


物种多样性由赤道向两极的递减是地球表面最显著的生态格局之一, 对其成因的探讨一直吸引着生态学家和生物地理学家的兴趣(Rosenzweig, 1995; Brown & Lomolino, 1998)。在过去几十年中, 人们提出了上百种关于物种多样性大尺度格局形成机制的假说(Palmer, 1994), 而且新的假说仍在不断地提出(Hubbell, 2001; Brown et al., 2004; Colwell et al., 2004)。其中, 以气候因素为基础的能量假说是目前讨论最多的假说之一。
生物地理学家很早就开始关注能量对物种多样性的可能影响, A. von Humboldt (1769-1859)在其植物地理学的研究中就认为, 能量影响着全球植物多样性的空间格局, 但他只给出了简单的描述, 而没有分析能量对物种多样性影响的机制(见Hawkins et al., 2003a)。在20世纪50年代以后, 随着生态系统研究的深入, 能量对物种多样性的影响受到越来越多生态学家的关注。
早期关于物种多样性能量假说的分析, 起源于对生态系统营养级(trophic level)的研究。比如, Hutchinson(1959)认为, 由于生态系统的食物链长度受其能量输入的控制, 因此能量在不同营养级之间的转化过程可能会控制生态系统内的物种多样性。但第一次使用严格的统计方法定量地分析能量对物种多样性地理格局影响的学者, 当属J. H. Brown及其同事(Brown, 1981; Wright, 1983)。Brown(1981)认为, 能量对物种多样性的影响是通过两个途径实现的, 即能量的多少(capacity rule)以及能量在不同物种及不同个体之间的分配(allocation rule)。之后, Wright (1983)提出, 一个营养级的物种多样性受从低营养级中获得的能量控制, 而植物物种多样性则受太阳辐射能量的控制, 他据此修改了经典的岛屿生物地理学理论, 以能量代替面积, 即S = kEz (其中, S为物种多样性, E为一个地区内某一营养级的能量输入, k、z均为常数)。他研究了全球36个岛屿上的植物和鸟类物种多样性, 发现实际观测数据很好地支持了他的理论(Wright, 1983)。据此, 他首先使用“物种—能量假说(species- energy theory)”这一术语(Wright, 1983), 这被人们认为是能量假说的正式提出。在此后的20多年中, 人们开展了大量关于能量假说的理论和实证研究(Adams & Woodward, 1989; Cousins, 1989; Fraser & Currie, 1996; Guegan et al., 1998; Kaspari et al., 2000; Lennon et al., 2000; Hurlbert, 2004; Storch et al., 2006; Davies et al., 2007; Evans et al., 2008)。
本文基于已有的研究, 综述了影响物种丰富度分布格局的能量形式以及不同能量假说的基本假设、理论预测和已有实证检验, 并在此基础上分析了不同能量假说的可能机制以及将来有待解决的问题。
1 能量的不同形式
能量是生物的重要资源, 能量的多少决定了一个地区对物种的承载能力(capacity) (Brown, 1981)。能量包括不同的形式, 而不同形式的能量对生物具有不同的作用。然而, 很多生态学家在研究中并没有区分不同能量形式之间的差异, 这可能是造成物种—能量假说研究中存在争议的原因之一。
Clarke和Gaston(2006)认为, 能量可以分为三种不同的形式: (1) 辐射能(radiation energy)或光合有效辐射(photosynthetically active radiation, PAR), 指波长介于400-700 nm的可见光, 可被植物用于光合作用; (2) 热能(thermal energy), 指一个地区的冷热气候特征, 通常用与温度相关的指标表征; (3) 吉布斯自由能(Gibbs free energy)或化学能(chemical energy), 指有机物被氧化所释放出来的能量, 通常指好氧生物的呼吸过程(对动物而言指分解代谢或异化作用), 是生物生长和各种生理活动的能量来源。
而Allen等(2007)则根据能量的作用机制, 将能量分为动能(kinetic energy)和势能(potential energy)两种形式。其中动能是指由于运动产生的能量。在生态学研究中, 一般可以区分两种不同的动能:一种是由分子运动产成的热量动能(thermal kinetic energy), 主要影响生物体内的化学反应过程, 通常用绝对温度(K)来衡量(Allen et al., 2002); 另一种则是太阳辐射, 主要被植物用来进行光合作用。因此动能包括了Clarke和Gaston (2006)划分方法的辐射能和热能。而势能则指通过光合作用或同化作用储存在生物体内的化学能, 通常以净初级生产力(net primary production, NPP)或与之相关的指标来衡量, 如年实际蒸散量(actual annual evapotranspiration, AET) (Currie et al., 2004)。
Evans等(2005)同样将能量划分为两种形式, 即太阳辐射能(solar energy matrics)和生产性能量(productive energy matrics)。前者指太阳辐射以及与之相关的其他变量, 比如年均温度和潜在蒸散量, 与Allen等(2007)的“动能”相对应; 后者则指一个地区NPP及与之相关的指标, 与Allen等(2007)的“势能”对应。
这三种对能量形式的区分方式在总体上来说是一致的, 这种区分有利于我们深入理解能量对物种多样性的影响机制。在下文的讨论中, 我们将更多地使用Allen等(2007)的划分(即动能和势能), 因为这种划分更具有物理学意义上的一般性。
一个地区的物种多样性是物种形成速率(speciation)、灭绝速率(extinction)、迁入速率以及迁出速率的直接结果。其中, 前两个过程在较长的时间尺度上起作用, 两者的差值被称为分化速率 (diversification rate); 而后二者则在较短的时间尺度上起作用(Rosenzweig, 1995; Brown & Lomolino, 1998)。在物种多样性分布格局的研究中, 前两者受到了更多的关注, 所有的能量假说均试图建立能量与物种形成/灭绝速率的关系, 并以此解释物种多样性的地理格局。
虽然太阳辐射是植物乃至整个生物圈最根本的能量来源, 但实际上, 植物仅利用了到达地球表面的光合有效辐射的极小部分, 一般认为小于1% (Öpik & Rolfe, 2005)。这说明, 太阳辐射本身可能并不是限制物种多样性的因子。因此, 作为一个单独的因子, 太阳辐射很难解释地球表面物种多样性的地理格局(Clarke & Gaston, 2006)。比如赤道和北极的太阳辐射仅相差4倍, 但两个地区的物种多样性却相差数十倍(Barthlott et al., 1996); 另一个极端的例子是世界屋脊——青藏高原地区, 虽然该地区太阳辐射很强, 且在东部的高寒草甸地区, 水分也不是明显的限制因子, 但物种多样性却很低(方精云等, 2004; 杨元合等, 2004)。由于太阳辐射本身难以解释物种多样性格局, 因而, 关于太阳辐射对物种多样性地理格局直接影响的研究较少, 大多研究集中在热量动能和势能(也即Clarke和Gaston (2006)提出的热能和化学能)对物种多样性格局的影响等方面。根据这两种能量形式对物种多样性的影响机制, 常见的能量假说包括以下五种类型: 生产力假说(productivity hypothesis)、水分—能量动态假说(water-energy dynamic hypothesis)、环境能量假说(ambient energy hypothesis)、寒冷忍耐假说(freezing tolerance hypothesis)以及生态学代谢假说(metabolic theory of ecology, MTE)。
2 生产力假说
生产力假说最早由J. H. Brown及其同事提出 (Brown, 1981; Wright, 1983), 在有些研究中也被称为更多个体假说(more individual hypothesis, MIH) (Gaston, 2000)或直接称为能量假说(Currie et al., 2004)。其主要观点是: 环境能量的增加会提高一个地区的净初级生产力(NPP), 增加生物量的积累, 从而为动物提供更多的食物; 其结果就是使该地区动植物的种群规模(population size)增大, 使更多的个体能够共存, 从而提高了该地区的物种多样性(Brown, 1981; Wright, 1983; Gaston, 2000; Clarke & Gaston, 2006; Evans et al., 2006)。因此, 这一假说中的能量是指一个地区的势能(Allen et al., 2007)或化学能 (Clarke & Gaston, 2006)。在最近的研究中, 能量的这一作用机制被重新表述为: “大蛋糕能被分为更多块(the larger pie can be divided into more pieces) (Fuhrman et al., 2008)”。
在对该假说的检验中, 一个地区的能量通常以NPP来表征(Adams & Woodward, 1989); 但由于NPP难以估算, 有的研究也用与之密切相关的其他指标来衡量, 比如实际蒸散量(AET) (Currie & Paquin, 1987)或干旱地区的年降雨/降水量(annual rainfall/precipitation) (Abramsky & Rosenzweig, 1984)以及归一化植被指数(normalized difference vegetation index, NDVI) (Hurlbert & Haskell, 2003; Evans et al., 2006)等。实际蒸散量反映了一个地区在一年内, 由地表(植被、土壤和水表面)向大气输送的实际水量, 其数值大小受能量和水分的双重控制(Thornthwaite & Hare, 1955), 被认为是表征NPP的最好的单变量气候指标(Lieth & Box, 1972; Lieth, 1975)。归一化植被指数反映了一个地区的植被覆盖程度(或绿度, greenness), 通常与植被生产力具有良好的线性关系(Hurlbert & Haskell, 2003; Evans et al., 2006)。
实测数据和模型模拟研究表明, 在全球尺度上, 群落的NPP由两极向赤道逐渐增加(Gurevitch et al., 2002), 这与物种多样性的纬度梯度具有高度一致性, 在一定程度上支持了生物多样性的生产力假说(Mittelbach et al., 2001; Willig et al., 2003)。比如, Currie和Paquin(1987)对北美洲树木多样性的研究发现, 实际蒸散量是影响其地理格局的主导因子, 解释了其多样性变化的70%以上(图1); 同时, 他们发现, 利用北美洲树木多样性与实际蒸散量的关系能很好地预测爱尔兰和英国的树木多样性。在全球尺度上, 树木多样性与NPP成显著的指数关系, NPP解释了树木多样性变化的84-97%(Adams & Woodward, 1989)。对动物的研究也很好地支持了该假说。比如, Hawkins等(2003a)对全球鸟类多样性的研究发现, 其分布格局主要受年实际蒸散量的影响 (图1)。
图1
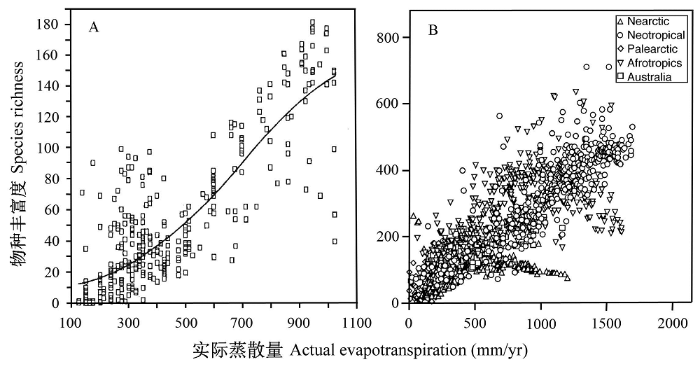
图1
北美洲树木(A, 引自Currie & Paquin, 1987)以及全球鸟类(B, 引自Hawkins et al., 2003a)物种丰富度与实际蒸散量(AET)的关系。其中实际蒸散量反映了一个地区的净初级生产力; Nearctic, 新北区; Neotropical, 新热带区; Palearctic, 古北区; Afrotropics, 古热带区; Australia, 澳洲区。
Fig. 1
Relationships between species richness and actual evapotranspiration (AET) for North American trees (A, from Currie & Paquin, 1987) and global birds (B, from Hawkins et al., 2003a). AET is a surrogate of net primary productivity (NPP).
虽然生产力假说得到了很多实际数据的支持, 但该假说仍面临两个关键问题。首先, 较高的生产力(或生物量)是否会增加一个地区的种群规模?这一问题对不同的类群可能有不同的答案(Gaston, 2000)。研究发现, 脊椎动物, 特别是鸟类和哺乳动物的种群规模呈现明显的纬度梯度, 由赤道向两极逐渐递减(Storch, 2003), 这与陆地表面植被NPP的纬度格局具有一致性。Evans等(2006)对北美洲鸟类种群大小与NDVI关系的研究也发现, 随NDVI增加, 鸟类种群增大。而对于植物, 一些生态学家认为, 森林群落的种群密度(指单位面积的个体总数)并没有明显的纬度梯度(Allen et al., 2002, 2007)。并且, 还有研究显示, 随着单位面积内生物量的提高, 成熟个体的数量并非增加而是减少(Tilman & Pacala, 1993), 这些发现均不符合生产力假说的预测。
第二, 为什么更大的种群(即更多的个体)会分化成更多的物种来分享能量等资源, 而不是形成一个“超级”物种(Wright, 1983; Blackburn & Gaston, 1996; Clarke & Gaston, 2006)。对这个问题, 以往的研究已经提出了多种不同的观点。Brown(1981)和Wright(1983)认为, 较大的种群规模会提高一个地区每个物种的种群大小, 使更多的物种能够抵御环境波动, 从而降低物种的灭绝概率(也见Terborgh & Winter, 1980; Lande, 1993; Srivastava & Lawton, 1998)。还有一些研究认为, 较大的种群规模包含更多的基因多样性, 使不同种群(或不同个体)能适应更多的新环境, 并进而可能在一定程度上增加不同种群之间的隔离程度; 从而提高了该地区的物种形成速率, 最终导致物种多样性的提高(Connell & Orias, 1964; Hubbell, 2001)。其他一些解释则认为, 一个群落的物种多样性与群落内的个体在不同种之间的分配有关。研究发现, 群落内的种群大小呈对数正态分布(Preston, 1962, 1968); 据此, 有些研究认为物种多样性是群落内的个体总数和最小生存种群的函数(Chave, 2004)。
在生产力假说提出之后, 很多生态学家开始研究生物多样性和生产力之间的关系(Guo & Berry, 1998; Weiher, 1999; Tilman et al., 2001; Schmid, 2002; Allcock & Hik, 2003; Hawkins et al., 2003a; Rajaniemi, 2003; Mulder et al., 2004; Hoffmann & Dodson, 2005; Grace et al., 2007; Sanders et al., 2007; Zobel & Pärtel, 2008); 同时, 由于物种多样性的维持受到越来越大的威胁, 而物种多样性的丧失可能会严重危及全球生态系统功能, 也使生态学家对生物多样性与生态系统生产力关系的讨论越来越多。但以往的研究并未发现生物多样性与生产力之间的统一关系(Loreau, 1998, 2000; Loreau et al., 2001)。在实际的观测中, 物种多样性沿生产力梯度的变化趋势大体可以分为四类: 单峰曲线、单调上升曲线、单调下降曲线以及没有显著关系 (Waide et al., 1999; Mittelbach et al., 2001; Hunt et al., 2005)。同时, 物种多样性沿生产力梯度的格局存在明显的尺度效应, 不同的类群之间也具有差异。在局地(local)尺度(比如局地的生物多样性实验)至景观尺度, 植物多样性与生产力多呈单峰曲线关系, 动物多样性与生产力则大多没有显著关系; 而在洲际至全球尺度, 植物和动物多样性与生产力的关系均以单调上升格局最为普遍(Waide et al., 1999)。
在物种多样性—生产力关系的研究中, 单调上升的曲线支持了生产力假说, 但单峰曲线的右半部分(即随生产力增加, 物种多样性下降的部分)以及多样性随生产力单调下降的格局, 并不符合生产力假说的预测, 因而在局地至景观尺度, 物种多样性与生产力的关系可能并不支持生产力假说。比如, Tilman(1982)通过施肥实验发现, 随着生产力的提高, 群落物种丰富度先呈现快速的增加, 但达到一个最大值之后则开始下降, 而非如理论预测继续上升。Abramsky和Rosenzweig(1984)对以色列干旱地区的啮齿类动物多样性的研究同样发现, 物种多样性与生产力(以降雨量作为一个替代指标)呈单峰关系。
对于物种多样性随生产力的提高而逐渐减小的现象, 人们提出了多种解释, 但尚未形成一致结论。比如, D. Tilman等人认为, 生产力提高到一定程度之后, 会导致一个地区生境和资源异质性的降低, 从而导致物种多样性的下降(Tilman, 1982; Tilman & Pacala, 1993); 而Abramsky和Rosenzweig (1984)则认为, 沿着生产力的梯度存在一个干扰梯度, 干扰的减弱使多样性呈现先增大后减小的单峰曲线格局(也见Rosenzweig, 1995); 另外, 物种多样性随生产力的提高而下降的格局也可能与竞争改变有关(Rosenzweig, 1995)。
3 环境能量假说
环境能量假说认为, 物种丰富度的地理格局主要是由能量对物种生理活动的直接控制引起的(Turner, 2004), 其主要代表人物有J. R. G. Turner和D. J. Currie等。这一假说有时也被称为“体温调节假说(thermoregulatory loads hypothesis)”(Lennon et al., 2000)或“分布区限制假说(range limitation hypothesis)”(Evans et al., 2005)。Currie(1991)则把这一假说总结为“良好的环境适合更多的物种生存(benign conditions permit more species)”。在这一假说中, 能量指热量动能(Allen et al., 2007), 通常以年均温、最大潜在蒸散量(potential evapotranspiration, PET)、太阳辐射或日照时数等指标来衡量。
与生产力假说不同, 环境能量假说认为, 能量(指势能或化学能)并非通过在营养级之间的流动影响物种多样性(间接影响), 而是通过直接影响生物个体的生理调节机制而改变物种多样性(Turner et al., 1987, 1988; Currie, 1991; Currie et al., 2004)。一般认为, 在环境能量较高的地区, 变温动物(比如两栖动物和爬行动物)的生理活动更加活跃, 繁殖及养育后代过程中的能量利用效率更高; 而在环境能量较低的地区, 其生理活动所消耗的能量增加, 从而降低繁殖及养育后代过程中的能量分配。与变温动物不同, 恒温动物需要消耗大量的能量维持其体温, 随着环境能量的提高, 这一过程消耗的能量逐渐降低, 从而使得这些物种能分配更多的能量用于繁殖和养育后代; 其结果可能会提高物种的种群规模, 降低物种的灭绝概率, 从而增加物种多样性(Terborgh & Winter, 1980; Brown, 1981; Srivastava & Lawton, 1998)。
环境能量对物种多样性的影响已经得到多个研究的支持。比如, 英国蝴蝶和蛾类的多样性均受夏季(5-9月)平均温度以及夏季日照的影响, 这可能主要是因为较高的环境温度和较强的光照, 会增强蝴蝶的生理活动(Turner et al., 1987)。而Turner等(1988)对英国鸟类的研究则发现, 夏候鸟的多样性随夏季均温升高而增大, 但与冬季均温不相关; 而冬候鸟的多样性则随冬季均温升高而增大, 与夏季温度不相关。这说明, 候鸟只受生活期内的温度影响, 而其他时间的温度对其影响很小; 随着生活期环境温度的提高, 其用于维持体温等过程的能量消耗降低, 这在另一个方面支持了环境能量假说。Currie(1991)对北美洲脊椎动物分布的研究发现, 最大潜在蒸散量(PET)对其多样性格局的解释量远高于实际蒸散量(AET), 他认为, PET主要是通过影响动物调节体温的过程, 进而影响其多样性的变化。这说明, 相对于生产力假说而言, 环境能量假说能更好地解释北美洲脊椎动物的多样性格局。对于伊比利亚半岛爬行动物(Schall & Pianka, 1977)以及北美蜥蜴多样性(Schall & Pianka, 1978)的研究, 也很好地支持了环境能量假说。一些关于植物多样性的研究也同样支持环境能量假说。例如, 南非树木多样性与最冷月的潜在蒸散量(PET)呈显著的抛物线关系, 即随最冷月PET增大, 树木多样性先增大至某一值后再逐渐减小(O’Brien, 1993, 1998; O’Brien et al., 1998; Field et al., 2005); 而温度和PET对全球种子植物科多样性的地理格局具有很大影响, 但其关系受水分的调节(Francis & Currie, 2003; Currie & Francis, 2004)。
虽然环境能量假说得到了大量实际研究支持, 但仍存在一些关键问题尚待解决。实际观测数据显示, 多样性与温度或PET之间的关系并非线性。很多研究发现, 随着环境能量的提高, 多样性逐渐增加, 但在环境能量达到某一个阈值后, 物种多样性也会达到一个上限; 此后, 随能量提高, 多样性或变化很小, 或逐渐下降。比如, 北美洲脊椎动物多样性(Currie, 1991)、南非树木多样性(O’Brien, 1993, 1998; O’Brien et al., 1998; Field et al., 2005)以及全球种子植物科多样性(Francis & Currie, 2003; Currie & Francis, 2004)沿环境能量梯度的变化均呈非线性趋势(图2)。合理解释这一关系, 将增进我们对于物种多样性格局成因的理解。
图2
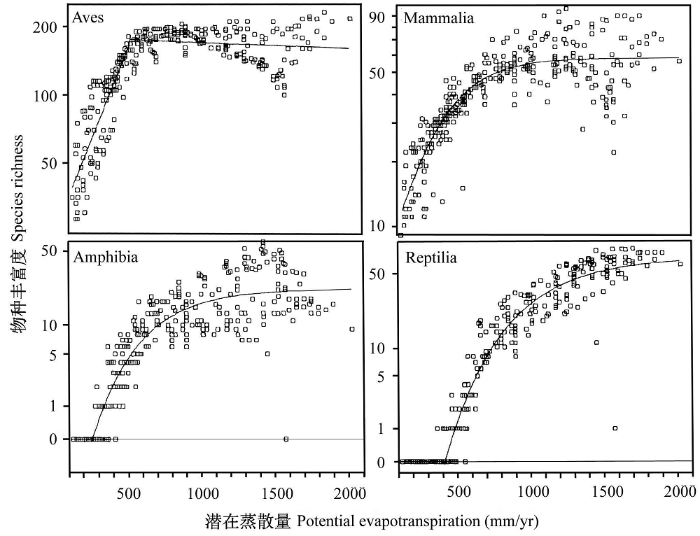
图2
北美洲脊椎动物多样性与环境能量(以最大潜在蒸散表示, PET)的关系(引自Currie, 1991)。Aves:鸟类; Mammalia:哺乳动物; Amphibia:两栖动物; Reptilia:爬行动物。
Fig. 2
Relationships between species richness of North American vertebrates and ambient energy measured by potential evapotranspiration (PET) (from Currie, 1991).
另外, 一些生态学家认为, 虽然温度对物种多样性具有重要影响, 但温度是衡量生物体获得或失去能量的趋势, 并不等同于能量(Huston, 2003; Clarke & Gaston, 2006)。对动物来说, 即使有合适的温度, 如果没有食物他们也无法生存。但值得说明的是, 绝大部分生态学研究的对象均存在于单一介质中, 也即大气(如鸟类、陆生哺乳动物, Jetz & Rahbek, 2002; Hawkins & Porter, 2003; McCain, 2004, 2005, 2007)、水(如鱼类、浮游生物, Macpherson & Duarte, 1994; Zhao et al., 2006; Walker & Cyr, 2007; Reyjol et al., 2007; Fuhrman et al., 2008)或土壤(如土壤动物, Andre et al., 2002)。虽然不同介质之间的比热相差很大, 但其自身的比热在不同地区相差很小, 因此在单独研究某一种介质中的物种多样性梯度时, 温度能在一定程度上反映不同地区热量动能的多少; 但当研究对象跨越不同的介质时, 温度也许不能很好地反映环境热量动能的变化梯度。
4 寒冷忍耐假说
寒冷忍耐假说认为, 在寒冷地区, 很多物种由于不能忍受冬季的寒冷而无法生存, 因此, 随着冬季温度的降低, 其物种多样性逐渐减少(Hawkins, 2001; Hawkins et al., 2003a)。这一假说可能来源于人们对环境“好坏”的直觉。很多人认为, 对于物种的生存来说, 冬季严寒的气候使北方地区的环境变得更加“恶劣”; 与此相反, 南方地区温暖的气候更适合物种的生存(Hutchinson, 1959; Clarke & Gaston, 2006)。这一假说有时也被称为“生理容忍假说(physiological tolerance hypothesis)”(Currie et al., 2004)或“低温限制假说”, 也有些生态学家将这一假说作为环境能量假说的一种。在寒冷忍耐假说中, 能量指热量动能, 通常用一个地区的冬季平均温度、最冷月均温或年极端低温表示。与前两类能量假说相比, 对于这一假说的研究相对较少。
对植物分布的研究为该假说提供了很多证据。如Sakai和Weiser(1973)研究发现, 北美洲部分树木的分布区主要受冬季低温的控制。在东亚地区, Sakai和Malla(1981)认为喜马拉雅山地区物种的垂直分布可能也受冬季低温的影响; Fang和Yoda (1991)指出中国的常绿阔叶林不能分布到平均极端最低温低于-3oC至-2oC的地区。Woodward(1987)总结了不同物种对冬季低温的忍耐性, 发现大部分热带木本植物无法在0oC以下的地区生存; 热带干旱地区落叶植物的生存温度下限介于0-10oC之间; 亚热带和温带地区的常绿阔叶木本植物虽然能忍受较低的温度, 但也无法忍受低于-15oC的冬季温度; 温带落叶阔叶树种能忍受-40oC的冬季低温; 针叶树种的生存温度下限则更低, 可达到-45oC; 一些温带树种或针叶树种甚至不存在低温限制, 因而能够在极端的低温环境下生存, 如杨属(Populus)、桦属(Betula)以及落叶松属(Larix)的某些物种(也见Sakai, 1979; Prentice et al., 1992)。
与此相反, Hawkins等(2003a)发现, 最冷月均温对全球鸟类多样性地理格局的解释量远低于实际蒸散量(AET)。因此他们认为鸟类多样性并不受冬季严寒的影响, 而受能量(指势能或化学能)输入的限制。Turner等(1987)的研究则显示, 英国蛾类多样性与冬季均温呈负相关关系, 即随冬季均温下降, 蛾类多样性逐渐增加, 这一结果也不支持寒冷忍耐假说。这可能是由于蛾在冬季处于冬眠状态, 较高的冬季温度会提高其呼吸速率, 减少其储存的能量, 并最终影响其物种多样性(Turner et al., 1987)。
环境的“恶劣”与否是相对的, 与水生生物进化为陆生生物时所面临的由水体到陆地的环境变化相比, 冬季严寒的恶劣程度可能远低于前者。因此, 这一假说面临的一个问题是: 既然某些物种能适应严寒的气候, 为什么其他物种不能(Hutchinson, 1959)?这一问题质疑了寒冷忍耐假说的一般性, 而这一问题的答案则可能涉及物种的进化历史以及研究的时间尺度。最近的一些研究认为, 在始新世(Ecocene)以前, 地球的气候类似于湿热的热带/亚热带地区, 在早第三纪(Paleogene)晚期, 全球的气候才逐渐变冷(Latham & Ricklefs, 1993; Qian & Ricklefs, 2000); 由于绝大部分物种的祖先在湿热环境中进化, 不具备抵御寒冷的机制, 因而分布受低温的控制(Latham & Ricklefs, 1993; Wiens & Donoghue, 2004; Ricklefs, 2007; Hawkins & DeVries, 2009)。
5 生态学代谢假说
最近, J. H. Brown及其同事提出了生态学代谢理论(也称代谢理论, Gillooly et al., 2001; Allen et al., 2002; Brown et al., 2004), 并尝试利用这一理论从机制上解释物种多样性的地理格局(包括纬度格局和海拔格局)与温度的关系。这种机制性解释主要基于个体新陈代谢速率(metabolic rate, B)与体形大小(M)及绝对温度(T)的关系(Allen et al., 2002; 也见Brown et al., 2004)。人们将这一基于代谢理论的物种多样性假说称为生态学代谢假说(或称代谢假说), 假说中的能量指热量动能, 一般用绝对温度来表示。该理论预测物种丰富度的对数与绝对温度的倒数(1/kT, k为Boltzmann常数)呈线性关系, 其斜率在-0.70至-0.60之间, 即:
ln(S) = C- E×(1/kT)
其中S为物种数量, T为绝对温度, E为生物进行新陈代谢的活化能, k为Boltzmann常数(k = 8.62×10-5eV K-1)。该假说提出之后, 引起了生态学家的广泛关注和争论 (Allen et al., 2003; Brown et al., 2003; Huston, 2003; Storch, 2003; Gillooly & Allen, 2007; Hawkins et al., 2007a, b)。对于生态学代谢理论的详细内容, 我们将另文详细论述(见王志恒等, 2009)。
6 水分–能量动态假说
水分–能量动态假说(有时简称为水热动态假说或水分假说)最早由E. M. O’Brien及其同事们提出, 其主要观点是认为物种多样性的大尺度格局由水分和能量共同决定(O’Brien, 1993; Hawkins et al., 2003b)。这一假说中的能量指热量动能(或热能) (Clarke & Gaston, 2006; Allen et al., 2007), 通常用潜在蒸散量(O’Brien, 1993, 1998; O’Brien et al., 1998)或温度表示; 而水分一般指液态水, 通常用一个地区的年降雨量表示(O’Brien, 1993, 2006)。近年来, 有些研究用实际蒸散量(AET)来衡量一个地区的水热动态(Kreft & Jetz, 2007)。实际蒸散量受能量和水分的共同影响, 在水分充足时(也即有足够的水分用于蒸散), 其大小受能量控制, 而在水分不足时则受水分控制。因此, 实际蒸散量反映了一个地区的水热动态平衡(O’Brien, 1993; Hawkins et al., 2003a)。
水热动态假说主要基于水分对生物体能量利用过程的影响。研究发现, 在植物的生理活动中, 液态水不仅是生物化学过程重要的溶剂, 也是很多生化反应(比如光合作用和呼吸作用等)的重要反应物或产物(Gurevitch et al., 2002; Clarke & Gaston, 2006; O’Brien, 2006)。同时, 液态水的运动是植物吸收和运输营养物质的动力(Gurevitch et al., 2002); 这些过程均对植物的能量利用过程(比如光合作用和呼吸作用)具有重要的影响。水在不同的能量条件下呈现不同的状态, 当能量太低时(温度0oC以下), 水以冰的形式存在; 当能量太高时(温度100oC以上), 水变为水蒸汽。以这两种状态存在的水分无法作为溶剂和反应物参与植物的能量利用过程。只有当水分以液态的形式存在时, 才能影响植物的能量利用过程。因此, O’Brien等人认为, 由于水分的作用, 能量对物种多样性的影响呈现抛物线形式, 即随能量升高, 多样性先升高后下降(O’Brien, 1993, 1998, 2006; O’Brien et al., 1998), 因而水分和能量的多少共同决定了植物光合作用的强弱以及生物量的积累, 并进而影响植物多样性格局(O’Brien, 1993; Gurevitch et al., 2002)。比如, 在沙漠地区, 虽然能量充足, 但由于水分的限制, 植物无法利用这些能量进行光合作用(O’Brien, 2006), 从而解释了为什么沙漠地区拥有极低的物种多样性。
早期, 这一假说主要被用来解释维管束植物, 特别是木本植物的多样性分布格局。比如, E. M. O’Brien及其同事利用这一假说解释南非树木物种丰富度的地理格局, 发现水热动态(降雨量和最冷月蒸散量)解释了南非树木多样性变化的79%(O’Brien, 1993; O’Brien et al., 1998)。她们利用南非树木数据建立了水热动态模型, 并用该模型预测了南、北美洲和中国木本植物的分布格局(O’Brien, 1998; O’Brien et al., 1998; Field et al., 2005)。但需要说明的是, 虽然这一模型解释了南非树木多样性格局, 但模型中的最冷月蒸散量在北温带大部分地区均为0, 因而无法反映实际的能量输入, 因此Hawkins等人建议使用其他的指标(比如最冷月均温等)代替最冷月蒸散量(Hawkins et al., 2007)。对北美、欧洲树木多样性以及全球种子植物科多样性的研究也很好地支持了这一假说(Francis & Currie, 2003; Currie & Francis, 2004; Hawkins et al., 2007c; Montoya et al., 2007)。如Francis和Currie等人对全球种子植物科多样性地理格局的分析显示, 水分亏缺(water deficit, WD, 反映了一个地区的干旱程度)和潜在蒸散量共同决定了种子植物的多样性格局, 且多样性与能量(以温度或潜在蒸散量表示)之间呈抛物线关系, 从而支持了这一假说 (Francis & Currie, 2003; Currie & Francis, 2004)。在最近的研究中, Hawkins等(2003a, 2005)用这一假说解释了鸟类多样性的分布格局, 但他们认为, 水热动态对动物的影响可能是间接的, 是通过对植物生产力和生物量的影响实现的。
虽然水分和能量均对物种多样性具有显著的影响, 但二者的相对重要性在不同地区具有很大差异。Hawkins等(2003b)对85个关于能量和水分效应的研究发现, 在能量较低的北方地区, 能量是限制物种多样性的主导因子; 而在南方地区, 能量并 不对物种的生理活动构成限制, 因而不是多样性 格局的主导因子, 相反, 水分则成为主导因子。Whittaker等(2007)对欧洲鸟类和蝴蝶多样性的研究很好地支持了这一结论。
7 小结
我们对5种不同的能量假说进行了简单的描述。在已有的研究中, 每一种假说都在不同程度上得到了实际观测数据的支持; 同时, 每种假说都存在自身的优点以及有待解决的问题。因此, 对于不同假说的优劣, 目前尚未形成一致结论。已有研究发现, 对于不同的生物类群, 主导其物种多样性地理格局的能量形式以及能量对多样性的影响机制可能是不同的, 因而不同假说的适用性可能也是不同的。Gaston(2000)认为, 植物可能受能量和水分的共同影响; 而动物则主要受能量的影响, 水分的作用较小。Allen等(2007)认为, 物种多样性的地理格局是由动能和势能共同决定的, 但二者通过两个不同的机制影响多样性格局; 同时, 对不同的类群, 二者的相对重要性也有很大差异(图3)。由于恒温动物的体温在不同地区基本保持不变, 其代谢速率不随环境温度而变化, 从而无法用生态学代谢理论解释其物种多样性格局; 而较高的生产力为恒温动物提供了更多的食物资源, 从而会提高种群规模, 因而其多样性格局可能主要受势能, 即不同地区的生产力控制。对变温动物和植物而言, 其新陈代谢速率随环境温度升高而升高, 直接结果就是缩短物种的世代时间和提高突变速率, 进而提高物种形成速率和物种多样性, 因此, 热量动能可能主导了变温动物和植物的多样性格局。基于此, 构建包含动能和势能的统一模型, 研究二者如何共同影响物种多样性, 将有助于我们理解物种多样性格局的成因。
图3
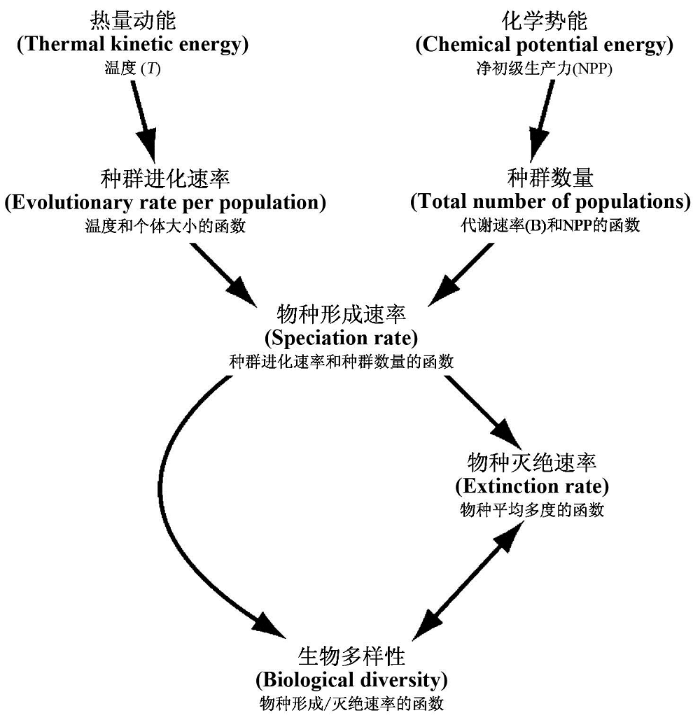
图3
热量动能和势能对物种多样性的影响机制(引自Allen et al., 2007)
Fig. 3
Collective mechanisms for influences of thermal kinetic energy and potential energy on species diversity (from Allen et al., 2007).
参考文献
Tilman's predicted productivity-diversity relationship shown by desert rodents
DOI:10.1038/309150a0
URL
PMID:6717592
[本文引用: 3]

Tilman has developed a model to predict the number of plant species that can coexist competitively on a limited resource base. Species diversity first increases over low resource supplies, then declines as the environment becomes richer. Although Tilman 's model was developed to describe interspecific interactions between plant species, it may also apply to animal species. Tilman questions whether animals specialize on particular proportions of nutrients. However, we believe animals probably specialize on relatively subtle microhabitat differences, especially in a multispecies competitive regime. Thus, microhabitats may act like nutrients. We hypothesize that animal species, too, show a peaked curve of diversity over productivity. The present data provide a confirmation of the hypothesis using rodent species. We have investigated the number of rodent species along a geographical gradient of increasing rainfall. The gradient extends from extremely poor desert habitats to those with annual rainfall over 300 mm. Because of the aridity , precipitation reflects productivity. The diversity pattern in desert rodents agrees with that predicted by Tilman for plants. It even possesses similar asymmetry, rising steeply then falling slowly. The pattern is duplicated in rocky and sandy habitats, each of which has a distinct and almost nonoverlapping assemblage of species. As mean precipitation is closely correlated with the variability of precipitation, the diversity pattern might also be caused by a decline in the frequency of disturbances, models for which have been proposed by several investigators.
Patterns in tree species richness as a test of the glacial extinction hypothesis
What determines disturbance-productivity-diversity relationships? The effect of scale, species and environment on richness patterns in an Australian woodland
Global biodiversity, biochemical kinetics, and the energetic-equivalence rule
Response to comment on “global biodiversity, biochemical kinetics, and the energetic-equivalence rule”
DOI:10.1126/science.1078627 URL PMID:12531999 [本文引用: 1]
Soil biodiversity: myth, reality or conning?
Global distribution of species diversity in vascular plants: towards a world map of phytodiversity
A sideways look at patterns in species richness, or why there are so few species outside the tropics
Two decades of homage to Santa Rosalia: toward a general theory of diversity
Response: heat and biodiversity
DOI:10.1126/science.299.5606.512 URL PMID:12546005 [本文引用: 1]
Toward a metabolic theory of ecology
Climate, energy and diversity
URL PMID:16928626 [本文引用: 11]
The mid-domain effect and species richness patterns: what have we learned so far?
DOI:10.1086/382056
URL
PMID:15026983
[本文引用: 1]
If species' ranges are randomly shuffled within a bounded geographical domain free of environmental gradients, ranges overlap increasingly toward the center of the domain, creating a
The ecological regulation of species diversity
Regional versus climatic effect on taxon richness in angiosperms: reply to Qian and Ricklefs
Large-scale biogeographical patterns of species richness of trees
Energy and large-scale patterns of animal- and plant-species richness
Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness
Topography, energy and the global distribution of bird species richness
DOI:10.1098/rspb.2006.0061
URL
PMID:17311781
[本文引用: 1]
A major goal of ecology is to determine the causes of the latitudinal gradient in global distribution of species richness. Current evidence points to either energy availability or habitat heterogeneity as the most likely environmental drivers in terrestrial systems, but their relative importance is controversial in the absence of analyses of global (rather than continental or regional) extent. Here we use data on the global distribution of extant continental and continental island bird species to test the explanatory power of energy availability and habitat heterogeneity while simultaneously addressing issues of spatial resolution, spatial autocorrelation, geometric constraints upon species' range dynamics, and the impact of human populations and historical glacial ice-cover. At the finest resolution (1 degree), topographical variability and temperature are identified as the most important global predictors of avian species richness in multi-predictor models. Topographical variability is most important in single-predictor models, followed by productive energy. Adjusting for null expectations based on geometric constraints on species richness improves overall model fit but has negligible impact on tests of environmental predictors. Conclusions concerning the relative importance of environmental predictors of species richness cannot be extrapolated from one biogeographic realm to others or the globe. Rather a global perspective confirms the primary importance of mountain ranges in high-energy areas.
Abundance, species richness and energy availability in the North American avifauna
Spatial scale, abundance and the species-energy relationship in British birds
Species-energy relationships at the macroecological scale: a review of the mechanisms
DOI:10.1017/s1464793104006517
URL
PMID:15727036
[本文引用: 2]
Correlations between the amount of energy received by an assemblage and the number of species that it contains are very general, and at the macro-scale such species-energy relationships typically follow a monotonically increasing curve. Whilst the ecological literature contains frequent reports of such relationships, debate on their causal mechanisms is limited and typically focuses on the role of energy availability in controlling the number of individuals in an assemblage. Assemblages from high-energy areas may contain more individuals enabling species to maintain larger, more viable populations, whose lower extinction risk elevates species richness. Other mechanisms have, however, also been suggested. Here we identify and clarify nine principal mechanisms that may generate positive species-energy relationships at the macro-scale. We critically assess their assumptions and applicability over a range of spatial scales, derive predictions for each and assess the evidence that supports or refutes them. Our synthesis demonstrates that all mechanisms share at least one of their predictions with an alternative mechanism. Some previous studies of species-energy relationships appear not to have recognised the extent of shared predictions, and this may detract from their contribution to the debate on causal mechanisms. The combination of predictions and assumptions made by each mechanism is, however, unique, suggesting that, in principle, conclusive tests are possible. Sufficient testing of all mechanisms has yet to be conducted, and no single mechanism currently has unequivocal support. Each may contribute to species-energy relationships in some circumstances, but some mechanisms are unlikely to act simultaneously. Moreover, a limited number appear particularly likely to contribute frequently to species-energy relationships at the macro-scale. The increased population size, niche position and diversification rate mechanisms are particularly noteworthy in this context.
Community structure of alpine sparse vegetation and effects of microtopography in Pushila, Everest-Choyu region, Tibet, China
Climate and vegetation of China V: effects of climate factors on the upper limit of distribution of evergreen broadleaf forest
Global models for predicting woody plant richness from climate: development and evaluation
A globally consistent richness-climate relationship for angiosperms
DOI:10.1086/368223
URL
PMID:12776882
[本文引用: 4]
Species richness, the simplest index of biodiversity, varies greatly over broad spatial scales. Richness-climate relationships often account for >80% of the spatial variance in richness. However, it has been suggested that richness-climate relationships differ significantly among geographic regions and that there is no globally consistent relationship. This study investigated the global patterns of species and family richness of angiosperms in relation to climate. We found that models relating angiosperm richness to mean annual temperature, annual water deficit, and their interaction or models relating richness to annual potential evapotranspiration and water deficit are both globally consistent and very strong and are independent of the diverse evolutionary histories and functional assemblages of plants in different parts of the world. Thus, effects of other factors such as evolutionary history, postglacial dispersal, soil nutrients, topography, or other climatic variables either must be quite minor over broad scales (because there is little residual variation left to explain) or they must be strongly collinear with global patterns of climate. The correlations shown here must be predicted by any successful hypothesis of mechanisms controlling richness patterns.
The species richness-energy hypothesis in a system where historical factors are thought to prevail: coral reefs
A latitudinal diversity gradient in planktonic marine bacteria
Global patterns in biodiversity
DOI:10.1038/35012228
URL
PMID:10821282
[本文引用: 4]
To a first approximation, the distribution of biodiversity across the Earth can be described in terms of a relatively small number of broad-scale spatial patterns. Although these patterns are increasingly well documented, understanding why they exist constitutes one of the most significant intellectual challenges to ecologists and biogeographers. Theory is, however, developing rapidly, improving in its internal consistency, and more readily subjected to empirical challenge.
Linking global patterns in biodiversity to evolutionary dynamics using metabolic theory
DOI:10.1890/06-1935.1 URL PMID:17824416 [本文引用: 1]
Effects of size and temperature on metabolic rate
DOI:10.1126/science.1061967
URL
PMID:11567137
[本文引用: 1]
We derive a general model, based on principles of biochemical kinetics and allometry, that characterizes the effects of temperature and body mass on metabolic rate. The model fits metabolic rates of microbes, ectotherms, endotherms (including those in hibernation), and plants in temperatures ranging from 0 degrees to 40 degrees C. Mass- and temperature-compensated resting metabolic rates of all organisms are similar: The lowest (for unicellular organisms and plants) is separated from the highest (for endothermic vertebrates) by a factor of about 20. Temperature and body size are primary determinants of biological time and ecological roles.
Does species diversity limit productivity in natural grassland communities?
DOI:10.1111/j.1461-0248.2007.01058.x
URL
PMID:17594423
[本文引用: 1]
Theoretical analyses and experimental studies of synthesized assemblages indicate that under particular circumstances species diversity can enhance community productivity through niche complementarity. It remains unclear whether this process has important effects in mature natural ecosystems where competitive feedbacks and complex environmental influences affect diversity-productivity relationships. In this study, we evaluated diversity-productivity relationships while statistically controlling for environmental influences in 12 natural grassland ecosystems. Because diversity-productivity relationships are conspicuously nonlinear, we developed a nonlinear structural equation modeling (SEM) methodology to separate the effects of diversity on productivity from the effects of productivity on diversity. Meta-analysis was used to summarize the SEM findings across studies. While competitive effects were readily detected, enhancement of production by diversity was not. These results suggest that the influence of small-scale diversity on productivity in mature natural systems is a weak force, both in absolute terms and relative to the effects of other controls on productivity.
Energy availability and habitat heterogeneity predict global riverine fish diversity
Species richness and biomass: dessection of the hump-shaped relationships
The Ecology of Plants
A global evaluation of metabolic theory as an explanation for terrestrial species richness gradients
DOI:10.1890/06-1444.1
URL
PMID:17824415
[本文引用: 3]
We compiled 46 broadscale data sets of species richness for a wide range of terrestrial plant, invertebrate, and ectothermic vertebrate groups in all parts of the world to test the ability of metabolic theory to account for observed diversity gradients. The theory makes two related predictions: (1) In-transformed richness is linearly associated with a linear, inverse transformation of annual temperature, and (2) the slope of the relationship is near -0.65. Of the 46 data sets, 14 had no significant relationship; of the remaining 32, nine were linear, meeting prediction 1. Model I (ordinary least squares, OLS) and model II (reduced major axis, RMA) regressions then tested the linear slopes against prediction 2. In the 23 data sets having nonlinear relationships between richness and temperature, split-line regression divided the data into linear components, and regressions were done on each component to test prediction 2 for subsets of the data. Of the 46 data sets analyzed in their entirety using OLS regression, one was consistent with metabolic theory (meeting both predictions), and one was possibly consistent. Using RMA regression, no data sets were consistent. Of 67 analyses of prediction 2 using OLS regression on all linear data sets and subsets, two were consistent with the prediction, and four were possibly consistent. Using RMA regression, one was consistent (albeit weakly), and four were possibly consistent. We also found that the relationship between richness and temperature is both taxonomically and geographically conditional, and there is no evidence for a universal response of diversity to temperature. Meta-analyses confirmed significant heterogeneity in slopes among data sets, and the combined slopes across studies were significantly lower than the range of slopes predicted by metabolic theory based on both OLS and RMA regressions. We conclude that metabolic theory, as currently formulated, is a poor predictor of observed diversity gradients in most terrestrial systems.
Tropical niche conservatism and the species richness gradient of North American butterflies
Metabolic theory and diversity gradients: where do we go from here?
DOI:10.1890/06-2141.1 URL PMID:17824418 [本文引用: 1]
Water links the historical and contemporary components of the Australian bird diversity gradient
Energy, water, and broad-scale geographic patterns of species richness
Global models for predicting woody plant richness from climate: comment
DOI:10.1890/0012-9658(2007)88[255:gmfpwp]2.0.co;2 URL PMID:17489474
Relative influences of current and historical factors on mammal and bird diversity patterns in deglaciated North America
Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds
Land use, primary productivity, and lake area as descriptors of zooplankton diversity
Species-energy relationship in the deep sea: a test using the quaternary fossil record
The effect of energy and seasonality on avian species richness and community composition
DOI:10.1086/345459
URL
PMID:12650464
[本文引用: 2]
We analyzed geographic patterns of richness in both the breeding and winter season in relation to a remotely sensed index of seasonal production (normalized difference vegetation index [NDVI]) and to measures of habitat heterogeneity at four different spatial resolutions. The relationship between avian richness and NDVI was consistent between seasons, suggesting that the way in which available energy is converted to bird species is similar at these ecologically distinct times of year. The number and proportion of migrant species in breeding communities also increased predictably with the degree of seasonality. The NDVI was a much better predictor of seasonal richness at finer spatial scales, whereas habitat heterogeneity best predicted richness at coarser spatial resolutions. While we find strong support for a positive relationship between available energy and species richness, seasonal NDVI explained at most 61% of the variation in richness. Seasonal NDVI and habitat heterogeneity together explain up to 69% of the variation in richness.
Species-energy relationships and habitat complexity in bird communities
Heat and biodiversity
DOI:10.1126/science.299.5606.512 URL PMID:12546005 [本文引用: 2]
Homage to Santa Rosalia or why are there so many kinds of animals?
Geographic range size and determinants of avian species richness
DOI:10.1126/science.1072779
URL
PMID:12202829
[本文引用: 2]
Geographic patterns in species richness are mainly based on wide-ranging species because their larger number of distribution records has a disproportionate contribution to the species richness counts. Here we demonstrate how this effect strongly influences our understanding of what determines species richness. Using both conventional and spatial regression models, we show that for sub-Saharan African birds, the apparent role of productivity diminishes with decreasing range size, whereas the significance of topographic heterogeneity increases. The relative importance of geometric constraints from the continental edge is moderate. Our findings highlight the failure of traditional species richness models to account for narrow-ranging species that frequently are also threatened.
Energy, density, and constraints to species richness: ant assemblages along a productivity gradient
DOI:10.1086/303313
URL
PMID:10686166
[本文引用: 1]
Global patterns and determinants of vascular plant diversity
Risks of population extinction from demographic and environmental stochasticity and random catastrophes
DOI:10.1086/285580
URL
PMID:29519140
[本文引用: 1]
Stochastic factors affecting the demography of a single population are analyzed to determine the relative risks of extinction from demographic stochasticity, environmental stochasticity, and random catastrophes. Relative risks are assessed by comparing asymptotic scaling relationships describing how the average time to extinction, T, increases with the carrying capacity of a population, K, under each stochastic factor alone. Stochastic factors are added to a simple model of exponential growth up to K. A critical parameter affecting the extinction dynamics is $$\tilde r,$$ the long-run growth rate of a population below K, including stochastic factors. If r is positive, with demographic stochasticity T increases asymptotically as a nearly exponential function of K, and with either environmental stochasticity or random catastrophes T increases asymptotically as a power of K. If r is negative, under any stochastic demographic factor, T increases asymptotically with the logarithm of K. Thus, for sufficiently large populations, the risk of extinction from demographic stochasticity is less important than that from either environmental stochasticity or random catastrophes. The relative risks of extinction from environmental stochasticity and random catastrophes depend on the mean and environmental variance of population growth rate, and the magnitude and frequency of catastrophes. Contrary to previous assertions in the literature, a population of modest size subject to environmental stochasticity or random catastrophes can persist for a long time, if r is substantially positive.
Global patterns of tree species richness in moist forests: energy-diversity theory does not account for variation in species richness
Bird diversity and environmental gradients in Britain: a test of the species-energy hypothesis
Evapotranspiration and primary productivity: CW Thornthwaite memorial model
Modeling the primary productivity of the world
Biodiversity and ecosystem functioning: a mechanistic model
Biodiversity and ecosystem functioning: recent theoretical advances
Biodiversity and ecosystem functioning: current knowledge and future challenges
DOI:10.1126/science.1064088
URL
PMID:11679658
[本文引用: 1]
The ecological consequences of biodiversity loss have aroused considerable interest and controversy during the past decade. Major advances have been made in describing the relationship between species diversity and ecosystem processes, in identifying functionally important species, and in revealing underlying mechanisms. There is, however, uncertainty as to how results obtained in recent experiments scale up to landscape and regional levels and generalize across ecosystem types and processes. Larger numbers of species are probably needed to reduce temporal variability in ecosystem processes in changing environments. A major future challenge is to determine how biodiversity dynamics, ecosystem processes, and abiotic factors interact.
Patterns in species richness, size, and latitudinal range of East Atlantic fishes
The mid-domain effect applied to elevational gradients: species richness of small mammals in Costa Rica
Area and mammalian elevational diversity
DOI:10.1890/0012-9658(2007)88[76:aamed]2.0.co;2
URL
PMID:17489456
Elevational gradients hold enormous potential for understanding general properties of biodiversity. Like latitudinal gradients, the hypotheses for diversity patterns can be grouped into historical explanations, climatic drivers, and spatial hypotheses. The spatial hypotheses include the species-area effect and spatial constraint (mid-domain effect null models). I test these two spatial hypotheses using regional diversity patterns for mammals (non-volant small mammals and bats) along 34 elevational gradients spanning 24.4 degrees S-40.4 degrees N latitude. There was high variability in the fit to the species-area hypothesis and the mid-domain effect. Both hypotheses can be eliminated as primary drivers of elevational diversity. Area and spatial constraint both represent sources of error rather than mechanisms underlying these mammalian diversity patterns. Similar results are expected for other vertebrate taxa, plants, and invertebrates since they show comparable distributions of elevational diversity patterns to mammalian patterns.
What is the observed relationship between species richness and productivity?
Contemporary richness of Holarctic trees and the historical pattern of glacial retreat
Species evenness and productivity in experimental plant communities
Climatic gradients in woody plant species richness: towards an explanation based on an analysis of southern Africa’s woody flora
Water-energy dynamics, climate, and prediction of woody plant species richness: an interim general model
Biological relativity to water-energy dynamics
Climate and woody plant diversity in southern Africa: relationships at species, genus and family levels
Variation in species richness: towards a unification of hypotheses
A global biome model based on plant physiology and dominance, soil properties and climate
The canonical distribution of commonness and rarity, Part I
Large-scale processes and the Asian bias in species diversity of temperate plants
DOI:10.1038/35025052
URL
PMID:11001054
[本文引用: 1]
An important issue in the study of biodiversity is the extent to which global patterns of species richness reflect large-scale processes and historical contingencies. Ecological interactions in local assemblages may constrain the number of species that can coexist, but differences in diversity in similar habitats within different regions (diversity anomalies) suggest that this limit is not firm. Variation in rate of species production could influence regional and perhaps local diversity independently of the ecological capacity of an area to support coexisting species, thereby creating diversity anomalies. Temperate Zone genera of plants that are disjunct between similar environments in eastern Asia and eastern North America (EAS-ENA) have twice as many species in Asia as in North America. Because lineages of these genera in Asia and North America are mostly sister pairs, they share a common history of adaptation and ecological relationship before disjunction. Thus, the diversity anomaly in EAS-ENA genera is not an artefact of taxon or habitat sampling but reflects differences in the net diversification (speciation-extinction) of the lineages in each of the continents. Here we propose that the most probable cause of the EAS-ENA anomaly in diversity is the extreme physiographical heterogeneity of temperate eastern Asia, especially compared with eastern North America, which in conjunction with climate and sea-level change has provided abundant opportunities for evolutionary radiation through allopatric speciation.
Explaining productivity-diversity relationships in plants
Patterns in species richness and endemism of European freshwater fish
History and diversity: explorations at the intersection of ecology and evolution
DOI:10.1086/519402
URL
PMID:17874385
[本文引用: 1]
Phylogenetic analysis provides an important tool for assessing the influence of historical and evolutionary processes on the structure of contemporary ecological systems. Patterns of diversity, for example, represent the regional buildup of species through immigration and diversification, their loss through extinction, and the sorting of species ecologically within the region. Colonization-extinction dynamics on islands can be inferred from lineage accumulation through time. Lineage branching within clades can be used to estimate rates of speciation and extinction. However, simulations of these processes show potential ambiguities in the interpretation of data. Clade size is unrelated to age in many studies, suggesting that speciation and extinction might be in long-term equilibrium and raising questions about unobserved past diversity. Among passerine birds and other groups, the size of similar-aged clades is positively related to the size of the region within which they have diversified, and it is greater in tropical than in temperate regions. There is no consensus on the causes of these patterns. Finally, the ecological interactions between populations within regions brings the timescale of species sorting and species production close to each other and emphasizes the important interaction of ecological and evolutionary processes in shaping ecological systems.
Winter hardiness of tree species at high altitudes in the east Himalaya, Nepal
Freezing resistance of trees in North America with reference to tree regions
Freezing avoidance mechanism of primordial shoots of conifer buds
Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains
Species densities of reptiles and amphibians on the Iberian Peninsuls
Geographical trends in numbers of species
DOI:10.1126/science.201.4357.679
URL
PMID:17750221
[本文引用: 1]
Geographic variation in the number of coexisting plant and animal species (species density) often follows repeated patterns; best known is the general increase in species richness from temperate to tropical latitudes. Here we undertake a quantitative analysis of geographic trends in species density for the terrestrial vertebrate faunas of the United States and Australia. Trends in numbers of species of amphibians, reptiles, birds, and mammals are described and are correlated with geographic variation in abiotic environmental measures. Intercontinental comparisons reveal general patterns as well as intriguing and profound differences in vertebrate distributions.
The species richness-productivity controversy
Why more productive sites have more species: an experimental test of theory using tree-hole communities
Comment on “global biodiversity, biochemical kinetics, and the energetic-equivalence rule”
Energy, range dynamics and global species richness patterns: reconciling mid-domain effects and environmental determinants of avian diversity
DOI:10.1111/j.1461-0248.2006.00984.x
URL
PMID:17118005
[本文引用: 1]
Spatial patterns of species richness follow climatic and environmental variation, but could reflect random dynamics of species ranges (the mid-domain effect, MDE). Using data on the global distribution of birds, we compared predictions based on energy availability (actual evapotranspiration, AET, the best single correlate of avian richness) with those of range dynamics models. MDE operating within the global terrestrial area provides a poor prediction of richness variation, but if it operates separately within traditional biogeographic realms, it explains more global variation in richness than AET. The best predictions, however, are given by a model of global range dynamics modulated by AET, such that the probability of a range spreading into an area is proportional to its AET. This model also accurately predicts the latitudinal variation in species richness and variation of species richness both within and between realms, thus representing a compelling mechanism for the major trends in global biodiversity.
Diversity and productivity in a long-term grassland experiment
DOI:10.1126/science.1060391
URL
PMID:11679667
[本文引用: 1]
Plant diversity and niche complementarity had progressively stronger effects on ecosystem functioning during a 7-year experiment, with 16-species plots attaining 2.7 times greater biomass than monocultures. Diversity effects were neither transients nor explained solely by a few productive or unviable species. Rather, many higher-diversity plots outperformed the best monoculture. These results help resolve debate over biodiversity and ecosystem functioning, show effects at higher than expected diversity levels, and demonstrate, for these ecosystems, that even the best-chosen monocultures cannot achieve greater productivity or carbon stores than higher-diversity sites.
Explaining the global biodiversity gradient: energy, area, history and natural selection
Does solar energy control organic diversity? Butterflies, moths and the British climate
British bird species distributions and the energy theory
Testing the standard neutral model of biodiversity in lake communities
Metabolic theory of ecology: an explanation for species richness patterns based on the metabolic processes of organisms
The relationship between productivity and species richness
The combined effects of scale and productivity on species richness
Geographical gradients of species richness: a test of the water-energy conjecture of Hawkins et al. (2003) using European data for five taxa
Historical biogeography, ecology, and species richness
DOI:10.1016/j.tree.2004.09.011
URL
PMID:16701326
[本文引用: 1]
Ecology and historical (phylogeny-based) biogeography have much to offer one another, but exchanges between these fields have been limited. Historical biogeography has become narrowly focused on using phylogenies to discover the history of geological connections among regions. Conversely, ecologists often ignore historical biogeography, even when its input can be crucial. Both historical biogeographers and ecologists have more-or-less abandoned attempts to understand the processes that determine the large-scale distribution of clades. Here, we describe the chasm that has developed between ecology and historical biogeography, some of the important questions that have fallen into it and how it might be bridged. To illustrate the benefits of an integrated approach, we expand on a model that can help explain the latitudinal gradient of species richness.
Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis
Species-energy theory: an extension of species-area theory
Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau
Patterns of fish species richness in China’s lakes
What determines the relationship between plant diversity and habitat productivity?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}