



城市化是一个由人口增长驱动的复杂社会-经济过程(United Nations, 2019), 在全球范围内发展迅速(Lin et al, 2020)。城市化改变土地利用类型(Lin et al, 2020), 导致物种栖息地质量下降, 栖息地破碎化以及丧失严重影响物种空间分布格局(Gippet et al, 2017; Wang et al, 2020), 同时降低栖息地间的景观连通性, 不利于个体扩散和基因流维持(Wang et al, 2008), 是目前野生动物生存的重要威胁之一(Grimm et al, 2008)。两栖动物有限的扩散能力(Smith & Green, 2005)和对栖息地功能、质量的高敏感性(Semlitsch, 2000; Hamer & McDonnell, 2008), 使其城市化区域的种群较易分化成为相对隔离的地方小种群(Wang, 2013)。保持栖息地景观连通性对于两栖动物集合种群的维持至关重要(Semlitsch, 2008)。此外, 两栖动物是受城市化威胁最严重的陆生脊椎动物类群(Seto et al, 2012), 是城市生态系统研究中重要的环境指示类群(Guzy et al, 2012)。因此, 研究两栖动物的潜在空间分布区域, 探究其栖息地景观连通性, 有助于我们深入理解城市化对野生动物种群的作用机制及造成的影响, 能够为当地生物多样性保护提供理论指导。
上海作为世界第三大城市(United Nations, 2019), 经历了近40年的高速城市化进程。城市化会严重影响本土物种的物种丰富度(Zhao et al, 2006)、种群数量(Zhang et al, 2016)和空间分布(Diao et al, 2022), 这使得生物多样性保护成为了上海生态城市建设的重要评价标准之一( 上海市规划和国土资源管理局 (2018) 上海市城市总体规划(2017-2035年).)。上海地区的本土两栖动物从20世纪80年代调查到的11种(黄正一等, 1980)锐减为如今的5种, 且现存两栖类物种的丰富度和多度与城市化水平呈显著负相关(Zhang et al, 2016)。然而, 这些两栖类物种的整体空间分布格局以及城市化对两栖类物种分布和扩散的影响机制却尚不明确。金线侧褶蛙(Pelophylax plancyi)是上海现存5个本土两栖类物种中数量最多且分布最广的②( 岳衢 (2019) 上海市公园无尾两栖类分布及其繁殖栖息地研究. 硕士学位论文, 华东师范大学, 上海.), 这使其成为反映城市中物种空间分布及景观连通性的理想物种之一。Wei等(2020)对金线侧褶蛙的种群遗传学研究发现, 金线侧褶蛙在上海地区表现出显著的遗传分化, 而这种分化的遗传格局与上海的现代城市化梯度高度一致。因此, 本研究以上海地区的金线侧褶蛙为例, 探究: (1)金线侧褶蛙在上海的潜在空间分布格局及其影响因素; (2)景观连通性对其地方小种群间个体扩散和基因交流的影响, 以期为城市生物多样性保护和生态空间规划建设提供科学指引和建议。
1 材料与方法
1.1 研究区域
上海市常住人口2,489万(上海市统计局, 2022)。上海的城市化格局是以黄浦江西岸的狭小区域为最初的城市中心(胡焕庸, 1987)不断向四周扩张形成的圈层结构(Kuang et al, 2014)以及不同功能区域( 上海市规划和国土资源管理局 (2018) 上海市城市总体规划(2017-2035年).)。其中, 外环高架以内的区域(约66,400 ha)人造地表面积占比80.95%, 人口密度约230人/ha (上海市统计局, 2022), 是上海城市化水平最高的区域。在外环高架路到绕城高速路之间的近郊区域内, 自20世纪90年代后期起, 城市化进程快速发展, 而在绕城高速路外侧的远郊区域内依然是以农业复合生态系统为主(图1)。Wei等(2020)根据上海的这种城市化圈层结构, 在不同区域内设置采样点, 研究了本地区城市化进程对两栖动物种群遗传格局的影响。结果显示, 这种圈层结构对上海地区金线侧褶蛙的种群遗传结构已经产生了显著的遗传隔离效应(Wei et al, 2020)。因此, 本研究利用Wei等(2020)设置的15个采样点(图1), 在以采样点为中心、半径2 km范围内的区域进行野外调查和采样工作。这些采样点中有9个为面积大于10 ha且保留原有水系的公园或城市绿地; 其余6个采样点位于由农业景观主导的郊野公园。
图1
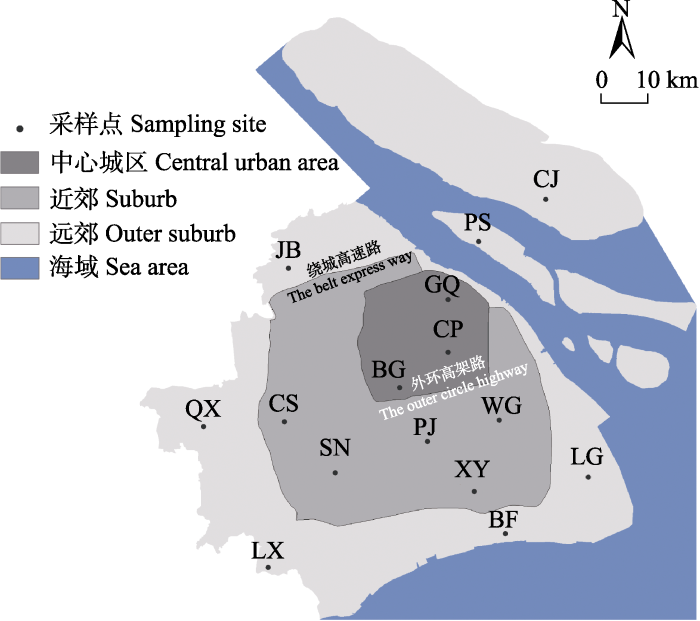
图1
上海地区金线侧褶蛙15个采样点分布图。CP: 世纪公园; BG: 上海植物园; GQ: 共青森林公园; CS: 辰山植物园; SN: 松南郊野公园; PJ: 浦江郊野公园; XY: 奉贤向阳村; WG: 浦东五灶港; JB: 嘉北郊野公园; QX: 青西郊野公园; LX: 廊下郊野公园; BF: 海湾郊野公园; LG: 南汇临港新城; PS: 长兴岛潘石镇; CJ: 崇明岛陈家镇。
Fig. 1
The map of 15 sampling sites of Pelophylax plancyi in Shanghai. CP, Century Park; BG, Shanghai Botanical Garden; GQ, Gongqing Forest Park; CS, Chenshan Botanical Garden; SN, Songnan Country Park; PJ, Pujiang Country Park; XY, Fengxian Xiangyang Village; WG, Pudong Wuzao Port; JB, Jiabei Country Park; QX, Qingxi Country Park; LX, Langxia Country Park; BF, Bay Country Park; LG, Nanhui Lingang New City; PS, Changxing Island Panshi Town; CJ, Chongming Island Chenjia Town.
1.2 金线侧褶蛙分布数据及组织标本采集
基于本研究的野外调查工作和实验室长期进行的上海地区两栖动物普查项目, 通过样线法对上海地区金线侧褶蛙进行种群调查。对于各城市公园和绿地, 由于实际面积普遍小于1 km2, 随机选择调查起始样点, 沿所在水体的岸线设定长200 m的样线, 为控制空间自相关性对采样的影响, 任意相邻样线的间距≥ 200 m。对于郊野公园, 由于面积较大, 因此设置1 km × 1 km的网格, 每个网格作为1个调查样方, 在每个样方中按照自南往北、自西往东的原则, 以发现的第一个水体(河流、池塘等)为起点, 设置1条200 m长的样线。2017年5-8月, 在本研究设定的15个采样点中的10个采样点共设置129条样线, 收集到金线侧褶蛙分布点GPS坐标数据共165条, 精度 ± 5 m (表1)。2017年5-8月和2018年5-8月, 在本研究设定的15个采样点上采集得到组织标本共计407份(表1)。调查于无雨的夜间(19:30至次日1:00) 进行, 调查人员沿样线以小于1 km/h的速度步行(Zhang et al, 2016), 使用强光手电筒搜寻, 捕捉金线侧褶蛙个体, 剪趾收集组织标本后放归。组织标本置于75%乙醇保存。本研究野外工作方案已通过华东师范大学实验动物伦理委员会审查(审查编号: AM20200201), 受其监督管理。
表1 上海市15个金线侧褶蛙采样点信息及两种分子标记使用到的样本信息
Table 1
| 采样点 Sampling site | 分布区域 Region | 城市化水平 Urbanization degree | 生境适宜 均值 Mean of habitat suitability | GPS坐标数 Number of GPS positions | 样本量(只) Sampling size | |
|---|---|---|---|---|---|---|
| 微卫星位点分析样本Samples for SSR analysis | 单核苷酸多肽位点分析样本 Samples for SNP analysis | |||||
| 世纪公园 Century Park (CP) | 中心城区 Central urban area | 0.734 | 0.236 | 15 | 32 | 10 |
| 上海植物园 Shanghai Botanical Garden (BG) | 中心城区 Central urban area | 0.773 | 0.147 | 10 | 28 | 10 |
| 共青森林公园 Gongqing Forest Park (GQ) | 中心城区 Central urban area | 0.705 | 0.135 | 7 | 16 | 10 |
| 辰山植物园 Chenshan Botanical Garden (CS) | 近郊 Suburb | 0.393 | 0.472 | - | 36 | 10 |
| 松南郊野公园 Songnan Country Park (SN) | 近郊 Suburb | 0.371 | 0.417 | 34 | 35 | 10 |
| 浦江郊野公园 Pujiang Country Park (PJ) | 近郊 Suburb | 0.469 | 0.526 | 12 | 15 | 10 |
| 奉贤向阳村 Fengxian Xiangyang Village (XY) | 近郊 Suburb | 0.412 | 0.372 | - | 25 | 10 |
| 浦东五灶港 Pudong Wuzao Port (WG) | 近郊 Suburb | 0.489 | 0.410 | - | 34 | 10 |
| 嘉北郊野公园 Jiabei Country Park (JB) | 远郊 Outer suburb | 0.530 | 0.455 | 14 | 17 | 10 |
| 青西郊野公园 Qingxi Country Park (QX) | 远郊 Outer suburb | 0.217 | 0.469 | 31 | 33 | 10 |
| 廊下郊野公园 Langxia Country Park (LX) | 远郊 Outer suburb | 0.313 | 0.463 | 25 | 39 | 10 |
| 海湾郊野公园 Bay Country Park (BF) | 远郊 Outer suburb | 0.262 | 0.519 | 3 | 11 | 10 |
| 南汇临港新城 Nanhui Lingang New City (LG) | 远郊 Outer suburb | 0.530 | 0.353 | - | 28 | 10 |
| 长兴岛潘石镇 Changxing Island Panshi Town (PS) | 远郊 Outer suburb | 0.085 | 0.280 | 14 | 30 | 10 |
| 崇明岛陈家镇 Chongming Island Chenjia Town (CJ) | 远郊 Outer suburb | 0.570 | 0.281 | - | 28 | 10 |
| 总计 Total | 165 | 407 | 150 | |||
SSR, simple sequence repeat; SNP, Single nucleotide polymorphism.
1.3 地理信息数据
本研究选用2017年8月24日Landsat 8卫星拍摄获取的Level 1T级别卫星影像, 取景号为LC811803 82017236LGN00和LC81180392017236LGN00。卫星影像数据的空间分辨率为30 m × 30 m、云量< 1%。卫星影像数据来源于中国科学院计算机网络信息中心地理空间数据云平台(
1.4 数据处理
1.4.1 金线侧褶蛙分布点数据
为控制位点空间自相关对预测结果的影响, 本研究利用ArcGIS 10.2 (ESRI)生成空间分辨率为30 m × 30 m的网格(fishnet)数据, 对处于同一网格的多个分布点, 只选取距离网格中心最近的1个分布点(Ward, 2007)。最终筛选保留了145个金线侧褶蛙的分布点数据。
1.4.2 遗传学数据
Wei等(2020)利用在上海地区15个采样点(图1)获取的全部407份样品分别提取13个微卫星(simple sequence repeat, SSR)位点数据(表1)。同时, 又从全部样品中随机抽取150份组织标本(每个采样点随机抽取10份标本), 经简化基因组测序获得71,910个单核苷酸多态性(single nucleotide polymorphism, SNP)位点(表1)。利用ARLEQUIN 3.5 (Excoffier et al, 2007)和vcftools 0.1.17 (Danecek et al, 2011)分别基于SSR和SNP数据集计算各采样点金线侧褶蛙地方小种群间的遗传分化系数(FST) (附录1), 并用FST/(1 - FST)对FST数据进行线性化处理(Rousset, 1997)。如此, 最终得到各小种群间分别用FST和FST/(1 - FST)计算的两组遗传距离矩阵。
1.4.3 环境变量
两栖动物的栖息地通常由水生环境和陆生环境两部分组成, 因此两栖动物对栖息地的完整性及质量均有较高要求。尤其是在城市化环境中, 地表类型、地表温度、植被盖度等因子对两栖动物的生存具有重要影响(Zhang et al, 2016; Li et al, 2018; Yang et al, 2022)。为了获得研究地区相关环境因子数据, 我们首先使用ENVI 5.2 (ESRI)对前述的两幅卫星影像分别进行辐射定标、大气校正、无缝镶嵌、研究区域裁剪等预处理工作。然后使用动态聚类算法(iterative self-organizing data analysis techniques algorithm, ISODATA; 邓书斌等, 2014)将研究区域的土地利用类型划分为7种: 内陆水域、淤泥质海滩、绿地、林地、耕地、人造地表、河口及海洋水域。使用大气校正法进行地表温度反演(邓书斌等, 2014)。此外, 使用归一化植被指数(normalized difference vegetation index, NDVI)反映具体区域的植被盖度, NDVI计算公式为:
其中, NIR指近红外波段的反射值, Red指红色波段的反射值(邓书斌等, 2014)。
1.5 潜在空间分布模型
由于野外调查无法穷尽上海地区全部区域, 因此使用最大熵模型(MaxEnt 3.4.1; Phillips et al, 2006), 基于卫星影像数据和金线侧褶蛙种群调查数据, 预测整个上海地区金线侧褶蛙的潜在空间分布格局。模型使用土地利用类型(7种地表类型)、地表温度、NDVI作为环境预测变量, 随机选取75%的种群调查获取的金线侧褶蛙分布点作为训练集用于模型构建, 剩余25%的分布点作为验证集用于模型验证。采用放回式取样法重复运算10次, 其余参数采用系统默认设置, 以运算10次所得模型平均值作为上海地区金线侧褶蛙的潜在空间分布格局。选择刀切法(Jackknife)分析每个变量对金线侧褶蛙潜在空间分布格局的重要性, 使用受试者操作特征曲线(receiver operating characteristic curve, ROC)下的面积值(area under the curve, AUC)进行模型总有效率的评价: 0.6-0.7为较差; 0.7-0.8为一般; 0.8-0.9为良好; 0.9-1.0为优秀(Swets, 1988)。
1.6 地理距离和阻力距离
已有的研究统计发现, 两栖动物的最大扩散距离多在2 km以内(Smith & Green, 2005), 前期的研究也发现上海地区2 km半径范围内的景观结构对两栖动物群落的丰富度和多度等具有显著的影响(Zhang et al, 2016)。因此, 本研究将以每个采样点为中心、半径2 km范围内的所有金线侧褶蛙划归在1个地方小种群中。通过ArcGIS计算得到两两采样点之间的欧式距离(Euclidean distance); 将MaxEnt模型模拟所得上海地区金线侧褶蛙生境适宜值(0-1)取倒数处理(Wang et al, 2013), 得到生境扩散阻力值。使用Circuitscape 4.0 (McRae & Beier, 2007), 选择成对模式(pairwise mode), 其余参数采用默认值, 计算各地方小种群间的阻力距离矩阵并生成扩散路径电流图。使用扩散路径电流图指示金线侧褶蛙在上海范围内的潜在扩散廊道, 电流值越高, 表示其扩散的可能性越大, 该区域成为其扩散廊道的可能性也越大。
1.7 生境适宜值与城市化水平之间的相关性
为进一步分析上海地区金线侧褶蛙生境适宜值与城市化水平之间的关系, 我们按照Zhang等(2016)的方法, 使用ArcGIS分别以15个采样点(图1)的中心为圆心, 为每个地方小种群建立2 km半径的区域, 基于土地利用类型图层, 计算该区域内人造地表的占比, 用来代表该小种群所在栖息地的城市化水平(Lin et al, 2020), 其值越高, 代表该区域城市化水平越高。同时使用ArcGIS提取该区域内金线侧褶蛙的生境适宜值并计算其平均值, 以生境适宜均值的大小衡量相应小种群所在栖息地的整体适宜性水平, 区域内该均值越大, 其栖息地适宜性水平越高。在R 4.1.3 (
1.8 地方小种群间的隔离模型
2 结果
2.1 上海地区金线侧褶蛙潜在空间分布格局
图2
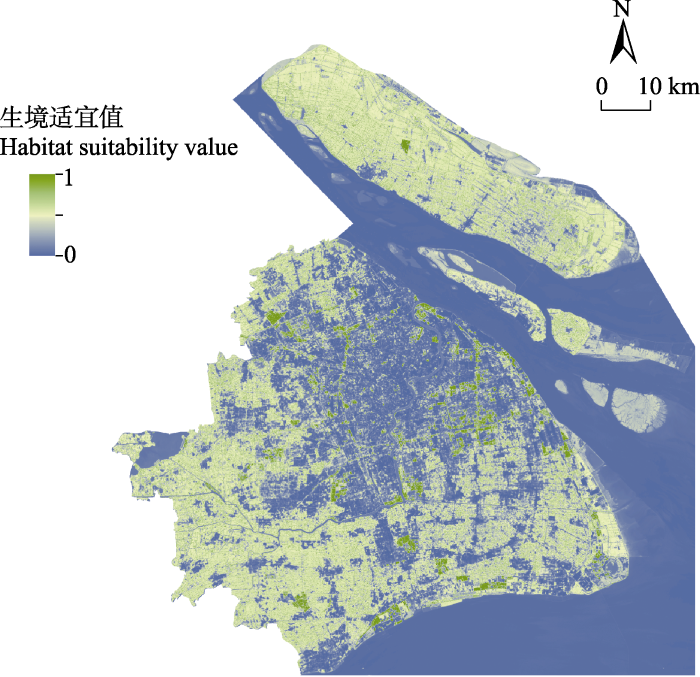
图2
上海金线侧褶蛙潜在空间分布
Fig. 2
The potential spatial distribution of Pelophylax plancyi in Shanghai
在使用的3种环境变量(附录2)中, NDVI对模型的百分比贡献率最大(51.833% ± 9.136%)。刀切法检验结果表明, 单独使用NDVI变量时, 训练集增益(training gain)最高(0.924 ± 0.080), 土地利用类型(0.904 ± 0.058)次之, 地表温度(0.887 ± 0.023)最低(图3a)。金线侧褶蛙的出现概率随着NDVI值的增加而上升(图3b); 在土地利用类型的选择上, 林地和绿地的预测概率值最高, 内陆水体和耕地次之, 人造地表、淤泥质海滩和河口及海洋水域的预测概率低于0.5 (图3c); 随着地表温度的升高, 其预测概率先增加后减小, 当地表温度在35℃左右时出现概率达到峰值(图3d)。
图3
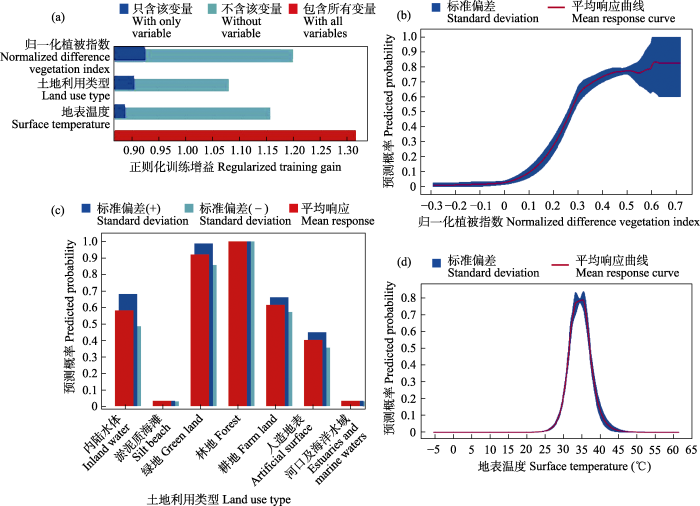
图3
环境因子对金线侧褶蛙潜在分布的相对贡献率(a)及响应图(b-d)
Fig. 3
Schematic diagram of the contribution rate of environmental factors to the potential distribution of Pelophylax plancyi (a) and the response of environmental factors to the potential distribution (b-d)
2.2 栖息地景观连通性
图4
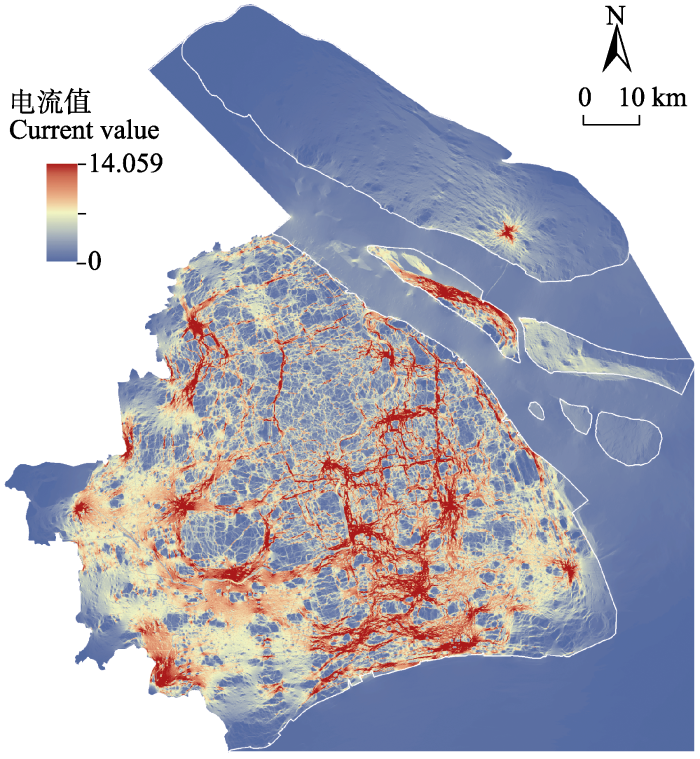
图4
上海地区金线侧褶蛙扩散路径电流图
Fig. 4
The disperse path map of Pelophylax plancyi in Shanghai based on current flows
表2 15个金线侧褶蛙地方小种群间的欧式距离系数(左下)及阻力距离系数(右上)
Table 2
| CP | BG | GQ | CS | SN | PJ | XY | WG | JB | QX | LX | BF | LG | PS | CJ | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CP | - | 8.921 | 10.583 | 7.182 | 6.970 | 7.395 | 7.299 | 5.691 | 8.779 | 8.835 | 7.912 | 6.766 | 6.943 | 12.184 | 19.107 |
| BG | 13.092 | - | 13.819 | 7.759 | 7.633 | 8.225 | 8.681 | 7.829 | 9.858 | 9.460 | 8.627 | 8.260 | 8.816 | 14.709 | 21.486 |
| GQ | 11.502 | 21.893 | - | 11.321 | 11.353 | 12.329 | 12.160 | 10.787 | 11.826 | 13.005 | 12.284 | 11.578 | 11.779 | 14.775 | 21.914 |
| CS | 38.632 | 26.020 | 44.400 | - | 3.223 | 6.173 | 6.094 | 5.651 | 5.873 | 4.033 | 4.085 | 5.543 | 6.352 | 12.049 | 18.753 |
| SN | 38.143 | 25.329 | 47.017 | 14.861 | - | 5.695 | 5.540 | 5.228 | 6.495 | 4.721 | 3.481 | 4.975 | 5.877 | 12.014 | 18.738 |
| PJ | 20.314 | 13.272 | 31.584 | 30.861 | 23.018 | - | 5.935 | 5.435 | 8.586 | 7.787 | 6.666 | 5.698 | 6.356 | 12.952 | 19.726 |
| XY | 30.711 | 27.821 | 42.069 | 43.910 | 32.645 | 15.007 | - | 4.826 | 8.526 | 7.674 | 6.447 | 4.485 | 5.339 | 12.598 | 19.361 |
| WG | 18.445 | 22.859 | 28.488 | 46.662 | 39.897 | 16.884 | 16.375 | - | 7.768 | 7.261 | 6.154 | 4.269 | 4.300 | 11.210 | 18.016 |
| JB | 38.090 | 34.177 | 34.380 | 32.553 | 45.087 | 46.981 | 61.885 | 55.283 | - | 7.615 | 7.402 | 7.972 | 8.546 | 12.503 | 19.053 |
| QX | 55.905 | 43.777 | 60.139 | 18.119 | 29.195 | 48.796 | 61.102 | 64.778 | 38.533 | - | 5.428 | 7.115 | 7.946 | 13.715 | 20.421 |
| LX | 60.891 | 48.251 | 69.988 | 31.233 | 22.971 | 43.667 | 48.216 | 59.805 | 63.727 | 33.378 | - | 5.858 | 6.782 | 12.937 | 19.659 |
| BF | 41.736 | 40.070 | 52.729 | 55.036 | 42.461 | 27.280 | 12.290 | 24.799 | 74.170 | 71.550 | 54.098 | - | 4.068 | 11.904 | 18.654 |
| LG | 40.987 | 46.146 | 49.087 | 68.159 | 58.566 | 37.302 | 26.395 | 23.480 | 78.639 | 86.020 | 74.354 | 22.382 | - | 11.801 | 18.530 |
| PS | 24.212 | 36.120 | 14.477 | 58.697 | 61.387 | 44.372 | 52.475 | 36.883 | 43.953 | 73.881 | 84.346 | 61.639 | 52.830 | - | 13.680 |
| CJ | 39.235 | 51.594 | 30.122 | 74.281 | 76.911 | 58.910 | 65.210 | 48.951 | 56.961 | 89.118 | 99.844 | 72.958 | 60.178 | 15.649 | - |
地方小种群缩写详见
The abbreviation of local populations corresponds to
2.3 地方小种群间隔离模式
基于SSR和SNP分子数据的FST和FST/(1-FST)分别作为遗传距离指标的Mantel检验结果(表3)表明: 欧式距离矩阵和遗传距离矩阵之间并未表现出显著的相关性; 而阻力距离矩阵和遗传距离矩阵之间均存在显著的正相关, 遗传分化程度随着阻力距离的增大而增大。
表3 距离隔离模型和阻力隔离模型的Mantel检验验证
Table 3
| 空间距离 Spatial distance | FST | FST/(1-FST) | ||
|---|---|---|---|---|
| SSR | SNP | SSR | SNP | |
| 欧氏距离 Euclidean distance | 0.030 | 0.005 | 0.031 | 0.005 |
| 阻力距离 Resistance distance | 0.593** | 0.420* | 0.598** | 0.419* |
SSR: 微卫星位点; SNP: 单核苷酸多态性位点; FST: 遗传分化系数。表内数值均为r值。* P < 0.05; ** P < 0.01。
SSR, Simple sequence repeat; SNP, Single nucleotide polymorphism; FST, Coefficient of inheritance differentiation. The table shows r values of Mantel test. * P < 0.05; ** P < 0.01.
3 讨论
两栖动物作为受威胁程度最高的陆生脊椎动物, 其41%的物种都处于受胁状态(threatened; IUCN, 2021), 而城市化是威胁其生存的最主要因素之一(Hamer & McDonnell, 2008)。相较于鸟类和哺乳类(Magle et al, 2012), 对城市化背景下两栖动物的研究仍然匮乏(Hamer & McDonnell, 2008; Magle et al, 2012)。与此同时, 全球因城市化而受威胁的两栖类物种中, 80%分布在热带和亚热带的发展中国家和地区(Hamer & McDonnell, 2008), 而两栖动物的城市生态学研究多集中于地处温带的发达国家(Botzat et al, 2016; Johnson & Munshi-South, 2017)。另一方面, 发展中国家和地区是当下和未来全球城市化高速发展的热点区域, 因此热带和亚热带地区两栖动物的城市生态学研究对全球两栖类生物多样性保护的影响不容忽视(Hamer & McDonnell, 2008)。上海地处亚热带地区, 作为正在经历高速城市化发展的代表性城市, 为我们研究城市化背景下的两栖动物提供了理想的研究区域。
上海以环状的形式向外进行城市化扩张, 拥有完整的“中心城区—近郊—远郊”城市化梯度(Kuang et al, 2014)。上海地区金线侧褶蛙的生境适宜性随着城市化水平的上升而显著下降, 证明高度城市化发展会给两栖动物带来负面影响(Lin et al, 2020)。人造地表占比高、植被覆盖低的区域, 城市热岛效应可使其地表温度高达43℃ (图2, 图3), 不利于金线侧褶蛙的生存。城市化通过改变自然栖息地的土地利用类型以及相关区域的自然温度变化规律等因素对两栖动物的分布产生影响(Yang et al, 2022)。水体对于两栖动物的生存和繁殖至关重要, 但是并非所有类型的水体均可以作为这类物种的适宜栖息地。例如, 穿越上海城区的主干河流黄浦江、上海西部的淀山湖、东部毗邻的东海等大型水体均不适宜金线侧褶蛙的生存(图2)。尽管如此, 城市发展过程中仍保有部分适宜两栖动物生存的栖息地空间。本研究发现, 植被覆盖度高、温度在35℃左右的小型水域更适宜金线侧褶蛙的生存(图3d)。在中国大部分城市群, 林地和草地的占比会对两栖动物的丰富度分布产生积极影响(Lin et al, 2020)。上海的林绿地空间结构较为复杂, 在林下、绿地常存在小型水体( 上海市绿化和市容管理局, 上海市园林设计研究总院有限公司 (2018) 绿地设计规范(修订).), 而这一结构恰好可满足金线侧褶蛙的生存需要。小型水体能够为两栖动物营造适宜其生存的微生境( 岳衢 (2019) 上海市公园无尾两栖类分布及其繁殖栖息地研究. 硕士学位论文, 华东师范大学, 上海.)。例如, 维持小型水体周边的植被覆盖度有利于金线侧褶蛙种群的延续(Zhang et al, 2016)。此外, 由于金线侧褶蛙对于栖息地水体具有高度依赖性, 其自身有限的扩散能力同样可能导致人造地表对其栖息地适宜性的影响较小。这提示我们, 在城市化水平较高的区域, 城市公园、绿化带等城市公共绿地仍然有可能成为适宜两栖动物生存的优良栖息地。
城市化不断扩张导致的栖息地面积减少和质量下降可能会使对栖息地依赖度高而扩散能力弱的两栖物种面临种群数量下降、遗传多样性丧失的风险(Tsuji et al, 2011), 甚至会影响种群遗传结构(Hamer & McDonnell, 2008)。栖息地连通性的保持对物种的长期生存至关重要(McRae & Beier, 2007)。通过保护栖息地之间的连通性, 可以减缓因栖息地质量变化和破碎化造成的物种数量减少(Tsuji et al, 2011), 也能促进遗传变异, 提升物种遗传多样性(Munshi-South et al, 2016)。金线侧褶蛙的扩散力较弱, 加上城市化造成的隔离效应(Wei et al, 2020)导致金线侧褶蛙种群易出现种群下水平的遗传分化, 形成具有不同遗传结构的空间上相对隔离的地方小种群(附录1)。传统IBD模型分析遗传分化与地理距离之间的相关性(Wright, 1943)。该模型是基于栖息地同质化的假设前提建立的(McRae, 2006)。然而个体从一个种群扩散到另一个种群时, 经常需要通过不同类型的景观, 不同的景观构成会对其扩散产生不同的阻力, 也会影响个体扩散路径的选择(McRae, 2006)。因此, 由IBD模型发展而来的IBR模型在前者的基础上进一步考虑了栖息地异质性对物种扩散产生的影响(McRae, 2006)。上海地区的城市景观异质性使得不同景观组成对金线侧褶蛙的个体扩散和基因交流产生差异化的阻碍效果, 符合IBR模型的使用场景(表3)。廊道被认为有利于增强栖息地间的连通性, 从而增强基因流, 改善地方小种群的遗传多样性, 进而提升城市化区域整体的生物多样性水平(Tewksbury et al, 2002)。上海的城市化对景观结构的改造所造成的异质性景观对金线侧褶蛙的个体扩散产生了显著的负面影响。相比于地理距离, 扩散阻力能够更好地反映异质性景观对金线侧褶蛙的个体扩散造成的影响, 地方种群间的连通性建设需要将景观构成列入评价范围。
了解野生动物的空间分布格局及景观连通性对个体扩散和基因交流的影响, 能够为针对性保护措施的制定提出科学建议。例如, 上海外环高架路(图1)沿线的外环绿带是规划中(2018-2035年)的重要市区野生动物栖息地和廊道( 上海市绿化和市容管理局, 上海市规划和自然资源局 (2020) 上海市生态空间专项规划(2018-2035) (草案公示稿).)。而本研究证实围绕外环高架路的栖息地对金线侧褶蛙具有较高生境适宜值(图2)和较高的扩散电流值(图4), 说明在该栖息地中的两栖动物不仅生存可能性高, 扩散可能性也较高。这意味着将外环绿带作为未来城市生态空间的规划方案是合理的, 科学地保护和规划外环绿带有利于野生动物的栖息与个体的扩散交流。此外, 上海陆域南部和西南部区域金线侧褶蛙的潜在空间分布(图2)和扩散(图4)可能性较高, 在未来上海生态城市建设过程中, 对这两部分区域应当以维持现有栖息地为主, 提供适宜两栖动物生存的具有较高植被覆盖度的小型水域, 比如池塘、湿地或溪流(Hamer & McDonnell, 2008), 辅以廊道建设以促进两栖动物种群的保护和扩散。与南部和西南部情况不同, 上海陆域北部和东北部区域包括部分中心城区, 受人类活动影响大, 城市化程度高, 适宜金线侧褶蛙生存的栖息地少(图2), 其自然扩散可能性也更低(图4)。考虑到上海总体区域的生物多样性, 未来对于该区域应以栖息地修复为先, 推进城市公园、楔形绿地等建设, 在此基础上再进行廊道构建, 促进个体的扩散和交流。总之, 本研究以两栖动物为例, 探讨了上海地区潜在野生动物栖息地的分布格局和野生动物扩散廊道的分布。本研究为上海市加强和提升生态文明建设提供了理论参考, 为上海市未来生态空间的规划建设提供了科学依据和研究案例。
附录 Supplementary Material
附录1 15个地方小种群间的遗传分化系数
Appendix 1 The pairwise fixation index (FST) between 15 local populations
附录2 用于上海地区金线侧褶蛙种群的潜在空间分布模型构建的环境因子
Appendix 2 Environmental factors used in the potential distribution model of Pelophylax plancyi in Shanghai, China
参考文献
Unexploited opportunities in understanding liveable and biodiverse cities. A review on urban biodiversity perception and valuation
DOI:10.1016/j.gloenvcha.2016.04.008 URL [本文引用: 1]
1000 Genomes Project Analysis Group (2011) The variant call format and VCFtools
DOI:10.1093/bioinformatics/btr330
URL
[本文引用: 1]

Summary: The variant call format (VCF) is a generic format for storing DNA polymorphism data such as SNPs, insertions, deletions and structural variants, together with rich annotations. VCF is usually stored in a compressed manner and can be indexed for fast data retrieval of variants from a range of positions on the reference genome. The format was developed for the 1000 Genomes Project, and has also been adopted by other projects such as UK10K, dbSNP and the NHLBI Exome Project. VCFtools is a software suite that implements various utilities for processing VCF files, including validation, merging, comparing and also provides a general Perl API.
Predicting current and future species distribution of the raccoon dog (Nyctereutes procyonoides) in Shanghai, China
DOI:10.1016/j.landurbplan.2022.104581 URL [本文引用: 1]
The ade 4 package: Implementing the duality diagram for ecologists
Arlequin (version 3.0): An integrated software package for population genetics data analysis
I’m not like everybody else: Urbanization factors shaping spatial distribution of native and invasive ants are species-specific
DOI:10.1007/s11252-016-0576-7 URL [本文引用: 1]
Global change and the ecology of cities
DOI:10.1126/science.1150195
PMID:18258902
[本文引用: 1]
Urban areas are hot spots that drive environmental change at multiple scales. Material demands of production and human consumption alter land use and cover, biodiversity, and hydrosystems locally to regionally, and urban waste discharge affects local to global biogeochemical cycles and climate. For urbanites, however, global environmental changes are swamped by dramatic changes in the local environment. Urban ecology integrates natural and social sciences to study these radically altered local environments and their regional and global effects. Cities themselves present both the problems and solutions to sustainability challenges of an increasingly urbanized world.
Urbanization interferes with the use of amphibians as indicators of ecological integrity of wetlands
DOI:10.1111/j.1365-2664.2012.02172.x
PMID:32367892
[本文引用: 1]
Wetlands are ecologically and economically important ecosystems but are threatened globally by many forms of human disturbance. Understanding the responses of wetland species to human disturbance is essential for effective wetland management and conservation. We undertook a study to determine (i) whether anurans can be used effectively to assess the ecological integrity of wetlands affected by groundwater withdrawal and, if so, (ii) what effect increasing urbanization might have on the utility of anurans as wetland indicators. We monitored the intensity of anuran calls at 42 wetlands in south-western Florida throughout 2001-2002 and 2005-2009. We first validated the use of anurans to assess wetland integrity using a small group of wetlands by comparing anuran calling and subsequent tadpole development with an established index employing vegetation composition and structure. We then verified that the results could be expanded to a variety of sites throughout the region. Finally, we focused on urbanized wetlands to determine whether urbanization could interfere with the use of anurans to assess wetland integrity. We used PRESENCE to estimate occupancy and detection probabilities and to examine the relationship between occupancy and five covariates expected to influence individual species occurrence. We used FRAGSTATS to calculate the mean proximity index for urbanized wetlands, which assesses the size and distribution of land use types within a specified area. Our results showed that the group of species including oak toad, southern cricket frog, pinewoods treefrog, barking treefrog, and little grass frog is a reliable indicator of wetland integrity. However, this same group of species, which is sensitive to wetland health, is selectively excluded from urbanized wetlands.. Although anurans are effective indicators of wetland health and complement vegetation surveys, the usefulness of this group for monitoring the ecological integrity of wetlands can be substantially reduced, or eliminated, as a consequence of urbanization. We urge for careful consideration of confounding factors in any studies examining the utility of indicator species.© 2012 The Authors. Journal of Applied Ecology © 2012 British Ecological Society.
Amphibian ecology and conservation in the urbanising world: A review
DOI:10.1016/j.biocon.2008.07.020 URL [本文引用: 7]
Amphibian and reptile species in Shanghai
上海地区的两栖爬行动物
Evolution of life in urban environments
A comparative analysis of megacity expansions in China and the U.S.: Patterns, rates and driving forces
DOI:10.1016/j.landurbplan.2014.08.015 URL [本文引用: 2]
Influence of breeding habitat characteristics and landscape heterogeneity on anuran species richness and abundance in urban parks of Shanghai, China
Urbanization effects on Chinese mammal and amphibian richness: A multi-scale study using the urban-rural gradient approach
DOI:10.1088/2515-7620/abd1c5
[本文引用: 5]
The scale and extent of global urbanization are unprecedented and increasing. As urbanization generally encroaches on natural habitats and the urban ecological footprint reaches far beyond the city limits, how urbanization affects biodiversity has received increasing attention from the scientific community. Nonetheless, the comprehensive syntheses of urbanization consequences for biodiversity, including diverse taxonomic groups, across multiple spatial scales and spanning a wide gradient range of urbanization intensity are still insufficient. Here, based on the urban-rural gradient approach, we assessed the effects of urbanization on Chinese mammal and amphibian richness across the entire urbanization gradient (i.e., urbanization level from 0 to 1) at the national, regional and urban agglomeration scales. We used the global mammal and amphibian distribution data along with corresponding background climate, habitat conditions and socioeconomic activities data for analysis. Our results revealed a detailed and diverse pattern of Chinese mammal and amphibian richness along the entire spectrum of urbanization gradient across three spatial scales. And an approximately monotonic decrease only existed in certain urban agglomerations. The imprint of urbanization on mammal and amphibian richness were largely masked by the overall primacy of background climate at the national and regional scales. As the scale of analysis shifting from the country to urban agglomerations, urbanization-associated variables and locally specific limiting factors started to play important roles in driving the richness patterns. Moreover, the environmental Kuznets curve hypothesis can explain the relationship between biodiversity pressure and urbanization activities in certain Chinese urban agglomerations. However, the findings of urbanization effects on biodiversity using the urban-rural gradient analysis should be interpreted with caution because many possible driving forces simultaneously present along the urban-rural gradient and are very challenging to attribute.
Urban wildlife research: Past, present, and future
DOI:10.1016/j.biocon.2012.06.018 URL [本文引用: 2]
The detection of disease clustering and a generalized regression approach
Isolation by resistance
Despite growing interest in the effects of landscape heterogeneity on genetic structuring, few tools are available to incorporate data on landscape composition into population genetic studies. Analyses of isolation by distance have typically either assumed spatial homogeneity for convenience or applied theoretically unjustified distance metrics to compensate for heterogeneity. Here I propose the isolation-by-resistance (IBR) model as an alternative for predicting equilibrium genetic structuring in complex landscapes. The model predicts a positive relationship between genetic differentiation and the resistance distance, a distance metric that exploits precise relationships between random walk times and effective resistances in electronic networks. As a predictor of genetic differentiation, the resistance distance is both more theoretically justified and more robust to spatial heterogeneity than Euclidean or least cost path-based distance measures. Moreover, the metric can be applied with a wide range of data inputs, including coarse-scale range maps, simple maps of habitat and nonhabitat within a species' range, or complex spatial datasets with habitats and barriers of differing qualities. The IBR model thus provides a flexible and efficient tool to account for habitat heterogeneity in studies of isolation by distance, improve understanding of how landscape characteristics affect genetic structuring, and predict genetic and evolutionary consequences of landscape change.
Circuit theory predicts gene flow in plant and animal populations
Population genomics of the Anthropocene: Urbanization is negatively associated with genome-wide variation in white-footed mouse populations
DOI:10.1111/eva.12357
PMID:27099621
[本文引用: 1]
Urbanization results in pervasive habitat fragmentation and reduces standing genetic variation through bottlenecks and drift. Loss of genomewide variation may ultimately reduce the evolutionary potential of animal populations experiencing rapidly changing conditions. In this study, we examined genomewide variation among 23 white-footed mouse (Peromyscus leucopus) populations sampled along an urbanization gradient in the New York City metropolitan area. Genomewide variation was estimated as a proxy for evolutionary potential using more than 10 000 single nucleotide polymorphism (SNP) markers generated by ddRAD-Seq. We found that genomewide variation is inversely related to urbanization as measured by percent impervious surface cover, and to a lesser extent, human population density. We also report that urbanization results in enhanced genomewide differentiation between populations in cities. There was no pattern of isolation by distance among these populations, but an isolation by resistance model based on impervious surface significantly explained patterns of genetic differentiation. Isolation by environment modeling also indicated that urban populations deviate much more strongly from global allele frequencies than suburban or rural populations. This study is the first to examine loss of genomewide SNP variation along an urban-to-rural gradient and quantify urbanization as a driver of population genomic patterns.
Maximum entropy modeling of species geographic distributions
DOI:10.1016/j.ecolmodel.2005.03.026 URL [本文引用: 1]
Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance
DOI:10.1093/genetics/145.4.1219
PMID:9093870
[本文引用: 1]
I reexamine the use of isolation by distance models as a basis for the estimation of demographic parameters from measures of population subdivision. To that aim, I first provide results for values of F-statistics in one-dimensional models and coalescence times in two-dimensional models, and make more precise earlier results for F-statistics in two-dimensional models and coalescence times in one-dimensional models. Based on these results, I propose a method of data analysis involving the regression of FST/(1-FST) estimates for pairs of subpopulations on geographic distance for populations along linear habitats or logarithm of distance for populations in two-dimensional habitats. This regression provides in principle an estimate of the product of population density and second moment of parental axial distance. In two cases where comparison to direct estimates is possible, the method proposed here is more satisfactory than previous indirect methods.
Principles for management of aquatic- breeding amphibians
Differentiating migration and dispersal processes for pond-breeding amphibians
Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools
Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations?
DOI:10.1111/eco.2005.28.issue-1 URL [本文引用: 2]
Measuring the accuracy of diagnostic systems
DOI:10.1126/science.3287615
PMID:3287615
[本文引用: 1]
Diagnostic systems of several kinds are used to distinguish between two classes of events, essentially "signals" and "noise". For them, analysis in terms of the "relative operating characteristic" of signal detection theory provides a precise and valid measure of diagnostic accuracy. It is the only measure available that is uninfluenced by decision biases and prior probabilities, and it places the performances of diverse systems on a common, easily interpreted scale. Representative values of this measure are reported here for systems in medical imaging, materials testing, weather forecasting, information retrieval, polygraph lie detection, and aptitude testing. Though the measure itself is sound, the values obtained from tests of diagnostic systems often require qualification because the test data on which they are based are of unsure quality. A common set of problems in testing is faced in all fields. How well these problems are handled, or can be handled in a given field, determines the degree of confidence that can be placed in a measured value of accuracy. Some fields fare much better than others.
Corridors affect plants, animals, and their interactions in fragmented landscapes
Paddy-associated frog declines via urbanization: A test of the dispersal-dependent-decline hypothesis
DOI:10.1016/j.landurbplan.2011.08.005 URL [本文引用: 2]
Examining the full effects of landscape heterogeneity on spatial genetic variation: A multiple matrix regression approach for quantifying geographic and ecological isolation
DOI:10.1111/evo.12134
PMID:24299396
[本文引用: 2]
Understanding the effects of landscape heterogeneity on spatial genetic variation is a primary goal of landscape genetics. Ecological and geographic variables can contribute to genetic structure through geographic isolation, in which geographic barriers and distances restrict gene flow, and ecological isolation, in which gene flow among populations inhabiting different environments is limited by selection against dispersers moving between them. Although methods have been developed to study geographic isolation in detail, ecological isolation has received much less attention, partly because disentangling the effects of these mechanisms is inherently difficult. Here, I describe a novel approach for quantifying the effects of geographic and ecological isolation using multiple matrix regression with randomization. I explored the parameter space over which this method is effective using a series of individual-based simulations and found that it accurately describes the effects of geographic and ecological isolation over a wide range of conditions. I also applied this method to a set of real-world datasets to show that ecological isolation is an often overlooked but important contributor to patterns of spatial genetic variation and to demonstrate how this analysis can provide new insights into how landscapes contribute to the evolution of genetic variation in nature. © 2013 The Author(s). Evolution © 2013 The Society for the Study of Evolution.
Quantifying the roles of ecology and geography in spatial genetic divergence
DOI:10.1111/ele.12025
PMID:23137142
Investigating the properties of ecological landscapes that influence gene flow among populations can provide key insights into the earliest stages of biological divergence. Both ecological and geographical factors can reduce gene flow, which can lead to population divergence, but we know little of the relative strengths of these phenomena in nature. Here, we use a novel application of structural equation modelling to quantify the contributions of ecological and geographical isolation to spatial genetic divergence in 17 species of Anolis lizards. Our comparative analysis shows that although both processes contributed significantly, geographical isolation explained substantially more genetic divergence than ecological isolation (36.3 vs. 17.9% of variance respectively), suggesting that despite the proposed ubiquity of ecological divergence, non-ecological factors play the dominant role in the evolution of spatial genetic divergence.© 2012 Blackwell Publishing Ltd/CNRS.
Plant diversity along the urban-rural gradient and its relationship with urbanization degree in Shanghai, China
DOI:10.3390/f11020171
URL
[本文引用: 1]
Urbanization is one of the major causes for plant diversity loss at the local and regional scale. However, how plant species distribute along the urban–rural gradient and what the relationship between urbanization degree and plant diversity is, is not very clear. In this paper, 134 sample sites along two 18 km width transects that run across the urban center of Shanghai were investigated. We quantified the spatial patterns of plant diversity along the urban–rural gradient and measured the relationship between plant diversity and urbanization degree, which was calculated using a land use land cover map derived from high spatial resolution aerial photos. We recorded 526 vascular plant species in 134 plots, 57.8% of which are exotic plant species. Six spatial distribution patterns of species richness were identified for different plant taxa along the rural to urban gradient. The native plant species richness showed no significant relationship to urbanization degree. The richness of the all plants, woody plants and perennial herbs presented significant positive relationship with urbanization degree, while the richness of annual herbs, Shannon-Wiener diversity and Heip evenness all exhibited a negative relationship to urbanization degree. Urbanization could significantly influence plant diversity in Shanghai. Our findings can provide insights to understand the mechanism of urbanization effects on plant diversity, as well as plant diversity conservation in urban areas.
Habitat suitability modelling to correlate gene flow with landscape connectivity
Modelling the potential geographic distribution of invasive ant species in New Zealand
DOI:10.1007/s10530-006-9072-y URL [本文引用: 1]
Long-term urbanization impacts the eastern golden frog (Pelophylax plancyi) in Shanghai City: Demographic history, genetic structure, and implications for amphibian conservation in intensively urbanizing environments
DOI:10.1111/eva.v14.1 URL [本文引用: 6]
Isolation by distance
DOI:10.1093/genetics/28.2.114 PMID:17247074 [本文引用: 2]
A global analysis of urbanization effects on amphibian richness: Patterns and drivers
DOI:10.1016/j.gloenvcha.2022.102476 URL [本文引用: 2]
Responses of anuran communities to rapid urban growth in Shanghai, China
Ecological consequences of rapid urban expansion: Shanghai, China
DOI:10.1890/1540-9295(2006)004[0341:ECORUE]2.0.CO;2 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}