
Biodiv Sci ›› 2008, Vol. 16 ›› Issue (6): 570-577. DOI: 10.3724/SP.J.1003.2008.08049 cstr: 32101.14.SP.J.1003.2008.08049
• Original Papers • Previous Articles Next Articles
Yangying Gan1,2, Shaohua Li1, Songquan Song1, Weiqing Wang1, Hongyan Cheng1,*( )
)
Received:2008-03-04
Accepted:2008-10-31
Online:2008-11-20
Published:2008-11-20
Contact:
Hongyan Cheng
About author:* E-mail: hycheng@ibcas.ac.cnYangying Gan, Shaohua Li, Songquan Song, Weiqing Wang, Hongyan Cheng. Seed dormancy and release of grapes from different proveniences[J]. Biodiv Sci, 2008, 16(6): 570-577.
| 种类 Category | 选用品种 Variety | 特性 Remarks |
|---|---|---|
| 东亚种 (S) East-Asia species | '通化一号' Tonghuayi '双优' Shuangyou '左山一' Zuoshanyi '左山二' Zuoshaner '双红' Shuanghong '双丰' Shuangfeng | 均为中国原生种山葡萄的选育品种, 产于中国东北、华北及华东部分地区, 分布海拔为200-2,100 m, 在葡萄属中抗寒性最强 (耐受-40℃) All of these varieties are selected from wild grape of V. amurensis, a Chinese origin species, distributed at altitudes of 200-2,100 m in northeast of China, northern China and parts of eastern China, the most cold-resistant species of grape, able to survive at -40℃. |
| 美洲种 (M) American species | '康可' Concord '贝达' Beta | 美洲葡萄实生苗选育品种 Selected from seedlings of V. labrusca, a wild American species 美洲种杂种 A cross-breed within American species (V. vulpine×V. labrusca) |
| 欧亚种 (O) Europe-Asia species | '京秀' Jingxiu '京玉' Jingyu '京蜜' Jingmi '玫瑰香' Muscat Hamburg '96-10' '意大利' Italia | 均为欧洲葡萄的选育品种。'京秀'、'京玉'、'京蜜'为早熟品种, '玫瑰香'为中熟品种, '96-10'、'意大利'为晚熟品种。 All of these varieties are selected from V.vinifera, the only existing species of Europe-Asia origin. Jingxiu, Jingyu, and Jingmi are early-maturing. Muscat Hamburg is medium-maturing, and 96-10 and Italia are late-maturing. |
| 欧山杂种 (O×S) Cross-breed of Europe-Asia species × East-Asia species | '北醇' Beichun '北红' Beihong | 欧洲葡萄与东亚种山葡萄的杂交后代 Cross-breed of Europe-Asia species × East-Asia species |
| 欧美杂种 (O×M) Cross-breed of Europe-Asia species × American species | '京亚' Jingya '高妻' Takqtsuma | 欧洲葡萄和美洲葡萄的杂交后代 Cross-breed of Europe-Asia species × American species |
| 蘡欧杂种 (Y×O) Cross-breed of East-Asia species×Europe-Asia species | '北紫' Beizi '北丰' Beifeng | 东亚种蘡薁葡萄和欧洲葡萄的杂交后代。蘡薁葡萄 (V. thunbergii) 为中国原生种, 产于华北、华中、华东、西南和华南地区, 分布于海拔150-2,500 m, 是中国分布最广的葡萄种。 Cross-breed of East-Asia species×Europe-Asia species.V. thunbergiiis a wild grape species of Chinese origin, most widely distributed in China. It distributes at altitudes of 150-2,500 m in northern China, central China, eastern China, southwest of China, and southern China. |
Table 1 Description of the experimental materials
| 种类 Category | 选用品种 Variety | 特性 Remarks |
|---|---|---|
| 东亚种 (S) East-Asia species | '通化一号' Tonghuayi '双优' Shuangyou '左山一' Zuoshanyi '左山二' Zuoshaner '双红' Shuanghong '双丰' Shuangfeng | 均为中国原生种山葡萄的选育品种, 产于中国东北、华北及华东部分地区, 分布海拔为200-2,100 m, 在葡萄属中抗寒性最强 (耐受-40℃) All of these varieties are selected from wild grape of V. amurensis, a Chinese origin species, distributed at altitudes of 200-2,100 m in northeast of China, northern China and parts of eastern China, the most cold-resistant species of grape, able to survive at -40℃. |
| 美洲种 (M) American species | '康可' Concord '贝达' Beta | 美洲葡萄实生苗选育品种 Selected from seedlings of V. labrusca, a wild American species 美洲种杂种 A cross-breed within American species (V. vulpine×V. labrusca) |
| 欧亚种 (O) Europe-Asia species | '京秀' Jingxiu '京玉' Jingyu '京蜜' Jingmi '玫瑰香' Muscat Hamburg '96-10' '意大利' Italia | 均为欧洲葡萄的选育品种。'京秀'、'京玉'、'京蜜'为早熟品种, '玫瑰香'为中熟品种, '96-10'、'意大利'为晚熟品种。 All of these varieties are selected from V.vinifera, the only existing species of Europe-Asia origin. Jingxiu, Jingyu, and Jingmi are early-maturing. Muscat Hamburg is medium-maturing, and 96-10 and Italia are late-maturing. |
| 欧山杂种 (O×S) Cross-breed of Europe-Asia species × East-Asia species | '北醇' Beichun '北红' Beihong | 欧洲葡萄与东亚种山葡萄的杂交后代 Cross-breed of Europe-Asia species × East-Asia species |
| 欧美杂种 (O×M) Cross-breed of Europe-Asia species × American species | '京亚' Jingya '高妻' Takqtsuma | 欧洲葡萄和美洲葡萄的杂交后代 Cross-breed of Europe-Asia species × American species |
| 蘡欧杂种 (Y×O) Cross-breed of East-Asia species×Europe-Asia species | '北紫' Beizi '北丰' Beifeng | 东亚种蘡薁葡萄和欧洲葡萄的杂交后代。蘡薁葡萄 (V. thunbergii) 为中国原生种, 产于华北、华中、华东、西南和华南地区, 分布于海拔150-2,500 m, 是中国分布最广的葡萄种。 Cross-breed of East-Asia species×Europe-Asia species.V. thunbergiiis a wild grape species of Chinese origin, most widely distributed in China. It distributes at altitudes of 150-2,500 m in northern China, central China, eastern China, southwest of China, and southern China. |
| 种类 Category | 品种 Variety | 千粒重 1000-seed weight (g) | 最初萌发率 Initial germination (%) |
|---|---|---|---|
| 东亚种 (S) | '双红' Shuanghong | 27.6d | 0 |
| '通化一号' Tonghuayi | 33.7c | 0 | |
| '双优' Shuangyou | 27.3d | 0 | |
| '左山一' Zuoshanyi | 36.7c | 0 | |
| '左山二' Zuoshaner | 28.9d | 0 | |
| '双丰' Shuangfeng | 28.7d | 0 | |
| 美洲种 (M) | '康可' Concord | 52.3b | 0 |
| '贝达' Beta | 38.6c | 0 | |
| 欧亚种 (O) | '京秀' Jingxiu | 56.8b | 46.3±6.0a |
| '京玉' Jingyu | 32.9d | 0 | |
| '玫瑰香' Muscat Hamburg | 40.9c | 0 | |
| '96-10' | 57.9b | 0 | |
| '意大利' Italia | 41.5c | 0 | |
| 欧山杂种 (O×S) | '北醇' Beichun | 41.9c | 16.7±2.6b |
| '北红' Beihong | 42.3c | 14.3±1.6b | |
| 欧美杂种 (O×M) | '京亚' Jingya | 90.0a | 0 |
| '高妻' Takqtsuma | 103.8a | 0 | |
| 蘡欧杂种 (Y×O) | '北紫' Beizi | 34.9c | 16±4b |
| '北丰' Beifeng | 38.4c | 20±6.9b |
Table 2 1000-seed weight and initial germination percentage of mature seeds of grape varieties from different proveniences
| 种类 Category | 品种 Variety | 千粒重 1000-seed weight (g) | 最初萌发率 Initial germination (%) |
|---|---|---|---|
| 东亚种 (S) | '双红' Shuanghong | 27.6d | 0 |
| '通化一号' Tonghuayi | 33.7c | 0 | |
| '双优' Shuangyou | 27.3d | 0 | |
| '左山一' Zuoshanyi | 36.7c | 0 | |
| '左山二' Zuoshaner | 28.9d | 0 | |
| '双丰' Shuangfeng | 28.7d | 0 | |
| 美洲种 (M) | '康可' Concord | 52.3b | 0 |
| '贝达' Beta | 38.6c | 0 | |
| 欧亚种 (O) | '京秀' Jingxiu | 56.8b | 46.3±6.0a |
| '京玉' Jingyu | 32.9d | 0 | |
| '玫瑰香' Muscat Hamburg | 40.9c | 0 | |
| '96-10' | 57.9b | 0 | |
| '意大利' Italia | 41.5c | 0 | |
| 欧山杂种 (O×S) | '北醇' Beichun | 41.9c | 16.7±2.6b |
| '北红' Beihong | 42.3c | 14.3±1.6b | |
| 欧美杂种 (O×M) | '京亚' Jingya | 90.0a | 0 |
| '高妻' Takqtsuma | 103.8a | 0 | |
| 蘡欧杂种 (Y×O) | '北紫' Beizi | 34.9c | 16±4b |
| '北丰' Beifeng | 38.4c | 20±6.9b |
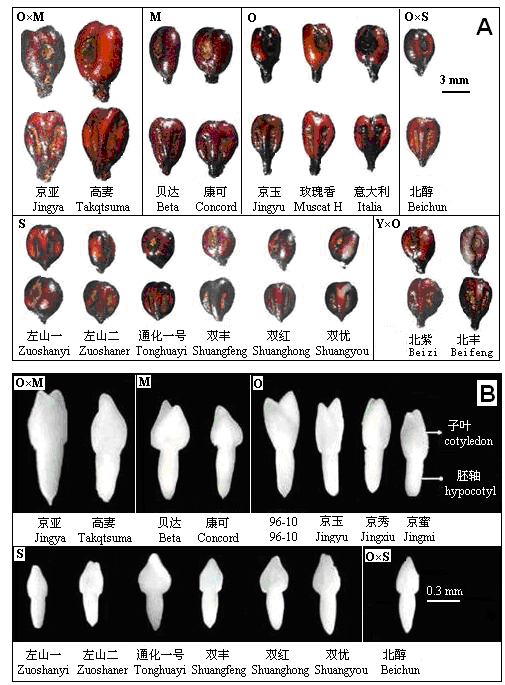
Fig. 1 Morphological appearance of mature seeds of grape varieties from different proveniences. A, Hilum side (above line) and back side (below line) of seeds; B, Embryos.
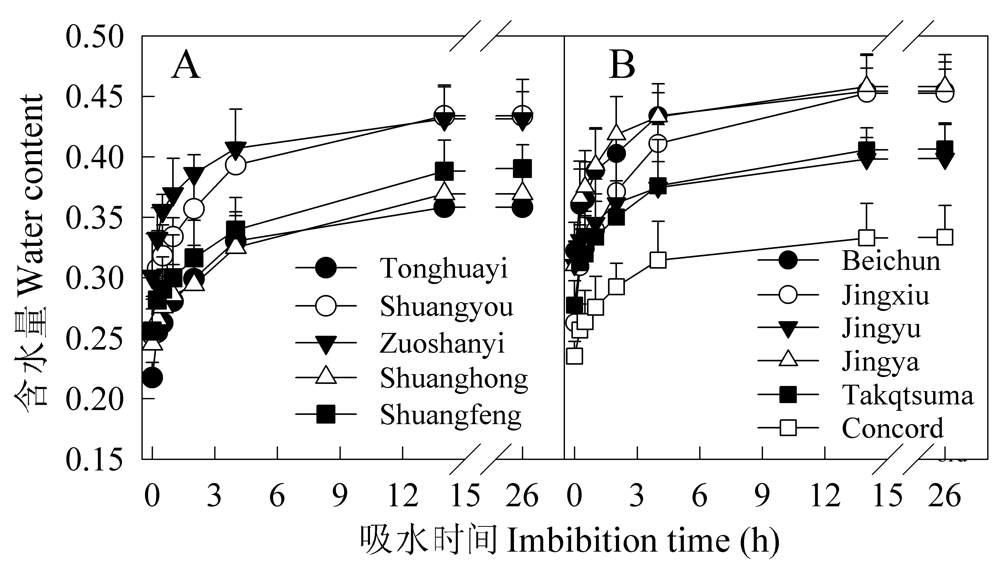
Fig. 2 Changes in seed water contents (g H2O g-1 DW) during imbibition of grape varieties from different proveniences. A, Varieties of East-Asia species (S); B, Beichun (O×S), Jingxiu and Jingyu (O), Jingya (O×M) and Concord (M).
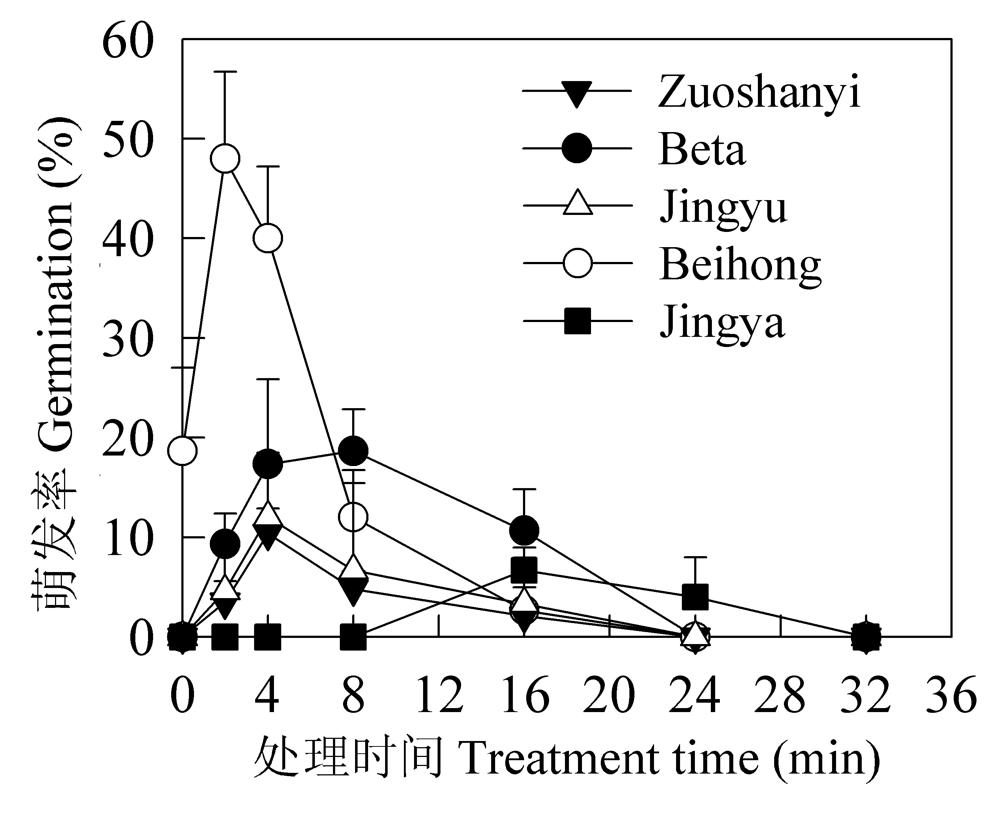
Fig. 3 Effects of sulfuric acid treatment on germination of grape seeds. Beta (M); Beihong (O×S); Zuoshanyi (S); Jingyu (O); Jingya (O×M).
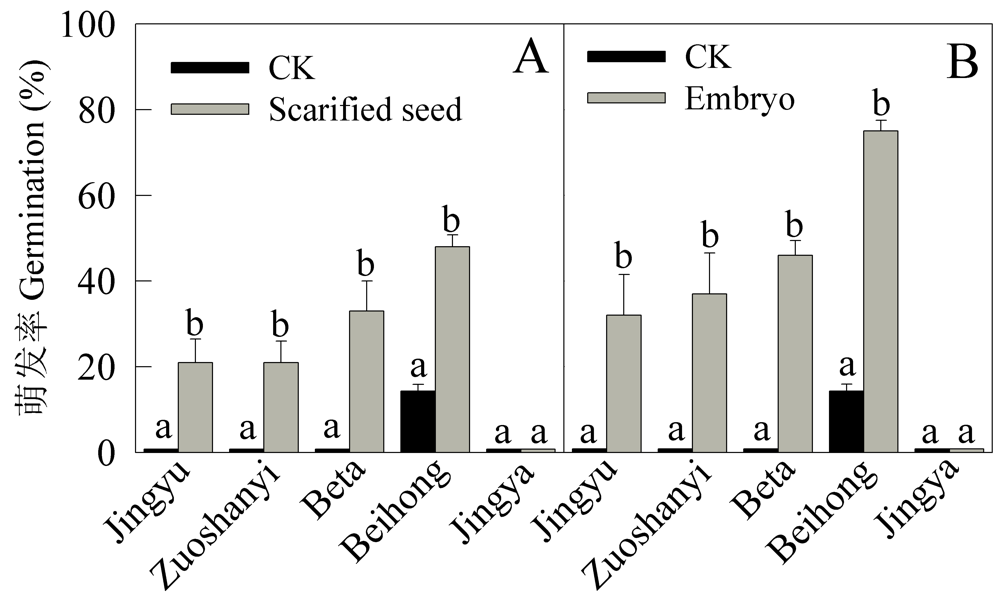
Fig. 4 Effects of removal of seed coat surrounding embryo on seed germination and germination of excised embryos. A, Scarified seeds (removed seed coat surrounding embryo) germinated on moistened filter paper; B, Excised embryos germinated on MS medium (no exogenous phytohormone). Different letters show significant difference at P = 0.05 level.

Fig. 5 Stratification effects at different temperatures on seed germination of grape varieties from different proveniences. (A) Varieties of East-Asia species; (B) Europe-Asia species; (C) Varieties of American species, cross-breeds of Europe-Asia species × East-Asia species, East-Asia species × Europe-Asia species, Europe-Asia species × American species; A, B, C: stratified for 2 months at 5°C, 25°C, 20/10°C and 30/20°C. (D) Varieties of American species and Europe-Asia species × American species, stratified for 6 months at 5°C and 20/10°C.
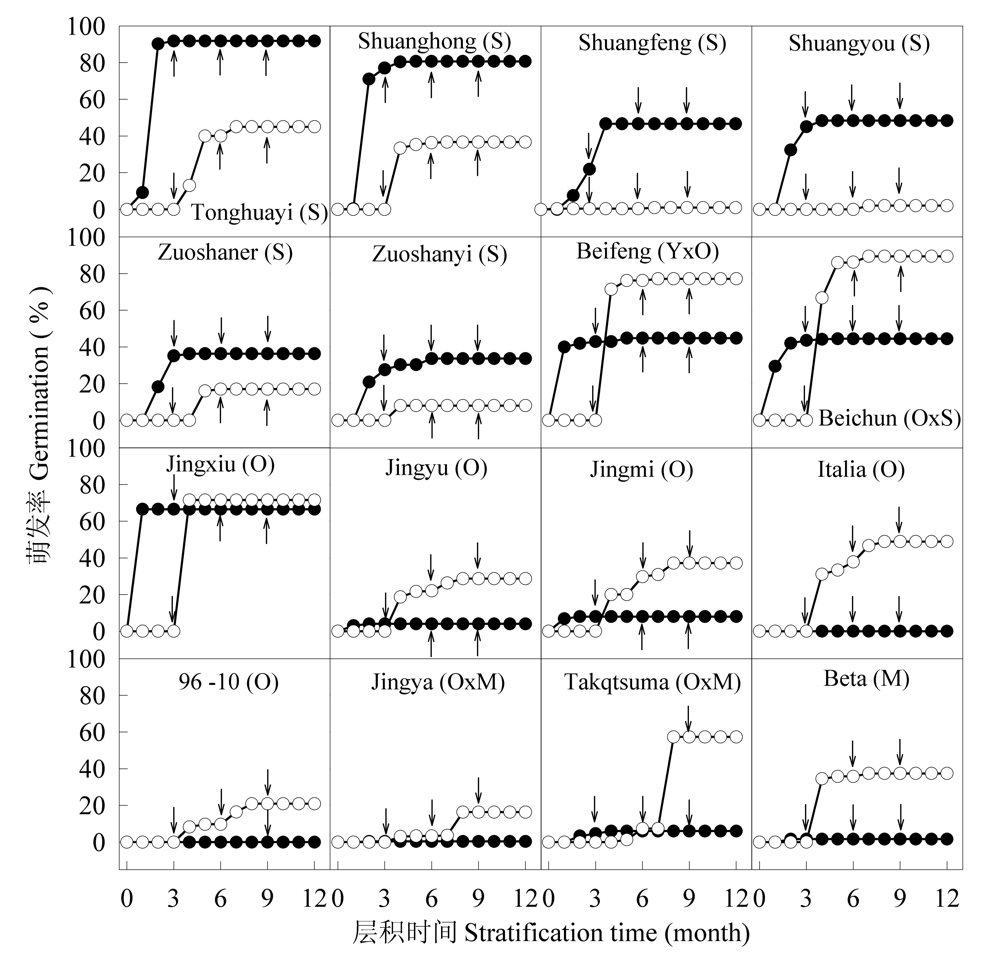
Fig. 6 Germination course of different species of grape seeds at alternate temperatures simulating the season changes. Cycle 1(●): Summer (3 months, 30/20°C)-autumn (3 months, 20/10°C)-winter (3 months, 5°C)-spring (3 months, 20/10°C); Cycle 2 (○): Winter (3 months, 5°C)-spring (3 months, 20/10°C)-summer (3 months, 30/20°C)-autumn (3 months, 20/10°C). Arrows indicate the simulating season changes at which seeds were moved from one temperature regime to the next.
| [1] | Alleweldt G, Speigel-Roy P, Reisch B (1990) Grapes (Vitis) genetic resources of temperate fruit and nut crops. Acta Horticulturae, 273,291-327. |
| [2] |
Baskin JM, Baskin CC (2004) A classification system for seed dormancy. Seed Science Research, 14,1-16.
DOI URL |
| [3] | Ellis RH, Hong D, Roberts EH (1983) A note on the development of a practical procedure for promoting the germination of dormant seed of grape (Vitis spp.). Vitis, 22,211-219. |
| [4] |
Fernandez GE, Clark JR, Moore JN (1991) Effect of seed coat manipulation on the germination of stenospermocarpic grape embryos cultured in ovule. HortScience, 26,1220.
DOI URL |
| [5] | He PC (贺普超) (1999) Taxonomy and resources of grapes (Vitis). In: Vitis (葡萄学)(ed. He PC (贺普超)), pp.8-10. China Agriculture Press, Beijing. (in Chinese) |
| [6] |
Jaillon O, Aury JM, Noel B, Policriti A, Clepet C, Casagrande A, Choisne N, Aubourg S, Vitulo N, Jubin C, Vezzi A, Legeai F, Hugueney P, Dasilva C, Horner D, Mica E, Jublot D, Poulain J, Bruyère C, Billault A, Segurens B, Gouyvenoux M, Ugarte E, Cattonaro F, Anthouard V, Vico V, Del Fabbro C, Alaux M, Di Gaspero G, Dumas V, Felice N, Paillard S, Juman I, Moroldo M, Scalabrin S, Canaguier A, Le Clainche I, Malacrida G, Durand E, Pesole G, Laucou V, Chatelet P, Merdinoglu D, Delledonne M, Pezzotti M, Lecharny A, Scarpelli C, Artiguenave F, Pè ME, Valle G, Morgante M, Caboche M, Adam-Blondon AF, Weissenbach J, Quétier F, Wincker P (2007) The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature, 449,463-467.
DOI URL PMID |
| [7] | Kong QS (孔庆山) (2004) Chinese Vitis (中国葡萄志), pp.16-18. China Agricultural Science and Technology Press, Beijing. (in Chinese) |
| [8] | Kucera B, Cohn MA, Leubner-Metzger G (2005) Plant hormone interaction during seed dormancy release and germination. Seed Science Research, 15,281-307. |
| [9] | Mao BR (毛伯韧), Wu ZS (吴兆苏) (1983) Study on the inheritance and mechanism of seed dormancy in wheat. Scientia Agricultura Sinica (中国农业科学), 16(6),53-60. (in Chinese with English abstract) |
| [10] | Nikolaeva MG (2004) On criteria to use in studies of seed evolution. Seed Science Research, 14,315-320. |
| [11] |
Rajasekaran K, Vine J, Mullins MG (1982) Dormancy in somatic embryos and seeds of Vitis: changes in endogenous abscisic acid during embryogeny and germination. Planta, 154,139-144.
URL PMID |
| [12] | Selim HH, Ibrahim FA, Fayek MA, Sari EI-Deen SA, Gama NM (1981) Effect of different treatments on germination of Romi red grape seeds. Vitis, 20,115-121. |
| [13] | Spiegel-Roy P, Shulman Y, Baron I, Ashbel E (1987) Effect of cyanamide in overcoming grape seed dormancy. HortScience, 22,208-210. |
| [14] | Thompson K, Ceriani RM, Bakker JP, Bekker RM (2003) Are seed dormancy and persistence in soil related? Seed Science Research, 13,97-100. |
| [15] | Zheng GH (郑光华) (2004) Seed Physiology Research (种子生理研究), pp.321-322. Science Press, Beijing. (in Chinese) |
| [1] | Ping Xie. How did nucleus and sexual reproduction come into being? [J]. Biodiv Sci, 2016, 24(8): 966-976. |
| [2] | Caroline A. Polgar, Richard B. Primack. Leaf out phenology in temperate forests [J]. Biodiv Sci, 2013, 21(1): 111-116. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
Copyright © 2026 Biodiversity Science
Editorial Office of Biodiversity Science, 20 Nanxincun, Xiangshan, Beijing 100093, China
Tel: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn