
生物多样性 ›› 2023, Vol. 31 ›› Issue (7): 23100. DOI: 10.17520/biods.2023100 cstr: 32101.14.biods.2023100
陈馥艳1,2, 游旨价1,*( ), 张秋月1, 黄健1, 星耀武1,*()
), 张秋月1, 黄健1, 星耀武1,*()
收稿日期:2023-04-03
接受日期:2023-07-17
出版日期:2023-07-20
发布日期:2023-07-22
通讯作者:
* E-mail: 作者简介:ywxing@xtbg.org.cn基金资助:
Fuyan Chen1,2, Chih-Chieh Yu1,*(), Qiuyue Zhang1, Jian Huang1, Yaowu Xing1,*()
Received:2023-04-03
Accepted:2023-07-17
Online:2023-07-20
Published:2023-07-22
Contact:
* E-mail: 摘要:
探究生物多样性在类群和空间不均一性的形成机制对于认识多样性的形成和演化规律具有重要意义, 也是生物学研究的热点之一。鬼臼亚科属于小檗科, 包含10属约89种, 属间物种数量差异巨大。本研究利用比较系统发育学的研究方法, 重建了鬼臼亚科的系统发育关系、分化时间和多样化历史, 并探讨了生物和非生物因素对净多样化速率的影响。研究结果表明鬼臼亚科起源于92.75 Myr, 净多样化速率在约23 Myr开始明显提升, 最显著的净多样化速率转变发生在21 Myr附近的淫羊藿属(Epimedium)分支上。基于性状的演化速率分析显示, 具有蜜距和分布在东亚亚热带的类群具有较高的净多样化速率, 但淫羊藿属特定的距长对其物种的净多样化速率没有影响; 果实类型、假种皮和气生茎的有无对类群间净多样化速率无影响。本研究表明功能性状的演化和中新世以来东亚地区季风气候的加强导致了鬼臼亚科内部多样性的分布不均。
陈馥艳, 游旨价, 张秋月, 黄健, 星耀武 (2023) 小檗科鬼臼亚科多样性格局的演化历史和成因. 生物多样性, 31, 23100. DOI: 10.17520/biods.2023100.
Fuyan Chen, Chih-Chieh Yu, Qiuyue Zhang, Jian Huang, Yaowu Xing (2023) The diversification history of Podophylloideae (Berberidaceae) and its underlying drivers. Biodiversity Science, 31, 23100. DOI: 10.17520/biods.2023100.
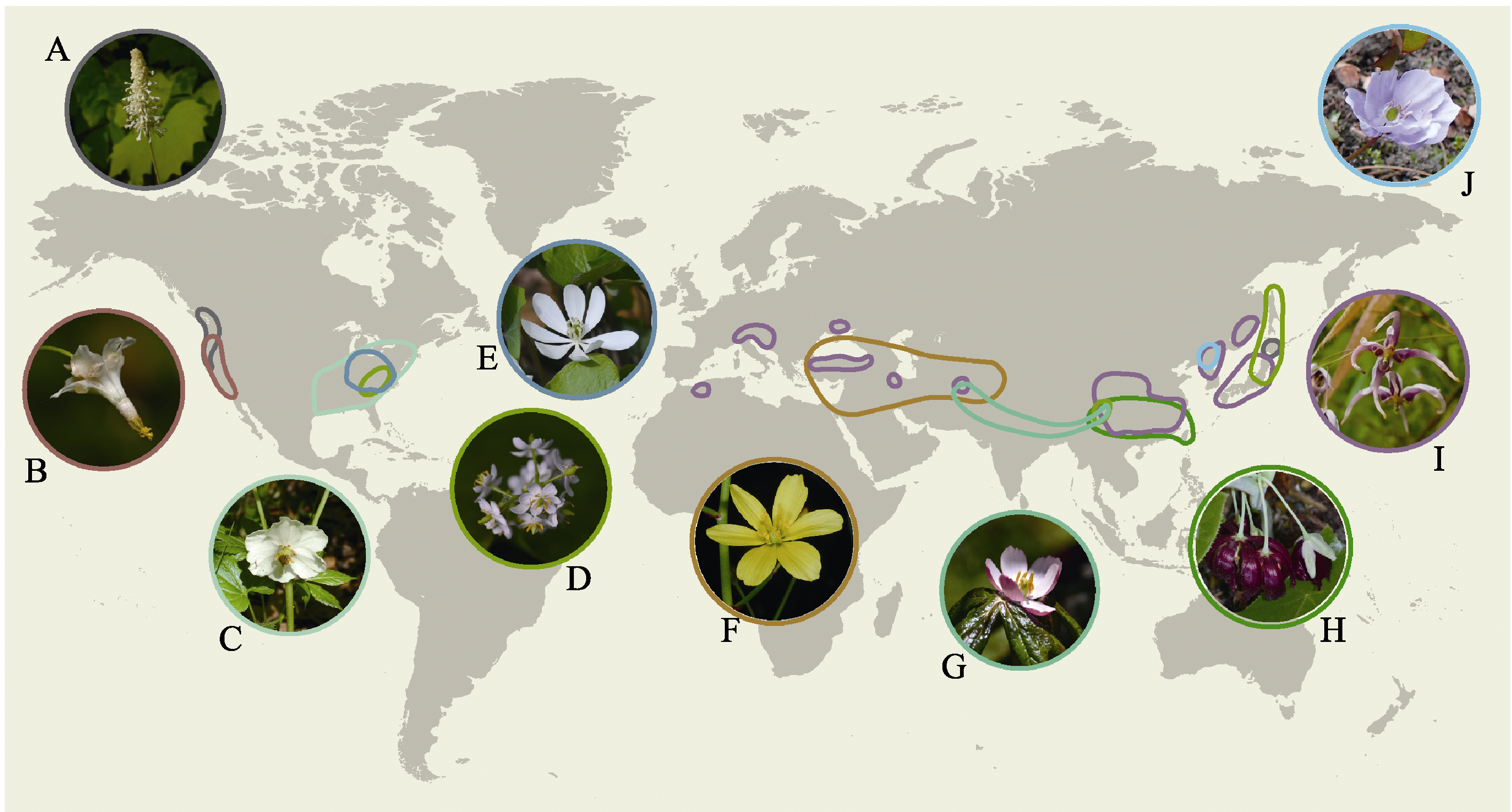
图1 鬼臼亚科不同属的分布及花的形态多样性。(A)裸花草属, Walter Siegmund摄, CC BY-SA 3.0; (B)折瓣花属, Walter Siegmund摄, CC BY-SA 3.0; (C)北美桃儿七属, Wasrts摄, CC BY-SA 4.0; (D)山荷叶属, 游旨价摄; (E)二叶鲜黄连属, Krzysztof Ziarnek摄, CC BY-SA 4.0; (F)山槐叶属, Ori Fragman-Sapir摄, CC BY-SA 3.0; (G)桃儿七属, 游旨价摄; (H)鬼臼属, Cathy DeWitt摄, CC BY 4.0; (I)淫羊藿属, 游旨价摄; (J)鲜黄连属, Krzysztof Ziarnek摄, CC BY-SA 4.0。地图来自 https://vemaps.com。
Fig. 1 Distribution and flower morphological diversity of each genus in Podophylloideae. (A) Achlys, photo by Walter Siegmund/CC BY-SA 3.0; (B) Vancouveria, photo by Walter Siegmund/CC BY-SA 3.0; (C) Podophyllum, photo by Wasrts/CC BY-SA 4.0; (D) Diphylleia, photo by Chih-Chieh Yu; (E) Jeffersonia, photo by Krzysztof Ziarnek /CC BY-SA 4.0; (F) Bongardia, photo by Ori Fragman-Sapir/CC BY-SA 3.0; (G) Sinopodophyllum, photo by Chih-Chieh Yu; (H) Dysosma, photo by Cathy DeWitt/CC BY 4.0; (I)Epimedium, photo by Chih-Chieh Yu; (J) Plagiorhegma, photo by Krzysztof Ziarnek /CC BY-SA 4.0. The map is taken from https://vemaps.com.
| 物种 Species | 采集地点 Sampling site | 序列号 Accession no. |
|---|---|---|
| 天全淫羊藿 E. flavum | 中国四川省天全县二郎山 Erlang Mountain, Tianquan County, Sichuan, China (29.87° N, 102.31° E) | OQ674756 |
| 短茎淫羊藿 E. brachyrrhizum | 中国贵州省岑巩县地郎 Dilang, Cengong County, Guizhou, China (27.32° N, 108.61° E) | OQ674753 |
| 黔岭淫羊藿 E. leptorrhizum | 中国重庆市黔江区仰头山森林公园 Yangtoushan Forest Park, Qianjiang District, Chongqing, China (29.56° N, 108.79° E) | OQ674758 |
| 紫距淫羊藿 E. epsteinii | 中国湖南省桑植县八大公山自然保护区 Badagong Mountain National Nature Reserve, Sangzhi County, Hunan, China (29.78° N, 110.09° E) | OQ674754 |
| 箭叶淫羊藿 E. sagittatum | 中国重庆市忠县刺竹沟 Cizhugou, Zhong County, Chongqing, China (30.10° N, 108.03° E) | OQ674759 |
| E. pubigerum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674751 |
| E. perralderianum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674757 |
| E. alpinum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674755 |
| E. diphyllum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674750 |
| E. grandiflorum var. thunbergianum | 日本岩手县八幡平市 Hachimantai, Iwate Prefecture, Japan (39.95° N, 140.88° E) | OQ674752 |
表1 本研究新采集的淫羊藿属信息及上传至GenBank的序列号
Table 1 Sampling information of newly sequenced Epimedium in this study
| 物种 Species | 采集地点 Sampling site | 序列号 Accession no. |
|---|---|---|
| 天全淫羊藿 E. flavum | 中国四川省天全县二郎山 Erlang Mountain, Tianquan County, Sichuan, China (29.87° N, 102.31° E) | OQ674756 |
| 短茎淫羊藿 E. brachyrrhizum | 中国贵州省岑巩县地郎 Dilang, Cengong County, Guizhou, China (27.32° N, 108.61° E) | OQ674753 |
| 黔岭淫羊藿 E. leptorrhizum | 中国重庆市黔江区仰头山森林公园 Yangtoushan Forest Park, Qianjiang District, Chongqing, China (29.56° N, 108.79° E) | OQ674758 |
| 紫距淫羊藿 E. epsteinii | 中国湖南省桑植县八大公山自然保护区 Badagong Mountain National Nature Reserve, Sangzhi County, Hunan, China (29.78° N, 110.09° E) | OQ674754 |
| 箭叶淫羊藿 E. sagittatum | 中国重庆市忠县刺竹沟 Cizhugou, Zhong County, Chongqing, China (30.10° N, 108.03° E) | OQ674759 |
| E. pubigerum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674751 |
| E. perralderianum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674757 |
| E. alpinum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674755 |
| E. diphyllum | 英国邱园(栽培) Kew Gardens, the United Kingdom (cultivated) | OQ674750 |
| E. grandiflorum var. thunbergianum | 日本岩手县八幡平市 Hachimantai, Iwate Prefecture, Japan (39.95° N, 140.88° E) | OQ674752 |
| 模型 Model | 性状状态 Trait state | 成种速率 Speciation rate (λ) | 灭绝速率 Extinction rate (μ) | 转化率 Turnover rate (q) |
|---|---|---|---|---|
| CID | 0, 1 | λ0 = λ1 | μ0 = μ1 | q01 ≠ q10 |
| BiSSE | 0, 1 | λ0 ≠ λ1 | μ0 ≠ μ1 | q01 ≠ q10 |
| CID-2 | 0A, 1A, 0B, 1B | λ0A = λ1A, λ0B = λ1B | μ0A = μ1A, μ0B = μ1B | q0A→1A = q1A→0A, q0B→1B = q1B→0B |
| CID-4 | 0A, 1A, 0B, 1B, 0C, 1C, 0D, 1D | λ0A = λ1A, λ0B = λ1B, λ0C = λ1C, λ0D = λ1D | μ0A = μ1A, μ0B = μ1B, μ0C = μ1C, μ0D = μ1D | q0A→1A = q1A→0A, q0B→1B = q1B→0B, q0C→1C = q1C→0C, q0D→1D = q1D→0D |
| HiSSE FULL | 0A, 1A, 0B, 1B, 0C, 1C, 0D, 1D | λ0A ≠ λ1A, λ0B ≠ λ1B, λ0C ≠ λ1C, λ0D ≠ λ1D | μ0A ≠ μ1A, μ0B ≠ μ1B, μ0C ≠ μ1C, μ0D ≠ μ1D | q0A→1A ≠ q1A→0A, q0B→1B ≠ q1B→0B, q0C→1C ≠ q1C→0C, q0D→1D ≠ q1D→0D |
表2 隐藏状态物种形成和灭绝分析使用的模型
Table 2 Selected models in Hidden State Speciation and Extinction analysis
| 模型 Model | 性状状态 Trait state | 成种速率 Speciation rate (λ) | 灭绝速率 Extinction rate (μ) | 转化率 Turnover rate (q) |
|---|---|---|---|---|
| CID | 0, 1 | λ0 = λ1 | μ0 = μ1 | q01 ≠ q10 |
| BiSSE | 0, 1 | λ0 ≠ λ1 | μ0 ≠ μ1 | q01 ≠ q10 |
| CID-2 | 0A, 1A, 0B, 1B | λ0A = λ1A, λ0B = λ1B | μ0A = μ1A, μ0B = μ1B | q0A→1A = q1A→0A, q0B→1B = q1B→0B |
| CID-4 | 0A, 1A, 0B, 1B, 0C, 1C, 0D, 1D | λ0A = λ1A, λ0B = λ1B, λ0C = λ1C, λ0D = λ1D | μ0A = μ1A, μ0B = μ1B, μ0C = μ1C, μ0D = μ1D | q0A→1A = q1A→0A, q0B→1B = q1B→0B, q0C→1C = q1C→0C, q0D→1D = q1D→0D |
| HiSSE FULL | 0A, 1A, 0B, 1B, 0C, 1C, 0D, 1D | λ0A ≠ λ1A, λ0B ≠ λ1B, λ0C ≠ λ1C, λ0D ≠ λ1D | μ0A ≠ μ1A, μ0B ≠ μ1B, μ0C ≠ μ1C, μ0D ≠ μ1D | q0A→1A ≠ q1A→0A, q0B→1B ≠ q1B→0B, q0C→1C ≠ q1C→0C, q0D→1D ≠ q1D→0D |
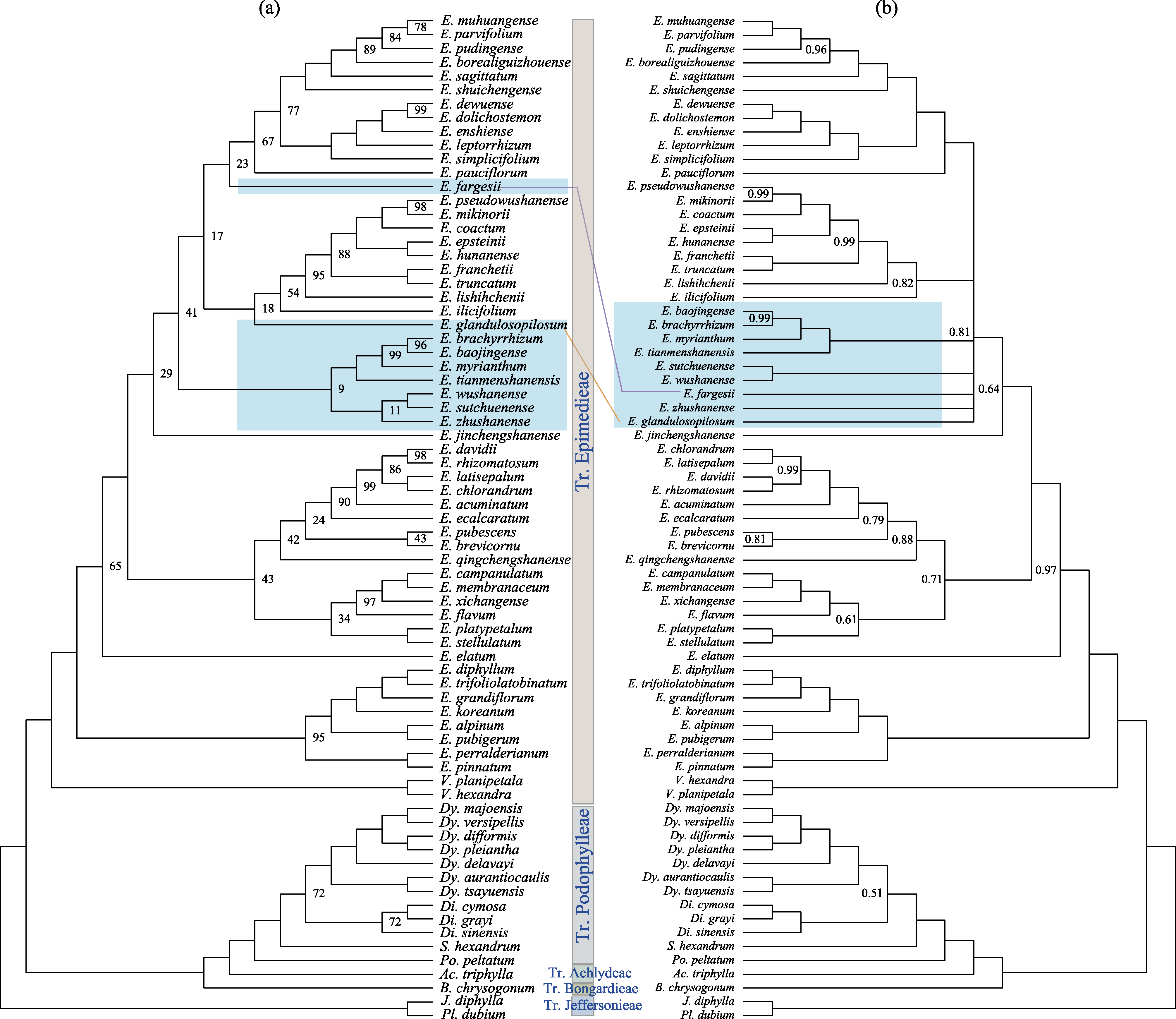
图2 鬼臼亚科系统关系(中文名称见附录1)。(a)和(b)分别为基于最大似然法和贝叶斯方法推断的拓扑结构, 节点处标出了小于100的自展值和小于1.00的后验概率。
Fig. 2 Phylogeny of Podophylloideae (See Chinese name in Appendix 1). (a) and (b) are topology of Maximum likelihood (ML) and Bayesian inference (BI). Bootstrap (BS) value from ML less than 100 and posterior probability (PP) from BI less than 1.00 are labeled on the nodes.
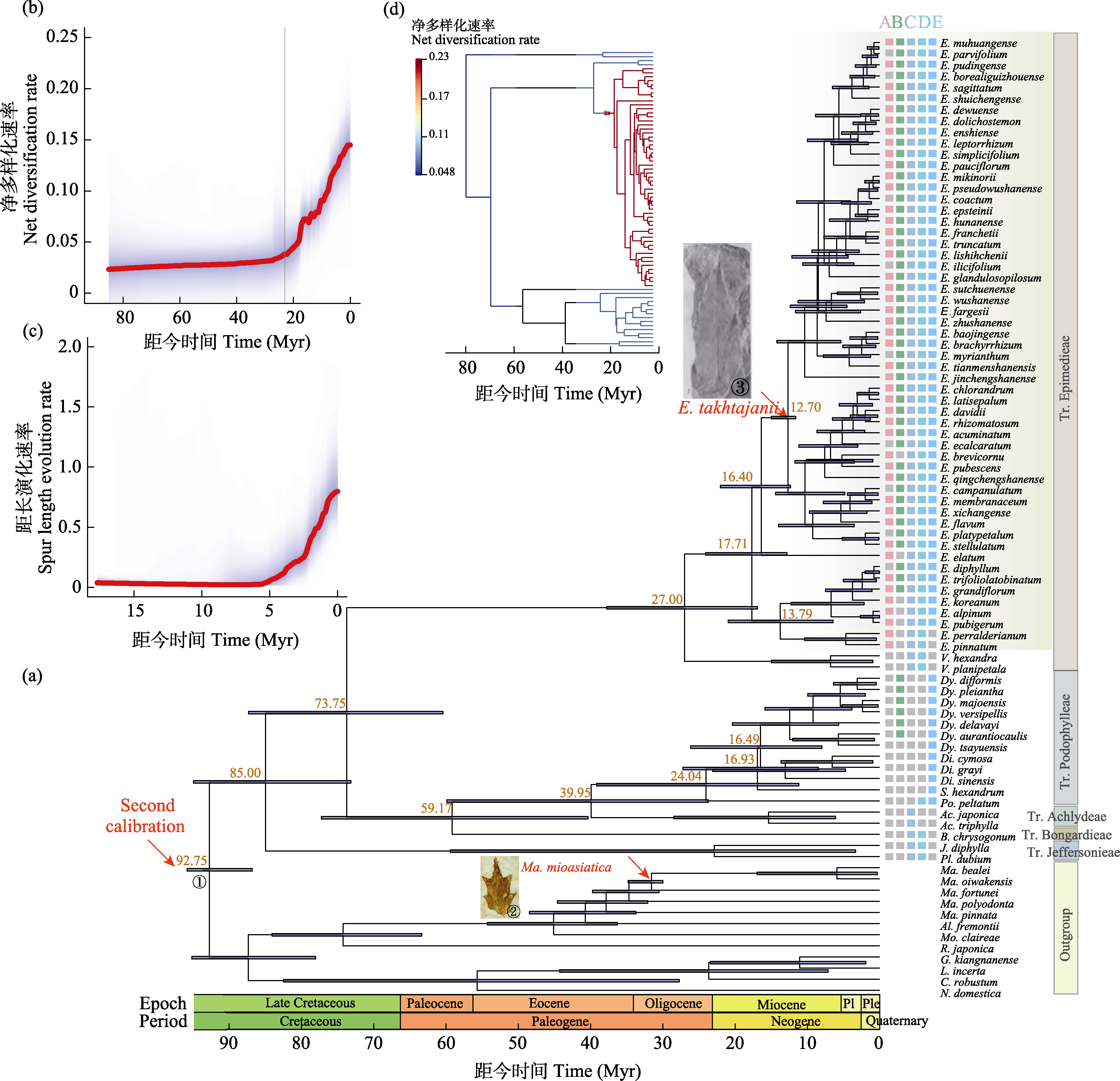
图3 鬼臼亚科的多样化历史。(a)鬼臼亚科分化时间树(节点处蓝色条带代表95%置信区间内的年龄范围), 以及用于HiSSE分析的4个形态性状和1个生境信息分布。其中, A表示有无蜜距(粉色为是, 灰色为否), B表示是否分布于东亚亚热带(绿色为是, 灰色为否), C表示果实类型(灰蓝色为干果, 灰色为浆果), D表示是否有假种皮(浅蓝色为是, 灰色为否), E表示是否有气生茎(深蓝色为是, 灰色为否)。(b)基于BAMM分析的鬼臼亚科的净多样化速率。(c)基于BAMM分析的淫羊藿属距长演化速率。(d)净多样化速率在系统树的分布。中文名称见附录1。
Fig. 3 Diversification history of Podophylloideae. (a) Dated phylogeny of Podophylloideae (horizontal bars at nodes indicate 95% credible intervals of the divergence time estimate), and 4 morphological characters and 1 habitat information used in HiSSE analysis. The colored squares labeled A?E corresponding to each species represents with or without spur (pink or grey), in or out of subtropical East Asia (green or grey), dry or fleshy fruit (bice or grey), with or without aril (light blue or grey), with or without aerial stem (dark blue or grey). (b) Diversification rate of Podophylloideae estimated by BAMM. (c) Rate of spur length evolution in Epimedium estimated by BAMM. (d) Diversification rate distribution on phylogenetic tree. See Chinese name in Appendix 1.
| 性状 Trait | 模型 Model | AIC | AICc | ΔAICc |
|---|---|---|---|---|
| 有距 vs. 无距 With vs. without spur | CID | 532.30 | 532.88 | 25.81 |
| BiSSE | 505.82 | 507.07 | ||
| CID-2 | 522.09 | 533.02 | 25.95 | |
| CID-4 | 510.59 | 513.40 | 6.33 | |
| FULL | 512.36 | 513.24 | 6.17 | |
| 东亚亚热带 vs. 非东亚亚热带 In vs. out of subtropical East Asia | CID | 531.12 | 531.70 | 27.54 |
| BiSSE | 502.91 | 504.16 | ||
| CID-2 | 512.13 | 513.01 | 8.85 | |
| CID-4 | 511.13 | 513.94 | 9.78 | |
| FULL | 521.24 | 532.16 | 28.00 | |
| 果实类型 Fruit type | CID | 495.95 | 496.53 | 20.94 |
| BiSSE | 494.72 | 495.97 | 20.38 | |
| CID-2 | 474.71 | 475.59 | ||
| CID-4 | 490.38 | 493.20 | 17.60 | |
| FULL | 502.27 | 513.20 | 37.61 | |
| 有假种皮 vs. 无假种皮 With vs. without aril | CID | 498.36 | 498.94 | 17.81 |
| BiSSE | 496.64 | 497.89 | 16.76 | |
| CID-2 | 480.25 | 481.13 | ||
| CID-4 | 508.39 | 511.20 | 30.07 | |
| FULL | 511.63 | 522.55 | 41.42 | |
| 有气生茎 vs. 无气生茎 With vs. without aerial stem | CID | 509.46 | 510.04 | 21.00 |
| BiSSE | 487.79 | 489.04 | ||
| CID-2 | 490.63 | 491.51 | 2.47 | |
| CID-4 | 491.49 | 494.31 | 5.47 | |
| FULL | 506.46 | 517.39 | 28.35 |
表3 HiSSE分析模型检测和各性状状态速率结果
Table 3 Model test and rates inferred from HiSSE analysis
| 性状 Trait | 模型 Model | AIC | AICc | ΔAICc |
|---|---|---|---|---|
| 有距 vs. 无距 With vs. without spur | CID | 532.30 | 532.88 | 25.81 |
| BiSSE | 505.82 | 507.07 | ||
| CID-2 | 522.09 | 533.02 | 25.95 | |
| CID-4 | 510.59 | 513.40 | 6.33 | |
| FULL | 512.36 | 513.24 | 6.17 | |
| 东亚亚热带 vs. 非东亚亚热带 In vs. out of subtropical East Asia | CID | 531.12 | 531.70 | 27.54 |
| BiSSE | 502.91 | 504.16 | ||
| CID-2 | 512.13 | 513.01 | 8.85 | |
| CID-4 | 511.13 | 513.94 | 9.78 | |
| FULL | 521.24 | 532.16 | 28.00 | |
| 果实类型 Fruit type | CID | 495.95 | 496.53 | 20.94 |
| BiSSE | 494.72 | 495.97 | 20.38 | |
| CID-2 | 474.71 | 475.59 | ||
| CID-4 | 490.38 | 493.20 | 17.60 | |
| FULL | 502.27 | 513.20 | 37.61 | |
| 有假种皮 vs. 无假种皮 With vs. without aril | CID | 498.36 | 498.94 | 17.81 |
| BiSSE | 496.64 | 497.89 | 16.76 | |
| CID-2 | 480.25 | 481.13 | ||
| CID-4 | 508.39 | 511.20 | 30.07 | |
| FULL | 511.63 | 522.55 | 41.42 | |
| 有气生茎 vs. 无气生茎 With vs. without aerial stem | CID | 509.46 | 510.04 | 21.00 |
| BiSSE | 487.79 | 489.04 | ||
| CID-2 | 490.63 | 491.51 | 2.47 | |
| CID-4 | 491.49 | 494.31 | 5.47 | |
| FULL | 506.46 | 517.39 | 28.35 |
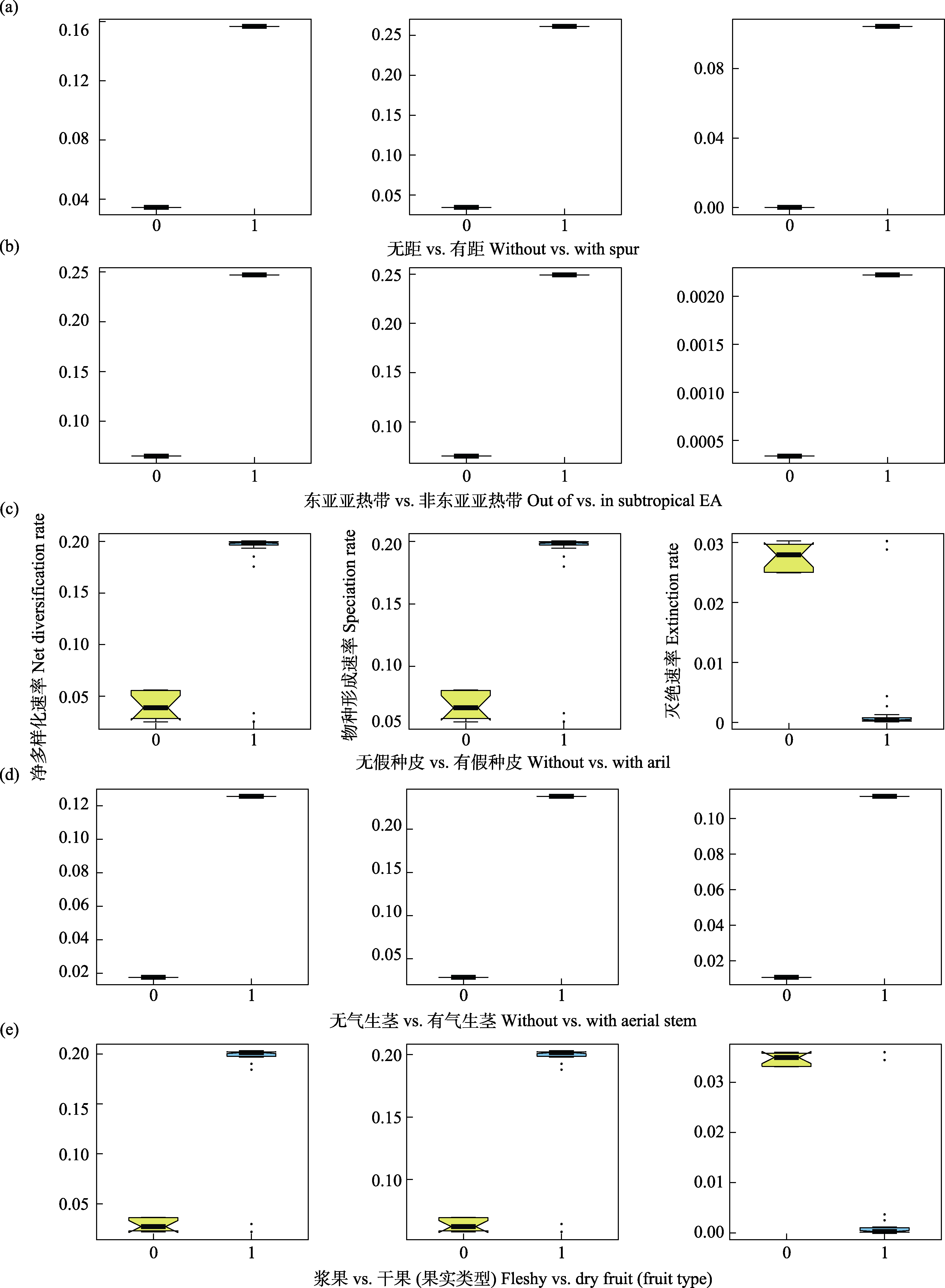
图4 基于二态性状进行多样化速率推断的HiSSE分析结果。(a)?(e)依次为对于是否有距, 是否分布在东亚亚热带, 是否有假种皮, 是否有气生茎和果实类型等性状状态下, HiSSE所推断最优模型下的分化、成种和灭绝速率。
Fig. 4 Result of Hidden State Speciation and Extinction analysis. (a)?(e) illustrate net diversification rate, speciation rate and extinction rate under the best models of the following 5 binary-state traits: the existence of spur, subtropical East Asian distribution, the existence of aril, the existence of aerial stem and fruit type.
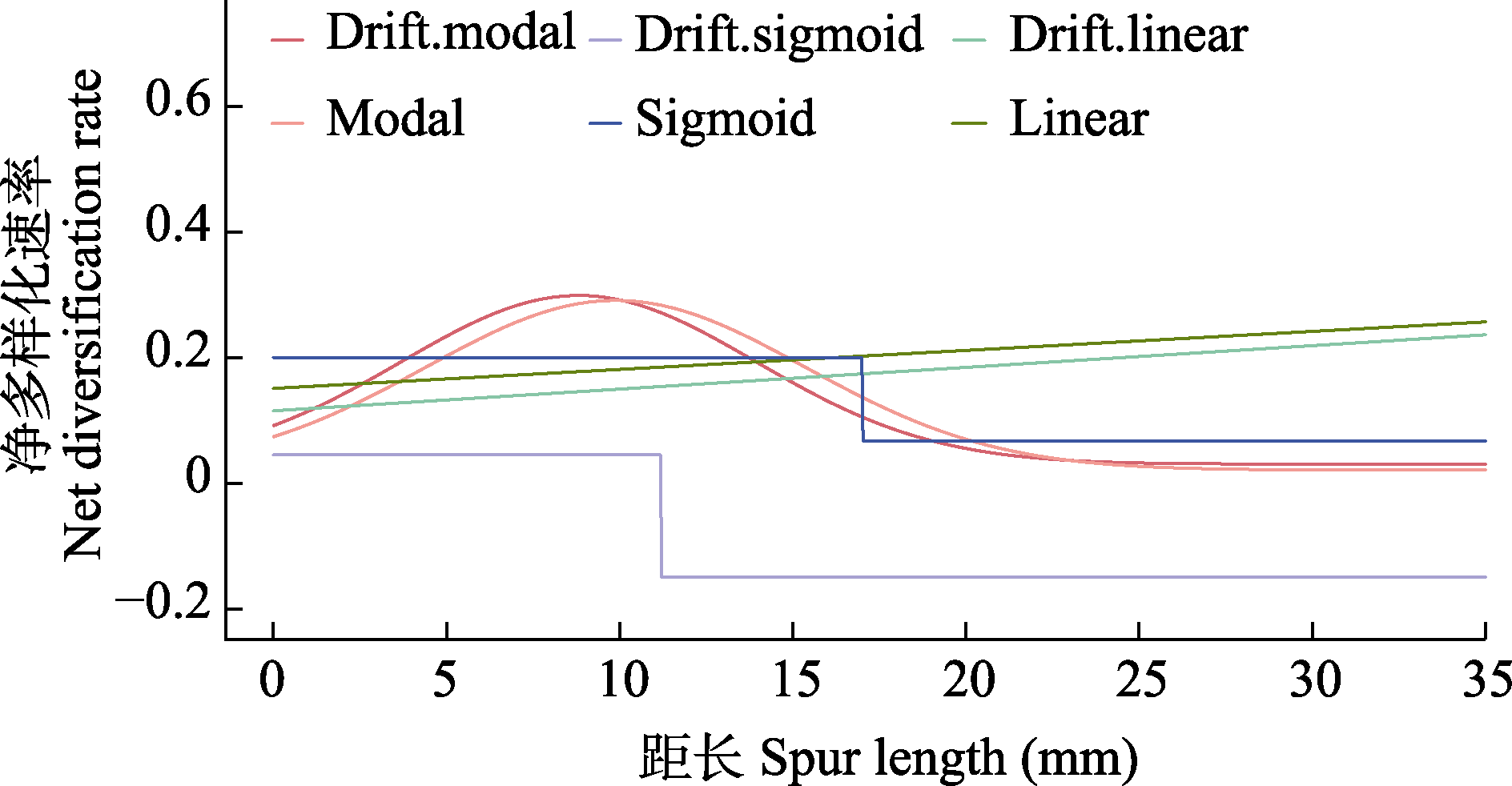
图5 基于定量性状进行多样化速率推断的QuaSSE分析结果: 距长-净多样化速率的分布, 最优模型为modal。图例从左到右分别显示单峰模型、逻辑斯蒂模型和线性模型, 上边一行有漂移率, 下边一行没有漂移率。
Fig. 5 Quantitative state-dependent diversification rate estimation based on QuaSSE analysis. Pattern of spur length- diversification rate, Modal is the best model. The legends showed, from left to right, the Modal, Sigmoid, and Linear models, with drift rates in the top row and no drift rates in the bottom row.
| [1] |
An ZS, Kutzbach JE, Prell WL, Porter SC (2001) Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since Late Miocene times. Nature, 411, 62-66.
DOI |
| [2] |
Armbruster WS, Muchhala N (2009) Associations between floral specialization and species diversity: Cause, effect, or correlation? Evolutionary Ecology, 23, 159-179.
DOI URL |
| [3] |
Beaulieu JM, Donoghue MJ (2013) Fruit evolution and diversification in campanulid angiosperms. Evolution, 67, 3132-3144.
DOI PMID |
| [4] |
Beaulieu JM, O'Meara BC (2016) Detecting hidden diversification shifts in models of trait-dependent speciation and extinction. Systematic Biology, 65, 583-601.
DOI PMID |
| [5] | Bouckaert R, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A, Heled J, Jones G, Kühnert D, Myrio ND, Matschiner M, Mendes FK, Müller NF, Ogilvie HA, Plessis LD, Popinga A, Rambaut A, Rasmussen D, Siveroni I, Suchard MA, Wu CH, Xie D, Zhang C, Stadler T, Drummond AJ (2019) BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Computational Biology, 15, e1006650. |
| [6] |
Byng J, Chase M, Christenhusz M, Fay M, Judd W, Mabberley D, Sennikov A, Soltis D, Soltis P, Stevens P (2016) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, 1-20.
DOI URL |
| [7] | Cho Y, Mower JP, Qiu YL, Palmer JD (2004) Mitochondrial substitution rates are extraordinarily elevated and variable in a genus of flowering plants. Proceedings of the National Academy of Sciences, USA, 101, 17741-17746. |
| [8] |
Christenhusz MJM, Byng JW (2016) The number of known plants species in the world and its annual increase. Phytotaxa, 261, 201-217.
DOI URL |
| [9] |
Coiro M, Doyle JA, Hilton J (2019) How deep is the conflict between molecular and fossil evidence on the age of angiosperms? New Phytologist, 223, 83-99.
DOI PMID |
| [10] |
Condamine FL, Rolland J, Höhna S, Sperling FAH, Sanmartín I (2018) Testing the role of the Red Queen and Court Jester as drivers of the macroevolution of Apollo butterflies. Systematic Biology, 67, 940-964.
DOI PMID |
| [11] |
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: More models, new heuristics and parallel computing. Nature Methods, 9, 772-772.
DOI PMID |
| [12] |
Ding WN, Ree RH, Spicer RA, Xing YW (2020) Ancient orogenic and monsoon-driven assembly of the world's richest temperate alpine flora. Science, 369, 578-581.
DOI URL |
| [13] |
Donoghue MJ (2005) Key innovations, convergence, and success: Macroevolutionary lessons from plant phylogeny. Paleobiology, 31, 77-93.
DOI URL |
| [14] | Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure from small quantity of fresh leaf material. Phytochemical Bulletin, 119, 11-15. |
| [15] |
Drouin G, Daoud H, Xia J (2008) Relative rates of synonymous substitutions in the mitochondrial, chloroplast and nuclear genomes of seed plants. Molecular Phylogenetics and Evolution, 49, 827-831.
DOI PMID |
| [16] | Ehlers J, Gibbard PL (2007) The extent and chronology of Cenozoic Global Glaciation. Quaternary International, 164/165, 6-20. |
| [17] | Enquist BJ, Feng X, Boyle B, Maitner B, Newman EA, Jørgensen PM, Roehrdanz PR, Thiers BM, Burger JR, Corlett RT, Couvreur TLP, Dauby G, Donoghue JC, Foden W, Lovett JC, Marquet PA, Merow C, Midgley G, Morueta-Holme N, Neves DM, Oliveira-Filho AT, Kraft NJB, Park DS, Peet RK, Pillet M, Serra-Diaz JM, Sandel B, Schildhauer M, Šímová I, Violle C, Wieringa JJ, Wiser SK, Hannah L, Svenning JC, McGill BJ (2019) The commonness of rarity: Global and future distribution of rarity across land plants. Science Advances, 5, eaaz0414. |
| [18] |
Fernández-Mazuecos M, Blanco-Pastor JL, Juan AN, Carnicero P, Forrest A, Alarcón M, Vargas P, Glover BJ (2019) Macroevolutionary dynamics of nectar spurs, a key evolutionary innovation. New Phytologist, 222, 1123-1138.
DOI PMID |
| [19] |
Filartiga AL, Klimeš A, Altman J, Nobis MP, Crivellaro A, Schweingruber F, Doležal J (2022) Comparative anatomy of leaf petioles in temperate trees and shrubs: The role of plant size, environment and phylogeny. Annals of Botany, 129, 567-582.
DOI PMID |
| [20] |
FitzJohn RG (2012) Diversitree: Comparative phylogenetic analyses of diversification in R. Methods in Ecology and Evolution, 3, 1084-1092.
DOI URL |
| [21] |
Fiz-Palacios O, Schneider H, Heinrichs J, Savolainen V (2011) Diversification of land plants: Insights from a family-level phylogenetic analysis. BMC Evolutionary Biology, 11, 341.
DOI PMID |
| [22] |
Gorb SN, Gorb EV (1995) Removal rates of seeds of five mamecochorous plants by the ant Formica polyctena (Hymenoptera: Formicidae). Oikos, 73, 367.
DOI URL |
| [23] |
Guo BL (2007) Two new species of Epimedium (Berberidaceae) from China. Acta Phytotaxonomica Sinica, 45, 813.
DOI URL |
| [24] |
Guo MY, Pang XH, Xu YQ, Jiang WJ, Liao BS, Yu JS, Xu J, Song JY, Chen SL (2022) Plastid genome data provide new insights into the phylogeny and evolution of the genus Epimedium. Journal of Advanced Research, 36, 175-185.
DOI PMID |
| [25] |
Gómez C, Espadaler X (2013) An update of the world survey of mamecochorous dispersal distances. Ecography, 36, 1193-1201.
DOI URL |
| [26] | He SZ, Wang YY, Xu WF, Guo BL, Yang XB, Sun QW (2014) The Genus Epimedium of China in Colour. Guizhou Publish Group, Guiyang. (in Chinese) |
| [何顺志, 王悦云, 徐文芬, 郭宝林, 杨相波, 孙庆文 (2014) 中国淫羊藿属植物彩色图鉴. 贵州科技出版社, 贵阳.] | |
| [27] | Hodges SA (1997) Floral nectar spurs and diversification. International Journal of Plant Sciences, 158, S81-S88. |
| [28] | Hodges SA, Arnold ML (1995) Spurring plant diversification:Are floral nectar spurs a key innovation? Proceedings of the Royal Society of London B: Biological Sciences, 262, 343-348. |
| [29] |
Hsieh CL, Yu CC, Huang YL, Chung KF (2022) Mahonia vs. Berberis unloaded: Generic delimitation and infrafamilial classification of Berberidaceae based on plastid phylogenomics. Frontiers in Plant Science, 12, 720171.
DOI URL |
| [30] |
Huang J, Su T, Jia LB, Spicer T, Zhou ZK (2018) A fossil fig from the Miocene of southwestern China: Indication of persistent deep time karst vegetation. Review of Palaeobotany and Palynology, 258, 133-145.
DOI URL |
| [31] |
Huang J, Su T, Lebereton-Anberrée J, Zhang ST, Zhou ZK (2016) The oldest Mahonia (Berberidaceae) fossil from East Asia and its biogeographic implications. Journal of Plant Research, 129, 209-223.
DOI PMID |
| [32] | Huang M, Wang D (2015) The role of elaiosome in seed dispersal of five ant-dispersed herbaceous plants. Acta Ecologica Sinica, 35, 5721-5727. (in Chinese with English abstract) |
| [黄曼, 王东 (2015) 油质体在5种蚁播植物种子散布中的作用. 生态学报, 35, 5721-5727.] | |
| [33] |
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics, 17, 754-755.
DOI PMID |
| [34] |
Jin JJ, Yu WB, Yang JB, Song Y, DePamphilis CW, Yi TS, Li DZ (2020) GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biology, 21, 241.
DOI |
| [35] | Joppa LN, Roberts DL, Pimm SL (2011) How many species of flowering plants are there? Proceedings of the Royal Society B: Biological Sciences, 278, 554-559. |
| [36] | Kutuzkina EF (1965) Discovery of Epimedium (Berberidaceae) in the Sarmatian sediments of the Northern Caucasus. Botanical Journal, 50, 1113-1115. |
| [37] |
Lagomarsino LP, Condamine FL, Antonelli A, Mulch A, Davis CC (2016) The abiotic and biotic drivers of rapid diversification in Andean bellflowers (Campanulaceae). New Phytologist, 210, 1430-1442.
DOI PMID |
| [38] |
Li Q, Sun H, Boufford DE, Bartholomew B, Fritsch PW, Chen JH, Deng T, Ree RH (2021) Grade of Membership models reveal geographical and environmental correlates of floristic structure in a temperate biodiversity hotspot. New Phytologist, 232, 1424-1435.
DOI PMID |
| [39] | Li XQ, Xiang XG, Jabbour F, Hagen O, Ortiz RD, Soltis PS, Soltis DE, Wang W (2022) Biotic colonization of subtropical East Asian caves through time. Proceedings of the National Academy of Sciences, USA, 119, e2207199119. |
| [40] |
Li YX, Quan QM, Sun GL (2009) Effect of floral morphology on fruit set in Epimedium sagittatum (Berberidaceae). Plant Systematics and Evolution, 279, 51-58.
DOI URL |
| [41] |
Magallon S, Sanderson MJ (2001) Absolute diversification rates in angiosperm clades. Evolution, 55, 1762-1780.
PMID |
| [42] |
Minelli A (2016) Species diversity vs. morphological disparity in the light of evolutionary developmental biology. Annals of Botany, 117, 781-794.
DOI PMID |
| [43] |
Pianka ER (1966) Latitudinal gradients in species diversity: A review of concepts. The American Naturalist, 100, 33-46.
DOI URL |
| [44] | Plummer M, Best N, Cowles K, Vines K (2006) CODA: Convergence diagnosis and output analysis for MCMC. R News, 6, 7-11. |
| [45] |
Qian H, Ricklefs RE (1999) A comparison of the taxonomic richness of vascular plants in China and the United States. The American Naturalist, 154, 160-181.
DOI PMID |
| [46] |
Quan C, Liu ZH, Utescher T, Jin JH, Shu JW, Li YX, Liu YS (2014) Revisiting the Paleogene climate pattern of East Asia: A synthetic review. Earth-Science Reviews, 139, 213-230.
DOI URL |
| [47] | Quan QM (2018) Effect of morphs on reproductive biology of Epimedium pubescens (Berberidaceae): A species endemic to China. International Journal of Agriculture and Biology, 20, 945-950. |
| [48] | Rabosky DL (2014) Automatic detection of key innovations, rate shifts, and diversity-dependence on phylogenetic trees. PLoS ONE, 9, e89543. |
| [49] |
Richardson JE, Pennington RT, Pennington TD, Hollingsworth PM (2001) Rapid diversification of a species-rich genus of neotropical rain forest trees. Science, 293, 2242-2245.
DOI PMID |
| [50] | Sauer JD (1988) Plant Migration:The Dynamics of Geographic Patterning in Seed Plant Species. University of California Press, Berkeley. |
| [51] |
Schluter D, Pennell MW (2017) Speciation gradients and the distribution of biodiversity. Nature, 546, 48-55.
DOI URL |
| [52] | Scholl JP, Wiens JJ (2016) Diversification rates and species richness across the Tree of Life. Proceedings of the Royal Society B: Biological Sciences, 283, 20161334. |
| [53] |
Sletvold N, Ågren J (2011) Nonadditive effects of floral display and spur length on reproductive success in a deceptive orchid. Ecology, 92, 2167-2174.
PMID |
| [54] | Song YC, Da LJ (2016) Evergreen broad-leaved forest of East Asia. In: Vegetation Structure and Function at Multiple Spatial, Temporal and Conceptual Scales (ed. Box EO), pp. 101-128. Springer International Publishing, Cham. |
| [55] |
Spriggs EL, Clement WL, Sweeney PW, Madriñán S, Edwards EJ, Donoghue MJ (2015) Temperate radiations and dying embers of a tropical past: The diversification of Viburnum. New Phytologist, 207, 340-354.
DOI URL |
| [56] |
Stamatakis A (2014) RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics, 30, 1312-1313.
DOI PMID |
| [57] | Stearn WT (2002) The Genus Epimedium and other Herbaceous Berberidaceae Including the Genus Podophyllum. Timber Press, Oregon. |
| [58] |
Sun YX, Moore MJ, Yue LL, Feng T, Chu HJ, Chen ST, Ji YH, Wang HC, Li JQ (2014) Chloroplast phylogeography of the East Asian Arcto-Tertiary relict—Tetracentron sinense (Trochodendraceae). Journal of Biogeography, 41, 1721-1732.
DOI URL |
| [59] |
Suzuki K (1984) Pollination system and its significance on isolation and hybridization in Japanese Epimedium (Berberidaceae). The Botanical Magazine, 97, 381-396.
DOI URL |
| [60] | Sweeting MM (1995) Karst in China: Its Geomorphology and Environment. Springer-Verlag, New York. |
| [61] |
Tang YF, Fang Y, Liu CQ, Lu QB, Hu XH (2020) The long spur of Impatiens macrovexilla may reflect adaptation to diurnal hawkmoth pollinators despite diversity of floral visitors. Flora, 266, 151599.
DOI URL |
| [62] |
Tian YM, Spicer RA, Huang J, Zhou ZK, Su T, Widdowson M, Jia LB, Li SH, Wu WJ, Xue L, Luo PH, Zhang ST (2021) New early Oligocene zircon U-Pb dates for the ‘Miocene' Wenshan Basin, Yunnan, China: Biodiversity and paleoenvironment. Earth and Planetary Science Letters, 565, 116929.
DOI URL |
| [63] | Tillich M, Lehwark P, Pellizzer T, Ulbricht-Jones ES, Fischer A, Bock R, Greiner S (2017) GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Research, 45, W6-W11. |
| [64] | Turner GF (2001) The ecology of adaptive radiation. Heredity, 86, 749-750. |
| [65] |
Vlašánková A, Padyšáková E, Bartoš M, Mengual X, Janečková P, Janeček Š (2017) The nectar spur is not only a simple specialization for long-proboscid pollinators. New Phytologist, 215, 1574-1581.
DOI PMID |
| [66] |
Wang W, Chen ZD, Liu Y, Li RQ, Li JH (2007) Phylogenetic and biogeographic diversification of Berberidaceae in the Northern Hemisphere. Systematic Botany, 32, 731-742.
DOI URL |
| [67] |
Whittall JB, Hodges SA (2007) Pollinator shifts drive increasingly long nectar spurs in columbine flowers. Nature, 447, 706-709.
DOI |
| [68] |
Wick RR, Schultz MB, Zobel J, Holt KE (2015) Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics, 31, 3350-3352.
DOI PMID |
| [69] | Xing YW, Ree RH (2017) Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proceedings of the National Academy of Sciences, USA, 114, E3444-E3451. |
| [70] | Ying TS (2001) Flora Reipublicae Popularis Sinicae (Tomus 29): Angiospermae, Dicotyledoneae (Lardizabalaceae,· Berberidaceae). Science Press, Beijing. (in Chinese) |
| [应俊生 (2001) 中国植物志(第二十九卷): 被子植物门, 双子叶植物纲 (木通科, 小檗科). 科学出版社, 北京.] | |
| [71] |
Zhang DG, Deng T, Kim CK, Zhang JW, Nie ZL, Sun H (2015) Epimedium tianmenshanensis (Berberidaceae), a new species from Hunan, China. Phytotaxa, 222, 33-43.
DOI URL |
| [72] |
Zhang SH, Chen TY, Zeng X, Yu Y, Zhang Y, Xie SP (2018) Plant-insect associations from the upper Miocene of Lincang, Yunnan, China. Review of Palaeobotany and Palynology, 259, 55-62.
DOI URL |
| [73] | Zhang X, Landis JB, Sun YX, Zhang HJ, Lin N, Kuang TH, Huang XH, Deng T, Wang HC, Sun H (2021) Macroevolutionary pattern of Saussurea (Asteraceae) provides insights into the drivers of radiating diversification. Proceedings of the Royal Society B: Biological Sciences, 288, 20211575. |
| [74] |
Zhang YJ, Zhang SJ, Dang HS, Zheng B, Li JQ, Wang Y (2016) Epimedium xichangense (Berberidaceae), a new species from Sichuan, China. Phytotaxa, 263, 286.
DOI URL |
| No related articles found! |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()