-1·s-1)等于通过该茎段的水流量(F, kg·s-1)与引起木质部水流的压力梯度(ΔP/ΔX, MPa·m-1)的比值(公式1)。水力导度除以该枝条茎段边材截面积(SA, m2), 即为枝条的比导率(Ksp, kg·m-1·MP a-1·s-1, 公式2)。
1.3 数据处理
选取平均海拔、凹凸度、坡度和坡向4个主要的地形参数, 测量数据来源于古田山24 ha样地第一次普查数据库。测量方法主要参照文献Yamakura等(1995)。其中, 平均海拔是指20 m×20 m样方4个顶点海拔的平均值; 坡度是指从该样方的4个顶点中每取3个顶点组成一个平面, 共组成4个不同的平面, 这4个平面与水平面夹角的平均值; 坡向是这4个平面与正北方向角度的平均值; 凹凸度是指该样方的平均海拔减去与该样方相邻的8个样方平均海拔的平均值, 处于样地边缘样方的凹凸度为样方中心的海拔减去4个顶点海拔的平均值。 凹凸度为正值, 说明目标样方比周围样方高, 反之则比周围样方低。
土壤参数选取土壤含水量、土壤全氮含量、土壤全磷含量、土壤全碳含量和土壤pH值, 数据来源于古田山24 ha监测样地土壤分析数据(张俪文, 2010)。
为了找到影响古田山植物功能性状最重要的地形因子, 使用软件CANOCO 4.5(TerBraak & Smilauer, 2002)对实验数据进行了RDA约束排序分析。RDA分析需要两个矩阵, 分别是物种数据和环境数据。物种数据是每个样方所测植物功能性状的平均值, 环境因子矩阵则是指4个地形参数和5个土壤参数。排序之前, 对所有量纲不同的参数都进行了标准化处理, 因此在排序图(biplot)中, 每个环境因子箭头长度所代表的特征向量的长度, 可以看作是环境因子对功能性状的解释量的相对大小。两个箭头的夹角可以看作是环境因子和功能性状的相关性大小。当夹角角度为0°-90°之间时, 两个变量之间呈正相关关系; 当夹角角度为90°-180°之间时, 二者之间呈负相关关系; 当夹角角度为90°时, 表示二者没有显著的相关关系(Lepš & Šmilauer, 2003)。
2 结果
2.1 功能性状的变化
从表1中可以看出, 147个样方中植物功能性状的差异比较大。比如, 叶片叶绿素含量、气孔密度(No./mm2)和叶片厚度(mm)的变化幅度分别为23.0-82.0, 76.63- 970.59 No./mm2和0.02-35.08 mm。
表1 古田山24-ha样地中所测功能性状的变化情况
Table 1
| 叶绿素 Chl | PSⅡ最大 光化学效率 Fv/Fm | 基于光能吸收的PSII光化学综合性能指数 PIABS | 比导率 Ksp (kg·m-1· MPa-1·s-1) | 气孔密度 SD (N·mm-2) | 叶片厚度 T (mm) | 比叶面积 SLA (m2·kg-1) | 叶长/宽比 LL/LW | |
|---|---|---|---|---|---|---|---|---|
| 最小值 Min. | 23.0 | 0.71 | 5.77 | 0.02 | 74.63 | 0.02 | 2.94 | 1.12 |
| 最大值 Max. | 82.0 | 0.84 | 75.13 | 5.01 | 970.59 | 0.60 | 35.08 | 6.86 |
| 均值±标准误 Mean±SE | 50.7±10.7 | 0.79±0.02 | 27.23±11.78 | 0.38±0.45 | 313.45±142.99 | 0.22±0.10 | 13.06±5.28 | 2.65±0.64 |
Chl, Chlorophyll content; Fv/Fm, Maximal photochemical efficiency of photo system II; PIABS, Performance index on basis of light energy absorption; Ksp, Stem sapwood xylem specific hydraulic conductivity; SD, Stomata density; T, Leaf thickness; SLA, Specific leaf area; LL/LW, Ratio of leaf length to leaf width.
2.2 地形和土壤特性的变化
从表2中可以看出, 147个样方的地形异质性非常大, 海拔、凹凸度和坡度的变化幅度分别为11.82-257.68 m、-16.30至18.62 m和14.48°-51.28°。而在土壤特性方面, 土壤全磷含量整体偏低, 土壤呈现较强的酸性。
表2 古田山24-ha样地中地形和土壤特性的变化情况
Table 2
| 地形 Topography | 土壤属性 Soil characteristics | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 平均海拔 Mean elevation (m) | 凹凸度Convexity (m) | 坡度 Slope (°) | 坡向 Aspect (°) | 全碳 TC (g/kg) | 全氮 TN (g/kg) | 全磷 TP (g/kg) | 酸碱度 pH | 含水量 Moisture (%) | ||
| 最小值 Min. | 11.82 | -16.30 | 14.48 | 95.89 | 24.61 | 1.23 | 0.06 | 4.33 | 10.73 | |
| 最大值 Max. | 257.68 | 18.62 | 51.28 | 255.53 | 90.57 | 4.29 | 0.31 | 5.40 | 35.28 | |
| 均值±标准误 Mean±SE | 125.13±69.44 | 1.03±8.30 | 33.44±7.98 | 181.43±38.46 | 39.10±10.98 | 2.04±0.66 | 0.13±0.05 | 4.71±0.19 | 19.65±6.07 | |
TP, TC and TN, Content of total soil carbon, nitrogen and phosphorus, respectively.
2.3 功能性状与地形和土壤特性的关系
对7个植物功能性状与4个地形因子和5个土壤因子的RDA约束排序分析结果见图2。结果表明, 地形因子能够解释10.4%的功能性状变异, 其中平均海拔的影响最大, 接下来依次是凹凸度、坡度和坡向; 土壤因子能解释13.9%的性状变异, 其中影响最大的是土壤含水量, 接下来依次是全氮、pH值、全磷、全碳和氮/磷比。
图2
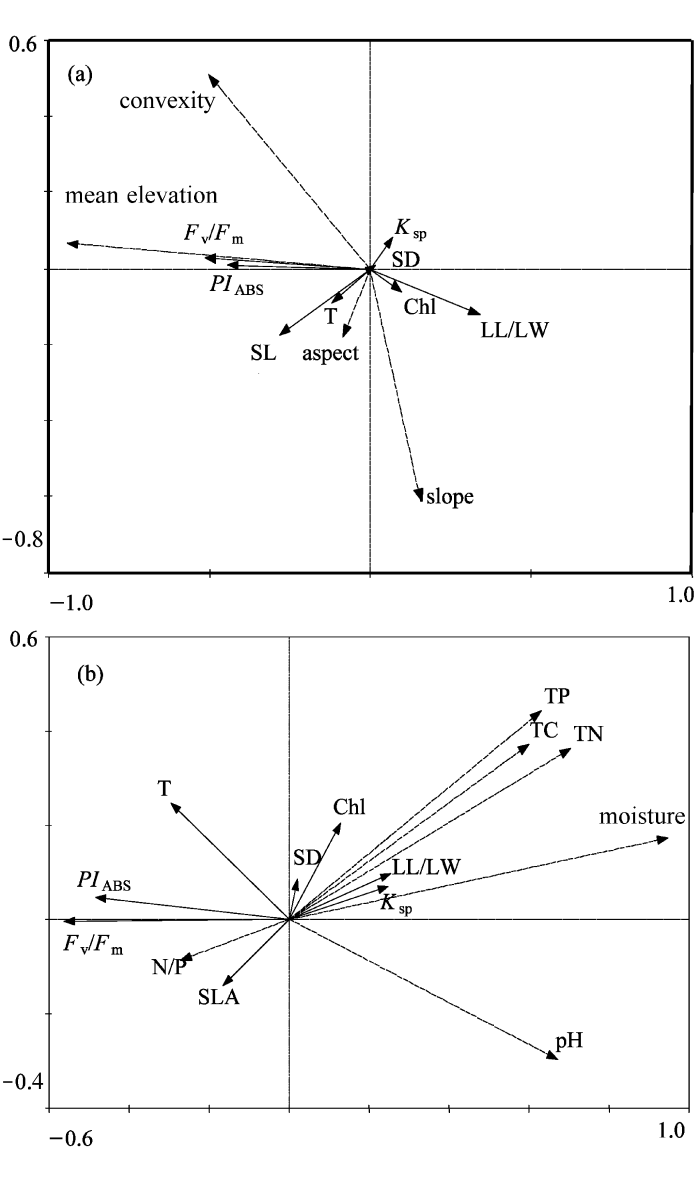
图2
功能性状与地形因子(a)和土壤因子(b)的RDA约束排序分析的双标图。
图中实线特征向量表示功能性状, 虚线特征向量表示地形因子,横纵坐标表示负荷量。Chl, 叶绿素含量; Fv/Fm, PSII的最大光化学效率; PIABS, 基于光能吸收的PSII光化学综合性能指数; Ksp, 枝条比导率; SD, 气孔密度; T, 叶片厚度; SLA, 比叶面积; LL/LW, 叶长/宽比; mean elevation, 平均海拔; convexity, 凹凸度; TC, TN, TP, 土壤全碳、全氮、全磷含量; moisture, 土壤含水量; N/P, 土壤氮/磷比。
Fig. 2
Biplots of RDA analysis between functional traits and topological variations (a), and soil parameters (b).
Functional traits are displayed in solid arrows and topological data in dashed arrows. Chl, Chlorophyll content; Fv/Fm, Maximal photochemical efficiency of photo system II; PIABS, Performance index on basis of light energy absorption; Ksp, Stem sapwood xylem specific hydraulic conductivity; SD, Stomata density; T, Leaf thickness; SLA, Specific leaf area; LL/LW, Ratio of leaf length to leaf width; TP, TC and TN, Content of total soil carbon, nitrogen and phosphorus, respectively; N/P, Ratio of nitrogen to phosphorus.
从排序图中可以看出, 叶绿素含量随海拔和凹凸度的上升而下降, 但与土壤中的氮含量和水分含量呈现正相关。叶绿素荧光参数Fv/Fm和PIABS则与土壤氮磷含量呈现出显著的负相关。枝条比导率与土壤含水量具有较为显著的正相关关系。比叶面积与海拔呈现显著的正比关系。
3 讨论
RDA分析结果表明, 在地形方面, 海拔和凹凸度是影响亚热带常绿阔叶林植物功能性状最重要的两个地形因子; 在土壤方面, 土壤含水量和氮含量则是最重要的因子。地形是多种环境因子的复合, 温度、水分、土壤养分的变化集中体现在地形梯度的变化上, 进而反映在植物功能性状的变化上。尤其在小尺度研究领域, 这些非生物因子的影响尤其明显(Loreau & Naeem, 2001)。
许多研究表明, 当土壤存在氮元素缺乏、干旱等胁迫时, 叶片叶绿素含量会随之下降(Reich et al., 1999; 杨燕等, 2005)。本研究中, 叶绿素含量随着海拔和凹凸度的上升而下降(图2a), 造成这一现象的原因是土壤氮素和含水量与海拔和凹凸度呈现显著的反比关系(表3), 即在海拔越高、凹凸度越大的地方, 由于冲刷等原因, 土壤含氮量和含水量就越低, 进而影响叶片叶绿素的合成。这与Criddle等(1994)的研究结论是一致的。
表3 地形因子与土壤因子Spearman相关分析
Table 3
| 全碳TC (g/kg) | 全氮TN (g/kg) | 全磷TP (g/kg) | 酸碱度 pH | 含水量Moist (%) | |
|---|---|---|---|---|---|
| 平均海拔 Mean elevation (m) | -0.510** | -0.579** | -0.479** | -0.174** | -0.564** |
| 凹凸度 Convexity (m) | -0.611** | -0.719** | -0.714** | -0.545** | -0.693** |
| 坡度 Slope (°) | 0.295** | 0.214** | 0.250** | -0.044 | 0.112** |
| 坡向 Aspect (°) | 0.014 | -0.061* | 0.004 | -0.046 | -0.062* |
TC, TN, TP, Content of total soil carbon, nitrogen and phosphorus, respectively. * P<0.05; ** P<0.01.
已有研究表明, 叶绿素吸收光能并用于驱动光合作用, 超出植物所利用部分的那些被激发的光能, 则以荧光的形式释放出去(Seaton & Walker, 1990)。Fv/Fm是指光系统Ⅱ的最大光化学效率(张守仁, 1999), 代表了植物潜在的太阳能利用效率(Roháček & Barták, 1999)。PIABS是基于光能吸收的光系统II光化学综合性能指数(李鹏民等, 2005)。两个荧光参数没有随着海拔和凹凸度的上升而下降, 表明尽管海拔越高、凹凸度越大的时候, 土壤氮素含量和含水量都越小, 但这样的养分环境对植物的光合作用仍然是适宜的, 古田山没有出现明显的氮素和水分胁迫。
有趣的是, 两个叶绿素荧光参数随着土壤氮素和磷素的上升而下降(图3a, b)。类似的结果也出现在英国的一个酸性土壤草地的长期氮沉降模拟实验中(Arróniz-Crespo et al., 2008)。世界上的许多森林地区, 磷都是最主要的一个限制因素(Attiwill & Adams, 1993; Wu et al., 2007)。Han等(2005)发现, 中国植物叶片中的磷含量低于世界平均水平, 氮/磷比显著高于世界水平, 这可能说明中国土壤中磷素缺乏状况比世界其他地区更为严重。汪涛等(2008)的研究进一步证实, 中国热带和亚热带地区的土壤全磷含量比其他地区都要低。而古田山24 ha样地土壤中全磷含量平均值只有0.14 g/kg, 更远远低于中亚热带地区土壤全磷含量0.39 g/kg的平均水平(张俪文, 2010)。由于磷素的严重缺乏, 古田山的土壤影响了植物光合作用的某些过程。已有研究表明, 磷胁迫能够通过影响氮元素向Rubisco的分配(Warren & Adam, 2002)、影响类囊体膜上的能量转导(Jacob & Lawlor, 1993)、抑制Calvin循环里几种关键酶的活性等方式(Natr, 1992), 限制植物的叶绿素荧光过程和光合作用(吴楚等, 2004)。此外, 磷素的缺乏也会影响植物对氮素的吸收(Asner et al., 1997; Aber et al., 1998), 因此在本研究中, 叶绿素荧光并未随着磷素的上升而恢复(Carroll et al., 2003; Phoenix et al., 2003)。
图3
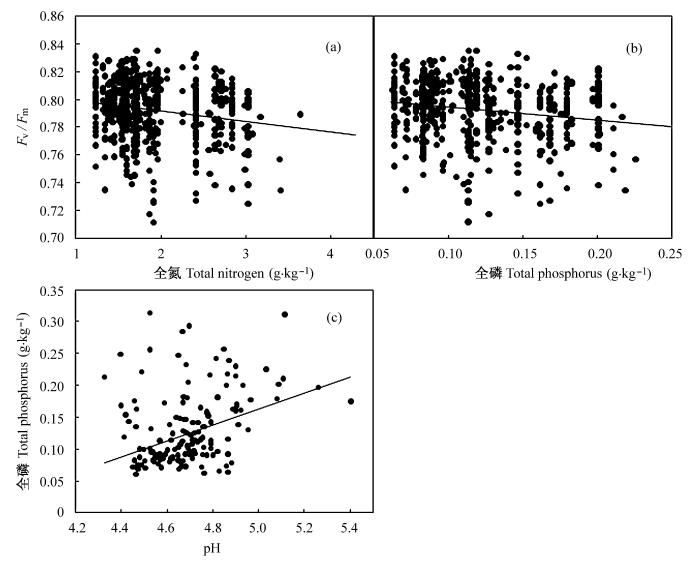
图3
叶绿素荧光参数Fv/Fm 与全氮含量(a)、全磷含量(b)呈负相关(P<0.01); 土壤全磷含量与土壤pH值成正比(P<0.01)。Fv/Fm, 光系统II的最大光化学效率。
Fig. 3
Fluorescence parameter Fv/Fm is negatively correlated with TN (a) and TP (b) (P<0.01); TP and soil pH are positively correlated (P<0.01). Fv/Fm, maximal photochemical efficiency of photosystem II.
磷元素在地壳中的含量虽然十分丰富, 但土壤中绝大多数磷是以无机盐和有机磷的形式存在的,不能被植物直接利用(Bieleski & Ferguson, 1983)。已有研究表明, 土壤中磷素的分布和可利用性首先取决于土壤pH值, 酸性土壤的磷含量通常也较为缺乏(Hinsinger, 2001)。土壤中磷的有效性随pH值的变化呈现出单峰曲线的变化模式, 在pH值为6.5-7.5之间时, 土壤中磷的有效性最佳(刘建玲和张凤华, 2000)。古田山样地中, 土壤pH值同植物叶片中的磷含量呈显著的正比关系(戴文燕, 2010)①(① 戴文燕 (2010) 古田山亚热带常绿阔叶林地上生物量估算与生态化学计量学特征. 硕士学位论文, 中国科学院植物研究所, 北京.)。而在本研究中, 土壤pH值同全磷含量呈现显著的正比关系(图3c, R = 0.487, P< 0.01), 结合古田山土壤pH值的范围(5.5-6.5), 可以预测, 当土壤pH值趋于中性时, 土壤中磷元素的可利用性会增强, 叶绿素荧光也有可能随之恢复。
另一个对植物生长发育有重要作用的环境因子是水分供应。枝条比导率是一个能够较好地反映植物水力结构和木质部水分传导效率的指标。生长在干旱环境下的植物倾向于发育出较窄的导管以适应干旱胁迫(Wright et al., 2004; Edwards, 2006), 这是植物为避免过多水分散失所采取的一种安全性策略。在本研究中, 枝条比导率随着土壤含水量和pH值的上升而上升, 说明在土壤含水量大、并且土壤酸碱度趋于中性的时候, 枝条水分传导效率较高。气孔密度是与植物蒸腾作用有关的一个形态指标, 但在本研究中, 气孔密度与地形和土壤因子的关系并不十分明显。事实上, 关于气孔密度与环境变量之间的关系目前还没有一致的结论(李芳兰等, 2005)。植物叶片气孔的发育受到多种因素的调控, 即使在相似的微环境下, 不同树种对环境变化的响应速度也不同。而古田山地区复杂的地形和气候特征又使得其森林群落内部的微环境更加变化多样。因此尽管在结构上气孔密度与植物水分利用有着天然的关系, 但它在反映地形对植物水分状况的影响方面, 可能并不是一个十分敏感的指标。
植物体会根据所生长和繁殖的环境来改变自己的形态特征。当坡度较大的时候, 植物倾向于长出较厚的叶子来抵御由于冲蚀引起的水分和养分流失。比叶面积(SLA)是反映植物生存策略的重要指示性状, 能够反映植物在不同环境下获取资源的能力。通常来讲, 生活在相对贫瘠的环境中的植物具有较小的SLA, 而将更多的干物质投入用于抵御不良环境(Westoby et al., 2002); 而生活在资源丰富的环境中的植物则具有较大的SLA, 用于获取更多的营养成分(Wilson et al., 1999)。各种环境因子综合作用、共同调控植物的SLA, 这使得SLA的变化更为复杂。此前的一些研究也表明, SLA随海拔的变化并没有统一的规律, 有时会呈现单峰变化的形式, 即在环境综合因子最优的地段, SLA出现峰值(Li et al., 2006)。
随着海拔的升高, 水热条件逐渐成为植物生长发育的限制性因子, 尤其在海拔较高、相对高程差较大的山区, 海拔对植物叶片发育的影响更为明显(李芳兰等, 2005; 祁建等, 2007)。在本研究中, SLA与海拔呈正相关关系, 这可能是由于古田山24 ha样地的相对高程差较小(268.6 m), 水热条件下降不是很明显, 还未成为植物生长的限制因子。而随着海拔升高, 人为干扰减少, 各种环境因子的综合条件有所好转, 加之高海拔地区云雾较多, 植物倾向于发育出大而薄的叶片, 以充分采集光照。植物在对生长和防御做出权衡的时候, 所采取的生存策略是对各种环境因子进行综合后所做出的一种最优选择。
致谢:
感谢中国科学院植物研究所宋凯、浙江省古田山自然保护区方腾、陈声文在野外试验中给予的帮助, 感谢中国科学院植物研究所张俪文博士提供的古田山土壤养分数据。
参考文献
Nitrogen saturation in temperate forest ecosystems: hypotheses revisited
Bryophyte physiological responses to, and recovery from, long-term nitrogen deposition and phosphorus fertilisation in acidic grassland
The decoupling of terrestrial carbon and nitrogen cycles
Vertical gradients in leaf trait diversity in a New Zealand forest
The interactions between plant growth, vegetation structure and soil processes in semi-natural acidic and calcareous grasslands receiving long-term inputs of simulated pollutant nitrogen deposition
Research in large, long term tropical forest plots
DOI:10.1016/s0169-5347(00)88955-7
URL
PMID:21236939
[本文引用: 1]

The past 15 years has seen the creation oflarge (>/16 ha) permanent inventory plots in each of the major tropical forest formations of the world. Currently, six such plots have been fully mapped, and five more and under way. A standardized methodology is used at all sites - a complete census of all trees and saplings down to 1 cm in diameter - thus assuring strict comparability between sites and allowing the development of general models for the dynamics of tropical forests. The inventories aim to gather demographic information on individual tree species, to provide long-term information on forest composition so that future changes can be detected, to estimate the economic value of forest resources, to generate models of sustainable extraction, and to provide data on underused native species for use in reforestation or plantation forestry. The plots also provide data from undisturbed forest to serve as a control for anthropological and management studies of harvested forests.
A handbook of protocols for standardized and easy measurement of plant functional traits worldwide
Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand
Plant distribution and the temperature coefficient of metabolism
Vive la différence: plant functional diversity matters to ecosystem processes
Plant functional traits and environmental filters at a regional scale
Climate change in the Arctic: using plant functional types in a meta-analysis of field experiments
Correlated evolution of stem and leaf hydraulic traits in Pereskia(Cactaceae)
DCCA analysis of plant species distribution in different strata of oak (Quercus liaotungensis) forest along an altitudinal gradient in Dongling Mountain, China
Structural and functional plasticity of Quercus ilex seedlings of different provenances in Italy
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
A test of the generality of leaf trait relationships on the Tibetan Plateau
Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review
Tree species diversity and soil patchiness in temperate broad-leaved forest with limited rooting space
In vivo photosynthetic electron transport does not limit photosynthetic capacity in phosphate-deficient sunflower and maize leaves
Functional traits and niche-based tree community assembly in an Amazonian forest
DOI:10.1126/science.1160662
URL
PMID:18948539
[本文引用: 1]
It is debated whether species-level differences in ecological strategy, which play a key role in much of coexistence theory, are important in structuring highly diverse communities. We examined the co-occurrence patterns of over 1100 tree species in a 25-hectare Amazonian forest plot in relation to field-measured functional traits. Using a null model approach, we show that co-occurring trees are often less ecologically similar than a niche-free (neutral) model predicts. Furthermore, we find evidence for processes that simultaneously drive convergence and divergence in key aspects of plant strategy, suggesting that at least two distinct niche-based processes are occurring. Our results show that strategy differentiation among species contributes to the maintenance of diversity in one of the most diverse tropical forests in the world.
Species Habitat Associations and Species Coexistence in Evergreen Broadleaved Forest in Gutianshan, Zhejiang (古田山常绿阔叶林物种生境关联及其对物种共存的贡献)
Leaf morphological and physiological responses of Quercus aquifolioides along an altitudinal gradient
Leaf characteristics and their Relationship of Cotinus coggygria in arid river valley located in the upper reaches of Minjiang River with environmental factors depending on its altitude gradients
An eco-anatomical study on leaves of Cotinus szechuanensis at gradient elevation in dry valley of the upper Minjiang River
Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study
The progress of phosphorus transformation in soil and its influencing factors
Effects of several environmental factors on plant physiology in Qinghai-Xizang Plateau
Biodiversity and ecosystem functioning: current knowledge and future challenges
DOI:10.1126/science.1064088
URL
PMID:11679658
[本文引用: 1]
The ecological consequences of biodiversity loss have aroused considerable interest and controversy during the past decade. Major advances have been made in describing the relationship between species diversity and ecosystem processes, in identifying functionally important species, and in revealing underlying mechanisms. There is, however, uncertainty as to how results obtained in recent experiments scale up to landscape and regional levels and generalize across ecosystem types and processes. Larger numbers of species are probably needed to reduce temporal variability in ecosystem processes in changing environments. A major future challenge is to determine how biodiversity dynamics, ecosystem processes, and abiotic factors interact.
Leaf traits and associated ecosystem characteristics across subtropical and timberline forests in the Gongga Mountains, Eastern Tibetan Plateau
Rebuilding community ecology from functional traits
DOI:10.1016/j.tree.2006.02.002
URL
PMID:16701083
[本文引用: 1]
There is considerable debate about whether community ecology will ever produce general principles. We suggest here that this can be achieved but that community ecology has lost its way by focusing on pairwise species interactions independent of the environment. We assert that community ecology should return to an emphasis on four themes that are tied together by a two-step process: how the fundamental niche is governed by functional traits within the context of abiotic environmental gradients; and how the interaction between traits and fundamental niches maps onto the realized niche in the context of a biotic interaction milieu. We suggest this approach can create a more quantitative and predictive science that can more readily address issues of global change.
Functional convergence in plant responses to the environment
Plant functional traits, environments and ecosystem functioning
Mineral nutrients: a ubiquitous stress factor for photosynthesis
Effects of enhanced nitrogen deposition and phosphorus limitation on nitrogen budgets of semi-natural grasslands
The altitudinal variation of leaf traits of Quercus liaotungensis and associated environmental explanations
Generality of leaf trait relationships: a test across six biomes
Predicting leaf physiology from simple plant and climate attributes, a global GLOPNET analysis
DOI:10.1890/06-1803.1
URL
PMID:17974336
[本文引用: 1]
Knowledge of leaf chemistry, physiology, and life span is essential for global vegetation modeling, but such data are scarce or lacking for some regions, especially in developing countries. Here we use data from 2021 species at 175 sites around the world from the GLOPNET compilation to show that key physiological traits that are difficult to measure (such as photosynthetic capacity) can be predicted from simple qualitative plant characteristics, climate information, easily measured (
Technique of the modulated chlorophyll fluorescence: basic concepts useful parameters and some applications
The development and patterning of stomata and glands in the epidermis of Peperomia
Chlorophyll fluorescence as a measure of photosynthetic carbon assimilation
A method for measuring hydraulic conductivity and embolism in xylem
DOI:10.1111/pce.1988.11.issue-1 URL [本文引用: 1]
CANOCO reference manual and CanoDraw for Windows user's guide: software for canonical community ordination
Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems
Storage, patterns and environmental controls of soil phosphorus in China
Phosphorus affects growth and partitioning of nitrogen to Rubisco in Pinus pinaster
DOI:10.1093/treephys/22.1.11
URL
PMID:11772551
[本文引用: 1]
We tested the hypothesis that photosynthetic and growth responses to phosphorus (P) are functions of differences in the partitioning of nitrogen (N) among different compounds, particularly ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco). We tested this hypothesis in: (a) a greenhouse experiment with mycorrhizal seedlings of Pinus pinaster Ait. grown in sand culture for 4 months with six factorial combinations of N (0.125 and 2.0 mM) and P (0.02, 0.08 and 0.34 mM); and (b) a field study in which P was applied at five rates (up to 175 kg ha-1) to 2-year-old P. pinaster growing on P-deficient siliceous sand. After 4 months of nutrient addition or 2 years after fertilizer application, we measured light-saturated rates of photosynthesis, growth, and N and P allocation in needles. Growth of P. pinaster increased significantly with increasing concentrations of P, as did the concentration of P in needles. Concentrations of P and Rubisco were positively related, whereas those of N and Rubisco were unrelated. At low-P supply, the Rubisco/Chl ratio varied between 8.5 and 12 mmol mol-1. With P supply in excess of requirement (needle N:P ratio = 2-12) the Rubisco/Chl ratio increased to between 24 and 26 mmol mol-1. Rates of light-saturated photosynthesis were unaffected by P supply because adequate concentrations of P were maintained in plants in all treatments. Orthophosphate accumulated in needles of plants receiving a high P supply, which may allow growth to continue for periods under P deficiency, provided that other nutrients also accumulate. In the case of N, Rubisco may fill this role.
Plant ecological strategies: some leading dimensions of variation between species
Specific leaf area and leaf dry matter content as alternative predictors of plant strategies
Modulation of leaf economic traits and trait relationships by climate
The worldwide leaf economics spectrum
DOI:10.1038/nature02403
URL
PMID:15103368
[本文引用: 1]
Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
Effect of phosphorus stress on chlorophyll biosynthesis, photosynthesis and biomass partitioning pattern of Fraxinus mandchurica seedlings
Phosphorus is essential to forest trees for their growth and development, because of its important role in many metabolic processes of trees. But, most part of phosphorus in soils of Northeastern China is immobile and unavailable, and thus, many forest trees often suffer from phosphorus stress which affects the biomass accumulation of forest trees. The aim of this study is to understand how the chlorophyll biosynthesis, photosynthesis and biomass partitioning of Fraxinus mandchurica seedlings response to phosphorus stress. Sand culture experiment showed that when the seedlings were under phosphorus stress, their 5-aminolevulinic acid (ALA) biosynthesis rate and porphobilinogen (PBG) synthase activity were reduced, but no significant changes in chlorophyll a and b, total chlorophyll and carotenoid contents were found. The nitrogen content in leaves and roots and the phosphorus content in all parts of seedlings were reduced under phosphorus stress, and the reduction of N content in leaves induced the decrease of maximum quantum efficiency of PS II (Fv/Fm) efficiency of PS II photochemistry (phi(PS II)), electron transport rate (E(TR)), and maximum photosynthesis rate (Amax). The effect of phosphorus stress on chlorophyll biosynthesis didn't affect photosynthesis. Under phosphorus stress, the total biomass decreased, but more biomass was partitioned into roots.
Mineral nutrient status of tree species in relation to environmental factors in the subtropical rain forest of Taiwan
Effects of different flush solutions on values of hydraulic conductivities and cavitation resistance abilities of tress of Populus tomentosa and Pinus tabulaeformis
Topography of a large-scale research plot established within a tropical rain forest at Lambir, Sarawak
Effects of water supply on the growth and eco-physiology of seedlings of the dragon spruce Picea asperata Mast
The Effect of Spatial Heterogeneity of Environmental Factors on Species Distribution and Community Structure (环境空间异质性对物种空间分布和群落结构的影响)
A discussion on chlorophyll fluorescence kinetics parameters and their significance
Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, east China
Hydraulic architecture of some diffuse-porous trees
DOI:10.1139/b78-274 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}