
|
|
||
|
基于基因组重测序揭示栽培欧菱遗传多样性及‘南湖菱’的起源驯化历史
生物多样性
2024, 32 (9):
24212-.
DOI: 10.17520/biods.2024212
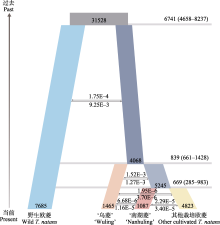
阐明栽培作物的起源驯化历史, 深入解析作物驯化改良的过程对于理解人类文明的进程至关重要。欧菱(Trapa natans)隶属于千屈菜科, 为一年生水生浮叶草本, 其果实俗称菱角, 是我国传统的水生蔬菜作物。然而, 相较于主粮作物, 我们对栽培欧菱种质资源的遗传多样性和起源驯化历史仍知之甚少。本研究基于31份不同品种(系)的二倍体栽培欧菱和23份二倍体野生欧菱的全基因组重测序(resequencing)数据, 利用群体基因组学分析方法, 对栽培欧菱的遗传多样性和我国特有的栽培品种‘南湖菱’的起源驯化历史进行研究。群体遗传结构与遗传分化的结果表明, 野生欧菱和栽培欧菱分属不同的基因池, 栽培欧菱起源于长江流域的二倍体野生欧菱。栽培欧菱遗传多样性水平整体低于野生欧菱(0.68 × 10−3 vs. 1.35 × 10−3), 有效群体大小相比野生欧菱也急剧收缩(~4,000 vs. ~7,700), 表明栽培欧菱经历了驯化瓶颈作用。栽培欧菱中, ‘南湖菱’的遗传多样性水平最低, 连锁不平衡的衰减速度最慢, 可能与其驯化程度高、选择强度大有关。基于FASTSIMCOAL2溯祖模拟的群体动态历史结果表明, ‘南湖菱’、‘乌菱’与其他栽培欧菱均发生了基因交流事件, ‘南湖菱’驯化发生在距今大约669年前, 这与明代文学记载信息相吻合。综上所述, 本研究通过探讨栽培欧菱的遗传多样性和‘南湖菱’的驯化历史, 对深入理解栽培欧菱驯化起源过程及其遗传演化特征具有重要意义。 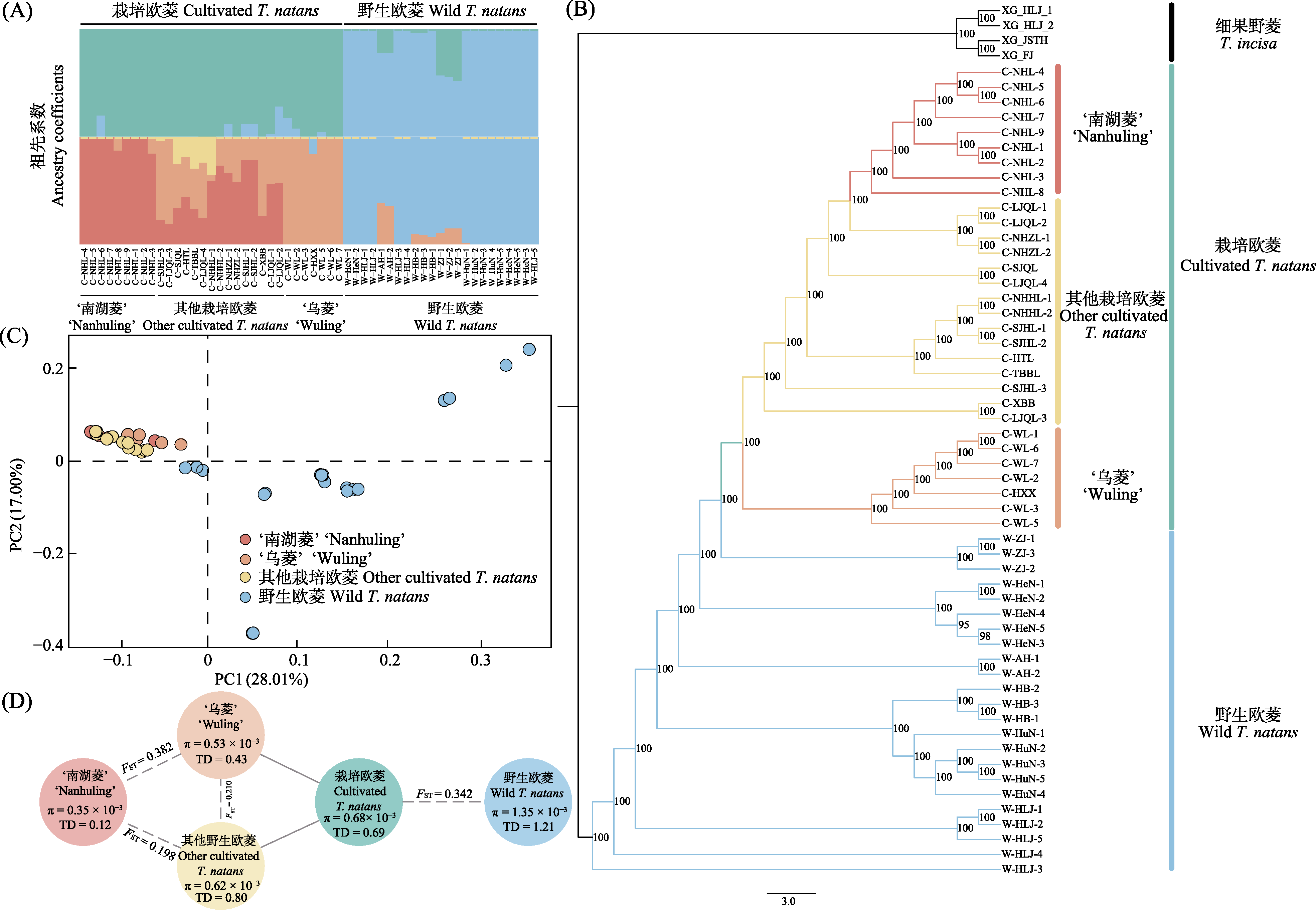 View image in article
图2
栽培欧菱和野生欧菱的群体遗传结构和遗传分化。A: 基于ADMIXTURE的群体遗传结构; B: 最大似然系统发育树; C: 主成分分析(PCA); D: 野生欧菱和栽培欧菱的核苷酸多态性(π)、Tajima’s D (TD)和遗传分化指数(FST)。
正文中引用本图/表的段落
基于ADMIXTURE分析的遗传结构结果表明,当K = 2时, 栽培欧菱和野生欧菱分别来源于两个基因池(图2A)。主成分分析也进一步支持了该结果, 在第一主成分(PC1)轴上将栽培欧菱和野生欧菱分开, PC1轴的解释比例为28.01% (图2C)。当K = 4时, 栽培欧菱内部进一步分为‘南湖菱’和‘乌菱’, 而其他栽培欧菱则包含多个遗传组分。通过IQ-TREE构建的系统发育树的结果与基于ADMIXTURE和PCA鉴定的结果基本一致, 分支支持率均高于95%, 栽培欧菱和野生欧菱在进化树上得到了较好的划分(图2B)。其中, 野生欧菱位于进化树的基部位置。来自宁波东钱湖(长江流域)的二倍体野生欧菱为栽培欧菱的基部类群。栽培欧菱主要分化成3支, ‘乌菱’首先从栽培欧菱中分化, 单系的‘南湖菱’和其他栽培欧菱也得到了较好的划分, 分支支持率均为100%。
对栽培欧菱和野生欧菱遗传多样性分析的结果(图2D)表明, 野生欧菱的遗传多样性水平(π = 1.35 × 10?3)整体高于栽培欧菱(π = 0.68 × 10?3)。对栽培欧菱而言, ‘南湖菱’的遗传多样性水平(π = 0.35 × 10?3)和Tajima’s D值最低(TD = 0.12), 遗传多样性不到野生欧菱的一半; ‘乌菱’的遗传多样性水平高于‘南湖菱’ (0.53 × 10?3 vs. 0.35 × 10?3)。遗传分化结果表明, 栽培欧菱与野生欧菱之间的遗传分化约为0.34 (FST = 0.34)。栽培欧菱群体中, ‘乌菱’和‘南湖菱’的遗传分化最高(FST = 0.38), ‘南湖菱’与其他栽培欧菱的遗传分化最低(FST = 0.20)。
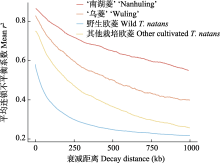
遗传分化的结果显示‘南湖菱’与其他栽培欧菱的遗传分化最低, 表明‘南湖菱’与其他栽培欧菱的亲缘关系可能较近。‘南湖菱’的驯化时间相对‘乌菱’较短(图4), 这可能导致其与其他栽培欧菱的分化略小于‘乌菱’与其他栽培欧菱。系统发育树上‘南湖菱’与其他栽培欧菱的拓扑结构进一步支持了该结果(图2B)。此外, 群体动态历史模拟检测到的‘南湖菱’与其他栽培欧菱的基因交流事件也进一步支持了二者较低的遗传分化水平。连锁不平衡的结果发现‘南湖菱’的连锁不平衡衰减速度最慢, 遗传多样性结果也表明其多样性水平最低, 可能与‘南湖菱’的驯化强度高、选择强度大有关(Doebley, 1989)。此外, 群体动态历史模拟的结果也表明‘南湖菱’的有效种群大小小于‘乌菱’, 进一步表明其高的驯化和选择强度。总之, 菱角自身的繁育系统和生活史特征导致其相比其他作物的遗传多样性水平较低, 而驯化瓶颈作用则导致菱角栽培群体的遗传多样性相比野生群体进一步降低。遗传多样性评估是自然资源与物种合理利用研究中必不可少的环节之一(Ellegren & Galtier, 2016)。本研究通过对菱角尤其是栽培欧菱的遗传多样性评估可为未来菱角的遗传选种与分子育种提供重要理论基础。
本文的其它图/表
|

{kind=link}