
|
|
||
|
巴尔通体在滇西南蝙蝠中高度流行并具有丰富的遗传变异特征
生物多样性
2021, 29 (9):
1245-1255.
DOI: 10.17520/biods.2021028
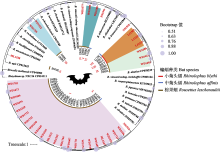
蝙蝠是很多病原微生物的自然宿主, 全球多项研究表明蝙蝠是巴尔通体(Bartonella species)的主要宿主。为了解滇西南地区蝙蝠中巴尔通体的流行特征, 我们于2015-2017年间在云南省4个地区应用网捕法捕获蝙蝠3种305只。经种类鉴定后采集肝脾组织, 提取核酸, 通过TaqMan实时荧光定量PCR方法检测巴尔通体的tmRNA基因ssrA, 并进行测序鉴定和系统发育分析。结果发现172只蝙蝠检出该基因, 总感染率为56.4%; 其中临沧、西双版纳、保山和瑞丽4个采样点的蝙蝠感染率分别为50.0% (22/44)、61.7% (29/47)、62.1% (18/29)和55.7% (103/185)。中菊头蝠(Rhinolophus affinis)、小菊头蝠(R. blythi)和棕果蝠(Rousettus leschenaultii)的感染率分别为50.0% (22/44)、62.1% (18/29)和56.9% (132/232), 差异没有统计学意义(χ2 = 1.135, P = 0.567), 表明巴尔通体在云南当地的蝙蝠种群中高度流行。定量PCR扩增产物2次扩增后测序获得37个巴尔通体ssrA序列, 属于10个系统发育分支, 其中1个为伊丽莎白巴尔通体(B. elizabethae)、特利波契巴尔通体(B. tribocorum)和克拉斯诺夫巴尔通体(B. krasnovii)的近缘种。其余序列与已知巴尔通体距离较远, 与亚洲、欧洲和美洲等其他地域来源于蝙蝠的巴尔通体近缘。遗传多样性分析显示, ssrA基因的核苷酸多样性指数(π)为0.11381 ± 0.00928, 基因型多样性指数(Hd)为0.985 ± 0.010, 形成29个基因型(单倍型), 说明云南蝙蝠巴尔通体具有丰富的遗传多样性。通过对本研究标本与全球相关序列的系统发育网络重建, 分析全球蝙蝠巴尔通体的地理和宿主分布特征, 可以看出巴尔通体与蝙蝠之间存在显著的宿主特异性关联。因此可初步确定蝙蝠-巴尔通体具有协同进化特征, 同时受到地理隔离的影响。 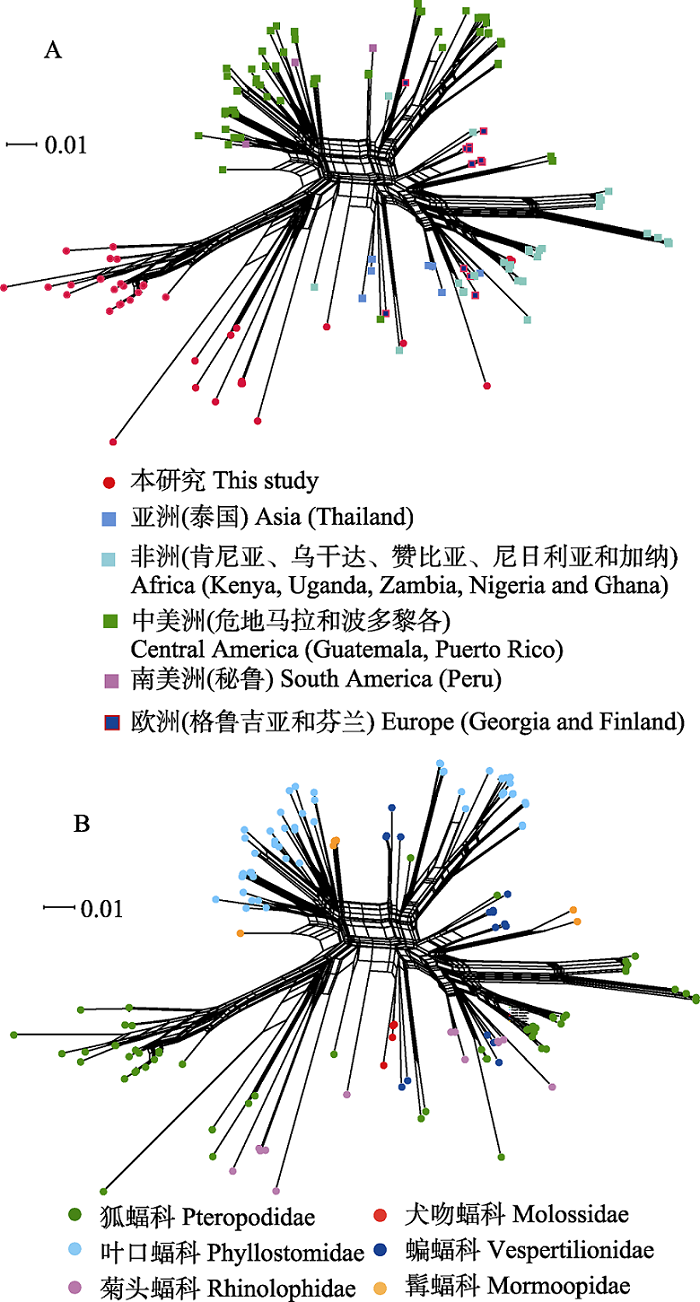 View image in article
图3
基于蝙蝠标本和GenBank中巴尔通体ssrA序列构建的邻接网系统发育网络图。网络图中有152个巴尔通体ssrA序列, 其中本研究序列37个, GenBank数据库下载序列115个。 图A和B分别为依据蝙蝠宿主的地理来源和其分类绘制的网络图。
正文中引用本图/表的段落
在文献和GenBank中搜索到115个来自蝙蝠宿主的巴尔通体ssrA序列, 结合本研究的数据所构建的系统发育网络图基部呈现明显的网状结构, 释放出网状分支间距离矩阵中相互矛盾的信号, 同时也显现出明显的簇状分支, 具有潜在的树结构, 末端分支的自展值均>85%, 多数为100%, 外部节点位置稳定(图3A)。依据蝙蝠的地理来源标识外部节点, 可以看出地理聚集的趋势, 本研究所涉及的中国云南地区序列形成明显的两大类群, 其余个别序列分散到其他地域中, 但没有与泰国序列聚集; 中南美洲形成较为明显的2大类群和一些小分支; 非洲和欧洲的序列也有各自在局部聚集的趋势。总体看, 整个网络图中序列的地理分区明显, 但是有个别序列相互渗透的现象, 比如南美洲秘鲁序列分散在中美洲的2个大分支中, 本研究的数个序列与非洲的聚集在一起, 中美洲也有数个序列靠近非洲分支。
为解析地理分布图中不同地域相互渗透掺杂的原因, 我们按照蝙蝠宿主分类重绘了网络图(图3B), 外部节点以彩色圆点标识, 网络图结构没有变化。从图3B可以看出, 云南菊头蝠、棕果蝠以及泰国犬吻蝠来源的序列处于不同分支, 而云南和泰国菊头蝠来源的序列仍没有聚在一起, 表明同一物种来源的巴尔通体在地理上被分隔开; 中南美洲来自秘鲁与来自危地马拉的蝙蝠科的序列聚在一起, 同样, 来自这2个地区的叶口蝠科的巴尔通体也聚在一起, 另外还有5个危地马拉的髯蝠科的巴尔通体分散形成3个分支, 表明在蝙蝠宿主的科水平上巴尔通体与其有一定的特异性关联, 但并不严格; 非洲及泰国的菊头蝠科来源的巴尔通体聚在一起, 与本研究中的菊头蝠科来源的巴尔通体彼此分开; 非洲狐蝠科来源的序列也形成了数个分支, 呈放射状; 欧洲只有蝙蝠科来源的序列, 形成了数个较为分散的分支, 个别序列与南美洲的同宿主来源的序列形成1个分支。
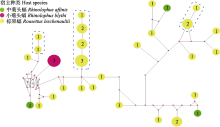
ssrA基因编码转运信使RNA (Transfer- messenger RNA), 是一种存在于细菌中的特殊结构小分子RNA, 长约260?430 bp。tmRNA的5′和3′端序列构成类似tRNA的结构, 中间序列编码肽标签(tag peptide), 兼有tRNA和mRNA的双重功能。tmRNA最主要作用是参与细菌蛋白质翻译过程, 可释放停滞在mRNA上的核糖体, 然后将肽标签加在新生肽链C末端, 使之利于被水解, 这一过程被称为“ribosome rescue” (Janssen & Hayes, 2012)。除了参与到细菌生命循环的质量控制系统中, 在其发育调控、环境应激反应以及致病机制等方面也起到重要作用。该基因按照功能可划分为2个部分: 其一为tRNA类似域, 包括5′-端(~50 nt)和3′-端(~70 nt), 序列相对保守; 另一部分为mRNA类似域, 是中间部分序列包含1个ORF, 序列差异较大。巴尔通体的ssrA基因最早被用于属特异性qPCR检测靶标(Diaz et al, 2012), 是巴尔通体检测较为常用的靶基因之一(Kosoy et al, 2018), 而用此基因序列构建巴尔通体系统发育树的研究相对较少(Sato et al, 2020)。巴尔通体的ssrA基因长约297-358 bp, 在多序列比对时, 我们发现该基因既有保守区也有高变异区, 且变异位点较多, 存在多段高变异和缺失区域, 为确定其是否适用于构建系统发育树, 对序列进行了饱和度检测, 结果为Iss (0.342) < Iss.c (0.780) (P = 0.000, 小于0.05), 核酸序列不趋于饱和, 可以用来建树。从序列相似性和系统发育树可以分辨出10个分支, 均与已知巴尔通体距离较远, 似乎形成了蝙蝠种群特有的巴尔通体, 整体上与猫、犬、灵长类和啮齿类等动物相关的巴尔通体距离较远。这10个分支的蝙蝠巴尔通体可初步判定为一些新种, 详细分类鉴定还需要分离菌株做进一步生化特征、细胞壁脂肪酸构成、多位点序列和基因组特征分析等。37个云南蝙蝠巴尔通体的ssrA基因型多样性丰富(0.985 ± 0.010), 核苷酸变异度较高(0.11381 ± 0.00928), 形成了29个基因型, 表明我国云南地区的蝙蝠巴尔通体遗传变异丰富, 与全球多地的调查结果一致。中性检验Tajima's D为负值, 没有统计检验的支持, 无法说明群体是否在扩张, 标本数量过少、采样不均匀可能是影响统计检验的主要原因。此外, 基因型网络图中, 小菊头蝠的3个序列形成了1个基因型, 而中菊头蝠和棕果蝠巴尔通体没有聚集现象, 不能完全展示出巴尔通体与宿主物种间的关联, 这可能是由于这2种蝙蝠相关巴尔通体遗传多样性丰富、宿主关联性不强, 也可能是由于样本量过少, 无法体现真实情况, 这是本研究中存在的缺陷。
在本研究中, 异地同种蝙蝠来源的巴尔通体呈现出聚集现象, 这反映出潜在的巴尔通体?宿主关联特征, 这一现象在我国山东以及全球其他地区的蝙蝠巴尔通体中也有发现(Han et al, 2017)。为进一步观察本研究中我国云南巴尔通体在系统地理发生、宿主关联性特征以及在全球蝙蝠巴尔通体中的系统发生位置, 我们构建了系统发育网络图, 外部节点分别以蝙蝠来源地域(洲水平)和宿主种类(科水平)标识, 用以判断不同来源的序列的分布模式。在地域分布的系统发育网络图中(图3A), 亚洲、美洲和非洲的聚集特征非常明显, 可以看出地域分化趋势, 欧洲序列数据由于较少, 分散在非洲和中美洲分支中, 很难看出趋势。这种系统地理发生分析有可能存在一个混杂因素, 即宿主因素(McKee et al, 2016)。因此, 我们在网络图(图3A)中看到不同地域的巴尔通体有相互渗透的现象。旧大陆和新大陆的蝙蝠物种分布不同, 这种宿主物种差异很可能是形成巴尔通体系统发育分化和网络图拓扑结构的主要原因, 进一步分析宿主类别差异对聚类所起的作用可以帮助我们辨识巴尔通体?蝙蝠的宿主特异性关联。在宿主分布网络图(图3B)中, 同科蝙蝠巴尔通体明显聚集, 可以解释不同地域聚集的原因, 如南美洲秘鲁和中美洲危地马拉的蝙蝠科及叶口蝠科巴尔通体形成的2个分支。在宿主分布网络图中还呈现出蝙蝠科、髯蝠科和菊头蝠科巴尔通体较为分散的样式, 在亚、非、欧、美及中南美洲的分支间相互渗透分布, 表明这些物种的巴尔通体遗传变异相当丰富, 且早已随宿主在全球扩散。总体来看, 巴尔通体与蝙蝠之间存在明显宿主特异性特征, 在更高一级的宿主分类水平, 这一特征就显得更为严格, McKee等(2016)构建的蝙蝠亚目分类水平的巴尔通体二叉树, 非常整齐地将巴尔通体划分为2大进化分支, 即Yinpterochiroptera亚目和Yangochiroptera亚目分支, 从此推断巴尔通体很早就与蝙蝠形成协同进化(coevolution)关系(Morse et al, 2012), 随着蝙蝠物种的分化而共同分化(codivergence), 呈现出适应性辐射(adaptive radiation)的进化模式, 蝙蝠宿主的同域分布与巴尔通体系统发生具有显著的一致性。
本文的其它图/表
|
{kind=link}