|
|
||||||||||||||||||||||||||||||
|
小檗科鬼臼亚科多样性格局的演化历史和成因
生物多样性
2023, 31 (7):
23100-.
DOI: 10.17520/biods.2023100
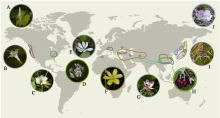
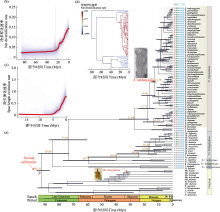
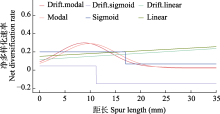
探究生物多样性在类群和空间不均一性的形成机制对于认识多样性的形成和演化规律具有重要意义, 也是生物学研究的热点之一。鬼臼亚科属于小檗科, 包含10属约89种, 属间物种数量差异巨大。本研究利用比较系统发育学的研究方法, 重建了鬼臼亚科的系统发育关系、分化时间和多样化历史, 并探讨了生物和非生物因素对净多样化速率的影响。研究结果表明鬼臼亚科起源于92.75 Myr, 净多样化速率在约23 Myr开始明显提升, 最显著的净多样化速率转变发生在21 Myr附近的淫羊藿属(Epimedium)分支上。基于性状的演化速率分析显示, 具有蜜距和分布在东亚亚热带的类群具有较高的净多样化速率, 但淫羊藿属特定的距长对其物种的净多样化速率没有影响; 果实类型、假种皮和气生茎的有无对类群间净多样化速率无影响。本研究表明功能性状的演化和中新世以来东亚地区季风气候的加强导致了鬼臼亚科内部多样性的分布不均。
表2
隐藏状态物种形成和灭绝分析使用的模型
正文中引用本图/表的段落
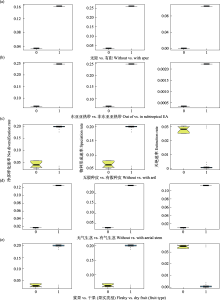
与常用的二元状态物种形成和灭绝(Binary State Speciation and Extinction, BiSSE)分析相比, 隐藏状态物种形成和灭绝(Hidden State Speciation and Extinction, HiSSE)分析可推断是否有隐藏性状影响类群演化速率以及各性状状态类群具体的演化速率(Beaulieu & O'Meara, 2016)。对于二态性状, 可以生成5个模型(表2), CID模型设定1个未知的二态性状; BiSSE模型设定1个已知二态性状作用于演化速率差异; CID-2模型设定1个未知二态性状作用于演化速率差异; CID-4模型设定2个未知二态性状作用于演化速率差异; FULL模型设定1个已知二态性状与2个未知二态性状共同作用于演化速率差异。结果可得不同模型的拟合度及各性状状态成种、灭绝和净多样化速率的后验分布, 据此判断是否有隐藏性状存在和可观测性状对演化速率差异化的作用程度。
其他3对二元状态的HiSSE分析结果中, 是否存在假种皮和果实类型的最优模型为可观测性状不发挥作用的CID-2模型, 且与次优模型的ΔAICc均大于2。表明不同性状状态物种演化速率的差异不能由这些可观测性状解释, 而是其他性状发挥了作用。是否存在气生茎的最优模型为BiSSE, 与次优模型CID-2的ΔAICc为2.47, 无法分辨气生茎存在是否促进了多样化速率。这3对性状在CID-2模型中的净多样化速率显示: 有气生茎 > 无气生茎, 干果 > 浆果, 有假种皮 > 无假种皮(图4)。
本文的其它图/表
|