



昆虫是草地生态系统中数量和种类最丰富的生物类群之一, 涵盖不同的形态和生活方式, 占据食物网的多个营养级(van Klink et al, 2020)。昆虫不仅在维持草地生态系统多样性和稳定性中发挥重要作用(Belovsky & Slade, 2000), 也可以提供一系列关键的生态服务功能(如授粉、害虫防控、物质分解等) (Losey & Vaughan, 2006; Stevens et al, 2018; David et al, 2019)。然而, 由于气候变化、人类活动以及草地资源的不合理利用, 昆虫的数量和多样性正在急剧下降(van Klink et al, 2020; Wagner, 2020; Wagner et al, 2021), 其多样性的下降速度远超植物与脊椎动物(Thomas et al, 2004)。在温带地区, 近40%的昆虫物种正面临灭绝风险(Basset & Lamarre, 2019)。昆虫种群数量和多样性的衰减会严重威胁当地的生态安全, 因此引起了人们的广泛关注。
工业革命以来, 随着化石燃料的大量燃烧和农业化肥的广泛施用, 越来越多的活性氮被排放到大气中并通过干湿沉降返回地球表面, 从而导致全球范围内氮沉降总量及速率不断增加, 对生物地球化学循环和生态系统稳定性产生了深刻影响(Galloway et al, 1995; Goulding et al, 1998)。目前亚洲已成为氮沉降增速最快的区域之一, 中国氮沉降速率增幅最高可达111.5 kg N·km−2·year−1 (Liu et al, 2013; Ackerman et al, 2019)。草地是全球分布最广、面积最大的陆地生态系统, 覆盖了约24%的地球陆地表面(李建东和方精云, 2017)。氮沉降会显著影响草地生态系统的结构和功能。例如, 氮沉降可以提高环境氮的可利用性, 促进初级生产力(Borer & Stevens, 2022); 同时氮沉降可以提高多年生禾草等植物的优势度, 通过加强植物对光和营养等资源的竞争, 降低植物多样性(Stevens et al, 2004; Silvertown et al, 2006; Simkin et al, 2016)。虽然人们关于氮沉降对草地植物群落的影响已有了深刻理解, 但对于昆虫群落受到的影响及作用机制尚不明确, 缺乏系统性认识。本文结合已有相关研究, 对氮沉降影响草地昆虫群落的作用模式及其调控机制进行梳理总结, 有助于全面理解氮沉降背景下草地昆虫群落的变化规律, 为保护草地生物多样性和维持草地生态系统健康提供理论依据。
1 氮沉降对草地昆虫群落的作用及其影响因素
植物和昆虫作为草地生态系统的重要组成部分, 形成了相互制约与相互依存的协同进化关系(王德利, 2004)。由于氮沉降可以对植物群落产生强烈影响, 因此可以间接地对更高营养级的昆虫群落产生深远影响(Nijssen et al, 2017; 孙玉诚等, 2017)。一些研究表明, 氮沉降可以通过促进初级生产力, 提高植物营养含量, 引起昆虫数量和生物量显著提高(Richardson et al, 2002; de Sassi et al, 2012; de Sassi & Tylianakis, 2012; Grinath, 2021)。但也有研究显示, 氮沉降可以通过降低植物适口性, 或改变植物群落组成和结构, 抑制昆虫种群数量(Chen et al, 2010; Cease et al, 2012; Asmus et al, 2018; Roth et al, 2021)。氮沉降会引起昆虫群落组成发生变化, 对昆虫多样性产生正向、负向或中性的影响。大多数研究显示, 随着环境氮浓度提高, 昆虫物种丰富度呈下降趋势(Haddad et al, 2000; Asmus et al, 2018)。Nessel等(2021)近期的meta分析表明, 氮富集使陆地昆虫多样性降低了33.8%。然而也有研究显示, 氮沉降不会改变昆虫多样性(Cuevas-Reyes et al, 2011)。例如, Meza-Lopez等(2018)研究发现, 氮沉降虽然提高了昆虫群落总多度, 但不会对昆虫物种丰富度产生显著影响。
氮沉降对昆虫群落的作用方向受到不同因素的影响, 包括模拟氮沉降处理水平的差异(氮沉降持续时间和强度)、不同的昆虫类群(摄食类群和功能群)以及研究的草地类型(附录1)。
1.1 氮沉降持续时间和强度
短期氮添加会对昆虫个体性能(如繁殖和生存)产生显著影响, 但对昆虫种群或群落水平的影响较为有限(Williams & Cronin, 2004; Huberty & Denno, 2006)。随着氮添加时间累积, 氮沉降对昆虫群落的影响会逐渐增强。Murphy等(2012)对比了不同氮添加时间对昆虫群落结构的影响, 发现捕食者和植食性昆虫飞虱的数量比例随施氮时间逐年增加, 表明昆虫群落对氮添加的响应可能需要数年时间才能显现。研究表明, 长期氮沉降(> 10年)可以对昆虫群落的组成和结构产生更强烈的影响(Haddad et al, 2000; Asmus et al, 2018)。然而也有研究表明, 氮添加对昆虫的影响不随时间积累持续加强。Meta分析的结果表明, 氮沉降对陆地无脊椎动物数量的负面影响在短期(< 0.5年)内甚至要高于中长期(3-10年)施氮处理(Nessel et al, 2021)。此外, 不同昆虫功能群对施氮时间的响应并不一致。Cuesta等(2008)发现, 植食者的数量在施氮3个月和15个月后均显著增加, 而捕食者数量在施氮3个月后增加, 但在施氮15个月后呈下降趋势。
随着施氮浓度提高, 氮沉降对昆虫数量和多样性的影响会逐渐加强。Haddad等(2000)的研究表明, 随着施氮浓度的增加(5 g N·m-2·year-1增加到30 g N·m-2·year-1), 昆虫群落物种丰富度呈线性下降, 而昆虫数量呈线性上升。然而草地昆虫对氮添加的响应并不全是呈“线性模式”, 有时也呈“单峰模式” (Westoby, 1978; Pöyry et al, 2006)。营养平衡假说(nutrient balance hypothesis)认为, 昆虫对食物营养具有特定的浓度需求, 营养过高或过低都会降低物种适合度(Raubenheimer & Simpson, 1993)。Raubenheimer和Simpson (2004)提出“几何框架”理论(geometric framework), 指出昆虫的营养需求具有种间差异性。例如, 亚洲小车蝗(Oedaleus asiaticus)偏爱碳水化合物含量高(低氮)的食物(Cease et al, 2012), 而棉铃虫(Heliothis zea)选择蛋白质含量高(高氮)的食物(Perner et al, 2005)。即使亲缘关系相近的昆虫也可能具有不同的营养需求(Braswell et al, 2019)。Zhu等(2023a)的研究表明, 不同强度氮添加处理会对两种同属的蝗虫(Euchorthippus cheui和E. unicolor)个体性能和种群数量产生相反的影响。因此, 昆虫的营养需求既取决于自身特定的发育阶段、性别和生理状态, 也是与环境长期协同进化的结果。
1.2 昆虫类群
不同昆虫类群具有不同的生活史, 以及不同的生理和生态学特征, 因此对氮沉降的响应具有显著差异。有研究表明, 氮沉降对半翅目(Gratton & Denno, 2003)及鞘翅目(Altfeld & Stiling, 2009)的物种存在正向影响, 而对鳞翅目(Roth et al, 2021)和直翅目(Cease et al, 2012)的物种存在负向影响。对于访花昆虫, 氮沉降会抑制植物群落中开花的杂类草类群, 减少这些昆虫的食物资源(主要为花粉/花蜜), 进而对其种群数量和丰富度产生负面影响(Potts et al, 2010)。Roth等(2021)通过分析瑞士383个地点的蝴蝶长期监测数据发现, 蝴蝶的数量(尤其是某些濒危物种)与氮沉降呈显著的负相关, 高氮沉降水平会显著减少当地蝴蝶的物种多样性。不同摄食类群的昆虫对氮沉降响应不同, 刺吸式昆虫(诸如叶蝉、飞虱、蝽、蚜虫)相比于咀嚼式昆虫, 对植物汁液的营养含量变化更为敏感, 因此施氮可以对其个体性能和种群数量产生更强的影响(Prestidge, 1982; Huberty & Denno, 2006; Cuesta et al, 2008; Altfeld & Stiling, 2009)。
氮沉降对昆虫的影响还依赖于昆虫的营养级位置。不同营养级昆虫的自身组织元素含量和摄入食物的元素含量具有差异(Sterner & Elser, 2003; González et al, 2018), 具有不同的养分需求, 因此对氮沉降具有不同的响应。低营养级昆虫受到的影响可能比高营养级昆虫更为强烈。例如, Asmus等(2018)发现, 昆虫群落受到氮沉降的影响主要体现在低营养级的植食者和分解者上, 这可能是由于低营养级昆虫对植物营养含量的变化更加敏感。相对于低营养级昆虫, 捕食者和寄生者与植物组织的氮含量差异较大, 因此对维持自身内稳态有更高的生理需求。有研究表明, 氮添加对植食者和分解者的数量和生物量产生积极影响, 但是对寄生者产生负面影响(Haddad et al, 2000)。
1.3 草地类型
氮沉降对昆虫群落的作用也会受到草地类型的影响。不同草地类型具有不同的初级生产力、植物群落组成以及养分循环速率(Bowman et al, 2008; 韩其飞等, 2021), 因此会进一步影响氮沉降对昆虫群落的作用方向和强度。不同草地类型具有不同的植物优势种和功能群比例, 对氮沉降的响应具有差异。例如, 氮添加会显著提高小灌木沼桦(Betula nana)的优势度而降低草本植物(如禾本科、莎草科)的优势度(Asmus et al, 2018)。然而对于昆虫来说, 草本植物比灌木的适口性更强, 也能提供更合适的生境条件, 因此虽然氮沉降造成的灌木入侵导致了系统更高的初级生产力, 但是并没有提高昆虫群落总数量和生物量(Asmus et al, 2018)。而在典型草原和盐沼的氮沉降研究中, 施氮一般会通过提升植物生物量以及植物适口性来增加昆虫的数量或生物量(Haddad et al, 2000; Huberty & Denno, 2006)。具体而言, 氮添加可以使典型草地植物群落组成向适口性更强的C3植物转变(Isbell et al, 2013), 对于单一植物米草属(Spartina)禾草占优势的盐沼系统来说, 氮添加不会改变植物群落组成, 但可以通过提高优势种的氮含量和适口性促进昆虫种群数量(Murphy et al, 2012)。Prather等(2021)在北美典型草原的研究样点发现氮添加会显著改变禾草的叶片氮含量而不会影响杂类草的叶片氮含量, 从而对昆虫群落产生影响。
2 氮沉降对昆虫群落的作用途径
氮沉降可以对昆虫产生直接的化学胁迫作用, 这一过程通常发生在水体或较为湿润的土壤环境中(Camargo & Alonso, 2006)。氮沉降引起的大量无机氮化合物(如NH4+、NO2-、NO3-等)增加可以对昆虫产生直接的毒性作用(Nijssen et al, 2017)。氮沉降也会降低土壤pH值, 引起土壤的酸化效应(Throop & Lerdau, 2004; Camargo & Alonso, 2006; 陈云等, 2021)。酸化效应会使土壤中H+、Al和Mn等毒性元素浓度增加, 活性增强, 从而直接影响昆虫的生理代谢过程(Roem & Berendse, 2000; Horswill et al, 2008; Tian et al, 2020)。例如, 土壤酸化会扰乱昆虫体内的离子调节、呼吸和代谢(Leuven & Oyen, 1987), 降低昆虫的孵化率和蜕皮成功率, 抑制昆虫的采食行为, 最终影响生长速率(Økland, 1992)。
图1
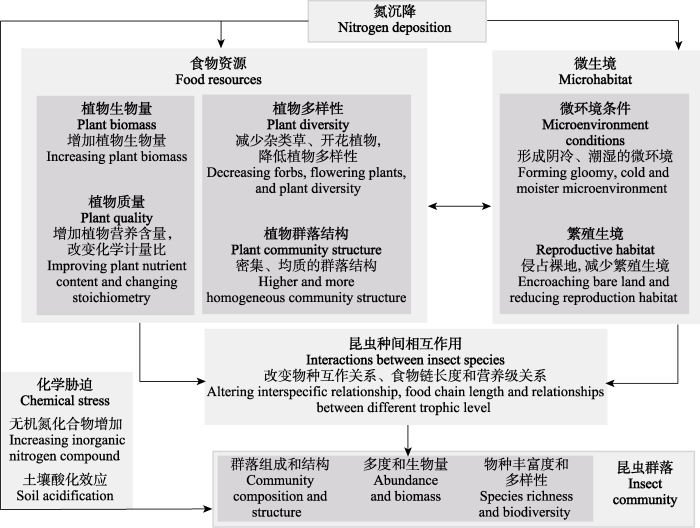
图1
氮沉降对草地昆虫群落作用途径的概念框架
Fig. 1
Conceptual framework for the impact pathways of nitrogen deposition on grassland insect communities
2.1 食物资源
2.1.1 植物多样性
氮沉降造成的植物群落多样性变化在全球不同的草地生态系统中普遍存在(Stevens et al, 2004; Silvertown et al, 2006)。氮沉降可以降低土壤养分资源异质性, 诱发不同植物对光、水分和养分资源竞争, 使得植物间产生不对称的种间竞争: 促进喜氮的高大禾本科或灌木物种快速生长, 抑制矮小的竞争力较弱的禾本科和杂类草物种, 导致植物群落组成发生改变, 引起植物多样性下降(张世虎等, 2022)。植物多样性发生变化会对昆虫群落数量和多样性产生显著影响(Wan et al, 2022)。Haddad等(2009)的研究结果表明, 植食性昆虫和捕食性昆虫物种丰富度与植物物种丰富度呈强烈的正相关关系。资源专食性假说(resource specialization hypothesis)预测, 植食性昆虫多样性随植物多样性的增加而增加, 而捕食性昆虫多样性随植食者多样性的增加而增加(Keddy, 1984)。因此, 氮沉降导致的植物多样性下降会通过营养级联作用对昆虫群落产生强烈影响。
2.1.2 植物生物量
氮沉降导致的植物生物量提高和枯落物积累可以显著提高输入地上食物网以及地下碎屑食物网的资源数量, 进而增加昆虫多度和丰富度。更多个体假说(more individuals hypothesis)认为, 生产力更高的植物群落可以为消费者提供更丰富的食物资源, 进而支持数量更多的消费者, 导致总体物种丰富度增加(Srivastava & Lawton, 1998)。然而许多昆虫物种依赖特定的植物物种或器官(植物鲜嫩组织、花蜜或花粉、种子等)生存, 氮沉降引起的这些特定寄主植物的减少或丧失会对这些昆虫类群产生严重威胁(David et al, 2019)。Öckinger等(2006)发现蝴蝶物种数与氮沉降量存在负相关, 并认为这种关系可能受寄主植物数量的调节。WallisDeVries等(2012)指出, 由于氮沉降导致了植物群落组成变化、 开花植物数量降低或是物候变化, 导致访花昆虫的数量发生大规模的下降。专食性昆虫物种通常比广食性物种更容易受到这些变化的影响, 因为它们的食物来源更加单一(Bernaschini et al, 2020)。虽然氮沉降可以引起食物资源数量发生变化进而影响昆虫多度, 但是验证两者之间因果关系的证据十分缺乏(Nijssen et al, 2017)。
2.1.3 植物质量
氮沉降可以通过两种途径改变昆虫食物资源的质量。一方面氮沉降可以直接影响植株个体的营养、次生代谢物和元素比例, 另一方面可以通过改变植物群落组成比例影响群落水平的植物质量(Nijssen et al, 2017)。氮含量被认为是表征植物质量的最佳指标, 氮沉降可以提高植物组织氮含量, 对昆虫个体性能和种群数量产生积极影响(Throop & Lerdau, 2004)。氮限制假说(nitrogen limitation hypothesis)认为, 植物和昆虫氮含量具有很大差异(植物: 2%-4%; 昆虫: 8%-14%), 因此氮沉降可以打破昆虫的氮限制, 有利于昆虫的生存和繁殖(Mattson, 1980)。但也有研究认为, 昆虫对植物氮含量具有特定的浓度需求, 氮含量不足或过剩都会降低其适合度(Zhu et al, 2019)。例如, 内蒙古草原的亚洲小车蝗(Oedaleus asiaticus)更偏向于选择采食低氮含量的食物, 施氮会抑制其个体性能和种群数量(Cease et al, 2012)。除了提高植物营养含量, 外源氮输入也会增加含氮次生代谢产物(如生物碱、非蛋白氨基酸等)的含量, 对昆虫产生负面影响(Mattson 1980; Li et al, 2016; David et al, 2019)。氮沉降会影响植物和昆虫的化学计量比(张世虎等, 2022)。虽然昆虫较植物具有更高的化学计量内稳态(stoichiometric homeostasis), 但维持内稳态的同时也会对其适合度造成负面影响。目前关于氮沉降对植物和昆虫化学计量关系的研究比较匮乏。有限的研究表明氮沉降可使草地从氮限制转向磷限制, 但是这种转变对昆虫群落的影响还不清楚。从化学计量关系的角度探讨氮沉降对昆虫群落的影响为环境-植物-昆虫互作关系提供了新的视角。综上, 氮沉降通过改变植物质量对昆虫产生的影响既受到植物特性(营养含量、防御水平、元素比例)的影响, 也受到昆虫自身特性的调控(Persson et al, 2010)。
2.1.4 植物群落结构
植物群落结构可表征为植物群落的平均高度或植物高度的异质性(heterogeneity of plant height) (Joern & Laws, 2013)。植物群落结构会影响昆虫的生境条件、食物资源和捕食风险。相较均质化群落, 复杂的植物群落结构可以支持更高的昆虫物种丰富度和数量(Jerrentrup et al, 2014; van Klink et al, 2015)。因为不同高度的植物可以满足不同昆虫类群对食物资源、微生境(繁殖或休眠)和躲避天敌的需求(Lawton, 1983; Treweek et al, 1997)。复杂的植物群落结构也可以缓冲外界环境的急剧变化, 为昆虫在极端气候或严重干扰条件下提供生存庇护(Nijssen et al, 2017)。Aranda和Graciolli (2015)在巴西热带稀树草原的研究表明, 植物群落结构驱动了膜翅目群落的变化。在中国东北松嫩草地的研究表明, 植物高度的异质性是调控昆虫多样性的重要因素(Zhu et al, 2012)。一般而言, 高大且复杂的植物群落结构可以为昆虫提供更多的生态位。因此氮沉降可以通过降低植物高度异质性对昆虫群落产生负面影响。
2.2 微生境
2.2.1 微环境条件
昆虫作为小型变温动物, 需要依赖于外部热量进行体温调节, 因此昆虫对微环境温度、湿度、光照的变化十分敏感(Jerrentrup et al, 2014)。氮沉降导致的微环境条件的变化会直接影响昆虫的发育、繁殖和生存(Nijssen et al, 2017)。氮沉降可以增加植物生物量, 导致地表形成更厚的枯落物层, 进而减弱光照辐射以及地表的空气循环, 形成更为阴冷、潮湿的微环境, 对昆虫产生不利影响(Klop et al, 2015; Nijssen et al, 2017)。在荷兰白垩草地(chalk grasslands)的研究表明, 氮沉降导致的植被快速生长显著降低了微环境温度, 抑制了切胸蚁(Temnothorax albipennis)的繁殖和种群数量(van Noordwijk et al, 2012)。在地表温度较低的条件下, 草地毛虫(Gynaephora alpherakjj)个体性能显著下降(Yang et al, 2017)。微环境的湿度变化会影响昆虫身体组织的水分平衡, 进而促进或抑制昆虫的生长发育速率(常晓娜等, 2008)。例如, 较低的环境相对湿度会减少亮斑扁角水虻(Hermetia illucens)的孵化率和成虫羽化率, 并延长其发育时间(Holmes et al, 2012)。昆虫的觅食行为和繁殖也受到光照条件(光强、光周期、波长)的强烈影响(Shi et al, 2017)。与短光照时长相比, 长光照可以显著提高西花蓟马(Frankliniella occidentalis)的觅食行为和产卵数量(Whittaker & Kirk, 2004)。此外, 氮沉降导致的微环境条件改变不仅会对昆虫生理和性能产生直接影响, 也会通过改变食物或天敌数量对昆虫产生间接作用, 但如何区分微环境变化对昆虫的直接和间接作用尚不清楚。
2.2.2 繁殖生境
2.3 昆虫种间相互作用
昆虫群落存在多种形式的种间相互作用(如竞争、捕食和互惠)。相互作用可以发生在相同营养级的物种间, 也存在于不同营养级的物种间, 如级联效应(cascade effect)。昆虫群落占据多个营养级, 一个营养级的变化会对其他营养级乃至整个食物网产生复杂的影响(Schmitz, 2010; van Klink et al, 2015; 朱玉等, 2017)。氮沉降可以影响同一营养级内的物种互作关系, 但研究案例多来自植物群落。例如内蒙古荒漠草原的研究表明, 氮沉降显著改变植物的竞争关系(Ma et al, 2022), 而关于氮沉降对相同营养级昆虫的种间竞争或互惠关系的研究比较少见。氮沉降对昆虫多营养级的研究十分匮乏(Nijssen et al, 2017)。根据理论假设, 氮沉降可能通过增加资源输入或提高植物营养水平对昆虫不同营养级产生正向效应, 增加食物链长度; 另一方面, 氮沉降也会引起寄主植物数量减少(如某些开花的杂类草), 降低植物多样性和群落结构异质性, 对昆虫不同营养级产生负面影响, 或者通过改变生境和食物资源适宜性抑制天敌数量, 从而缩短食物链长度(Nijssen et al, 2017)。例如, 氮添加通过降低寄主植食性象甲(Mecinus pascuorum)的数量, 显著地抑制了寄生蜂(Mesopolobus incultus)的数量(Hancock et al, 2013)。
氮沉降可以对昆虫食物网产生强烈的级联效应。级联效应可以通过上行作用(bottom-up effect), 即通过植食者数量的变化影响更高营养级的天敌数量, 也可以表现为下行作用(top-down effect), 即较高营养级的天敌数量可以控制植食者的种群数量(天敌假说, Russell, 1989)。一般认为草地食物网中植物驱动的上行作用占优势(Scherber et al, 2010; Wan et al, 2022)。氮沉降可以通过改变植物多样性(资源专食性假说)、植物质量(氮限制假说、营养平衡假说)、植物生物量(更多个体假说)影响昆虫不同营养级的种间相互作用(图2)。例如, Zhu等(2023b)的研究表明, 昆虫不同营养级受不同调控机制影响: 植食者对植物质量变化更为敏感, 地表捕食者对植物多样性变化较为敏感, 冠层捕食者对植物生物量变化较为敏感。而下行作用的研究案例相对非常有限。厘清氮沉降背景下上行作用和下行作用在调控草地昆虫群落及其相对贡献中的作用仍然是一项艰巨的挑战。未来亟需基于食物网的方法(food-web approaches)探究昆虫复杂的种间互作关系。
图2

图2
植物调节不同营养级昆虫间关系的5种主要假说
Fig. 2
Five hypotheses regarding how plants regulate the relationships among insects at different trophic levels
3 展望
综上所述, 当前氮沉降对草地昆虫群落的影响已积累了一定的理论认识和实验证据, 但仍存在诸多不足。第一, 外源氮输入可以同时改变植物群落的多个属性(包括物种丰富度、生产力、群落组成和植物质量), 因此在单一植物群落内很难分离植物群落不同属性对昆虫群落的影响。未来的研究应充分考虑草地植物群落中不同物种或功能群对昆虫群落的影响, 以厘清植物数量、植物质量以及其他植物性状对昆虫群落的独立和联合作用, 有利于梳理氮沉降对昆虫群落的作用机制。第二, 大多数研究关注了氮沉降对低营养级昆虫的影响, 但关于氮沉降对高营养级(如捕食者、寄生者)尤其是昆虫食物网的影响及调控机制还十分薄弱。在自然环境下,昆虫多营养级相互作用尤为复杂。随着研究者认识的深入, 尤其是食物网理论发展的需要, 亟需加强氮沉降对草地生物多营养级效应的探索。第三, 氮沉降可以通过改变土壤理化性质对生活在其中的昆虫类群产生影响, 然而氮沉降对土壤昆虫影响的研究比较少见。土壤昆虫与很多地下生态学过程有关, 在维持生态系统养分循环和能量流动方面发挥重要作用。未来应加强氮沉降对土壤昆虫以及地下食物网的影响及调控机制的研究。第四, 功能性状决定了生物的能量代谢、生长、繁殖、扩散等一系列属性并且可以有效指示环境变化或干扰对生物群落组成的影响, 对于研究群落构建机制具有重要的理论意义, 因此基于性状的研究方法(trait-based approaches)在过去几年发展迅速。但关于氮沉降如何影响昆虫功能性状还尚不清楚, 未来应该加强昆虫关键功能性状(如体型大小和移动特性等)对氮沉降响应的研究。第五, 未来的研究应更多考虑氮沉降的长期效应以及更大空间尺度的作用效应。因为氮沉降对昆虫的影响存在滞后效应, 尤其是高营养级昆虫物种对氮沉降的响应可能需要更长的时间才能显现。同时模拟氮沉降的处理多位于面积较小的样地内, 难以判定氮沉降对昆虫群落的影响是来自昆虫短距离内随机迁移的结果还是昆虫种群数量实际发生的变化。第六, 氮沉降和其他全球变化因子(如CO2浓度升高、增温、降水变化等)存在复杂的交互作用。例如, 氮沉降和大气CO2浓度增高的协同作用可以使植物产生更高的化学防御水平(同时增加氮基次级代谢物和碳基次级代谢物), 从而对植食性昆虫产生潜在的影响。氮沉降和增温的交互作用可能会加速昆虫的发育速率, 导致昆虫种群数量的暴发。因此未来需要综合考虑多种全球变化因子对草地昆虫群落的影响及机制。
附录 Supplementary Material
附录1 氮沉降对草地昆虫群落的作用及其影响因素
Appendix 1 Effects of nitrogen deposition on grassland insect community and influence factors
参考文献
Global estimates of inorganic nitrogen deposition across four decades
Effects of aphid-tending Argentine ants, nitrogen enrichment and early-season herbivory on insects hosted by a coastal shrub
DOI:10.1007/s10530-008-9223-4 URL [本文引用: 2]
Spatial-temporal distribution of the Hymenoptera in the Brazilian Savanna and the effects of habitat heterogeneity on these patterns
DOI:10.1007/s10841-015-9832-z URL [本文引用: 1]
Long-term nutrient addition alters arthropod community composition but does not increase total biomass or abundance
DOI:10.1111/oik.2017.v127.i3 URL [本文引用: 6]
Toward a world that values insects
DOI:10.1126/science.aaw7071 PMID:31249044 [本文引用: 1]
Insect herbivory accelerates nutrient cycling and increases plant production
Edge effects on insect-plant food webs: Assessing the influence of geographical orientation and microclimatic conditions
DOI:10.1111/een.v45.4 URL [本文引用: 1]
Nitrogen deposition and climate: An integrated synthesis
DOI:10.1016/j.tree.2022.02.013 URL [本文引用: 1]
Negative impact of nitrogen deposition on soil buffering capacity
Helicoverpa zea (Lepidoptera: Noctuidae) preference for plant structures, and their location, within Bt cotton under different nitrogen and irrigation regimes
DOI:10.1093/jee/toz105
PMID:31329903
[本文引用: 1]

Helicoverpa zea Boddie is a common economic pest of cotton (Gossypium hirsutum L.), including transgenic cotton varieties that express Bacillus thuringiensis (Bt). Helicoverpa zea oviposition is similar in Bt and non-Bt cotton, but behavior of H. zea larvae can be different in the presence of Bt, with neonates moving away from terminals faster in single-toxin Bt than non-Bt cotton or avoiding Bt-treated diet in the lab. We quantified H. zea oviposition and larval distribution on structures within cotton plants in small plot experiments of Cry1Ac + Cry1F cotton for 2 yr under different irrigation and nitrogen treatments. More eggs were oviposited on plants receiving nitrogen application during 2016 and on leaves in the top section of irrigated plants during 2017, but other treatment effects on eggs or larvae were minimal. Helicoverpa zea eggs were most common on leaves in the top third of plants at position zero and middle section of cotton plants throughout the season, but some oviposition occurred on fruiting structures as well. First and second instars were more common on squares in the top section of plants during 2016 and bolls in the middle and lower sections during 2017 due to oviposition lower in the canopy during 2017. During both years, third through fifth instars were more common on bolls in the middle and lower section of plants closer to the main stem. These findings have resistance management implications as extended larval feeding on bolls could optimize nutrition, decrease Bt susceptibility, and potentially influence behavioral resistance.© The Author(s) 2019. Published by Oxford University Press on behalf of Entomological Society of America. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment
We provide a global assessment, with detailed multi-scale data, of the ecological and toxicological effects generated by inorganic nitrogen pollution in aquatic ecosystems. Our synthesis of the published scientific literature shows three major environmental problems: (1) it can increase the concentration of hydrogen ions in freshwater ecosystems without much acid-neutralizing capacity, resulting in acidification of those systems; (2) it can stimulate or enhance the development, maintenance and proliferation of primary producers, resulting in eutrophication of aquatic ecosystems; (3) it can reach toxic levels that impair the ability of aquatic animals to survive, grow and reproduce. Inorganic nitrogen pollution of ground and surface waters can also induce adverse effects on human health and economy. Because reductions in SO2 emissions have reduced the atmospheric deposition of H2SO4 across large portions of North America and Europe, while emissions of NOx have gone unchecked, HNO3 is now playing an increasing role in the acidification of freshwater ecosystems. This acidification process has caused several adverse effects on primary and secondary producers, with significant biotic impoverishments, particularly concerning invertebrates and fishes, in many atmospherically acidified lakes and streams. The cultural eutrophication of freshwater, estuarine, and coastal marine ecosystems can cause ecological and toxicological effects that are either directly or indirectly related to the proliferation of primary producers. Extensive kills of both invertebrates and fishes are probably the most dramatic manifestation of hypoxia (or anoxia) in eutrophic and hypereutrophic aquatic ecosystems with low water turnover rates. The decline in dissolved oxygen concentrations can also promote the formation of reduced compounds, such as hydrogen sulphide, resulting in higher adverse (toxic) effects on aquatic animals. Additionally, the occurrence of toxic algae can significantly contribute to the extensive kills of aquatic animals. Cyanobacteria, dinoflagellates and diatoms appear to be major responsible that may be stimulated by inorganic nitrogen pollution. Among the different inorganic nitrogenous compounds (NH4+, NH3, NO2-, HNO2NO3-) that aquatic animals can take up directly from the ambient water, unionized ammonia is the most toxic, while ammonium and nitrate ions are the least toxic. In general, seawater animals seem to be more tolerant to the toxicity of inorganic nitrogenous compounds than freshwater animals, probably because of the ameliorating effect of water salinity (sodium, chloride, calcium and other ions) on the tolerance of aquatic animals. Ingested nitrites and nitrates from polluted drinking waters can induce methemoglobinemia in humans, particularly in young infants, by blocking the oxygen-carrying capacity of hemoglobin. Ingested nitrites and nitrates also have a potential role in developing cancers of the digestive tract through their contribution to the formation of nitrosamines. In addition, some scientific evidences suggest that ingested nitrites and nitrates might result in mutagenicity, teratogenicity and birth defects, contribute to the risks of non-Hodgkin's lymphoma and bladder and ovarian cancers, play a role in the etiology of insulin-dependent diabetes mellitus and in the development of thyroid hypertrophy, or cause spontaneous abortions and respiratory tract infections. Indirect health hazards can occur as a consequence of algal toxins, causing nausea, vomiting, diarrhoea, pneumonia, gastroenteritis, hepatoenteritis, muscular cramps, and several poisoning syndromes (paralytic shellfish poisoning, neurotoxic shellfish poisoning, amnesic shellfish poisoning). Other indirect health hazards can also come from the potential relationship between inorganic nitrogen pollution and human infectious diseases (malaria, cholera). Human sickness and death, extensive kills of aquatic animals, and other negative effects, can have elevated costs on human economy, with the recreation and tourism industry suffering the most important economic impacts, at least locally. It is concluded that levels of total nitrogen lower than 0.5-1.0 mg TN/L could prevent aquatic ecosystems (excluding those ecosystems with naturally high N levels) from developing acidification and eutrophication, at least by inorganic nitrogen pollution. Those relatively low TN levels could also protect aquatic animals against the toxicity of inorganic nitrogenous compounds since, in the absence of eutrophication, surface waters usually present relatively high concentrations of dissolved oxygen, most inorganic reactive nitrogen being in the form of nitrate. Additionally, human health and economy would be safer from the adverse effects of inorganic nitrogen pollution.
Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content
DOI:10.1126/science.1214433
PMID:22282812
[本文引用: 4]
Current paradigms generally assume that increased plant nitrogen (N) should enhance herbivore performance by relieving protein limitation, increasing herbivorous insect populations. We show, in contrast to this scenario, that host plant N enrichment and high-protein artificial diets decreased the size and viability of Oedaleus asiaticus, a dominant locust of north Asian grasslands. This locust preferred plants with low N content and artificial diets with low protein and high carbohydrate content. Plant N content was lowest and locust abundance highest in heavily livestock-grazed fields where soils were N-depleted, likely due to enhanced erosion. These results suggest that heavy livestock grazing and consequent steppe degradation in the Eurasian grassland promote outbreaks of this locust by reducing plant protein content.
Effects of environmental moisture and precipitation on insects: A review
Climatic factors are closely related to the individual’s development and growth, and the population dynamics of insect pests. Environmental moisture (including atmospheric relative humidity and soil moisture content) can directly cause the changes of insects’ water content and destroy their water balance in vivo, and consequently, affects their individual’s development and populationoccurrence. As resulting in the changes of environmental moisture, precipitation can also affects field population dynamics of insects by physically washing out. Therefore, the studies on the impacts of environmental moisture and precipitation on insects have a wider significance for the integrated pest management (IPM). This paper reviewed the impacts of environmental moisture and precipitation, as well as other environmental factors (<EM>e.g</EM>., temperature), on the growth and development, survival, behavior, reproduction, and population ecology of insects, and introduced the applied studies on the regulation of environmental moisture (<EM>e.g</EM>., via irrigation) to control the occurrences of insect pests (<EM>e.g</EM>., <EM>Helicoverpa armigera</EM>) in fields.
环境湿度和降雨对昆虫的影响
Advances in ecological stoichiometry in typically and ecologically vulnerable regions of China
中国典型生态脆弱区生态化学计量学研究进展
Effects of nitrogen fertilization on tritrophic interactions
DOI:10.1007/s11829-010-9092-5 URL [本文引用: 1]
Short- and medium-term effects of experimental nitrogen fertilization on arthropods associated with Calluna vulgaris heathlands in north-west Spain
We studied the short- and medium-term effects of experimental nitrogen fertilization (3 and 15 months after the treatment) on the arthropods of Calluna vulgaris heathlands in NW Spain. Three heathland sites were selected with two permanent plots per site: control and fertilized. Ammonium nitrate fertilizer (56 kg N ha(-1) yr(-1)) was applied monthly and insects were caught using pitfall traps. We found mainly species-level responses to nitrogen addition. Seven species (e.g. Lochmaea suturalis) showed a consistent trend (benefited or harmed) in both periods and were proposed as possible reliable indicators of the effects of nitrogen deposition in these ecosystems. We also found variable arthropod trophic-group responses: (a) herbivores (leaf beetles, true bugs) increased in abundance on a short-term scale; (b) predators (carabid beetles, true bugs) showed opposite and less clear responses in both periods. Further long-term studies are needed to determine the mechanisms underlying the observed arthropod responses.
Abundance of gall-inducing insect species in sclerophyllous savanna: Understanding the importance of soil fertility using an experimental approach
DOI:10.1017/S0266467411000368
URL
[本文引用: 1]
Although many studies have now demonstrated that both richness and abundance of gall-inducing insect species are directly and indirectly (via the host plant) influenced by soil quality, the empirical evaluation of it in the field remains anecdotal at best. The effects of soil fertility on richness and abundance of gall-inducing insects associated with a widespread savanna species, Eremanthus glomerulatus, were evaluated under experimental field conditions in Brasilia, central Brazil. The effect of soil fertility on gall-inducing insects species richness was evaluated using three treatments: (1) plots fertilized with nitrogen; (2) plots fertilized with phosphorus; and (3) control plots: soils without fertilization. Species richness of gall-inducing insects (six species of Cecidomyiidae) did not differ among the treatments. Leaves with galls had higher nitrogen concentrations (mean = 15.0 ± 0.5 mg g−1), compared with leaves without galls (mean = 9.0 ± 0.7 mg g−1) on plants that occurred in soils with addition of nitrogen. Similarly, leaves with galls had higher foliar phosphorus concentration (mean = 1.0 ± 0.04 mg g−1) than leaves without galls (mean = 0.6 ± 0.05 mg g−1) in plots with addition of phosphorus. In galled leaves, a negative relationship between gall density and nitrogen concentration was found for one gall-inducing insect species, while three species showed a positive relationship between gall density and leaf nitrogen concentration. A negative relationship between gall density and concentration of leaf phosphorus was observed for four of the six gall-inducing insect species studied. No relationship was found between gall density and leaf nitrogen and phosphorus concentration in ungalled leaves. We argue that foliar nitrogen and phosphorus concentration respond to gall density in galled leaves and therefore, gall-inducing insect species are capable of manipulating their host plant, modifying the foliar nutrients of E. glomerulatus in sclerophyllous savanna.
Understanding how changing soil nitrogen affects plant-pollinator interactions
DOI:10.1007/s11829-019-09714-y [本文引用: 3]
Plant-mediated and nonadditive effects of two global change drivers on an insect herbivore community
Warmer temperatures can alter the phenology and distribution of individual species. However, differences across species may blur community-level phenological responses to climate or cause biotic homogenization by consistently favoring certain taxa. Additionally, the response of insect communities to climate will be subject to plant-mediated effects, which may or may not overshadow the direct effect of rising temperatures on insects. Finally, recent evidence for the importance of interaction effects between global change drivers suggests that phenological responses of communities to climate may be altered by other drivers. We used a natural temperature gradient (generated by elevation and topology), combined with experimental nitrogen fertilization, to investigate the effects of elevated temperature and globally increasing anthropogenic nitrogen deposition on the structure and phenology of a seminatural grassland herbivore assemblage (lepidopteran insects). We found that both drivers, alone and in combination, severely altered how the relative abundance and composition of species changed through time. Importantly, warmer temperatures were associated with biotic homogenization, such that herbivore assemblages in the warmest plots had more similar species composition than those in intermediate or cool plots. Changes in herbivore composition and abundance were largely mediated by changes in the plant community, with increased nonnative grass cover under high treatment levels being the strongest determinant of herbivore abundance. In addition to compositional changes, total herbivore biomass more than doubled under elevated nitrogen and increased more than fourfold with temperature, bearing important functional implications for herbivores as consumers and as a prey resource. The crucial role of nonnative plant dominance in mediating responses of herbivores to change, combined with the frequent nonadditive (positive and negative) effects of the two drivers, and the differential responses of species, highlight that understanding complex ecosystem responses will benefit from multifactor, multitrophic experiments at community scales or larger.
Climate change disproportionately increases herbivore over plant or parasitoid biomass
Nitrogen fixation: Anthropogenic enhancement-environmental response
DOI:10.1029/95GB00158
URL
[本文引用: 1]
In the absence of human activities, biotic fixation is the primary source of reactive N, providing about 90–130 Tg N yr−1 (Tg = 1012 g) on the continents. Human activities have resulted in the fixation of an additional ≈140 Tg N yr−1 by energy production (≈20 Tg N yr−1 ), fertilizer production (≈80 Tg N yr−1), and cultivation of crops (e.g., legumes, rice) (≈40 Tg N yr−1 ). We can only account for part of this anthropogenic N. N2O is accumulating in the atmosphere at a rate of 3 Tg N yr−1. Coastal oceans receive another 41 Tg N yr−1 via rivers, much of which is buried or denitrified. Open oceans receive 18 Tg N yr−1 by atmospheric deposition, which is incorporated into oceanic N pools (e.g., NO3−, N2). The remaining 80 Tg N yr−1 are either retained on continents in groundwater, soils, or vegetation or denitrified to N2. Field studies and calculations indicate that uncertainties about the size of each sink can account for the remaining anthropogenic N. Thus although anthropogenic N is clearly accumulating on continents, we do not know rates of individual processes. We predict the anthropogenic N‐fixation rate will increase by about 60% by the year 2020, primarily due to increased fertilizer use and fossil‐fuel combustion. About two‐thirds of the increase will occur in Asia, which by 2020 will account for over half of the global anthropogenic N fixation.
Ecological mechanisms and phylogeny shape invertebrate stoichiometry: A test using detritus-based communities across Central and South America
DOI:10.1111/fec.2018.32.issue-10 URL [本文引用: 1]
Nitrogen deposition and its contribution to nitrogen cycling and associated soil processes
DOI:10.1046/j.1469-8137.1998.00182.x
URL
[本文引用: 1]
Human activity has greatly perturbed the nitrogen cycle through increased fixation by legumes, by energy and fertilizer production, and by the mobilization of N from long‐term storage pools. This extra reactive N is readily transported through the environment, and there is increasing evidence that it is changing ecosystems through eutrophication and acidification. Rothamsted Experimental Station, UK has been involved in research on N cycling in ecosystems since its inception in 1843. Measurements of precipitation composition at Rothamsted, made since 1853, show an increase of nitrate and ammonium N in precipitation from 1 and 3 kg N ha−1 yr−1, respectively, in 1855 to a maximum of 8 and 10 kg N ha−1 yr−1 in 1980, decreasing to 4 and 5 kg N ha−1 yr−1 today. Nitrogen inputs via dry deposition do, however, remain high. Recent measurements with diffusion tubes and filter packs show large concentrations of nitrogen dioxide of c. 20 μg m−3 in winter and c. 10 μg m−3 in summer; the difference is linked to the use of central heating, and with variations in wind direction and pollutant source. Concentrations of nitric acid and particulate N exhibit maxima of 1·5 and 2 μg m−3 in summer and winter, respectively. Concentrations of ammonia are small, barely rising above 1 μg m−3.
Seasonal shift from bottom-up to top-down impact in phytophagous insect populations
Although many studies now examine how multiple factors influence the dynamics of herbivore populations, few studies explicitly attempt to document where and when each is important and how they vary and interact. In fact, how temporal variation in top-down (natural enemies) and bottom-up (host plant resources) factors affect herbivore dynamics has been suggested as a particularly important yet poorly understood feature of terrestrial food webs. In this study we examined how temporal changes in predator density (wolf spiders, sheet-web builders, and mirid egg predators) and host-plant resources (plant quality and structural complexity) influence the population dynamics of the dominant phytophagous insects on Atlantic-coast salt marshes, namely Prokelisia planthoppers (Homoptera: Delphacidae). We designed a factorial experiment in meadows of Spartina alterniflora to mimic natural variation in vegetation quality and structure by establishing two levels of plant nutrition (leaf nitrogen content) by fertilization, and two levels of habitat complexity by adding leaf litter (thatch). We then assessed seasonal changes in the strength of bottom-up (plant quality) and top-down (predator) impacts on planthopper populations. Planthopper populations responded positively to increased plant quality treatments in late summer. Despite the greater number of planthopper adults colonizing fertilized Spartina plots compared to unfertilized controls, the offspring of these colonists were much less abundant at the end of the season in fertilized plots, particularly those with thatch. The initial colonization effect was later erased because arthropod predators selectively accumulated in fertilized plots where they inflicted significant mortality on all stages of planthoppers. Predators rapidly colonized fertilized plots and reached high densities well in advance of planthopper colonization, a response we attribute to their rapid aggregation in complex-structured habitats with readily available alternative prey. Our results suggest that plant resources not only mediate the strength of predator impacts on herbivore populations, but they also promote the coupling of predator and prey populations and thus influence when enemy impacts are realized.
Chronic, low-level nitrogen deposition enhances abundances of ant-protected herbivores inhabiting an imperiled foundation species
DOI:10.1016/j.actao.2021.103706 URL [本文引用: 1]
Plant species loss decreases arthropod diversity and shifts trophic structure
DOI:10.1111/j.1461-0248.2009.01356.x
PMID:19702636
[本文引用: 1]
Plant diversity is predicted to be positively linked to the diversity of herbivores and predators in a foodweb. Yet, the relationship between plant and animal diversity is explained by a variety of competing hypotheses, with mixed empirical results for each hypothesis. We sampled arthropods for over a decade in an experiment that manipulated the number of grassland plant species. We found that herbivore and predator species richness were strongly, positively related to plant species richness, and that these relationships were caused by different mechanisms at herbivore and predator trophic levels. Even more dramatic was the threefold increase, from low- to high-plant species richness, in abundances of predatory and parasitoid arthropods relative to their herbivorous prey. Our results demonstrate that, over the long term, the loss of plant species propagates through food webs, greatly decreasing arthropod species richness, shifting a predator-dominated trophic structure to being herbivore dominated, and likely impacting ecosystem functioning and services.
The effects of long-term nitrogen loading on grassland insect communities
DOI:10.1007/s004420050026
PMID:28308414
[本文引用: 5]
Just as long-term nitrogen loading of grasslands decreases plant species richness and increases plant biomass, we have found that nitrogen loading decreases insect species richness and increases insect abundances. We sampled 54 plots that had been maintained at various rates of nitrogen addition for 14 years. Total insect species richness and effective insect diversity, as well as herbivore and predator species richness, were significantly, negatively related to the rate of nitrogen addition. However, there was variation in trophic responses to nitrogen. Detritivore species richness increased as nitrogen addition increased, and parasitoids showed no response. Insect abundances, measured as the number of insects and insect biovolume (an estimate of biomass), were significantly, positively related to the rate of nitrogen addition, as were the abundances of herbivores and detritivores. Parasitoid abundance was negatively related to the rate of nitrogen addition. Changes in the insect community were correlated with changes in the plant community. As rates of nitrogen addition increased, plant species richness decreased, plant productivity and plant tissue nitrogen increased, and plant composition shifted from C to C grass species. Along this gradient, total insect species richness and effective insect diversity were most strongly, positively correlated with plant species richness. Insect biovolume was negatively correlated with plant species richness. Responses of individual herbivores varied along the nitrogen gradient, but numbers of 13 of the 18 most abundant herbivores were positively correlated with their host plant biomass. Although insect communities did not respond as strongly as plant communities, insect species richness, abundance, and composition were impacted by nitrogen addition. This study demonstrates that long-term nitrogen loading affects the entire food chain, simplifying both plant and insect communities.
Effects of atmospheric nitrogen deposition on net primary productivity of grassland ecosystem in Central Asia
大气氮沉降对中亚草地生态系统净初级生产力的影响
Fertilizer application decreases insect abundance on Plantago lanceolata: A large-scale experiment in three geographic regions
DOI:10.1007/s11829-012-9237-9 URL [本文引用: 1]
Relative humidity effects on the life history of Hermetia illucens (Diptera: Stratiomyidae)
DOI:10.1603/EN12054 URL [本文引用: 1]
Base cation depletion, eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition
DOI:10.1016/j.envpol.2007.11.006
PMID:18164110
[本文引用: 1]
Pollutant nitrogen deposition effects on soil and foliar element concentrations were investigated in acidic and limestone grasslands, located in one of the most nitrogen and acid rain polluted regions of the UK, using plots treated for 8-10 years with 35-140 kg N ha(-2)y(-1) as NH(4)NO(3). Historic data suggests both grasslands have acidified over the past 50 years. Nitrogen deposition treatments caused the grassland soils to lose 23-35% of their total available bases (Ca, Mg, K, and Na) and they became acidified by 0.2-0.4 pH units. Aluminium, iron and manganese were mobilised and taken up by limestone grassland forbs and were translocated down the acid grassland soil. Mineral nitrogen availability increased in both grasslands and many species showed foliar N enrichment. This study provides the first definitive evidence that nitrogen deposition depletes base cations from grassland soils. The resulting acidification, metal mobilisation and eutrophication are implicated in driving floristic changes.
Consequences of nitrogen and phosphorus limitation for the performance of two planthoppers with divergent life-history strategies
Phytophagous insects have a much higher nitrogen and phosphorus content than their host plants, an elemental mismatch that places inherent constraints on meeting nutritional requirements. Although nitrogen limitation is well documented in insect herbivores, phosphorus limitation is poorly studied. Using factorial experiments in the laboratory and field, in which levels of soil nitrogen and phosphorus were manipulated, we studied the relative consequences of macronutrient limitation for two herbivores, namely the phloem-feeding planthoppers Prokelisia dolus and P. marginata. These planthoppers inhabit the salt marshes of North America where large stands of their Spartina host plant are found. Notably, these congeners differ in their dispersal abilities; P. marginata is dispersive whereas P. dolus is sedentary. Both nitrogen and phosphorus subsidies enhanced the nitrogen and phosphorus content of Spartina. When P. dolus and P. marginata were raised on plants with an enriched nitrogen signature, they exhibited greater survival, grew to a larger size, developed more rapidly, and achieved higher densities than on nitrogen-deficient plants. However, P. marginata experienced greater fitness penalties than P. dolus on nitrogen-deficient plants. Phosphorus limitation and associated fitness penalties were not as severe as nitrogen limitation for P. marginata, and were not detected in P. dolus. The tempered response of P. dolus to N- and P-deficient Spartina is probably due to its greater investment in feeding musculature and hence ability to compensate for nutrient deficiencies with increased ingestion. To cope with deteriorating plant quality, P. dolus employs compensatory feeding, whereas P. marginata disperses to higher quality Spartina. When its option of dispersal is eliminated and P. marginata is confined on nutrient-deficient plants, its performance is drastically reduced compared with P. dolus. This research highlights the importance of interfacing herbivore life-history strategies with ecological stoichiometry in order to interpret the consequences of macronutrient limitation on herbivore performance and population dynamics.
Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity
Grazing intensity affects insect diversity via sward structure and heterogeneity in a long-term experiment
DOI:10.1111/jpe.2014.51.issue-4 URL [本文引用: 2]
Ecological mechanisms underlying arthropod species diversity in grasslands
DOI:10.1146/annurev-ento-120811-153540
PMID:22830354
[本文引用: 1]
Arthropods are an important component of grassland systems, contributing significantly to biodiversity and ecosystem structure and function. Climate, fire, and grazing by large herbivores are important drivers in grasslands worldwide. Arthropod responses to these drivers are highly variable and clear patterns are difficult to find, but responses are largely indirect with respect to changes in resources, species interactions, habitat structure, and habitat heterogeneity resulting from interactions among fire, grazing, and climate. Here, we review these ecological mechanisms influencing grassland arthropod diversity. We summarize hypotheses describing species diversity at local and regional scales and then discuss specific factors that may affect arthropod diversity in grassland systems. These factors include direct and indirect effects of grazing, fire, and climate, species interactions, above- and belowground interactions, and landscape-level effects.
Plant zonation on lakeshores in Nova Scotia: A test of the resource specialization hypothesis
DOI:10.2307/2259532 URL [本文引用: 1]
Impact of nitrogen deposition on larval habitats: The case of the Wall Brown butterfly Lasiommata megera
DOI:10.1007/s10841-014-9748-z URL [本文引用: 1]
Plant architecture and the diversity of phytophagous insects
DOI:10.1146/ento.1983.28.issue-1 URL [本文引用: 1]
Impact of acidification and eutrophication on the distribution of fish species in shallow and lentic soft waters of the Netherlands: An historical perspective
DOI:10.1111/jfb.1987.31.issue-6 URL [本文引用: 1]
Responses of tree and insect herbivores to elevated nitrogen inputs: A meta-analysis
DOI:10.1016/j.actao.2016.10.008 URL [本文引用: 1]
Enhanced nitrogen deposition over China
DOI:10.1038/nature11917 [本文引用: 1]
The economic value of ecological services provided by insects
DOI:10.1641/0006-3568(2006)56[311:TEVOES]2.0.CO;2 URL [本文引用: 1]
Precipitation variations, rather than N deposition, determine plant ecophysiological traits in a desert steppe in Northern China
DOI:10.1016/j.ecolind.2022.109144 URL [本文引用: 1]
Herbivory in relation to plant nitrogen content
DOI:10.1146/ecolsys.1980.11.issue-1 URL [本文引用: 2]
A test for clinal variation in Artemisia californica and associated arthropod responses to nitrogen addition
Nutrient presses and pulses differentially impact plants, herbivores, detritivores and their natural enemies
Nitrogen and phosphorus enrichment cause declines in invertebrate populations: A global meta-analysis
DOI:10.1111/brv.v96.6 URL [本文引用: 2]
Pathways for the effects of increased nitrogen deposition on fauna
DOI:10.1016/j.biocon.2017.02.022 URL [本文引用: 11]
The relationship between local extinctions of grassland butterflies and increased soil nitrogen levels
DOI:10.1016/j.biocon.2005.10.024 URL [本文引用: 1]
Effects of acidic water on freshwater snails: Results from a study of 1000 lakes throughout Norway
This paper summarizes some results from a monograph which focused on ecological and biogeographical aspects of the biota in Norwegian lakes and rivers with particular reference to the snail fauna. Field studies were made in the course of 20 summers within the period of 1953-1987. Acidic water is the main reason why snails are absent from some 4000 lakes in a low-calcium area in southern Norway ('Acid south'). This area is heavily influenced by acidic precipitation. Snails were not detected in lakes with pH below 5.2. In one lake, three snail species disappeared in parallel with acidification from pH 5.2 to 4.2. The calcium level modified the effect of low pH.
Effects of plant diversity, plant productivity and habitat parameters on arthropod abundance in montane European grasslands
DOI:10.1111/eco.2005.28.issue-4 URL [本文引用: 1]
To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs
DOI:10.1111/j.1600-0706.2009.18545.x URL [本文引用: 1]
Global pollinator declines: Trends, impacts and drivers
DOI:10.1016/j.tree.2010.01.007 URL [本文引用: 1]
Different responses of plants and herbivore insects to a gradient of vegetation height: An indicator of the vertebrate grazing intensity and successional age
DOI:10.1111/oik.2006.115.issue-3 URL [本文引用: 1]
Trophic differences regulate grassland food webs: Herbivores track food quality and predators select for habitat volume
The influence of nitrogenous fertilizer on the grassland Auchenorrhyncha (Homoptera)
DOI:10.2307/2403278 URL [本文引用: 1]
The geometry of compensatory feeding in the locust
DOI:10.1006/anbe.1993.1114 URL [本文引用: 1]
Organismal stoichiometry: Quantifying non-independence among food components
DOI:10.1890/02-0248 URL [本文引用: 1]
How do nutrients and warming impact on plant communities and their insect herbivores? A 9-year study from a sub-Arctic heath
DOI:10.1046/j.1365-2745.2002.00681.x URL [本文引用: 1]
Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities
DOI:10.1016/S0006-3207(99)00049-X URL [本文引用: 1]
Negative effects of nitrogen deposition on Swiss butterflies
DOI:10.1111/cobi.13744
PMID:33829544
[本文引用: 4]
Nitrogen (N) deposition caused by agriculture and combustion of fossil fuels is a major threat to plant diversity, but the effects on higher trophic levels are less clear. In this study we investigated how N deposition may affect species richness and abundance (number of individuals per species) in butterflies. We started with reviewing the literature and found that vegetation parameters might be as important as climate and habitat variables in explaining variation in butterfly species richness. It thus seems likely that increased N deposition indirectly affects butterfly communities via its influence on plant communities. To test this prediction, we analyzed data from the Swiss biodiversity monitoring program surveying species diversity of vascular plants and butterflies in 383 study sites of 1 km that are regularly distributed over Switzerland, covering a modelled N deposition gradient from 2 to 44 kg N ha yr. Using traditional linear models and structural equation models, we found that high N deposition was consistently linked to low butterfly diversity, suggesting a net loss of butterfly diversity through increased N deposition. At low elevations, N deposition may contribute to a reduction in butterfly species richness via microclimatic cooling due to increased plant biomass. At higher elevations, negative effects of N deposition on butterfly species richness may also be mediated by reduced plant species richness. In most butterfly species, abundance was negatively related to N deposition, but the strongest negative effects were found for species of conservation concern. We conclude that in addition to factors such as intensified agriculture, habitat fragmentation and climate change, N deposition is likely to play a key role in negatively affecting butterfly diversity and abundance. Article Impact Statement: Nitrogen deposition negatively affects butterfly species richness and butterfly abundance, particularly in species of conservation concern. This article is protected by copyright. All rights reserved.This article is protected by copyright. All rights reserved.
Enemies hypothesis: A review of the effect of vegetational diversity on predatory insects and parasitoids
Worldwide decline of the entomofauna: A review of its drivers
DOI:10.1016/j.biocon.2019.01.020 URL [本文引用: 1]
Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment
DOI:10.1038/nature09492 [本文引用: 1]
Adult tea green leafhoppers, Empoasca onukii (Matsuda), change behaviors under varying light conditions
The Park Grass Experiment 1856-2006: Its contribution to ecology
DOI:10.1111/jec.2006.94.issue-4 URL [本文引用: 2]
Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States
Why more productive sites have more species: An experimental test of theory using tree-hole communities
DOI:10.1086/286187 URL [本文引用: 1]
Atmospheric nitrogen deposition in terrestrial ecosystems: Its impact on plant communities and consequences across trophic levels
DOI:10.1111/fec.2018.32.issue-7 URL [本文引用: 1]
Impact of nitrogen deposition on the species richness of grasslands
A transect of 68 acid grasslands across Great Britain, covering the lower range of ambient annual nitrogen deposition in the industrialized world (5 to 35 kg Nha-1 year-1), indicates that long-term, chronic nitrogen deposition has significantly reduced plant species richness. Species richness declines as a linear function of the rate of inorganic nitrogen deposition, with a reduction of one species per 4-m2 quadrat for every 2.5 kg Nha-1 year-1 of chronic nitrogen deposition. Species adapted to infertile conditions are systematically reduced at high nitrogen deposition. At the mean chronic nitrogen deposition rate of central Europe (17 kg Nha-1 year-1), there is a 23% species reduction compared with grasslands receiving the lowest levels of nitrogen deposition.
The role of a mound-building ecosystem engineer for a grassland butterfly
DOI:10.1007/s10841-014-9670-4 URL [本文引用: 1]
Progress in research on the responses of insects to global climate change
昆虫对全球气候变化的响应与适应性
Comparative losses of British butterflies, birds, and plants and the global extinction crisis
Effects of nitrogen deposition on insect herbivory: Implications for community and ecosystem processes
Below- ground-mediated and phase-dependent processes drive nitrogen-evoked community changes in grasslands
DOI:10.1111/jec.v108.5 URL [本文引用: 1]
Integration of sheep production and nature conservation: Experimental management
DOI:10.1006/jema.1996.0102 URL [本文引用: 1]
Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances
DOI:10.1126/science.aax9931
PMID:32327596
[本文引用: 2]
Recent case studies showing substantial declines of insect abundances have raised alarm, but how widespread such patterns are remains unclear. We compiled data from 166 long-term surveys of insect assemblages across 1676 sites to investigate trends in insect abundances over time. Overall, we found considerable variation in trends even among adjacent sites but an average decline of terrestrial insect abundance by ~9% per decade and an increase of freshwater insect abundance by ~11% per decade. Both patterns were largely driven by strong trends in North America and some European regions. We found some associations with potential drivers (e.g., land-use drivers), and trends in protected areas tended to be weaker. Our findings provide a more nuanced view of spatiotemporal patterns of insect abundance trends than previously suggested.Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Effects of large herbivores on grassland arthropod diversity
DOI:10.1111/brv.2015.90.issue-2 URL [本文引用: 2]
Life-history strategies as a tool to identify conservation constraints: A case-study on ants in chalk grasslands
DOI:10.1016/j.ecolind.2011.06.028 URL [本文引用: 1]
Insect declines in the Anthropocene
DOI:10.1146/annurev-ento-011019-025151
PMID:31610138
[本文引用: 1]
Insect declines are being reported worldwide for flying, ground, and aquatic lineages. Most reports come from western and northern Europe, where the insect fauna is well-studied and there are considerable demographic data for many taxonomically disparate lineages. Additional cases of faunal losses have been noted from Asia, North America, the Arctic, the Neotropics, and elsewhere. While this review addresses both species loss and population declines, its emphasis is on the latter. Declines of abundant species can be especially worrisome, given that they anchor trophic interactions and shoulder many of the essential ecosystem services of their respective communities. A review of the factors believed to be responsible for observed collapses and those perceived to be especially threatening to insects form the core of this treatment. In addition to widely recognized threats to insect biodiversity, e.g., habitat destruction, agricultural intensification (including pesticide use), climate change, and invasive species, this assessment highlights a few less commonly considered factors such as atmospheric nitrification from the burning of fossil fuels and the effects of droughts and changing precipitation patterns. Because the geographic extent and magnitude of insect declines are largely unknown, there is an urgent need for monitoring efforts, especially across ecological gradients, which will help to identify important causal factors in declines. This review also considers the status of vertebrate insectivores, reporting bias, challenges inherent in collecting and interpreting insect demographic data, and cases of increasing insect abundance.
Insect decline in the Anthropocene: Death by a thousand cuts
Global warming and excess nitrogen may induce butterfly decline by microclimatic cooling
DOI:10.1111/gcb.2006.12.issue-9 URL [本文引用: 1]
Changes in nectar supply: A possible cause of widespread butterfly decline
DOI:10.1093/czoolo/58.3.384
URL
[本文引用: 1]
Recent studies have documented declining trends of various groups of flower-visiting insects, even common butterfly species. Causes of these declines are still unclear but the loss of habitat quality across the wider countryside is thought to be a major factor. Nectar supply constitutes one of the main resources determining habitat quality. Yet, data on changes in nectar abundance are lacking. In this study, we provide the first analysis of changes in floral nectar abundance on a national scale and link these data to trends in butterfly species richness and abundance. We used transect data from the Dutch Butterfly Monitoring Scheme to compare two time periods: 1994-1995 and 2007-2008. The results show that butterfly decline can indeed be linked to a substantial decline in overall flower abundance and specific nectar plants, such as thistles. The decline is as severe in reported flower generalists as in flower specialists. We suggest that eutrophication is a main cause of the decline of nectar sources.
Plant genetic diversity affects multiple trophic levels and trophic interactions
DOI:10.1038/s41467-022-35087-7
[本文引用: 2]
Intraspecific genetic diversity is an important component of biodiversity. A substantial body of evidence has demonstrated positive effects of plant genetic diversity on plant performance. However, it has remained unclear whether plant genetic diversity generally increases plant performance by reducing the pressure of plant antagonists across trophic levels for different plant life forms, ecosystems and climatic zones. Here, we analyse 4702 effect sizes reported in 413 studies that consider effects of plant genetic diversity on trophic groups and their interactions. We found that that increasing plant genetic diversity decreased the performance of plant antagonists including invertebrate herbivores, weeds, plant-feeding nematodes and plant diseases, while increasing the performance of plants and natural enemies of herbivores. Structural equation modelling indicated that plant genetic diversity increased plant performance partly by reducing plant antagonist pressure. These results reveal that plant genetic diversity often influences multiple trophic levels in ways that enhance natural pest control in managed ecosystems and consumer control of plants in natural ecosystems for sustainable plant production.
Progress in the coadaptation and coevolution between plants and herbivores
植物与草食动物之间的协同适应及进化
What are the biological bases of varied diets?
DOI:10.1086/283303 URL [本文引用: 1]
The effect of photoperiod on walking, feeding, and oviposition in the western flower thrips
DOI:10.1111/eea.2004.111.issue-3 URL [本文引用: 1]
Response of a gall-forming guild (Hymenoptera: Cynipidae) to stressed and vigorous prairie roses
DOI:10.1603/0046-225X-33.4.1052 URL [本文引用: 1]
N addition suppresses the performance of grassland caterpillars (Gynaephora alpherakjj) by decreasing ground temperature
Mechanisms underlying loss of plant biodiversity by atmospheric nitrogen deposition in grasslands
大气氮沉降影响草地植物物种多样性机制研究综述
The effects of large herbivore grazing on meadow steppe plant and insect diversity
DOI:10.1111/jpe.2012.49.issue-5 URL [本文引用: 1]
Contrasting effects of nitrogen fertiliser application on the performance of closely related grasshoppers through changes in plant nutrient concentrations
DOI:10.1111/een.v48.3 URL [本文引用: 1]
Large mammalian herbivores affect arthropod food webs via changes in vegetation characteristics and microclimate
DOI:10.1111/jec.v111.9 URL [本文引用: 1]
Characteristics, causes, and consequences of trait-mediated indirect interactions in ecosystems
生态系统基于性状调节的物种间接作用: 特征、成因及后果
Negative effects of vertebrate on invertebrate herbivores mediated by enhanced plant nitrogen content
DOI:10.1111/1365-2745.13100
[本文引用: 1]
Classic theory holds that the main interaction within the herbivore guild is competition, based on research focused on co-occurring, similarly sized species that reduce the quantity of shared plant resources. However, plant quality may also be crucial in mediating herbivore interspecific interactions. This is especially true when competition occurs between distantly related herbivore species, given that small terrestrial herbivores (e.g., insect herbivores) appear to be more sensitive to alterations of plant quality than plant quantity. In this study, we first tested in the field whether large vertebrate herbivores (cattle Bos taurus) exerted a negative effect on smaller insect herbivores (grasshopper Euchorthippus unicolor) through their overlapping foraging preferences for a dominant grass Leymus chinensis. We measured changes in grass quantity, grass quality, and microclimatic conditions in response to vertebrate grazing and conducted additional manipulative studies in the field and the laboratory to identify potential mechanisms underlying the interaction. Our results showed that grazing by large herbivores caused a significant decline in grasshopper population density and individual performance (survival, size, and weight of both female and male E. unicolor), despite a 38% increase in grass nitrogen (N) content in grazed plots. Experiments manipulating N levels of L. chinensis in the field and the laboratory confirmed that enriching plant N had a negative effect on grasshopper individual performance and population size. Therefore, enhanced quality (N content) of plant resources appears to be an important driver in mediating the negative effect of vertebrate grazing on grasshoppers. Synthesis. We document that phylogenetic relatedness and trait similarity can be poor predictors of interaction strength in some cases, since distantly related herbivores of disparate size can interact indirectly via changes in plant quality. Counter-intuitively, the observed negative effect of cattle on grasshoppers was mediated, at least in part, by an increase in plant quality in cattle grazed areas. The implication is that light to moderate grazing, a common management strategy, may contribute to suppression of grasshoppers in the Eurasian steppe grassland system by altering plant nutrient supplies.

{kind=link}
{kind=link}
{kind=link}
{kind=link}