


生物多样性是指生物及其与周边环境形成的生态复合体以及与此相关的各种生态过程的总和(Mori et al, 2017; 文志等, 2020), 生态系统服务通过生物地球化学循环实现, 是人类从生态系统中获得的各种惠益(Carpenter et al, 2009), 二者在生态系统中均表现出受限于自然基底, 并被气候变化和人类活动所影响的特征(Paul et al, 2020)。高水平的生物多样性和生态系统服务不仅可为当前社会发展提供物质保障, 还有利于物质保障的长期稳定供给, 是人类社会可持续发展的重要基石。但千年生态系统评估(MA)、生物多样性和生态系统服务政府间科学政策平台(IPBES)的全球评估报告(Carpenter et al, 2009; Bongaarts, 2019)等权威研究共同证实了世界各国普遍面临着生物多样性下降和生态系统退化的境况, 保护生物多样性和增强生态系统服务成为人类社会实现可持续发展的必然选择。
生物多样性与生态系统服务密切相关, 二者不仅影响因素重叠, 且它们本身也具有交互作用(Mace et al, 2012), 厘清二者间的复杂关系和反馈机制是实现生物多样性与生态系统服务综合提升的基础。生物多样性是物种相互作用和各种生态过程共同作用的结果, 受气候变化和人类活动干扰(井新等, 2022), 并在生态过程中产生生态系统服务, 如植被生长带来的固碳释氧和土壤保持(Huang et al, 2018; Wen et al, 2019)。来自生物多样性-生态系统功能(BEF)的研究表明, 生态功能随着生物多样性水平的提高而增加(Rey Benayas et al, 2009; Paul et al, 2020)。但并非所有生态系统服务都完全依赖于生态功能, 生态系统服务由生物多样性、生态功能和非生物驱动因素单独或共同决定(Midgley, 2012; 井新和贺金生, 2021), 因此生态功能的增加未必能带来生态系统服务的提升(文志等, 2020)。如Lange等(2015)、Wen等(2019)的研究发现土壤微生物多样性和植物多样性增加有利于土壤保持和固碳服务的提高, 但Adler等(2011)、Chen等(2022)、Wallis等(2023)的研究发现植物多样性与净初级生产力间并没有明确的关系。同时, 气候变化和人类活动会产生或影响生态系统服务(Paul et al, 2020), 包括气候变化对净初级生产力和水源供给的影响, 人类活动对粮食供给和生境质量的影响, 这些生态系统服务的变化会导致生物多样性的改变, 如区域内粮食供给的提升造成生物多样性的降低(Clark & Tilman, 2017), 城市发展过程中生境质量变化对生物多样性的影响(Bongaarts, 2019)等。目前已有较多研究关注生物多样性与生态系统服务的复杂联系, 但涉及的生态系统服务种类较少, 不能全面反映二者间的复杂关系和反馈机制, 有待进一步开展系统研究, 从而为生物多样性与生态系统服务的综合提升提供可靠的理论依据。
乌江流域位于我国西南地区中部, 生物多样性丰富, 承担着长江上游生态屏障功能, 但该地区目前仅有关于生物多样性或生态系统服务的研究。鉴于此, 本文基于乌江流域内植物物种空间分布格局, 应用MaxEnt模型、Zonation软件评估了乌江流域的生物多样性空间分布格局特征, 并使用InVEST模型核算了流域内三类六种生态系统服务, 再运用随机森林模型、部分依赖图模型(partial dependence plot, PDP模型)分析了生物多样性与各生态系统服务间的相互关系, 以期为当地生物多样性与生态系统服务的管理提升提供理论基础和参考。
1 材料与方法
1.1 研究区域概况
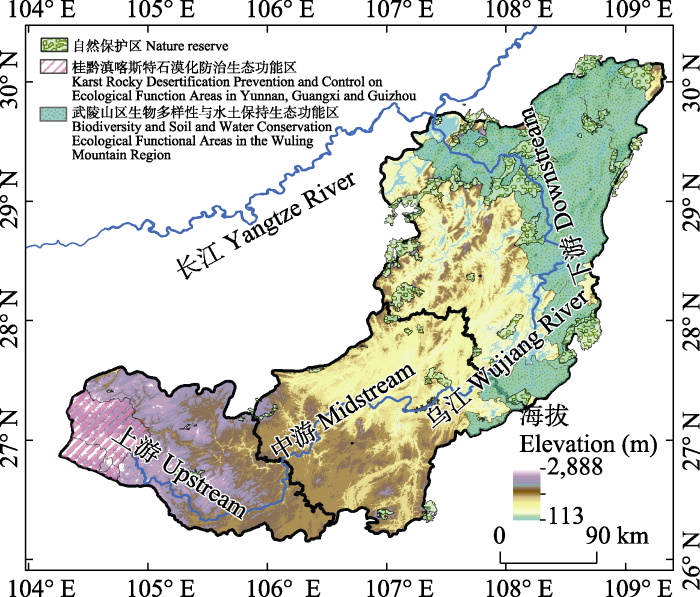
乌江是长江上游南岸的最大支流, 发源于贵州省赫章县, 自西向东汇集了多条支流, 在重庆市涪陵区汇入长江。乌江流域(104°18′-109°22′ E, 26°07′-30°22′ N)是典型的喀斯特地区, 位于我国第二阶梯向第三阶梯的过渡区, 海拔由西向东逐渐降低, 平均海拔为1,100 m, 面积为8.8 × 104 km2。乌江流域属亚热带季风气候, 年均气温13-18℃, 年均降水量900-1,400 mm; 化屋脊以上为乌江上游, 化屋脊至思南为乌江中游, 思南以下为乌江下游(杨俊毅等, 2023)。乌江流域的植物区系以温带成分为主, 热带成分为辅, 主要由阔叶林、针叶林、灌丛和草甸组成; 因地势起伏较大, 地貌类型多样, 植被垂直分异明显, 常绿阔叶林是该流域山地垂直地带谱中典型的基带, 也是优势垂直带, 植被水平分布呈西南低东北高的特点。境内生物多样性丰富, 生态功能重要, 拥有各类保护区53个以及两个国家重点生态功能区(图1)。
图1
1.2 数据来源及处理
1.2.1 物种分布数据
植物物种分布数据来源于中国数字植物标本馆(
1.2.2 环境因子
初步选取19种生物气候因子(附录1)、3种地形因子(海拔、坡度和坡向)、3种土壤因子(土壤有机质、土壤酸碱度和土壤质地)作为影响物种分布的环境变量。生物气候因子数据来源于WorldClim (
生态系统服务核算所使用的土地利用数据来源于Earth System Science Data (
1.3 研究方法
1.3.1 生物多样性评估
Zonation是一个用于大尺度生物多样性保护规划的软件, 是根据景观连通性和生物多样性特征(如物种分布、土地利用等), 逐步移除具有最小边际损失的栅格, 得到一个高连通性景观结构的嵌套栅格, 以此表征生物多样性保护等级, 并将此作为生物多样性评价指标。生物多样性保护等级的边际损失计算公式如下(Lehtomäki & Moilanen, 2013):
其中,
运行Zonation软件, 导入基于MaxEnt模型所得的各物种潜在分布, 以2020年的土地利用数据为条件层, 移除规则为附加效益函数(additive benefit function), 翘曲因子设置为1, 其余参数为默认值。
1.3.2 生态系统服务核算
基于千年生态系统评估中所包含的4类生态系统服务, 考虑到数据的可获得性、乌江流域的生态功能定位, 本文选取三类六种生态系统服务开展研究: 供给服务中的水源供给和粮食供给; 调节服务中的固碳服务和水源涵养; 支持服务中的土壤保持和生境质量。水源供给在InVEST模型的产水模块中计算(Huang et al, 2022); 粮食供给基于NPP计算(Zhang et al, 2020); 固碳服务在InVEST模型的碳模块结合空间校正的方法计算(杨俊毅等, 2023); 水源涵养基于水源供给, 综合地形指数、土壤饱和导水率和流速系数计算(曹明等, 2021); 土壤保持在InVEST模型的泥沙淤积模块中计算(Huang et al, 2022); 生境质量在InVEST模型的生境质量模块中进行评估(刘园等, 2019)。
1.3.3 生物多样性与生态系统服务的相互关系
随机森林模型是一种包含多个决策树的分类器算法, 多决策树的训练方式使其克服了过拟合问题, 且随机森林模型既能处理离散型数据, 也能处理连续型数据, 适用于包含多类变量的分析(Dai et al, 2022)。在使用随机森林模型进行回归分析的过程中, 将70%的数据作为训练数据集, 30%的数据作为验证数据集, 节点数设为500, 以得到各生态系统服务对生物多样性的贡献度。
部分依赖图模型是一种全局解释模型的方法, 可分析单个因子对模型结果的边际效应, 展示解释变量与目的变量间复杂的线性或非线性响应(Friedman, 2001)。本文利用部分依赖图模型定量分析了生物多样性与生态系统服务之间的响应特征。
2 结果
2.1 生物多样性空间分布格局
图2
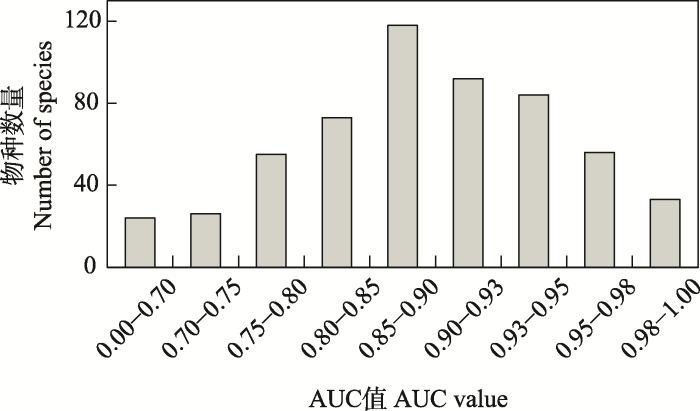
图2
MaxEnt模型所得561种植物物种AUC值的分布
Fig. 2
Distribution of the number of AUC values for 561 plant species from the MaxEnt model
图3
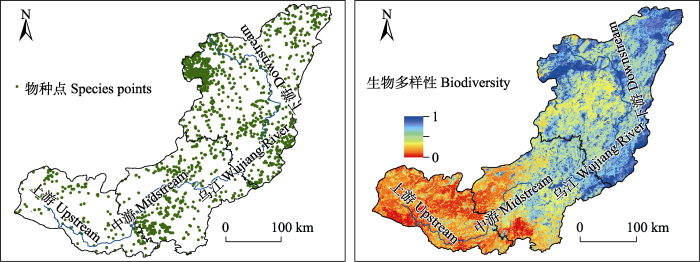
图3
乌江流域物种点分布和基于Zonation评估的生物多样性空间格局
Fig. 3
Species point distribution and spatial pattern of biodiversity based on Zonation assessment in the Wujiang River Basin
2.2 生态系统服务的空间格局
乌江流域的调节服务与支持服务均呈东北高西南低的分布格局, 最高区域均位于下游, 除土壤保持的最低区域位于中游外, 其余最低区域均位于上游; 供给服务中, 水源供给呈东北高西南低的分布格局, 但粮食供给呈西南高东北低的分布格局(图4, 表1)。2020年, 乌江流域的水源供给总量为7.12 × 1010 m3, 单位均值为810.57 mm/m2; 粮食供给总量为6.46 × 106 t, 单位均值为735.08 kg/ha; 固碳服务总量为1.15 × 103 Tg, 单位均值为131.30 Mg/ha; 水源涵养总量为2.00 × 1010 m3, 单位均值为227.98 mm/m2; 土壤保持总量为3.53 × 109 t, 单位均值为401.30 t/ha; 生境质量单位均值为0.68 (生境质量为指数, 统计中仅计算其均值)。
图4
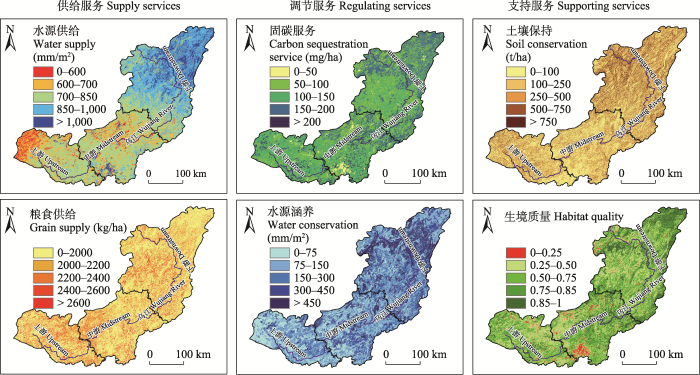
图4
2020年乌江流域6种生态系统服务的空间格局
Fig. 4
Spatial pattern of six ecosystem services in the Wujiang River Basin in 2020
表1 2020年乌江流域六种生态系统服务统计
Table 1
| 供给服务 Supply services | 调节服务 Regulating services | 支持服务 Supporting services | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 水源供给 Water supply | 粮食供给 Grain supply | 固碳服务 Carbon sequestration service | 水源涵养 Water conservation | 土壤保持 Soil conservation | 生境质量 Habitat quality | ||||||
| 均值 Mean (mm/m2) | 总量 Total (× 108 m3) | 均值 Mean (kg/ha) | 总量 Total (× 104 t) | 均值 Mean (Mg/ha) | 总量 Total (Tg) | 均值 Mean (mm/m2) | 总量 Total (× 108 m3) | 均值 Mean (t/ha) | 总量 Total (× 108 t) | 均值 Mean | |
| 上游 Upstream | 709.85 | 137.94 | 826.30 | 160.60 | 127.30 | 247.41 | 146.13 | 28.37 | 329.24 | 6.40 | 0.63 |
| 中游 Midstream | 741.94 | 187.51 | 737.81 | 186.49 | 129.74 | 328.00 | 203.06 | 51.28 | 272.42 | 6.88 | 0.66 |
| 下游 Downstream | 896.37 | 386.86 | 692.40 | 298.88 | 134.00 | 578.40 | 279.42 | 120.52 | 509.21 | 21.98 | 0.72 |
| 全流域 Total basin | 810.57 | 712.31 | 735.08 | 645.97 | 131.30 | 1,153.81 | 227.98 | 200.18 | 401.30 | 35.26 | 0.68 |
2.3 生物多样性与生态系统服务的相互关系
以生物多样性为因变量, 各项生态系统服务为自变量, 基于随机森林模型定量分析各生态系统服务对生物多样性的贡献度。供给服务中, 水源供给和粮食供给对生物多样性的贡献度分别为66.24%和27.43%; 调节服务中, 固碳服务和水源涵养对生物多样性的贡献度分别为19.91%和44.72%; 支持服务中, 土壤保持和生境质量对生物多样性的贡献度分别为26.18%和42.82%。
部分依赖图模型分析结果表明(图5), 除粮食供给外, 各生态系统服务与生物多样性均呈正相关。水源供给超过790 mm/m2时, 生物多样性大幅提高, 达到850 mm/m2后, 生物多样性趋于稳定; 粮食供给在0-700 kg/ha时, 生物多样性基本稳定, 超过700 kg/ha后, 生物多样性明显下降; 固碳服务在135-165 Mg/ha时, 生物多样性最高且保持稳定, 超过165 Mg/ha后, 生物多样性略有下降; 水源涵养超过90 mm/m2后, 生物多样性显著提高, 当超过260 mm/m2后, 生物多样性趋于稳定; 土壤保持在300-380 t/ha时生物多样性较高, 超过380 t/ha时生物多样性反而下降, 达到450 t/ha后, 生物多样性才呈增加趋势, 直至1,000 t/ha, 生物多样性趋于稳定; 生境质量超过0.3后生物多样性显著提高, 超过0.85后生物多样性趋于稳定。
图5
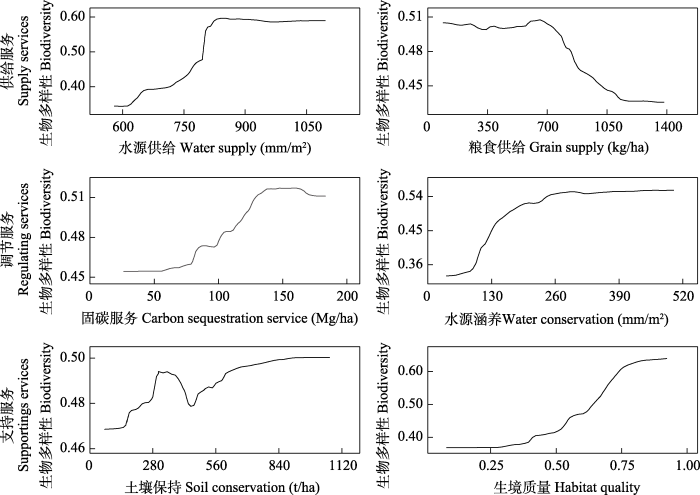
图5
2020年乌江流域生物多样性与6种生态系统服务间的非线性关系
Fig. 5
Nonlinear relationship between biodiversity and six ecosystem services in the Wujiang River Basin in 2020
3 讨论
本研究基于InVEST模型等遥感手段核算了生态系统服务, 但仅靠模型易出现数量级的差异, 因此核算结果需与实地数据进行对比分析。与2020年《长江流域及西南诸河水资源公报》(
从乌江流域各项生态系统服务的空间分布看, 总体为下游生态环境较好, 提供的生态系统服务较多, 中游次之, 上游较差, 但土壤保持与粮食供给的空间分布与此并不一致(图4)。土壤保持表现为下游 > 上游 > 中游, 可能是因为上游为桂黔滇喀斯特石漠化防治生态功能区所在区域, 是退耕还林工程、石漠化综合治理工程等大规模生态修复的重点实施区(Hu et al, 2022), 生态工程的实施会带来植被面积的增加, 而植被覆盖度和根系的增加有利于土壤保持服务的提高, 即使仍不如自然条件较好的下游, 但会稍高于中游地区。粮食供给表现为上游 > 中游 > 下游, 这是因为粮食是人类耕作生产的, 其空间分布会与人口密度相一致(赵敏等, 2022)。
基于随机森林所得的各生态系统服务对生物多样性的贡献度可以理解为二者间相互影响的程度, 但区分施加影响方和被影响方还需结合生态系统过程来分析。从各生态系统服务对生物多样性的贡献度来看, 供给服务、调节服务和支持服务并无明显差异, 但与水相关的生态系统服务表现出高于其他生态系统服务的影响度, 水源供给和水源涵养对生物多样性的贡献度分别为66.24%和44.72%。与水相关的生态系统服务受自然影响强烈, 有研究表明, 降水对水源供给的贡献度高达90%, 是典型的单气象因子驱动的生态系统服务(付刚, 2022)。在生态系统中, 降水直接影响水源供给的高低, 而生物多样性虽然会被降水所影响, 如降水过少导致的水源不足会降低生物多样性(曹明等, 2021), 但生物对降水变化的响应是滞后的, 因此受自然影响较大的生态系统服务不仅先于生物多样性改变, 其供给水平的高低也成为生物多样性的调节因素之一。粮食供给受人为干扰强烈, 人类是否耕作决定了农田是否有产出, 在农田生态系统中, 人类活动使得单一的粮食作物占据了其他植物的生存空间, 破坏了动物的栖息环境(Clark & Tilman, 2017), 造成生物多样性的下降, 这类生态系统服务供给水平的高低也是影响生物多样性变化的因素之一。对于固碳服务和土壤保持等来说, 生物多样性是它们的基础。生物多样性与生物功能多样性直接相关, 并影响生态系统功能, 生态位互补理论认为, 由于生态位差异, 高生物多样性群落中的生物能够占有更广的“功能空间”, 生物功能更丰富, 使群落的资源利用效率得到提高(Letten et al, 2017), 这就意味着生物多样性高的植物群落光能利用效率更高, 使得区域内光合作用增强, 进而提高固碳服务(Huang et al, 2018), 同时, 植物多样性丰富的地区地下根系更密集, 生态位分化更明显, 使得地上植物叶片功能多样性增加, 这些均有利于土壤保持服务的提高, Wen等(2019)在海南地区的研究中同样发现了植物叶片多功能可提高土壤保持服务。可见对于依赖生物功能的生态系统服务来说, 生物多样性是其调节因素。综上可知, 生态系统通过生物地球化学循环实现生态系统服务, 分析生物多样性与生态系统服务的关系要根据生态系统过程来甄别。生物多样性调节着依赖于生物功能而存在的生态系统服务; 对于受自然影响和人为干扰较大、先于生物多样性改变的生态系统服务, 生物多样性被它们所影响。
生物多样性与生态系统服务是相互影响的, 生态系统服务的变化因直接影响人类社会福祉而更易被大众感知(Paul et al, 2020), 如降水减少, 人类会普遍感知到水源供给的匮乏, 但难以察觉到生物多样性的丧失, 因此, 分析生物多样性随生态系统服务的变化对监管更有参考价值。本文基于乌江流域生物多样性与生态系统服务的响应特征, 发现当生态系统服务处于较低和较高水平时, 对生物多样性的影响均趋于稳定, 而当生态系统服务处于中间水平, 较小的变化就会显著影响生物多样性。乌江流域上游的生态系统服务和生物多样性水平较低, 如果通过适当的管理手段提高上游的生态系统服务水平, 会对生物多样性有较高的增益; 中游的生态系统服务和生物多样性处于中间水平, 生物多样性对生态系统服务变化较敏感, 生态系统服务的轻微变化就会对生物多样性产生较大影响, 因此需关注此地区生态系统服务的变化, 防止生态环境恶化影响生态系统服务进而造成生物多样性的大幅降低; 乌江下游的生态系统服务和生物多样性水平较高, 即使生态系统服务水平稍有降低, 生物多样性仍能保持较高的水平。
除粮食供给这种严重依赖人为活动的生态系统服务外, 乌江流域内的生物多样性与各生态系统服务均呈正相关。土壤保持服务在较低水平时, 会有较高的生物多样性, 这是因为土壤保持在核算时用的是土壤潜在侵蚀量减去实际土壤侵蚀量(Huang et al, 2022), 当部分地区自然环境较好、土壤潜在侵蚀量低时, 也会核算出低的土壤保持, 而这类地区会有较高的生物多样性。在固碳服务极高的地区, 生物多样性反而稍有下降, 这是因为乌江流域实施过多轮大规模生态修复工程, 种植了部分物种单一的人工林, 这类植被生长年限不长, 与成熟的天然林相比, 它们的光合作用更高(Hu et al, 2022), 但食物链和营养等级较少, 因此在固碳服务高的区域, 生物多样性反而稍有下降。水源供给、粮食供给等强烈受自然影响和人为干扰的生态系统服务是影响生物多样性的重要因素, 根据其与生物多样性的非线性关系可提出具有实际意义的管理意见。从响应特征来看, 当水源供给低于790 mm/m2时生物多样性会迅速降低, 因此在区域生物多样性保护中, 应随时关注降水量, 在易干旱地区, 应实施适当的人为干预, 如确保生态林的比例提高生态蓄水能力、搭配种植针阔叶树种提高流域产流等, 保证水源供给, 从而保护生物多样性; 生物多样性在粮食生产低于700 kg/ha时基本不变, 但当超过700 kg/ha后, 生物多样性急速下降, 目前流域内粮食产量为735.08 kg/ha, 在满足人类需求的前提下, 不宜再扩大粮食耕种面积, 并可适当减少。
附录 Supplementary Material
附录1 本文使用的19个生物气候因子
Appendix 1 19 bioclimatic factors used in this study
参考文献
Productivity is a poor predictor of plant species richness
DOI:10.1126/science.1204498
PMID:21940895
[本文引用: 1]

For more than 30 years, the relationship between net primary productivity and species richness has generated intense debate in ecology about the processes regulating local diversity. The original view, which is still widely accepted, holds that the relationship is hump-shaped, with richness first rising and then declining with increasing productivity. Although recent meta-analyses questioned the generality of hump-shaped patterns, these syntheses have been criticized for failing to account for methodological differences among studies. We addressed such concerns by conducting standardized sampling in 48 herbaceous-dominated plant communities on five continents. We found no clear relationship between productivity and fine-scale (meters(-2)) richness within sites, within regions, or across the globe. Ecologists should focus on fresh, mechanistic approaches to understanding the multivariate links between productivity and richness.
Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services
Assessing the effectiveness of water retention ecosystem service in Qinling National Nature Reserve based on InVEST and propensity score matching model
DOI:10.17520/biods.2020271 URL [本文引用: 2]
基于InVEST与倾向评分匹配模型评估秦岭国家级自然保护区水源涵养服务保护成效
Science for managing ecosystem services: Beyond the Millennium Ecosystem Assessment
Species diversity has a positive interrelationship with aboveground biomass and a mismatch with productivity in a subtropical broadleaf forest on the Wuyi Mountains, China
DOI:10.3390/d14110952
URL
[本文引用: 1]
The Wuyi Mountains in China are one of the largest and most complete subtropical forest ecosystems at the same latitude in the world. However, our limited understanding of the function of this distinctive ecosystem means that the interrelationship between species diversity and ecosystem functioning needs to be investigated if evergreen broadleaf forests (EBLFs) are to be protected. Here, we calculated species richness, the Shannon–Wiener index, and the Pielou index in large-, medium-, and small-scale (20 m × 20 m, 10 m × 10 m, and 5 m × 5 m) forest dynamics plots (FDPs). We also investigated the interrelationships between species diversity and aboveground biomass (AGB)/productivity in FDPs at the different scales using the generalized linear regression model. The results showed that species diversity and AGB positively interrelated with each other, whereas there was no species diversity-productivity interrelationship in the subtropical BLF on the Wuyi Mountains. The structural equation model (SEM) showed that elevation was a key factor directly affecting AGB. These findings lead to the conclusion that productivity is not solely correlated with species diversity and that abiotic factors probably mediate the positive interrelationship between species diversity and ecosystem functioning. This study provides theoretical basis that can be used to protect EBLFs in subtropical areas and the results can be used to improve the construction of artificial forest communities and vegetation restoration at the same latitude.
Comparative analysis of environmental impacts of agricultural production systems, agricultural input efficiency, and food choice
DOI:10.1088/1748-9326/aa6cd5 URL [本文引用: 2]
Influence of soil properties, topography, and land cover on soil organic carbon and total nitrogen concentration: A case study in Qinghai-Tibet Plateau based on random forest regression and structural equation modeling
DOI:10.1016/j.scitotenv.2022.153440 URL [本文引用: 1]
Greedy function approximation: A gradient boosting machine
The Trade-off and Synergy Between Landscape Pattern Evolution and Ecosystem Services in China in the Past 40 Years
近40年中国陆域景观格局演变与生态系统服务权衡和协同关系研究
博士学位论文, 北京师范大学, 北京.]
Vegetation restoration facilitates belowground microbial network complexity and recalcitrant soil organic carbon storage in southwest China karst region
DOI:10.1016/j.scitotenv.2022.153137 URL [本文引用: 3]
Generalized additive model reveals nonlinear trade-offs/synergies between relationships of ecosystem services for mountainous areas of Southwest China
DOI:10.3390/rs14122733
URL
[本文引用: 3]
Ecosystem services (ESs) are an essential link between ecosystems and human well-being, and trade-offs/synergies happen in ESs at different temporal and spatial scales. It is crucial to explore patterns of trade-offs/synergies among ESs, and their nonlinear relationships with changes in ESs. The primary objective of this study was to evaluate five ESs in 2000 and 2018: namely, water yield, food production, carbon sequestration, soil conservation, and habitat quality in mountainous regions of Southwest China. The mean values of the five ESs increased by 365.8 m3/ha, 13.92 t/hm2, 497.09 TgC/yr2, 138.48 t/km2, and 0.002, respectively. Using spatial statistics and analysis, an ES trade-off synergy model (ESTD) was constructed for the five ESs change values. Overall, soil conservation has a trade-off with all five ESs, except habitat quality; this trade-off is increasing slightly. Water yield is in synergy with all ESs except soil conservation, with decreasing synergy; habitat quality is in synergy with all ESs except food production, with increasing synergy. Finally, the nonlinear relationship between the value of the change in the ES and ESTD was analyzed using a generalized additive model. Changes in water yield showed the greatest impact on ESTD except for food production, wherein changes in all three ESs had minimal impacts on ESTD. Food production dominates its trade-offs/synergies relationship with soil conservation; carbon sequestration is the dominant player in its trade-offs/synergies relationship with soil conservation. Habitat quality has a secondary position of influence, except in the trade-offs/synergies involving food production. By exploring the drivers of trade-offs/synergies among ESs, this study can provide guidance for the effective implementation of policies related to ecological protection and restoration.
Impacts of species richness on productivity in a large-scale subtropical forest experiment
DOI:10.1126/science.aat6405
PMID:30287660
[本文引用: 2]
Biodiversity experiments have shown that species loss reduces ecosystem functioning in grassland. To test whether this result can be extrapolated to forests, the main contributors to terrestrial primary productivity, requires large-scale experiments. We manipulated tree species richness by planting more than 150,000 trees in plots with 1 to 16 species. Simulating multiple extinction scenarios, we found that richness strongly increased stand-level productivity. After 8 years, 16-species mixtures had accumulated over twice the amount of carbon found in average monocultures and similar amounts as those of two commercial monocultures. Species richness effects were strongly associated with functional and phylogenetic diversity. A shrub addition treatment reduced tree productivity, but this reduction was smaller at high shrub species richness. Our results encourage multispecies afforestation strategies to restore biodiversity and mitigate climate change.Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Relationship between biodiversity, ecosystem multifunctionality and multiserviceability: Literature overview and research advances
DOI:10.17521/cjpe.2020.0154
[本文引用: 1]
Over the recent decade, biodiversity and ecosystem multifunctionality (BEMF) has aroused as an emerging reserach hotspot in the filed of biodiversity and ecosystem functioning. Ecosystem multifunctionality is defined as the capacity of an ecosystem to provide multiple ecosystem functions simulateneously, it has received broad consideration by community and ecosystem ecologists. In this study, we first conducted a literature review of the research history in biodiversity and ecosystem multifunctionality. Next, we summarized the major trends in biodiversity and ecosystem multifunctionality research including the impacts of biodiversity dimensions, global change drivers and spatial-temporal scales on ecosystem multifunctionality. We reviewed the new research methods and research directions emerged in the field. We also defined a new concept, i.e., ecosystem multiserviceability (EMS) based on the distinction between ecosystem functions and ecosystem services. Finally, we briefly summarized the limitations in current research of biodiversity and ecosystem multifunctionality/multiserviceability (BEMF/BEMS) and presented the outlook for future study.
生物多样性与生态系统多功能性和多服务性的关系: 回顾与展望
DOI:10.17521/cjpe.2020.0154
[本文引用: 1]
近10年来, 生物多样性与生态系统多功能性(BEMF)的关系是生物多样性与生态系统功能领域新兴的热点研究方向。生态系统多功能性是指生态系统同时提供多重生态系统功能的能力, 受到群落和生态系统生态学研究者的广泛关注。该文简要回顾了生物多样性与生态系统多功能性关系研究历史, 侧重介绍了生态系统多功能性量化方法发展历程, 并总结了生物多样性与生态系统多功能性研究的主要趋势, 包括生物多样性维度、时空尺度和全球变化驱动因子等对生态系统多功能性的影响。同时, 回顾了近5年生物多样性与生态系统多功能性关系研究的新方法、新方向; 根据生态系统服务和生态系统功能的区别, 提出了生态系统多服务性(ecosystem multiserviceability, EMS)概念。最后简要介绍了生物多样性与生态系统多功能性、生物多样性与生态系统多服务性(BEMS)研究存在的不足及对未来的展望。
Complex relationships and feedback mechanisms between climate change and biodiversity
DOI:10.17520/biods.2022462
[本文引用: 1]
Background and Aims: Climate change and biodiversity loss are two major changes that human society is experiencing. Climate change affects all aspects of biodiversity and is a major driver of biodiversity loss; in turn, biodiversity loss exacerbates climate change. Therefore, halting or even reversing climate change and biodiversity loss is a global issue that needs to be addressed by human society. However, we lack a clear understanding of the complex relationships and feedback mechanisms between climate change and biodiversity. Here, we summarize the research on climate and biodiversity change in the last decade by focusing on studies investigating the responses and feedback of biodiversity to climate change at different organizational levels, spatial scales, and diversity dimensions. Progress: Our results showed that most studies focus on the direct impacts of climate change on biodiversity, involving different organizational levels and dimensions and trophic levels of biodiversity. Studies on the indirect impacts of climate change were rare, and we suggested that mechanistic studies need to be strengthened. The mechanisms and quantification of the effects of biodiversity on ecosystem multifunctionality were challenges for current research. There was no consensus on how biodiversity contributes to ecosystem response to climate change; the positive and negative feedback effects of biodiversity in the context of climate change were a blind spot in domestic and international research. Prospects: The future direction and key scientific issues that need to be solved in the field of climate change and biodiversity change are numerous. We identify 4 main areas of future research: understanding (1) the impacts of multi-factor climate change on biodiversity, (2) how mitigation and adaptation measures to climate change can benefit biodiversity conservation, (3) how the theory of biodiversity and ecosystem function can be applied to the real-world ecosystems and (4) what is the contribution of biodiversity conservation to carbon neutrality goals.
气候变化与生物多样性之间的复杂关系和反馈机制
DOI:10.17520/biods.2022462
[本文引用: 1]
气候变化与生物多样性丧失是人类社会正在经历的两大变化。气候变化影响生物多样性的方方面面, 是导致生物多样性丧失的一个主要驱动因子; 反过来, 生物多样性丧失会加剧气候变化。因此, 阻止甚至扭转气候变化和生物多样性丧失是当前人类社会亟需解决的全球性问题,但我们对气候变化与生物多样性之间的复杂关系和反馈机制尚缺乏清晰认识。本文总结了近年气候变化与生物多样性变化的研究进展, 重点概述了不同组织层次、空间尺度和维度的生物多样性对气候变化的响应和反馈等相关领域的研究进展和存在的主要问题。结果发现多数研究关注气候变化对生物多样性的直接影响, 涉及到生物多样性的不同组织层次、维度和营养级, 但针对气候变化间接影响的研究仍然较少, 机理研究同样需要加强; 生物多样性对生态系统功能影响的环境依赖和尺度推演、生物多样性对生态系统多功能性的作用机理和量化方法是当前研究面临的挑战; 生物多样性对生态系统响应气候变化的作用机制尚无统一的认识; 生物多样性对气候变化的正、负反馈效应是国内外研究的盲点。最后, 本文展望了未来发展方向和需要解决的关键科学问题, 包括多因子气候变化对生物多样性的影响; 减缓和适应气候变化的措施如何惠益于生物多样性保护; 生物多样性与生态系统功能的理论如何应用到现实世界; 生物多样性保护对实现碳中和目标的贡献。
Plant diversity increases soil microbial activity and soil carbon storage
DOI:10.1038/ncomms7707
PMID:25848862
[本文引用: 1]
Lange, Markus; Sierra, Carlos A.; Mellado-Vazquez, Perla G.; Malik, Ashish A.; Steinbeiss, Sibylle; Trumbore, Susan E.; Gleixner, Gerd Max Planck Inst Biogeochem, D-07701 Jena, Germany. Eisenhauer, Nico Univ Jena, Inst Ecol, D-07743 Jena, Germany. Eisenhauer, Nico German Ctr Integrat Biodivers Res (iDiv) Halle Je, D-04103 Leipzig, Germany. Eisenhauer, Nico Univ Leipzig, Inst Biol, D-04103 Leipzig, Germany. Bessler, Holger; Engels, Christoph Humboldt Univ, Fac Life Sci, Div Plant Nutr, D-10099 Berlin, Germany. Griffiths, Robert I.; Thomson, Bruce C. Ctr Ecol & Hydrol, Wallingford OX10 8BB, Oxon, England. Roy, Jacques Ctr Natl Rech Sci, Ecotron UPS 3248, F-34980 Montferrier Sur Lez, France. Scheu, Stefan Univ Gottingen, JF Blumenbach Inst Zool & Anthropol, D-37073 Gottingen, Germany.
Methods and workflow for spatial conservation prioritization using Zonation
Linking modern coexistence theory and contemporary niche theory
DOI:10.1002/ecm.2017.87.issue-2 URL [本文引用: 1]
Identifying conservation priority areas for gymnosperm species under climate changes in China
DOI:10.1016/j.biocon.2020.108914 URL [本文引用: 2]
Study on the spatio-temporal patterns of habitat quality and its terrain gradient effects of the middle of Yangtze River economic belt based on the InVEST model
基于InVEST模型的长江中游经济带生境质量的时空分异特征及其地形梯度效应
Biodiversity and ecosystem services: A multilayered relationship
DOI:10.1016/j.tree.2011.08.006 URL [本文引用: 1]
Biodiversity and ecosystem function
DOI:10.1126/science.1217245 PMID:22246761 [本文引用: 1]
Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology
DOI:10.1111/1365-2664.12669 URL [本文引用: 1]
On the functional relationship between biodiversity and economic value
DOI:10.1126/sciadv.aax7712
URL
[本文引用: 4]
This study reviews the conditions under which a positive link between biodiversity and economic value may hold.
Maximum entropy modeling of species geographic distributions
DOI:10.1016/j.ecolmodel.2005.03.026 URL [本文引用: 2]
Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change
DOI:10.1016/j.jnc.2014.03.007 URL [本文引用: 2]
Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis
DOI:10.1126/science.1172460
PMID:19644076
[本文引用: 1]
Ecological restoration is widely used to reverse the environmental degradation caused by human activities. However, the effectiveness of restoration actions in increasing provision of both biodiversity and ecosystem services has not been evaluated systematically. A meta-analysis of 89 restoration assessments in a wide range of ecosystem types across the globe indicates that ecological restoration increased provision of biodiversity and ecosystem services by 44 and 25%, respectively. However, values of both remained lower in restored versus intact reference ecosystems. Increases in biodiversity and ecosystem service measures after restoration were positively correlated. Results indicate that restoration actions focused on enhancing biodiversity should support increased provision of ecosystem services, particularly in tropical terrestrial biomes.
Remotely sensed carbon content: The role of tree composition and tree diversity
DOI:10.1016/j.rse.2022.113333 URL [本文引用: 1]
Research progress on the relationship between biodiversity and ecosystem services
DOI:10.13287/j.1001-9332.202001.003
[本文引用: 2]
The relationship between biodiversity and ecosystem services is one of the hot topics in ecological research. The ways that the relationship is applied to the development of ecosystem management and policy have gradually gained attention after the Millennium Ecosystem Services Assessment in 2005. However, applying theoretical understanding of the relationship into practical management still faces challenges. Examining recent progresses may help to guide practices and po-licies. We summarized recent progress in researches on the relationship between biodiversity and ecosystem services in terms of the impacts of biodiversity on single ecosystem service, ecosystem multifunctionality and trade-offs of ecosystem services, as well as how environmental changes affected these relationships. Moreover, we outlined the application of these relationships in nature reserve management, forest management, degraded ecosystem restoration, and agro-ecosystem improvement. We further analyzed the shortcomings in relationship studies and then emphasized that future research and practice trends are the interaction of different biodiversity components and multiple trophic levels on ecosystem services, the coupling effects of environmental changes on the relationships, and practical approaches to biodiversity for improving ecosystem services.
生物多样性与生态系统服务关系研究进展
DOI:10.13287/j.1001-9332.202001.003
[本文引用: 2]
生物多样性与生态系统服务关系是当前生态学研究的热点之一,2005年千年生态系统评估之后如何将两者关系应用到生态系统管理和政策制定中逐渐受到重视。然而,从理论上的两者关系认知到应用上的管理实践仍面临着严峻挑战,梳理已有研究成果有利于更好地指导管理实践。本文基于近年生物多样性与生态系统服务关系的主要研究成果,从生物多样性对生态系统单种服务、多种服务、生态系统服务权衡的影响以及环境变化如何影响这些关系等方面,总结了两者关系的主要研究进展,概述了两者关系在自然保护区管理、森林生态系统管理、退化生态系统恢复和农业生态系统改善等领域中的应用,分析了两者关系研究中依然存在的不足之处,强调未来需加强的研究方向主要包括: 生物多样性不同组分相互作用和多营养级生物多样性相互关系对生态系统服务的影响、环境耦合变化对两者关系的复合效应以及生物多样性改善生态系统服务的实践途径。
Functional diversity overrides community-weighted mean traits in linking land-use intensity to hydrological ecosystem services
DOI:10.1016/j.scitotenv.2019.05.160 URL [本文引用: 3]
Estimations of forest water retention across China from an observation site-scale to a national-scale
DOI:10.1016/j.ecolind.2021.108274 URL
Spatio-temporal pattern and driving mechanism of ecosystem carbon sequestration services in the Wujiang River Basin
乌江流域固碳服务时空格局及驱动机制
DayCent model predictions of NPP and grain yields for agricultural lands in the contiguous U.S.
Evolution and coupling between economic and population spatial pattern in Wujiang River Basin
乌江流域经济与人口空间格局演变及耦合特征
Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China
DOI:10.1016/j.scitotenv.2022.156867 URL

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}