


一个多世纪以来, 生物多样性的形成和维持机理即群落构建(community assembly)机制一直是生态学研究的核心论题。传统的观点认为, 各物种特征的权衡(trade-offs)和组合的不同决定了其生活史对策(life historical strategy, 如资源利用方式)的不同, 由此决定了各物种在群落中所占有的生态位(niche)不同, 进而决定了多物种的稳定共存(Hutchinson, 1959; Vandermeer, 1972; Silvertown, 2004)。也就是说, 物种在群落中的共存是以生态位的分化为前提的, 生态位相同的物种可能因竞争共同的资源而发生竞争排除, 不能稳定共存(Vandermeer, 1972)。然而, 当解释生境中限制性资源不多, 多物种生态位分化不明显, 而依然稳定同存的现象(如热带雨林)时, 传统的生态位构建理论遇到了巨大的挑战(Bell, 2000; Hubbell, 2001)。并且, 以生态位理论为基础的确定性因素无法有效地预测群落结构动态, 使得许多生态学家开始考虑非确定性因子的作用。
以Hubbell为代表的生态学家根据种群遗传学的中性理论(neutral theory), 提出了群落中性理论(Bell, 2000; Hubbell, 2001)。群落中性理论以其简约性和可预测能力迅速赢得许多生态学家的青睐, 成为过去10多年来最受关注的生态学理论热点(Volkov et al., 2003; Chave, 2004; Alonso et al., 2006; Holt, 2006)。一些生态学家甚至认为, 群落中性理论是将生物多样性和生物地理学相统一的理论(Hubbell, 2001; Bell, 2003)。然而, 大量的实验研究表明, 群落各物种并非如中性理论所假设的那样在生态功能上等价(functional equivalence), 基于功能性状的物种生态位分化和种间作用在群落构建和多样性维持中有着决定性作用(Harpole & Tilman, 2006; Kraft et al., 2008); 而且由于中性理论的模型中群落大小难以界定, 物种分化速率、迁移率和死亡率等各参数可变性很大, 而模型模拟结果对这些参数设置的变化又极其敏感(Zhang & Lin, 1997; Yu et al., 1998), 因而该理论对群落结构的预测也只能是微弱的(weak test)(Wootton, 2005; McGill et al., 2006)。因此, 最近几年来, 越来越多的生态学家倾向于认为, 在群落构建中生态位理论和中性理论并非对立, 将生态位理论和中性理论整合可以更好地理解群落构建的机理(Tilman, 2004; Chase, 2005; Gravel et al., 2006; Leibold & McPeek, 2006)。
本文试图在简要回顾群落中性理论和生态位理论发展的基础上, 梳理其最新研究进展, 并探讨整合生态位理论和中性理论, 解释群落构建和多样性维持的可能途径。
1 群落构建的生态位理论
关于群落多样性的形成和维持机理, 主要存在两类不同的观点。一种观点认为, 在群落构建和多样性维持中, 共存物种间的生态位分化等确定性因素占主导地位; 而另一种观点则认为扩散和随机作用是主要决定因子。关于群落构建的确定性和非确定性因素, 达尔文在其巨著《物种起源》中早有所提及。他认为看似偶然生长在一起的群落物种, 其背后有着深刻的必然性。而对确定性因素和随机作用在群落构建中的地位较为系统的讨论, 可以追溯到20世纪初的群落超有机体论(super-organism concept) (Clements, 1916; Tansley, 1935)和个体论(individualistic concept)(Gleason, 1926)的争论。以Clements为代表的群落有机体学派认为, 群落构建是确定性过程, 群落之间有着可分辨的边界, 群落在受干扰后能够逐渐演变到原来的状态, 即群落演替; 在群落组分种的相互作用下, 演替是从一个方向有规律地向另一个方向变化, 演替的最后阶段是稳定的单元顶极(monoclimax)(Clements, 1916)或多元顶极(polyclimax)(Tansley, 1935)。对此, 以Gleason为代表的群落个体学派提出了猛烈的批评, 他们认为群落只是一些物种的随机组合, 群落之间并没有明显的界限, 群落结构变化也没有明确的方向性, 各物种以其独特的方式响应着环境的时空变化。其后, 岛屿生物地理学的出现、群落构建规则的提出以及群落中性理论的蓬勃发展, 将群落构建中确定性作用和随机过程的相对重要性的讨论推向了高潮。
1.1 生态位和物种共存机制
早在1910年Johnson就率先提出了生态位概念。之后, Grinnell正式定义生态位(niche)为正好被一个种(或亚种)所占据的环境限制性因子单元; 其后Elton指出生态位不仅表现物种对其生活环境的需求(requirements), 而且表明物种在群落关系中的角色以及其对生境的影响(Chase & Leibold, 2003)。Gause(1934)提出竞争排除原理(competitive exclusion principle), 证实了生态位分化是维持物种共存的必要条件。Hutchinson(1957)从多维空间和资源利用等方面提出多维超体积生态位(N-dimensional hyper-volume niche), 并进一步区分了理想条件下的基础生态位(fundamental niche)和实际生境中的现实生态位(realized niche)。Grubb(1977)进一步补充提出更新生态位(regeneration niche)和时间生态位(time niche); 之后Pianka(1983)和Colinvaux(1986)等相继对Hutchinson的多维超体积生态位理论作了不同程度的修正和补充。Litvak和Hansell(1990)认为生态位重叠可以反映群落物种之间的关系。
近年来, 随着生态学趋向量化发展, 对生态位的定义和理解也更加量化和成熟。Shea和Chesson(2002)重新将生态位定义为物种对每个生态位空间点的反应和效应。生态位空间点是指特定时空中环境温度和湿度等物理因子与食物、被捕食者等生物因素的综合; 而种群数量变化则可以反映出物种对空间点的反应结果。Chase和Leibold (2003) 在总结以往研究的基础上, 提出群落生态位研究中应该更加注重物种在大尺度时间和多维空间资源上的综合需求和影响; Silvertown(2004)则更加强调生态位的优先占据原则和种间竞争效应; Timan(2004)最近又提出了随机生态位理论(见后文)。
竞争排除原理证实由于对限制性环境资源的竞争, 生态位相似的物种不能够稳定共存(Gause, 1934); 因此, 在生态位理论的框架下研究者们发展了多种假说来解释群落多物种共存的现象。这些假说包括: Lotka-Volterra竞争模型(Leigh, 1968)、竞争-拓殖权衡(Levine & Rees, 2002)、资源比例假说(Tilman, 1982)、更新生态位理论(Grubb, 1977)、贮存效应(Chesson & Warner, 1981)、微生物介导假说(Zobel et al., 1997), 详见Box 1。

总之, 如今坚持生态位理论的生态学家们认为, 除生境中的可利用资源外, 环境因子等其他因素也影响生态位分化, 促使多物种共存(Chase, 2003; Chase & Leibold, 2003)。这些因素如: (1) 时间的内在和外在变化, 即时间生态位的分化(Grubb, 1977; Chesson, 2000); (2) 空间的内在和外在变化, 当空间本身被看作一种资源时, 物种对空间资源的利用存在着权衡(Murrell & Law, 2003); (3)干扰和环境波动阻碍竞争排除的发生, 从而促进物种的共存(Silvertown, 2004); (4)环境因子以及以上各因素之间的交互作用使得多物种共存(Mouquet et al., 2004)。然而,在自然群落中, 这些确定性的因素依然远远不足以解释复杂的群落结构动态。这使得我们不得不考虑非确定性因素在群落结构动态中的作用。
1.2 基于生态位理论的群落构建规则
对于群落构建机理, Diamond(1975)首次正式提出了群落构建规则(community assembly rules), 认为群落构建是大区域物种库中物种经过多层环境过滤(filtering)和生物作用选入小局域的筛选过程。之后, Wilson和Gitay(1995)将植物群落构建规则定义为在特定生境中一系列限制物种(或一组物种)出现或增多的潜在规则。这些规则是由群落中生物间相互作用决定的非随机性模式(non-random patterns)。Keddy(1992)、Diaz(1998)以及Weiher和Keddy(2001)都对群落构建规则的规范和完善做出了重要贡献。如Keddy(1992)所述, 群落构建可以被认为是物种的筛选过程, 环境条件和生物间的相互作用可以被看作是多个嵌套的筛子(nested sieves); 群落构建将区域物种库中的物种经过这些嵌套筛子的过滤, 只有那些具有特定性状并符合各环境筛的特定物种才能进入小局域群落。图1表示了这种群落构建的筛选过程。
图1
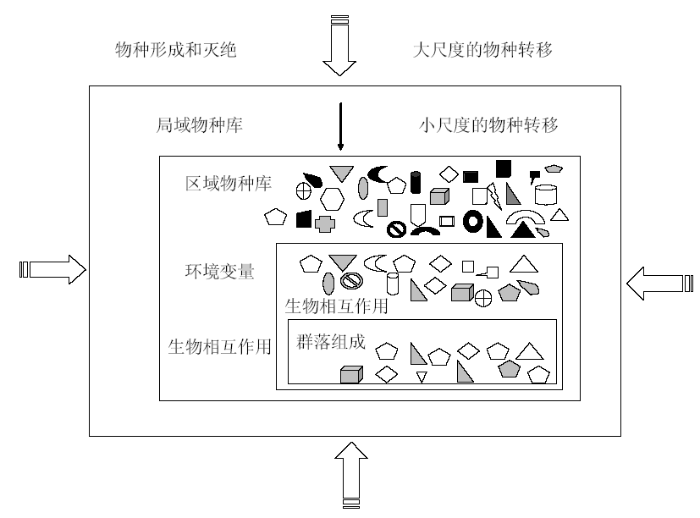
图1
图示群落构建的筛选过程(仿自Keddy, 1992)
Fig. 1
A graphic representation of community assembly process (Simulated from Keddy, 1992)
早在1968年, 种群数量遗传学家Kimura就指出, 在等位基因上发生的突变绝大多数是中性的, 不受环境选择的作用。他认为大多数的宏观进化结果都来自于中性等位基因在遗传漂变中的随机固定(见Box 2)。可以说, Hubbell (2001)的群落中性理论是分子进化中性理论在宏观层次上的推广(Hu et al., 2006)。Hubbell的群落中性理论与分子进化中性理论的不同之处只是在于: 种群遗传学的中性理论不寻求推翻或替代适应性进化的重要性, 而是采用随机过程来补充或完善进化过程中确定性突变-选择理论所不能够解释的环节; 而Hubbell的群落中性理论则试图在忽略达尔文以来建立的“生物性状的适应性分化”思想的前提下解释群落构建机理, 与传统生态位构建思想形成对立(Leigh, 2007)。此外, 这种不依赖物种属性和环境选择来解释群落物种多样性的思想, 还可以追溯到MacArthur和Wilson(1967)的岛屿生物地理学理论。岛屿生物地理学预测, 岛屿(或类似生境)上的物种数取决于物种迁入速率与岛屿上物种灭绝速率之间的动态平衡。迁入速率由岛屿和大陆的距离决定, 灭绝速率由岛屿的面积决定。该理论假设不同物种具有相同的出生/死亡率、迁入/灭绝速率, 并且不考虑物种属性差异以及种间竞争和生态位差异。

2.1 群落中性理论的基本内涵
群落中性理论有两个基本假设: (1)个体水平的生态等价性: 群落中相同营养级所有个体在生态上等价, 更确切的说是有等同的统计(demographic)属性, 即在群落动态中, 所有个体具有相同的出生、死亡、迁入和迁出概率, 甚至物种形成概率; (2) 群落饱和性: 群落动态是一个随机的零和(zero-sum)过程, 也就是说群落中某个个体死亡或迁出马上会伴随着另外一个随机个体的出现以填充其空缺, 这样群落大小不变, 景观中每个局域群落都是饱和的(此后, 中性理论的解析解放宽了这个假设, 详见Volkov et al., 2003)。基于这两个基本假设, Hubbell(2001)认为群落动态实际上是在随机作用下个体的随机生态漂变过程。由此, 群落中性理论能够得出两个主要理论推测: (1) 群落物种多度分布符合零和多项式分布(zero-sum multinomial distribution, ZSM); (2) 扩散限制对群落结构有着决定性作用(图2)。群落中性理论认为其物种多度分布的零和多项式分布综合了Fisher对数级数分布与Preston 对数正态分布, 即在区域集合群落(region)中, 物 种—多度关系符合对数级数分布; 而在局域(local) 群落内, 该关系接近对数正态分布, 数学表达见 Box 3。
图2
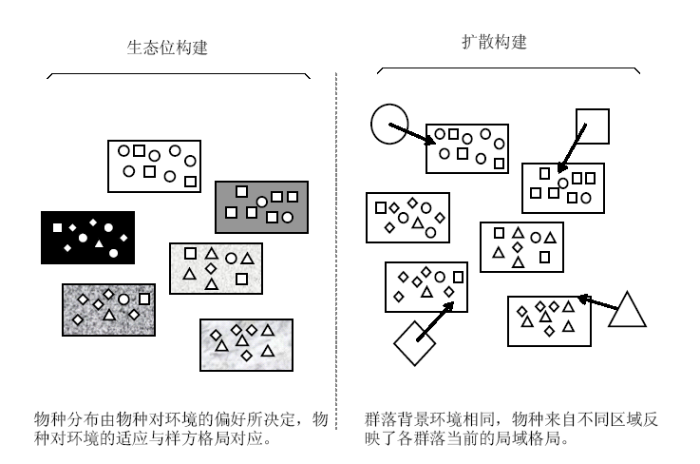
图2
生态位构建和扩散构建下的物种理论分布示意(其中○□◇△代
Fig. 2
A graphic representation of species distribution in different local community (○□◇△ indicate four species and six communities and larger rectangles indicate six communities, respectively. Simulated from Chave, 2008)

群落中性模型明确了Fisher提出的参数α的意义, 即区域个体数与物种形成速率的乘积。另一个参数m则是扩散限制程度的指标。局域群落物种多样性一方面取决于种库的多样性水平(即γ多样性), 另一方面受物种空间分布的不均匀性(即β多样性)影响。群落中性理论明确强调扩散限制对群落物种多样性的影响, 并通过参数模拟(区域多样性由θ表示, 扩散过程由m表示), 将局域群落物种多样性的两种影响因素区分开来。在此基础上, 群落中性理论推导出物种—多度曲线和物种—面积曲线, 以及β多样性格局(Hubbell, 2001)。
2.2 群落中性理论的验证
群落中性理论的提出, 激发了一大批针对特定生态系统群落构建的验证。根据对中性理论检验的角度不同, 我们总结了近年来的一些主要验证研究, 发现相关验证主要集中在中性理论物种等价性假设、零和多项的物种多度分布预测和扩散限制这样三个方面(表1)。
表1 群落中性理论的检验总结
Table 1
| 验证项 | 待检验假设 | 检验区域 | 主要结论 | 相关文献 |
|---|---|---|---|---|
| 物 种 等 价 性 | 检验群落物种的出生率、死亡率和拓殖能力是否相同 | 热带雨林乔木 | 大部分物种的出生率、死亡率和拓殖能力不符合中性假设 | Chave, 2004; Hubbell, 2006 |
| 自然群落中, 物种的功能属性与群落结构是否有关 | 高寒草地、温带草原、热带雨林 | 群落组分种对环境响应各异, 物种功能性状与群落结构变化有关, 不符合中性假设 | Fargione et al., 2003; Adler, 2004; Harpole & Tilman, 2006; Chu et al., 2007; Kraft et al., 2008; Niu et al., 2008 | |
| 物种是否可相互替代以及物种间相互作用是否等价性 | 热带雨林乔木 | 物种不可替代, 种间作用不等价, 如同属内物种相互作用强于异属物种。 | Kelly et al., 2008 | |
| 物种入侵和拓殖成功率是否与入侵物种的属性以及群落结构有关 | 各类入侵物种和被入侵植物群落 | 大多研究表明物种的成功拓殖与物种本身属性和入侵群落属性有密切关系; 也有研究表明入侵与物种和群落属性无关。 | Fargione et al., 2003; Cadotte et al., 2006; MacDougall et al., 2009 | |
| 零 和 多 项 模 型 | 自然群落物种分布模式是否与中性理论预测一致。 | 热带雨林、温带针叶林、亚高寒草地、鱼类、腕足类生物化石、鸟类。 | 大多数的群落中, 物种多度分布符合中性理论预测。 | Condit et al., 2002; Volkov et al., 2003; Alonso & McKane, 2004; Etienne & Olff, 2004a; Olszewski & Erwin, 2004; He, 2005; Houlahan et al., 2007; Volkov et al., 2007; Muneepeerakul et al., 2008; 杜晓光和周淑荣, 2008 |
| 比较中性模型和其他模型种对物种多度分布的拟合。 | 重新检验之前经验检验数据 | 中性模型的拟合符合程度并不比其他模型拟合的好。 | McGill, 2003; McGill et al., 2006;Volkov et al., 2007 | |
| 扩 散 限 制 | 扩散对现实群落构建有作用 | 热带雨林、亚热带常绿阔叶林乔木、温带落叶阔叶林 | 不同的群落类型、不同研究尺度, 生态位构建和扩散限制相对作用贡献不同, 如 | Condit et al., 2002; Tuomisto, 2003; Gilbert et al., 2004; Karst et al., 2005; Jones et al., 2006; Cielo et al., 2007; Chave, 2008; Jones et al., 2008; Legendre et al., 2009 |
| 岛屿中的物种数是否与物种属性有关。 | 岛屿生物的重新拓殖, 植物和蝴蝶种群等。 | 岛屿植物拓殖能力与其叶片坚韧程度相关; 蝴蝶多度与其寄主植物生物量相关 | Yamamoto et al., 2007; Burns & Neufeld, 2009 |
物种生态等价性: 中性理论假设群落内所有个体在生态功能上等价, 这一假设与传统的生态位理论相悖, 也因此受到了多方面的质疑。根据群落内个体的生态等价性假设, 可以推测各物种在生态上也是等价的(如群落物种入侵和拓殖机会同等) (Hubbell, 2001, 2006)。但大量前人研究和针对中性理论物种生态等价性的验证都表明各群落组分种的生态功能并不对称, 各组分种对环境变化的响应也不尽相同, 群落结构维持和变化与组分种的属性密切相关(表1)。
零和多项模型: 在物种等价和群落饱和的基本假设以及物种扩散限制前提下, 群落中性理论预测群落多度应该符合零和多项分布。由于比较群落物种多度分布是否与零和多项分布预测相符, 直观而又易操作, 使其成为早期中性理论验证的重要方面。早期简单的比较发现对于大部分的群落, 零和多项模型都能给出较好的预测(Hubbell, 2001; Bell, 2000)。但McGill等(2003, 2006, 2007)对以往检验采取更为严格多模型比较重新评估后, 发现多种模型都能对种多度做出预测, 零和多项模型的拟合并不总是占优势。在此, 值得我们思考的是, 模型预测与实际群落格局是否符合可能并不是判断群落是否中性的充分条件, 而只是一种必要条件, 因为不同的机制可能导致同样的格局(He, 2005; Purves & Pacala, 2005; Hubbell, 2006)。此外, 按中性理论的零和多项分布预测, 群落物种之间将会出现负的时间相关, 然而, Houlahan等(2007)分析了大量的数据后发现, 自然群落中物种间并不存在负的时间相关。
扩散限制: 中性理论的一个重要推论是扩散限制(距离效应)显著影响群落结构: 随着空间距离的增加, 群落的相似性降低, 两者具体的数量关系可以由种子扩散曲线所推导出, 这种扩散限制对于每个物种都是相同的(Hubbell, 2001; Condit et al., 2002)。与此对应, 生态位理论认为, 每个物种生活在其最适宜的环境内, 即拥有自己的生态位, 因而物种间的生态位分化将导致不同环境条件下的群落组成不同, 两个群落环境条件越相似, 则其群落组成越相似(两种不同模式构建群落的方式如图2示意)。近来, 研究者越来越倾向于认为生态位分化与扩散限制对群落的构建都有作用, 而两者的相对贡献与研究尺度和生态系统类型有关(Legendre et al., 2009) (详见表1及图3)。
图3
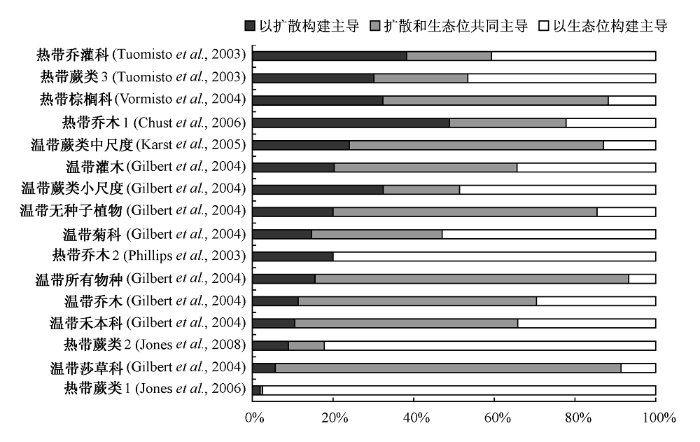
图3
扩散限制验证的8个生态系统的16个研究对象中扩散主导、生态位主导以及两者共同作用的相对贡献
Fig. 3
Summary of the relative importance of dispersal and niche assembly in 16 ecosystems from eight studies
2.3 群落中性理论的改进
在Hubbell提出群落中性理论之后, 学者们提出了十多个类似的群落中性模型。这些模型在对零和假设的定义、集合群落模式以及模型参数的模拟途径等方面进行了不同程度的改进和修正, 发展了群落中性理论。其中Volkov等(2003)使用前推法, 采用马尔可夫链描述状态和转移概率, 以主方程(master equation)控制模型, 分别推导出集合群落和局域群落物种—多度分布的解析解。解析解的求出为预测给定参数的种群结构和种群动态提供了便利(Vallade & Houchmandzadeh, 2003; Alonso & McKane, 2004; McKane et al., 2004)。Etienne等则利用后推法(即在扩散限制条件下, 每个个体都被追溯到迁入群落时的祖先, 从而得到模型参数的概率分布, 也可得到给定群落物种-多度分布的中性模型参数)提出了多元联合模型(multivariate coalescence model)和抽样规则(sampling formula) (Etienne & Olff, 2004b; Etienne, 2005)。前推法和后推法从各自不同的角度完善了Hubbell(2001)中性模型的解析分析。
此外, 基于对热带雨林的持续观测和实验结果, Hubbell等(2008)对早期的中性理论模型进行了修正和拓展。Volkov等(2005)将密度限制因子c加入中性模型, 以替代原模型中的扩散因子m, 发现拟合效果与原模型不相上下。密度限制属于经典的生态位理论, 反映了资源环境容量限制, 或自上而下的捕食者控制。因此, 这个模型将群落模式和关键物种的种群动态联系起来, Hubbell (2008)坚持认为只要密度制约效应(density dependence effect)对 每个物种的作用相同, 群落仍然可以被看作是中 性的。
与以前对模型的参数修正不同, 近几年的研究主要致力于群落中性模型的“真实化”。如一些科学家以种子扩散曲线反映扩散限制, 提出更接近真实群落的空间显式中性模型(spatially explicit neutral models), 来替代Hubbell(2001)对扩散限制的空间隐性处理(Chave & Leigh, 2002; Zillio & Condit, 2007)。Rosindell和Cornell(2007)又完善了显式中性模型在大区域集合群落抽样面积问题。但由于空间显式模型的复杂性, 而且模型没有解析解, 甚至还没有较好的模拟方法, 使其实用性受到限制, 有待进一步发展。
由于中性模型中群落个体生态等价假设与实际状况不符, Zhou和Zhang(2008)以及Lin等(2009)最近提出近中性模型(nearly neutral models), 通过在模型参数中纳入微小的随机种间差别来放宽该假设, 使得群落中性模型更为稳定。Zhou和Zhang(2008)在其近中性模型中, 让群落中新迁入或新种的竞争力(以结实率表达)不再严格遵守中性假设, 而是服从具有微小方差的正态分布。这样在不引进生态位分化的前提下, 随机波动与扩散限制一起维持了近中性群落的物种结构。Lin等(2009)假定物种的出生率和死亡率等统计参数在权衡上存在种间差异, 但最终使种间的适合度一致; 当在一个近中性抽彩模型(lottery model)中包含出生–死亡率权衡和种间差异的情况下模拟适合度等同的物种共存动态, 结果发现这种权衡使得物种共存的时间更长, 并且群落中稀有种的死亡率往往低于那些优势种。这一结果与Volkov等(2007)在热带雨林中的研究是一致的。
3 中性理论与生态位理论的整合
群落中性理论作为对生态学基本概念最激进的挑战之一, 就目前的发展来看并未颠覆经典的生态位群落构建理论, 但它揭示了随机过程对群落构建的影响, 进而对群落构建理论产生了重大影响。近年来, 越来越多的批评者开始接受群落中性理论的合理部分, 如通过群落构建的零模型来检验随机作用对群落构建的影响。生态位理论的支持者试图将中性理论的合理部分整合至基于生态位的群落构建理论体系中(见下文), 而中性理论也在简约原则下纳入确定性因子, 以弥补其关键的缺陷(Hubbell, 2006; Zhou & Zhang, 2008; Lin et al., 2009)。如今, 我们看到群落生态学中已经产生了一种明显的趋势, 即通过整合中性理论与生态位理论来完善群落构建的机理探讨(Alonso et al., 2006; Adler et al., 2007)。
3.1 群落中性理论的质疑
群落中性理论以其简约结构和预测能力, 对群落构建机理的生态位理论提出了重大的挑战, 甚至可以说为群落生态学带来了一场革命。但由于中性理论的物种生态等价性假设与实际不符, 使其饱受质疑。由于否定物种差别, 群落中性理论至少无法解释如下事实: (1)群落组分种对环境变化的响应因种而异, 尤其是物种对生境破碎化的响应; (2)不同物种生物入侵能力有显著差异, 群落抗入侵能力也各不相同(Cadotte et al., 2006; MacDougall et al., 2009); (3)物种多样性与生态系统功能之间的正相关关系, 如生产力因物种数增加而上升的现象(Naeem et al., 2002)。此外, 如表1所示, 群落中性理论对群落模式的预测结果也值得商榷(McGill et al., 2006)。
上述质疑以及群落中性理论面对现实生态问题时的无能为力, 使其有效性和理论价值被严重削弱。如在过去10年里, 有关中性理论的讨论一方面成为理论生态学界的焦点, 另一方面却很少得到Conservation Biology这类应用性刊物的关注(Clark, 2009)。 最近, Clark等(2007, 2009)从认识论的角度对群落中性理论提出了批评。认为在对随机性的认识和处理方式上, 群落中性理论错误地以随机论掩盖未知过程(stochasticity stands in for ignorance)。他们认为, 科学认识的一般途径是通过挖掘确定性过程(即机制), 来降低模型预测的随机误差; 而中性理论放弃关于过程的现有知识, 用随机动态来替代未知过程。这种剥离过程(process-free)的方法论可能会对群落生态学乃至整个生态学的发展带来负面影响(Clark et al., 2007; Clark, 2009)。但Clark的这种说法也值得商榷, 因为群落中性理论对种群动态的概率假设并不意味着就是剥离过程(即并不是增加误差项)。另外, Clark等的质疑还主要针对关于中性理论的“生态等同性”前提, 他认为其中存在三种概念混淆, 即用等价化机制(equalizing mechanism)解释竞争物种的共存, 用模型的对称性推断自然群落的中性, 将扩散限制视为功能或统计等价性。
3.2 中性理论与生态位理论的整合
如前所述, 尽管群落中性理论饱受争议, 但依然显示出强大的生命力, 这不仅是由于群落中性理论的简约性和预测力, 为追求简约通则的现代生态学带来了一丝曙光, 更重要的是它强调了群落构建中随机过程的作用。如今许多生态学家认为这两种理论其实是相容互补而不是绝对对立的, 两者从不同的侧面反映和描述了群落的内禀特征。群落构建是随机的生态漂变和生态位分化共同作用的过程, 如今更多的研究开始重视整合生态位和中性理论探究群落构建中随机作用和确定性作用的相对贡献。研究者们从不同的视角试图将中性理论的合理部分整合到生态位的框架中, 以期推动理解群落构建的机理。例如, 一些研究者主张将中性理论作为群落构建的零模型(Harte, 2004; Gaston & Chown, 2005; Gotelli & McGill, 2006); Gravel(2006)等则提出中性-生态位连续体假说。
3.2.1 中性理论作为群落构建的零模型
群落生态学中零模型的说法由来已久(Caswell, 1976; Harvey et al., 1983), 而将中性理论作为群落构建的零模型则是由Harte(2004)正式提出的。他认为群落中性理论可能因其许多缺陷而是“错误”的, 然而由于中性理论所表述的是在纯随机作用下的群落构建状态, 所以可以被看作群落构建理论的零模型。此后, Gaston和Chown(2005)也指出, 中性理论由于忽视实际存在的物种生态对策和生态位分化在群落构建中的作用, 显然是错误的, 然而群落中性模型的预测与许多观测结果相符, 这也意味着群落中性理论所描述的随机作用在群落构建中也有着相应的作用, 只是它仅在特定的条件下适用, 故可以被作为群落构建的零模型。
Gotelli和McGill(2006)则较为系统地分析了群落中性理论和零模型之间的异同, 认为虽然由于群落中性理论考虑了随机过程的作用并且可以检验实际数据, 可以被当作是一种特殊的零模型, 但是中性理论与标准零模型之间存在三个重要的不同: (1)多数零模型包含了一些基于生态位分化的物种水平参数, 而中性理论则假设所有物种在功能上等价; (2)零模型中的一些参数从实际数据本身直接获得, 而中性模型中的物种形成和灭绝率以及转移率几乎都不可能直接测得, 只能通过拟合或推测; (3)零模型常被认为只是一种统计描述式, 而中性理论则明确指出随机作用下的生态漂变是群落构建的机理性过程。
总之, 从定性来看, 中性理论确实与典型的零模型相似, 然而由于中性理论重要参数的不可测性, 它作为零模型用于实验验证的价值就非常有限。尽管如此, 将中性模型拟合的模式与其他非中性机理模型拟合的结果作比较, 仍有助于促进对群落构建机理的理解, 同时, 以简单的零模型评估中性模型就显得很重要。
3.2.2 群落生态位—中性连续体
Gravel等(2006)在整合群落中性理论和生态位理论的基础上提出生态位—中性连续体。他们认为: (1) 竞争和随机漂变可以同时作用于群落构建和群落动态中, 它们作用的相对大小决定了群落构建是一个从纯生态位构建到纯中性构建的连续体; (2) 竞争和扩散的共同影响足以产生这种连续体, 物种间的竞争不对称性决定了种群动态对繁殖体供给的影响程度; (3)竞争和随机排除之间的平衡决定了群落内物种相对多度的分布模式。在相对独立的群落中, 从集合群落中的迁入并不足以平衡这两种排除过程。在物种多样性低的群落中, 生态位分化是影响物种多度分布的主导因子, 物种之间生态位重叠很少或几乎没有, 物种相对多度决定于环境资源的分布状况; 而在多样性高的群落内, 物种间生态位重叠极大, 高迁入率又使得群落限制性和相似性增加, 也抑制了物种丰富度的增大, 群落动态则由随机排除所主导。中性—生态位连续体模型预测, 生态位重叠的增加和群落中性会有助于物种丰富度的增加; 而低的环境异质性同样能够使物种数目不多的群落中物种生态位重叠增大, 使群落趋于中性。总之, Gravel等(2006)预测当生态位互补使群落达到饱和时, 繁殖体迁入维持了物种冗余(redundancy), 生态漂变能够增加群落的物种丰富度。生态位重叠和扩散限制的交互作用是中性—生态位连续体的基本特征。
值得一提的是, 许多实验研究为这种中性—生态位连续体假说提供了有力的支持。Hubbell(2001)指出温带森林—热带森林的群落动态表现了这种连续状态; 青藏高原高寒草地上的次生演替也为此提供了有力的支持(Chu et al., 2007)。 当然, 关于中性—生态位连续体的实验验证依然十分缺乏, 其内在机理尚有待于进一步的探究(图3表示了已有的相关验证)。
3.2.3 其他整合假说
Tilman(2004)从研究群落物种入侵的问题出发, 提出随机生态位理论(stochastic niche theory), 将随机过程的作用加入到生态位权衡的群落动态中。随机生态位理论认为, 群落的构建过程实质是一个繁殖体不断入侵的过程。物种入侵群落的过程是随机的, 而是否在群落中成功拓殖则取决于入侵种和已有物种之间的资源竞争等。Tilman(2004)认为其随机生态位理论能够很好地解释群落多样性维持、物种多度分布和物种入侵等现象, 弥补了经典生态学物种权衡理论和中性理论的许多不足。
Purves和Pacala(2005)提出中性作用存在于生态位群落构建中, 认为群落构建的生态位理论和中性理论之争, 是由于错误地以群落结构是否与中性模型预测一致作为判断群落是否中性的标准(也有一些科学家认为这种做法仅仅是验证中性理论的必要条件, 但不是充分条件, 详见McGill et al., 2006; McGill et al., 2007)。Purves和Pacala(2005)在Hubbell经典的零和多项中性漂变模型中加入演替生态位、抽彩生态位和生境特化后, 发现在生物多样性高的群落中, 中性漂变模型模拟的物种多度分布依然符合实际的群落结构。他们认为这一结果揭示, 群落随机漂变中存在生态位分化依然能够产生中性群落; 反之, 随机漂变可以发生在生态位群落构建过程中, 群落的中性格局并不意味着物种在生态学上是等价, 更不意味着群落动态是中性的。他们预测在多样性高的群落中, 物种多度分布决定于生态漂变, 而不依赖于生态位结构; 与此相反, 生态系统生物地化循环功能则完全取决于群落的物种生态位结构, 并不依赖于生态漂变。Purves和Pacala的这些观点看似新颖有理, 但是缺乏直接的实验支持。
Hérault(2007)提出了资源群(emergent group)概念, 试图调和生态位和中性整合之间的矛盾。他定义资源群为一系列功能上相似并且生态对策趋同的物种, 群落由为数不多的功能群组成。在功能群内, 物种数量和多度的变化更倾向于中性的过程, 物种丰富度由零和多项式下的个体随机丧失和迁入平衡决定; 而在功能群间, 由于功能差异明显, 生态位构建是决定物种丰富度的主导作用。虽然这种资源群的提法由来已久(Lavorel & Garnier, 2002), 然而, 这一理论为我们以物种功能性状量化生态位和中性作用在群落构建中的相对贡献提供了非常有价值的新思路。
此外, Adler等(2007) 认为群落中性过程类似于多物种共存中的同一化(equaling)机制, 而基于生态位的群落构建与多物种共存中的稳定化(stablizing)机制类似。这两种过程相互补充, 相互作用, 共同决定着物种共存和生物多样性维持(Chesson, 2000; Adler et al., 2007)。这更意味着在群落构建的实验验证中, 我们不应评价生态位或中性过程各自的重要性, 而应检验二者的相对重要性。如McGill等(2006)以量化的物种属性指标, 如基础代谢率、种子或卵的大小、植物元素的化学计量学等替代描述性的“种”的分类指标等等。因此, 检验物种性状如何促进群落的稳定化和同一化就显得更加重要, 这些思想都体现了这种整合的思路。
综上所述, 在两种理论融合的问题上, 倾向于生态位理论的学者认为, 中性的模式属于缺少稳定机制并且各物种拥有相同适合度的特殊状态(Adler et al., 2007); 倾向于中性理论的学者则认为生态位决定的状态是群落动态过程处于平衡点的特殊状态, 一旦这个暂时的平衡状态被打破, 群落又会向平衡点两侧变化(Gewin, 2006)。融合生态位分化和中性作用的假说都认为, 环境变化是驱动连续体从一极向另一极移动的主要驱动力。例如在土壤贫瘠、物种组成相对简单的草地生态系统中, 生态位分化的贡献可能更大; 而在物种丰富的热带雨林群落中, 多样性的维持可能主要由中性作用所决定(Gravel et al., 2006)。总之, 生态位和中性理论争论的核心问题是生态位分化和随机作用在群落构建和生物多样性维持中的相对贡献大小问题; 换言之, 就是在环境梯度上, 物种功能性状和随机作用在决定群落物种相对多度分布中的相对贡献问题。
4 总结
关于群落构建机理的探究由来已久, 与许多宇宙现象和生命现象本身一样, 随机作用和确定性因素相互依存, 共同决定了生命形式和多样性的维持。基于物种生态位分化的群落构建理论虽然历经了长达近一个世纪的发展, 但依然不完善, 尤其对随机作用的关注不足, 对现实自然群落的预测力不强, 更不能对生物多样性模式、物种多度分布、物种—面积关系等群落模式作出准确的预测, 这就给群落中性理论的发展留下了空间。群落中性理论虽因其基本假设和预测不尽符合实际而饱受争议, 却因其结构的简约性和预测能力受到了生态学家的重视。更重要的是, 群落中性理论激发了人们对随机作用在群落构建和生物多样性维持中作用的重新思考和定位, 科学家们也越来越认识到群落构建是随机漂变和生态位构建的共同作用。例如,尽管Hubbell(2001)最初宣称中性理论是“群落与生物地理的统一理论”, 但后来也不得不接受物种不可能生态等价的事实(Hubbell, 2005, 2006)。
我们发现, 经过了近20年的争论,生态学中群落中性理论和生态位理论之争的声音已经是那么的微弱, 更多的生态学家则在致力于将二者整合于一体, 以更好地理解群落构建机理。然而, 复杂的自然世界又为我们提出了新的科学难题, 即基于生态位分化的确定性过程和中性作用的随机过程怎样耦合在一起, 共同形成和维持物种多样性?在这个问题上, 人们的探索才刚刚开始, 而群落构建的零模型、中性-生态位连续体以及随机生态位等假说的提出为尝试这种整合建立了良好的开端, 今后的研究可能需要更加关注将检验拓展到除植物以外的其他生物群落, 如土壤动物和微生物群落当中。在不同类型群落的检验可以帮助我们更好地理解确定性过程和随机过程在决定群落构建中的作用。
致谢:
北京师范大学张大勇教授以及两位匿名审稿人对本文初稿修改提供了具有建设性的意见, 特致谢忱!
参考文献
Niche evolution and adaptive radiation: testing the order of trait divergence
Neutral models fail to reproduce observed species-area and species-time relationships in Kansas grasslands
A niche for neutrality
The merits of neutral theory
DOI:10.1016/j.tree.2006.03.019
URL
PMID:16766082
[本文引用: 2]

Hubbell's neutral theory of biodiversity has challenged the classic niche-based view of ecological community structure. Although there have been many attempts to falsify Hubbell's theory, we argue that falsification should not lead to rejection, because there is more to the theory than neutrality alone. Much of the criticism has focused on the neutrality assumption without full appreciation of other relevant aspects of the theory. Here, we emphasize that neutral theory is also a stochastic theory, a sampling theory and a dispersal-limited theory. These important additional features should be retained in future theoretical developments of community ecology.
Sampling Hubbell’s neutral theory of biodiversity
The distribution of abundance in neutral communities
DOI:10.1086/303345
URL
PMID:10777433
[本文引用: 3]
The patterns of abundance generated by a simple stochastic birth-death-immigration model are described in order to characterize the diversity of neutral communities of ecologically equivalent species. Diversity is described by species number S and the variance of frequency or log abundance q approximately . The frequency distribution of abundance is very generally lognormal, skewed to the left by immigration and resembling descriptions of natural communities. Increased immigration and community size always cause S to increase. Their effect on q approximately is more complicated, but given biologically reasonable assumptions, S and q approximately will be positively correlated in most circumstances. Larger samples contain more species; the graph of log S on log individuals, equivalent to a species-area curve, is generally convex upward but becomes linear with a slope of about +0.25 when immigration is low and births exceed deaths. When individuals invade a new, vacant environment, both S and q approximately increase through time. Thus, a positive correlation between S and q approximately will usually be generated when sites of differing size or age are surveyed. At equilibrium, communities maintain roughly constant levels of S and q approximately but change in composition through time; composition may remain similar, however, for many generations. Many prominent patterns observed in natural communities can therefore be generated by a strictly neutral model. This does not show that community structure is determined exclusively by demographic stochasticity, but rather demonstrates the necessity for an appropriate null model when functional hypotheses are being tested.
The interpretation of biological surveys
Plant extinction dynamics in an insular metacommunity
Conceptual Ecology and Invasion Biology: Reciprocal Approaches to Nature
Community structure: a neutral model analysis
Community assembly: when should history matter?
DOI:10.1007/s00442-003-1311-7
URL
PMID:12836009
[本文引用: 2]
Community assembly provides a conceptual foundation for understanding the processes that determine which and how many species live in a particular locality. Evidence suggests that community assembly often leads to a single stable equilibrium, such that the conditions of the environment and interspecific interactions determine which species will exist there. In such cases, regions of local communities with similar environmental conditions should have similar community composition. Other evidence suggests that community assembly can lead to multiple stable equilibria. Thus, the resulting community depends on the assembly history, even when all species have access to the community. In these cases, a region of local communities with similar environmental conditions can be very dissimilar in their community composition. Both regional and local factors should determine the patterns by which communities assemble, and the resultant degree of similarity or dissimilarity among localities with similar environments. A single equilibrium in more likely to be realized in systems with small regional species pools, high rates of connectance, low productivity and high disturbance. Multiple stable equilibria are more likely in systems with large regional species pools, low rates of connectance, high productivity and low disturbance. I illustrate preliminary evidence for these predictions from an observational study of small pond communities, and show important effects on community similarity, as well as on local and regional species richness.
Towards a really unified theory for metacommunities
A spatially explicit neutral model of beta-diversity in tropical forests
DOI:10.1006/tpbi.2002.1597
URL
PMID:12167354
[本文引用: 1]
To represent species turnover in tropical rain forest, we use a neutral model where a tree's fate is not affected by what species it belongs to, seeds disperse a limited distance from their parents, and speciation is in equilibrium with random extinction. We calculate the similarity function, the probability F(r) that two trees separated by a distance r belong to the same species, assuming that the dispersal kernel P(r), the distribution of seeds about their parents and the prospects of mortality and reproduction, are the same for all trees regardless of their species. If P(r) is radially symmetric Gaussian with mean-square dispersal distance sigma, F(r) can be expressed in closed form. If P(r) is a radially symmetric Cauchy distribution, then, in two-dimensional space, F(r) is proportional to 1/r for large r. Analytical results are compared with individual-based simulations, and the relevance to field observations is discussed.
Mechanisms of maintenance of species diversity
On the balance between niche and neutral processes as drivers of community structure along a successional gradient: insights from alpine and sub-alpine meadow communities
DOI:10.1093/aob/mcm166
URL
PMID:17720679
[本文引用: 2]
90 species was collected in alpine and sub-alpine meadows of the Tibet Plateau (China). A successional gradient (1, 3, 15 and 30 years after abandonment) was established in a sub-alpine meadow. The relationships between species traits and their abundance were evaluated using regression models. KEY RESULTS: Seed mass was negatively related to both species density (r = -0.6270, P 0.07 in the 3-, 15- and 30-year meadows). Data gathered in all sites showed a significant negative relationship between the average individual biomass of a given species and its density within the community (r
Position on slope, disturbance, and tree species coexistence in a seasonal semideciduous forest in SE Brazil
Beyond neutral science
DOI:10.1016/j.tree.2008.09.004
URL
PMID:19026462
[本文引用: 3]
Biodiversity science is unusual in that an emerging paradigm is not based on a specific process, but rather depends largely on stochastic elements, perceived as neutral forces. Here I suggest that these forces, which have been justified, in part, by the concepts of symmetry and equalizing mechanisms, have application to the understanding of stochastic models but do not constitute forces that operate in nature. Another process now regularly classified as a neutral force, limited dispersal, represents a fundamental demographic process that is not neutral with respect to species differences, but rather differs among species in important ways. Finally, I suggest that the dramatic shift in ecological research to focus on neutrality could have a cost in terms of scientific understanding and relevance to real biodiversity threats.
Resolving the biodiversity paradox
DOI:10.1111/j.1461-0248.2007.01041.x
URL
PMID:17594418
[本文引用: 2]
The paradox of biodiversity involves three elements, (i) mathematical models predict that species must differ in specific ways in order to coexist as stable ecological communities, (ii) such differences are difficult to identify, yet (iii) there is widespread evidence of stability in natural communities. Debate has centred on two views. The first explanation involves tradeoffs along a small number of axes, including 'colonization-competition', resource competition (light, water, nitrogen for plants, including the 'successional niche'), and life history (e.g. high-light growth vs. low-light survival and few large vs. many small seeds). The second view is neutrality, which assumes that species differences do not contribute to dynamics. Clark et al. (2004) presented a third explanation, that coexistence is inherently high dimensional, but still depends on species differences. We demonstrate that neither traditional low-dimensional tradeoffs nor neutrality can resolve the biodiversity paradox, in part by showing that they do not properly interpret stochasticity in statistical and in theoretical models. Unless sample sizes are small, traditional data modelling assures that species will appear different in a few dimensions, but those differences will rarely predict coexistence when parameter estimates are plugged into theoretical models. Contrary to standard interpretations, neutral models do not imply functional equivalence, but rather subsume species differences in stochastic terms. New hierarchical modelling techniques for inference reveal high-dimensional differences among species that can be quantified with random individual and temporal effects (RITES), i.e. process-level variation that results from many causes. We show that this variation is large, and that it stands in for species differences along unobserved dimensions that do contribute to diversity. High dimensional coexistence contrasts with the classical notions of tradeoffs along a few axes, which are often not found in data, and with 'neutral models', which mask, rather than eliminate, tradeoffs in stochastic terms. This mechanism can explain coexistence of species that would not occur with simple, low-dimensional tradeoff scenarios.
Fecundity of trees and the colonization-competition hypothesis
Plant Succession: An Analysis of the Development of Vegetation
Beta-diversity in tropical forest trees
DOI:10.1126/science.1066854
URL
PMID:11809969
[本文引用: 3]
The high alpha-diversity of tropical forests has been amply documented, but beta-diversity-how species composition changes with distance-has seldom been studied. We present quantitative estimates of beta-diversity for tropical trees by comparing species composition of plots in lowland terra firme forest in Panama, Ecuador, and Peru. We compare observations with predictions derived from a neutral model in which habitat is uniform and only dispersal and speciation influence species turnover. We find that beta-diversity is higher in Panama than in western Amazonia and that patterns in both areas are inconsistent with the neutral model. In Panama, habitat variation appears to increase species turnover relative to Amazonia, where unexpectedly low turnover over great distances suggests that population densities of some species are bounded by as yet unidentified processes. At intermediate scales in both regions, observations can be matched by theory, suggesting that dispersal limitation, with speciation, influences species turnover.
Biological invasions and the neutral theory
Plant functional traits and environmental filters at a regional scale
Testing the neutral theory of plant communities in subalpine meadow
A new sampling formula for neutral biodiversity
How dispersal limitation shapes species-body size distributions in local communities
DOI:10.1086/380582
URL
PMID:14767837
[本文引用: 1]
A critical but poorly understood pattern in macroecology is the often unimodal species-body size distribution (also known as body size-diversity relationship) in a local community (embedded in a much larger regional species pool). Purely neutral community models that assume functional equivalence among species are incapable of explaining this pattern because body size is the key determinant of functional differences between species. Several niche-based explanations have been offered, but none of them is completely satisfactory. Here we develop a simple model that unites a neutral community model with niche-based theory to explain the relationship. In the model, species of similar size are assumed to belong to the same size guild. Within a size guild, all individuals are equivalent in their competition for resources, sensu Hubbell's neutral community model; they have the same speciation rate and dispersal capacities. Between size guilds, however, the total number of individuals, the speciation rate, and the dispersal capacities differ, but using known allometric scaling laws for these properties, we can describe the differences between size guilds. Our model predicts that species richness reaches an optimum at an intermediate body size, in agreement with observations. The optimum at intermediate body size is basically the result of a trade-off between, on the one hand, allometric scaling laws for the number of individuals and the speciation rate that decrease with body size and, on the other hand, the scaling law for active dispersal that increases with body size.
A novel genealogical approach to neutral biodiversity theory
Sampling theory of selectively neutral alleles
DOI:10.1016/0040-5809(72)90035-4 URL PMID:4667078
Community assembly and invasion: an experimental test of neutral versus niche processes
Neutrality, niches, and dispersal in a temperate forest understory
The individualistic concept of the plant association
Environmental variability promotes coexistence in lottery competitive systems
Null versus neutral models: what’s the difference?
Reconciling niche and neutrality: the continuum hypothesis
Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences
The maintenance of species richness in plant communities: the importance of the regeneration niche
Non-neutral patterns of species abundance in grassland communities
DOI:10.1111/j.1461-0248.2005.00836.x
URL
PMID:16958864
[本文引用: 2]
Although the distribution of plant species abundance in a Minnesota grassland was consistent with neutral theory, niche but not neutral mechanisms were supported by the ability of species traits to predict species abundances in three experimental grassland communities. In particular, data from 27 species grown in monoculture showed that species differed in a trait, R*, which is the level to which each species reduced the concentration of soil nitrate, the limiting soil nutrient and which is predicted to be inversely associated with competitive ability for nitrogen (N). In these N-limited habitats, species abundance ranks correlated with their predicted competitive ranks: low R* species, on average dominated. These correlations were significantly different than expected for neutral theory, which assumes the exchangeability of species traits. Additionally, we found that changes in relative abundance after environmental change (N-addition or disturbance) were not neutral but also were significantly associated with R*.
Null models in ecology
Deriving a neutral model of species abundance from fundamental mechanisms of population dynamics
Reconciling niche and neutrality through the emergent group approach
Neutral theory in population genetics and macroecology
Emergent neutrality
DOI:10.1016/j.tree.2006.08.003
URL
PMID:16901580
[本文引用: 1]
Community ecology is in a current state of creative ferment, stimulated by the development of neutral models of community organization. Here, I reflect on recent papers by Scheffer and van Nes, and by Gravel et al., which illuminate how neutrality can emerge from ecological and evolutionary processes, thus suggesting ways to unify neutral and niche perspectives.
Compensatory dynamics are rare in natural ecological communities
Neutral theory in community ecology and the hypothesis of functional equivalence
The neutral theory and evolution of ecological equivalence
Concluding remarks: population studies, animal ecology and demography
Homage to santa rosalia or why are there so many kinds of animals?
Explaining variation in tropical plant community composition: influence of environmental and spatial data quality
DOI:10.1007/s00442-007-0923-8
URL
PMID:18064493
[本文引用: 1]
The degree to which variation in plant community composition (beta-diversity) is predictable from environmental variation, relative to other spatial processes, is of considerable current interest. We addressed this question in Costa Rican rain forest pteridophytes (1,045 plots, 127 species). We also tested the effect of data quality on the results, which has largely been overlooked in earlier studies. To do so, we compared two alternative spatial models [polynomial vs. principal coordinates of neighbour matrices (PCNM)] and ten alternative environmental models (all available environmental variables vs. four subsets, and including their polynomials vs. not). Of the environmental data types, soil chemistry contributed most to explaining pteridophyte community variation, followed in decreasing order of contribution by topography, soil type and forest structure. Environmentally explained variation increased moderately when polynomials of the environmental variables were included. Spatially explained variation increased substantially when the multi-scale PCNM spatial model was used instead of the traditional, broad-scale polynomial spatial model. The best model combination (PCNM spatial model and full environmental model including polynomials) explained 32% of pteridophyte community variation, after correcting for the number of sampling sites and explanatory variables. Overall evidence for environmental control of beta-diversity was strong, and the main floristic gradients detected were correlated with environmental variation at all scales encompassed by the study (c. 100-2,000 m). Depending on model choice, however, total explained variation differed more than fourfold, and the apparent relative importance of space and environment could be reversed. Therefore, we advocate a broader recognition of the impacts that data quality has on analysis results. A general understanding of the relative contributions of spatial and environmental processes to species distributions and beta-diversity requires that methodological artefacts are separated from real ecological differences.
Effects of mesoscale environmental heterogeneity and dispersal limitation on floristic variation in rain forest ferns
Fern community assembly: the roles of chance and the environment at local and intermediate scales
Assembly and response rules: two goals for predictive community ecology
Phylogeny, niches, and relative abundance in natural communities
DOI:10.1890/07-0322.1
URL
PMID:18481521
[本文引用: 1]
Community structure refers to the number of species in a community and the pattern of distribution of individuals among those species. We use a novel way of representing community structure to show that abundance within closely related pairs of co-occurring tree species in a highly diverse Mexican forest is more equitable than is abundance within more distantly related pairs. This observation is at odds with the fundamental assumption of neutral models of community structure, i.e., that species are interchangeable. The observed patterns suggest niche apportionment, in which interaction is focused pairwise between congeners but falls away from the phylogenetic structure above the genus level. Thus niche processes may significantly affect community structure through regulating relative abundance in a substantial proportion of species, which in turn potentially enhances community stability. One such mechanism of stable coexistence has already been shown to be active in this forest.
Evolutionary rate at the molecular level
DOI:10.1038/217624a0 URL PMID:5637732
Functional traits and niche-based tree community assembly in an Amazonian forest
DOI:10.1126/science.1160662
URL
PMID:18948539
[本文引用: 2]
It is debated whether species-level differences in ecological strategy, which play a key role in much of coexistence theory, are important in structuring highly diverse communities. We examined the co-occurrence patterns of over 1100 tree species in a 25-hectare Amazonian forest plot in relation to field-measured functional traits. Using a null model approach, we show that co-occurring trees are often less ecologically similar than a niche-free (neutral) model predicts. Furthermore, we find evidence for processes that simultaneously drive convergence and divergence in key aspects of plant strategy, suggesting that at least two distinct niche-based processes are occurring. Our results show that strategy differentiation among species contributes to the maintenance of diversity in one of the most diverse tropical forests in the world.
Assessing the scale-specific importance of niches and other spatial processes on beta diversity: a case study from a temperate forest
DOI:10.1007/s00442-008-1214-8
URL
PMID:19018575
Niche processes and other spatial processes, such as dispersal, may simultaneously control beta diversity, yet their relative importance may shift across spatial and temporal scales. Although disentangling the relative importance of these processes has been a continuing methodological challenge, recent developments in multi-scale spatial and temporal modeling can now help ecologists estimate their scale-specific contributions. Here we present a statistical approach to (1) detect the presence of a space-time interaction on community composition and (2) estimate the scale-specific importance of environmental and spatial factors on beta diversity. To illustrate the applicability of this approach, we use a case study from a temperate forest understory where tree seedling abundances were monitored during a 9-year period at 40 permanent plots. We found no significant space-time interaction on tree seedling composition, which means that the spatial abundance patterns did not vary over the study period. However, for a given year the relative importance of niche processes and other spatial processes was found to be scale-specific. Tree seedling abundances were primarily controlled by a broad-scale environmental gradient, but within the confines of this gradient the finer scale patchiness was largely due to other spatial processes. This case study illustrates that these two sets of processes are not mutually exclusive and can affect abundance patterns in a scale-dependent manner. More importantly, the use of our methodology for future empirical studies should help in the merging of niche and neutral perspectives on beta diversity, an obvious next step for community ecology.
Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail
Coexistence of the niche and neutral perspectives in community ecology
DOI:10.1890/0012-9658(2006)87[1399:cotnan]2.0.co;2
URL
PMID:16869414
[本文引用: 1]
The neutral theory for community structure and biodiversity is dependent on the assumption that species are equivalent to each other in all important ecological respects. We explore what this concept of equivalence means in ecological communities, how such species may arise evolutionarily, and how the possibility of ecological equivalents relates to previous ideas about niche differentiation. We also show that the co-occurrence of ecologically similar or equivalent species is not incompatible with niche theory as has been supposed, because niche relations can sometimes favor coexistence of similar species. We argue that both evolutionary and ecological processes operate to promote the introduction and to sustain the persistence of ecologically similar and in many cases nearly equivalent species embedded in highly structured food webs. Future work should focus on synthesizing niche and neutral perspectives rather than dichotomously debating whether neutral or niche models provide better explanations for community structure and biodiversity.
Neutral theory: a historical perspective
DOI:10.1111/j.1420-9101.2007.01410.x
URL
PMID:17956380
[本文引用: 1]
To resolve a panselectionist paradox, the population geneticist Kimura invented a neutral theory, where each gene is equally likely to enter the next generation whatever its allelic type. To learn what could be explained without invoking Darwinian adaptive divergence, Hubbell devised a similar neutral theory for forest ecology, assuming each tree is equally likely to reproduce whatever its species. In both theories, some predictions worked; neither theory proved universally true. Simple assumptions allow neutral theorists to treat many subjects still immune to more realistic theory. Ecologists exploit far fewer of these possibilities than population geneticists, focussing instead on species abundance distributions, where their predictions work best, but most closely match non-neutral predictions. Neutral theory cannot explain adaptive divergence or ecosystem function, which ecologists must understand. By addressing new topics and predicting changes in time, however, ecological neutral theory can provide probing null hypotheses and stimulate more realistic theory.
Demographic trade-offs in a neutral model explain death-rate-bundance-rank relationship
DOI:10.1890/07-2079.1
URL
PMID:19294910
[本文引用: 3]
The neutral theory of biodiversity has been criticized for its neglect of species differences. Yet it is much less heeded that S. P. Hubbell's definition of neutrality allows species to differ in their birth and death rates as long as they have an equal per capita fitness. Using the lottery model of competition we find that fitness equalization through birth-death trade-offs can make species coexist longer than expected for demographically identical species, whereas the probability of monodominance for a species under zero-sum neutral dynamics is equal to its initial relative abundance. Furthermore, if newly arising species in a community survive preferentially they are more likely to slip through the quagmire of rareness, thus creating a strong selective bias favoring their community membership. On the other hand, high-mortality species, once having gained a footing in the community, are more likely to become abundant due to their compensatory high birth rates. This unexpected result explains why a positive association between species abundance and per capita death rate can be seen in tropical-forest communities. An explicit incorporation of interspecific trade-offs between birth and death into the neutral theory increases the theory's realism as well as its predictive power.
A community perspective on the multidimensional niche
Partitioning beta diversity in a subtropical broad- leaved forest of China
DOI:10.1890/07-1880.1
URL
PMID:19341137
[本文引用: 2]
The classical environmental control model assumes that species distribution is determined by the spatial variation of underlying habitat conditions. This niche-based model has recently been challenged by the neutral theory of biodiversity which assumes that ecological drift is a key process regulating species coexistence. Understanding the mechanisms that maintain biodiversity in communities critically depends on our ability to decompose the variation of diversity into the contributions of different processes affecting it. Here we investigated the effects of pure habitat, pure spatial, and spatially structured habitat processes on the distributions of species richness and species composition in a recently established 24-ha stem-mapping plot in the subtropical evergreen broad-leaved forest of Gutianshan National Nature Reserve in East China. We used the new spatial analysis method of principal coordinates of neighbor matrices (PCNM) to disentangle the contributions of these processes. The results showed that (1) habitat and space jointly explained approximately 53% of the variation in richness and approximately 65% of the variation in species composition, depending on the scale (sampling unit size); (2) tree diversity (richness and composition) in the Gutianshan forest was dominantly controlled by spatially structured habitat (24%) and habitat-independent spatial component (29%); the spatially independent habitat contributed a negligible effect (6%); (3) distributions of richness and species composition were strongly affected by altitude and terrain convexity, while the effects of slope and aspect were weak; (4) the spatial distribution of diversity in the forest was dominated by broad-scaled spatial variation; (5) environmental control on the one hand and unexplained spatial variation on the other (unmeasured environmental variables and neutral processes) corresponded to spatial structures with different scales in the Gutianshan forest plot; and (6) five habitat types were recognized; a few species were statistically significant indicators of three of these habitats, whereas two habitats had no significant indicator species. The results suggest that the diversity of the forest is equally governed by environmental control (30%) and neutral processes (29%). In the fine-scale analysis (10 x 10 m cells), neutral processes dominated (43%) over environmental control (20%).
Coexistence and relative abundance in annual plant communities: the roles of competition and colonization
DOI:10.1086/342073
URL
PMID:18707522
[本文引用: 1]
Although an interspecific trade-off between competitive and colonizing ability can permit multispecies coexistence, whether this mechanism controls the structure of natural systems remains unresolved. We used models to evaluate the hypothesized importance of this trade-off for explaining coexistence and relative abundance patterns in annual plant assemblages. In a nonspatial model, empirically derived competition-colonization trade-offs related to seed mass were insufficient to generate coexistence. This was unchanged by spatial structure or interspecific variation in the fraction of seeds dispersing globally. These results differ from those of the more generalized competition-colonization models because the latter assume completely asymmetric competition, an assumption that appears unrealistic considering existing data for annual systems. When, for heuristic purposes, completely asymmetric competition was incorporated into our models, unlimited coexistence was possible. However, in the resulting abundance patterns, the best competitors/poorest colonizers were the most abundant, the opposite of that observed in natural systems. By contrast, these natural patterns were produced by competition-colonization models where environmental heterogeneity permitted species coexistence. Thus, despite the failure of the simple competition-colonization trade-off to explain coexistence in annual plant systems, this trade-off may be essential to explaining relative abundance patterns when other processes permit coexistence.
The limiting similarity, convergence, and divergence of coexisting species
Plant invasions and the niche
A test of the unified neutral theory of biodiversity
DOI:10.1038/nature01583
URL
PMID:12692564
[本文引用: 2]
One of the fundamental questions of ecology is what controls biodiversity. Recent theory suggests that biodiversity is controlled predominantly by neutral drift of species abundances. This theory has generated considerable controversy, because it claims that many mechanisms that have long been studied by ecologists (such as niches) have little involvement in structuring communities. The theory predicts that the species abundance distribution within a community should follow a zero-sum multinomial distribution (ZSM), but this has not, so far, been rigorously tested. Specifically, it remains to be shown that the ZSM fits the data significantly better than reasonable null models. Here I test whether the ZSM fits several empirical data sets better than the lognormal distribution. It does not. Not only does the ZSM fail to fit empirical data better than the lognormal distribution 95% of the time, it also fails to fit empirical data better even a majority of the time. This means that there is no evidence that the ZSM predicts abundances better than the much more parsimonious null hypothesis.
Empirical evaluation of neutral theory
DOI:10.1890/0012-9658(2006)87[1411:eeont]2.0.co;2
URL
PMID:16869415
[本文引用: 6]
We describe a general framework for testing neutral theory. We summarize similarities and differences between ten different versions of neutral theory. Two central predictions of neutral theory are that species abundance distributions will follow a zero-sum multinomial distribution and that community composition will change over space due to dispersal limitation. We review all published empirical tests of neutral theory. With the exception of one type of test, all tests fail to support neutral theory. We identify and perform several new tests. Specifically, we develop a set of best practices for testing the fit of the zero-sum multinomial (ZSM) vs. a lognormal null hypothesis and apply this to a data set, concluding that the lognormal outperforms neutral theory on robust tests. We explore whether a priori parameterization of neutral theory is possible, and we conclude that it is not. We show that non-curve-fitting predictions readily derived from neutral theory are easily falsifiable. In toto, there is a current overwhelming weight of evidence against neutral theory. We suggest some next steps for neutral theory.
Species abundance distributions: moving beyond single prediction theories to integration within an ecological frame-work
DOI:10.1111/j.1461-0248.2007.01094.x
URL
PMID:17845298
[本文引用: 2]
Species abundance distributions (SADs) follow one of ecology's oldest and most universal laws--every community shows a hollow curve or hyperbolic shape on a histogram with many rare species and just a few common species. Here, we review theoretical, empirical and statistical developments in the study of SADs. Several key points emerge. (i) Literally dozens of models have been proposed to explain the hollow curve. Unfortunately, very few models are ever rejected, primarily because few theories make any predictions beyond the hollow-curve SAD itself. (ii) Interesting work has been performed both empirically and theoretically, which goes beyond the hollow-curve prediction to provide a rich variety of information about how SADs behave. These include the study of SADs along environmental gradients and theories that integrate SADs with other biodiversity patterns. Central to this body of work is an effort to move beyond treating the SAD in isolation and to integrate the SAD into its ecological context to enable making many predictions. (iii) Moving forward will entail understanding how sampling and scale affect SADs and developing statistical tools for describing and comparing SADs. We are optimistic that SADs can provide significant insights into basic and applied ecological science.
Analytic solution of Hubbell’s model of local community dynamics
DOI:10.1016/j.tpb.2003.08.001
URL
PMID:14642345
[本文引用: 1]
Recent theoretical approaches to community structure and dynamics reveal that many large-scale features of community structure (such as species-rank distributions and species-area relations) can be explained by a so-called neutral model. Using this approach, species are taken to be equivalent and trophic relations are not taken into account explicitly. Here we provide a general analytic solution to the local community model of Hubbell's neutral theory of biodiversity by recasting it as an urn model, i.e. a Markovian description of states and their transitions. Both stationary and time-dependent distributions are analysed. The stationary distribution-also called the zero-sum multinomial-is given in closed form. An approximate form for the time-dependence is obtained by using an expansion of the master equation. The temporal evolution of the approximate distribution is shown to be a good representation for the true temporal evolution for a large range of parameter values.
Immigration and local competition in herbaceous plant communities: a three-year seed-sowing experiment
Neutral metacommunity models predict fish diversity patterns in Mississippi-Missouri basin
DOI:10.1038/nature06813
URL
PMID:18464742
[本文引用: 1]
River networks, seen as ecological corridors featuring connected and hierarchical dendritic landscapes for animals and plants, present unique challenges and opportunities for testing biogeographical theories and macroecological laws. Although local and basin-scale differences in riverine fish diversity have been analysed as functions of energy availability and habitat heterogeneity, scale-dependent environmental conditions and river discharge, a model that predicts a comprehensive set of system-wide diversity patterns has been hard to find. Here we show that fish diversity patterns throughout the Mississippi-Missouri River System are well described by a neutral metacommunity model coupled with an appropriate habitat capacity distribution and dispersal kernel. River network structure acts as an effective template for characterizing spatial attributes of fish biodiversity. We show that estimates of average dispersal behaviour and habitat capacities, objectively calculated from average runoff production, yield reliable predictions of large-scale spatial biodiversity patterns in riverine systems. The success of the neutral theory in two-dimensional forest ecosystems and here in dendritic riverine ecosystems suggests the possible application of neutral metacommunity models in a diverse suite of ecosystems. This framework offers direct linkage from large-scale forcing, such as global climate change, to biodiversity patterns.
Heteromyopia and the spatial coexistence of similar competitors
The role of biomass allocation strategy on diversity loss due to fertilization
The nearly neutral theory of molecular evolution
Dynamic response of Permian brachiopod communities to long-term environmental change
DOI:10.1038/nature02464
URL
PMID:15085129
[本文引用: 1]
The fossil record preserves numerous natural experiments that can shed light on the response of ecological communities to environmental change. However, directly observing the community dynamics of extinct organisms is not possible. As an alternative, neutral ecological models suggest that species abundance distributions reflect dynamical processes like migration, competition, recruitment, and extinction. Live-dead comparisons suggest that such distributions can be faithfully preserved in the rock record. Here we use a maximum-likelihood approach to show that brachiopod (lamp shell) abundance distributions from four temporally distinct ecological landscapes from the Glass Mountains, Texas (of the Permian period), exhibit significant differences. Further, all four are better fitted by zero-sum multinomial distributions, characteristic of Hubbell's neutral model, than by log-normal distributions, as predicted by the traditional ecological null hypothesis. Using the neutral model as a guide, we suggest that sea level fluctuations spanning about 10 Myr altered the degrees of isolation and exchange among local communities within these ecological landscapes. Neither these long-term environmental changes nor higher-frequency sea level fluctuations resulted in wholesale extinction or major innovation within evolutionary lineages.
Species-area relationships from a spatially explicit neutral model in an infinite landscape
DOI:10.1111/j.1461-0248.2007.01050.x
URL
PMID:17542937
[本文引用: 1]
We use recently developed technical methods to study species-area relationships from a spatially explicit extension of Hubbell's neutral model on an infinite landscape. Our model includes variable dispersal distances and exhibits qualitatively different behaviour from the cases of nearest-neighbour dispersal and finite periodic landscapes that have previously been studied. We show that different dispersal distances and even different dispersal kernels produce identical species-area curves up to rescaling of the two axes. This scaling property provides a straightforward method for fitting the model to empirical data. The species-area curves display all three phases observed empirically and enable the exponent describing the power law relationship for species-area curves to be identified as the gradient at the central phase. This exponent can take all values between 0 and 1 and is given by a simple function of the speciation rate, independent of all other model variables.
Community ecology theory as a framework for biological invasions
DOI:10.1016/S0169-5347(02)02495-3 URL [本文引用: 1]
Plant coexistence and the niche
Species turnover and equilibrium island biogeography
DOI:10.1126/science.194.4265.572 URL PMID:17818409
Natural variability and the manifold mechanisms of ecological communities
The use and abuse of vegetational concepts and terms
Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion and community assembly
Dispersal, environment, and floristic variation of western Amazonian forests
DOI:10.1126/science.1078037
URL
PMID:12522248
[本文引用: 1]
The distribution of plant species, the species compositions of different sites, and the factors that affect them in tropical rain forests are not well understood. The main hypotheses are that species composition is either (i) uniform over large areas, (ii) random but spatially autocorrelated because of dispersal limitation, or (iii) patchy and environmentally determined. Here we test these hypotheses, using a large data set from western Amazonia. The uniformity hypothesis gains no support, but the other hypotheses do. Environmental determinism explains a larger proportion of the variation in floristic differences between sites than does dispersal limitation; together, these processes explain 70 to 75% of the variation. Consequently, it is important that management planning for conservation and resource use take into account both habitat heterogeneity and biogeographic differences.
Analytical solution of a neutral model of biodiversity
Density dependence explains tree species abundance and diversity in tropical forests
DOI:10.1038/nature04030
URL
PMID:16319890
[本文引用: 1]
The recurrent patterns in the commonness and rarity of species in ecological communities--the relative species abundance--have puzzled ecologists for more than half a century. Here we show that the framework of the current neutral theory in ecology can easily be generalized to incorporate symmetric density dependence. We can calculate precisely the strength of the rare-species advantage that is needed to explain a given RSA distribution. Previously, we demonstrated that a mechanism of dispersal limitation also fits RSA data well. Here we compare fits of the dispersal and density-dependence mechanisms for empirical RSA data on tree species in six New and Old World tropical forests and show that both mechanisms offer sufficient and independent explanations. We suggest that RSA data cannot by themselves be used to discriminate among these explanations of RSA patterns--empirical studies will be required to determine whether RSA patterns are due to one or the other mechanism, or to some combination of both.
Neutral theory and relative species abundance in ecology
DOI:10.1038/nature01883
URL
PMID:12944964
[本文引用: 4]
The theory of island biogeography asserts that an island or a local community approaches an equilibrium species richness as a result of the interplay between the immigration of species from the much larger metacommunity source area and local extinction of species on the island (local community). Hubbell generalized this neutral theory to explore the expected steady-state distribution of relative species abundance (RSA) in the local community under restricted immigration. Here we present a theoretical framework for the unified neutral theory of biodiversity and an analytical solution for the distribution of the RSA both in the metacommunity (Fisher's log series) and in the local community, where there are fewer rare species. Rare species are more extinction-prone, and once they go locally extinct, they take longer to re-immigrate than do common species. Contrary to recent assertions, we show that the analytical solution provides a better fit, with fewer free parameters, to the RSA distribution of tree species on Barro Colorado Island, Panama, than the lognormal distribution.
Patterns of relative species abundance in rainforests and coral reefs
DOI:10.1038/nature06197
URL
PMID:17972874
[本文引用: 3]
A formidable many-body problem in ecology is to understand the complex of factors controlling patterns of relative species abundance (RSA) in communities of interacting species. Unlike many problems in physics, the nature of the interactions in ecological communities is not completely known. Although most contemporary theories in ecology start with the basic premise that species interact, here we show that a theory in which all interspecific interactions are turned off leads to analytical results that are in agreement with RSA data from tropical forests and coral reefs. The assumption of non-interacting species leads to a sampling theory for the RSA that yields a simple approximation at large scales to the exact theory. Our results show that one can make significant theoretical progress in ecology by assuming that the effective interactions among species are weak in the stationary states in species-rich communities such as tropical forests and coral reefs.
Phylogenies and community ecology
Limitations to species coexistence: evidence for competition from field observations, using a patch model
Field parameterization and experimental test of the neutral theory of biodiversity
DOI:10.1038/nature03211
URL
PMID:15662423
[本文引用: 1]
Ecologists would like to explain general patterns observed across multi-species communities, such as species-area and abundance-frequency relationships, in terms of the fundamental processes of birth, death and migration underlying the dynamics of all constituent species. The unified neutral theory of biodiversity and related theories based on these fundamental population processes have successfully recreated general species-abundance patterns without accounting for either the variation among species and individuals or resource-releasing processes such as predation and disturbance, long emphasized in ecological theory. If ecological communities can be described adequately without estimating variation in species and their interactions, our understanding of ecological community organization and the predicted consequences of reduced biodiversity and environmental change would shift markedly. Here, I introduce a strong method to test the neutral theory that combines field parameterization of the underlying population dynamics with a field experiment, and apply it to a rocky intertidal community. Although the observed abundance-frequency distribution of the system follows that predicted by the neutral theory, the neutral theory predicts poorly the field experimental results, indicating an essential role for variation in species interactions.
Relative resource abundance explains butterfly biodiversity in island communities
Can high tree species richness be explained by Hubbell's null model?
The effects of competitive asymmetry on the rate of competitive displacement: How robust is Hubbell’s community drift model?
A nearly neutral model of biodiversity
DOI:10.1890/06-1817.1
URL
PMID:18376566
[本文引用: 3]
S. P. Hubbell's unified neutral theory of biodiversity has stimulated much new thinking about biodiversity. However, empirical support for the neutral theory is limited, and several observations are inconsistent with the predictions of the theory, including positive correlations between traits associated with competitive ability and species abundance and correlations between species diversity and ecosystem functioning. The neutral theory can be extended to explain these observations by allowing species to differ slightly in their competitive ability (fitness). Here, we show that even slight differences in fecundity can greatly reduce the time to extinction of competitors even when the community size is large and dispersal is spatially limited. In this case, species richness is dramatically reduced, and a markedly different species abundance distribution is predicted than under pure neutrality. In the nearly neutral model, species co-occur in the same community not because of, but in spite of, ecological differences. The more competitive species with higher fecundity tend to have higher abundance both in the metacommunity and in local communities. The nearly neutral perspective provides a theoretical framework that unites the sampling model of the neutral theory with theory of biodiversity affecting ecosystem function.
The impact of neutrality, niche differentiation and species input on diversity and abundance distributions,
Plant coexistence in the interactive environment: arbuscular mycorrhiza should not be out of mind

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}