



滨海特有植物是指仅分布于海岛或滨海地区的具有明显海岛特色的维管植物(陈征海等, 1995; 施德法等, 1996)。海岛作为滨海特有植物的主要栖息地, 因其特殊的生境、与大陆的长期隔离, 形成了与相近纬度的大陆差异化的种质资源。滨海特有植物受分布范围狭窄、种群规模小等内在因素制约, 濒危或灭绝的风险远高于陆地上分布的物种(Francisco-Ortega et al, 2000), 而气候变化、外来物种入侵、生境片段化等外在影响又加剧了其潜在灭绝风险。因此, 探明海岛环境中特有植物种质资源, 尤其是滨海特有植物的种质资源和分布机制, 将有助实现基于生物多样性的海岛滨海特有植物种质资源保护和利用的重要愿景。
海岛的种-面积关系(island species-area relation-ship, ISAR)描述了海岛物种数量随着采样面积增加而增加的现象(Whittaker & Fernández-Palacios, 2007; Kreft et al, 2008)。影响海岛的种-面积关系斜率和截距的因素陆续被提出(Rosenzweig, 1995), 这其中主要包括迁移和灭绝之间的平衡、面积导致的扰动速率(MacArthur & Wilson, 1967; Whittaker et al, 2008; Pinheiro et al, 2017)等。海岛的物种丰富度除了遵循ISAR外, 隔离程度(Ricklefs, 1987; Fahrig, 2013)、海拔(Triantis et al, 2003; Rojas-Sandoval et al, 2020)、温度(Kalmar & Currie, 2006; Kubota et al, 2015; Yu et al, 2019)、降水(Heatwole, 1991; Valli et al, 2019; Wang et al, 2019)、风速(Irl et al, 2021)以及与生境异质性相关的海岛形状指数(shape index, SI)和周长面积比(perimeter area ratio, PAR) (Honnay et al, 2003; Sfenthourakis & Triantis, 2009)等因素也会对其产生影响(Borges & Hortal, 2009; Cabral et al, 2014; Kubota et al, 2015; Liu et al, 2018)。但在许多情况下, 环境因子对海岛物种丰富度的预测能力在不同空间尺度、地理区位中存在明显差异。
除环境因素外, 物种功能特征的差异也可能是影响物种丰富度的重要因素(Ewers & Didham, 2006)。由于不同生活型植物的生境偏好、物种扩散能力不同(Chisholm et al, 2016), 导致其对生境片段化的敏感性存在差异(Cagnolo et al, 2009; Irl et al, 2020)。对滨海特有植物不同生活型物种丰富度的比较研究可以提供其对特定环境因子的响应、空间利用以及在群落中可能存在的竞争关系等信息(Schrader et al, 2020)。因此, 研究环境因子对滨海特有植物不同生活型物种丰富度的影响, 可为了解面积、景观、气候以及人类行为如何影响其分布格局提供重要依据。
福建海岛众多, 面积和隔离距离跨度广, 岛屿形状、海岸线长度、海拔等景观特征多样, 是开展岛屿植物分布和环境驱动因子研究的理想区域。福建地处亚热带海洋性季风气候区, 能够反映我国亚热带海岛的植物多样性特征。同时由于福建海岛上具有丰富的濒危和特有植物资源, 因此成为开展海岛生物多样性和物种资源保护利用研究的重要地区之一。然而, 频繁的人为活动和海岛经济开发对福建海岛的植物多样性和生态系统产生了威胁。因此, 探明滨海特有植物物种丰富度与环境因子之间的关系, 将为相关部门开展海岛植物资源的保护和开发利用提供建议。本研究以福建53个海岛为对象, 首先描述了福建滨海特有植物的组成; 其次讨论了滨海特有植物及不同生活型植物的种-面积关系; 最后着重分析了表征景观、气候以及人为干扰3个方面的10个环境因子在驱动滨海特有植物及不同生活型植物物种丰富度中的作用, 以解释福建海岛滨海特有植物及不同生活型植物的分布格局。
1 材料与方法
1.1 研究区概况
福建省地处中国东南沿海地区, 临近台湾海峡。福建的海岛均为大陆岛(continental island), 地貌以丘陵、台地、平原、风成沙地为主。土壤主要为赤红壤、红壤、风沙土和滨海盐土。气候为亚热带海洋性季风气候, 闽江口及其以南为南亚热带海洋性季风气候, 闽江口以北为中亚热带海洋性季风气候(福建省海岛资源综合调查委员会, 1996)。年均温15.2-21.0℃, 年均降水量1,000-1,600 mm。海岛植被类型主要由常绿针叶林、常绿阔叶林和灌草丛群落等组成。福建的海岛数量超过1,500个, 但许多海岛为面积小、有植被覆盖的出水岛礁, 且福建海岛分布零散, 多数无人岛登岛困难。因此本研究结合实际情况只选取福建海域(117°31′61″-120°68′66″ E, 23°60′90″-26°93′08″ N)范围内具有登岛条件和植被覆盖的海岛, 为反映景观、气候、人为干扰的差异, 选取了不同面积、海拔、隔离程度、不同人为活动程度以及不同气候条件等具有环境梯度的海岛。
本文共选取了53个海岛(图1), 包括塔屿、对面屿、黄干岛、大坠岛、黄湾屿等在内的44个无居民海岛以及海坛岛、惠屿、琅岐岛、大练岛、岱嵩岛等在内的9个有居民海岛。海岛平均面积13.39 km2, 最小的仅0.007 km2, 最大的267.13 km2 (海坛岛), 其中84.90% (45个)的海岛面积小于1 km2。海岛平均海拔70.34 m (13.98-438.20 m) (附录1)。
图1
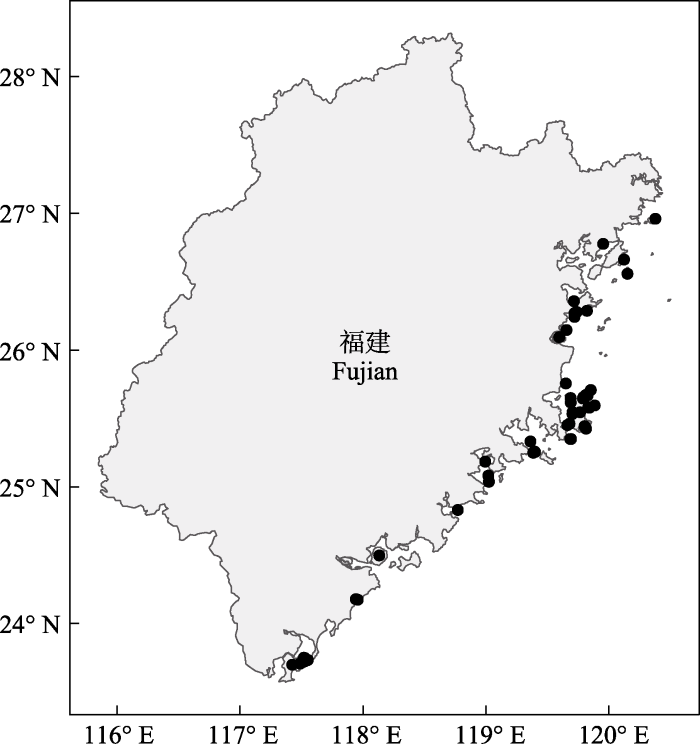
图1
本研究选取的福建53个海岛的分布图(海岛名称见附录1)
Fig. 1
Distribution of the 53 islands in Fujian selected for this study (see Appendix 1 for islands name)
1.2 物种数据获取
分别在2018年和2022年的生长季节(5-7月、9-11月)对53个海岛上的所有植物物种进行调查。调查方式以样线调查为主、样方调查为辅, 同时在海岛的边缘以及悬崖等无法进入的地带, 采用无人机航拍进行辅助调查。对面积不超过1 km2的小型海岛(n = 45)进行全岛踏查(王金旺等, 2017): 每个海岛布设3-10条样线, 每条样线至少步行2次(Yu et al, 2012)。在面积1-10 km2的海岛(n = 3)上, 每个海岛建立6-20条样线, 每个海岛样线总长度均大于12 km。在面积大于10 km2的海岛(n = 5)上, 同样进行了如上所述的调查, 但同时以每个海岛上突出的山体为中心再进行样线的布设(Hu et al, 2011), 每个大岛的样线总长度均大于25 km, 详细记录样线内的物种信息。
为保证物种记录详尽, 在样线调查后确定每个海岛的典型群落类型, 再以标准样方法进行样方设置, 样方设置涵盖不同类型的植物群落。依据海岛典型植物群落情况设置20 m × 20 m的森林样方, 53个海岛共设置了675个。于森林样方的对角选取4个5 m × 5 m的灌木样方, 同时在样方四角以及中心点选取5个1 m × 1 m的草本样方。详细记录乔木层出现的全部树种, 对胸径(DBH) ≥ 3 cm或树高 ≥ 3 m的植株测量胸径和高度, 记录种名、数量、地径、冠幅和生长状况等, 记录灌木层(DBH < 3 cm或树高 ≤ 3 m)的种名、数量、地径、盖度、高度和生长状况, 记录草本层的种名、多度、平均高度和盖度。同时详细记录每个样方的经纬度、海拔、坡度、地形、郁闭度、生境类型等信息(方精云等, 2009), 现场拍摄照片并采集标本, 凭证标本存放于福建农林大学风景园林与艺术学院标本室。
参考《中国植物志》对植物生境的描述, 筛选具有滨海特色的滨海特有植物(地理分布上通常为海岛和海岸带), 获得福建53个海岛的滨海特有植物数据, 并依据生活型将其划分为乔木、灌木和草本3类。
1.3 海岛环境因子
为分析海岛环境因子对滨海特有植物及不同生活型植物物种丰富度的影响, 共选取景观、气候、人为干扰3类共10个变量, 其中面积(area)、海拔(elevation)、距大陆距离(distance to the mainland, DM)、距最近大岛距离(distance to the nearest big island, DN)、周长面积比、形状指数6个变量为景观因子; 年均风速(annual mean wind, AMW)、年均温(annual mean temperature, AMT)、年降水量(annual precipitation, AP) 3个变量为气候因子; 建筑与农田面积占比(proportion of buildings and farmland area, BFA)为人为干扰因子(附录1)。
海岛的面积和周长、距大陆距离、距最近大岛距离、建筑与农田面积占比数据来源于中国科学院空天信息创新研究院发布的2020年全球30 m精度地表覆盖矢量地图(
年均风速数据来源于NOAA 10 m精度的风速矢量地图(
建筑与农田面积占比为海岛建筑与农田面积之和与海岛面积的比值。海岛的面积、周长这两个变量用于生成周长面积比和形状指数。周长面积比是海岛的边缘占面积的比值, 表示海岛边缘生境相对于海岛内部生境的相对数量, 表征海岛的边缘效应(Wu, 2004)。形状指数表征海岛形状的复杂程度, 当海岛形状为圆形时, 其形状指数为1, 随着形状越来越复杂和不规则, 形状指数也会随之升高。其公式为SI = C/[2 × (π × A)0.5] (Hoffmeister et al, 2005), 其中C为海岛的周长, A为海岛的面积。距大陆距离(DM)、距最近大岛距离(DN)用以表征海岛的隔离程度(Yu et al, 2012)。
1.4 数据分析
在构建模型前使用方差膨胀因子(VIF)检测环境因子之间的共线性, 10个环境因子的VIF均小于6, 低于最大阈值10 (Neter et al, 1996), 说明其共线性较弱。采用物种累计曲线判断对福建海岛滨海特有植物的调查是否充分。
基于R的sars包, 使用对数转换的幂函数模型logS = logc + z × logA来表征滨海特有植物以及不同生活型植物物种丰富度与海岛面积之间的种-面积关系。其中S是物种丰富度, c和z是常数。z表示物种丰富度随面积变化的速率, 通常用作衡量物种、群落对片段化生境的敏感性(Yu et al, 2012)。
使用Pearson相关系数检测海岛环境因子之间的相关性, 通过Bonferroni校正调整显著性水平。采用Global Moran’s I检测环境因子的空间自相关, 并采用Mantel检验基于欧式距离分析海岛环境因子矩阵与滨海特有植物及不同生活型植物的物种有无矩阵之间的相关性。
为探讨海岛环境因子对滨海特有植物以及不同生活型物种丰富度的影响, 对自变量和因变量进行标准化处理, 使用全子集回归, 根据小样本校正后的赤池信息量准则(AIC)即AICc等对子集排序, 从中获取最优子集的方法作为最佳模型选择的标准。在此基础上, 使用层次分割对最佳拟合模型中的环境因素进行评估, 定量分析不同环境变量的解释率。
以上数据分析均在R 4.2.2中完成。
2 结果
2.1 滨海特有植物组成
本研究调查的福建53个海岛共有滨海特有植物40科85属101种(含种下分类单位), 包括乔木5种, 占总物种数的4.95%; 灌木26种, 占25.74%; 草本70种, 占69.31%。其中国家重点保护植物有珊瑚菜(Glehnia littoralis) (极危, CR)和海滨藜(Atriplex maximowicziana) (濒危, EN); 中国特有植物有滨海白绒草(Leucas chinensis)、福建胡颓子(Elaeagnus oldhamii)、海岸扁担杆(Grewia piscatorum)、华南画眉草(Eragrostis nevinii)、九里香(Murraya exotica)、上狮紫珠(Callicarpa siongsaiensis)等, 而上狮紫珠、海滨藜也是福建特有种。
物种累计曲线(图2)表明, 随着岛屿数量的增加, 滨海特有植物以及乔木、灌木、草本物种数的曲线均呈先上升后增速放缓的趋势。虽然由于乔木在海岛上分布数量较少导致其曲线表现较为平缓, 但总体数据合理, 表明53个海岛滨海特有植物调查充分。
图2
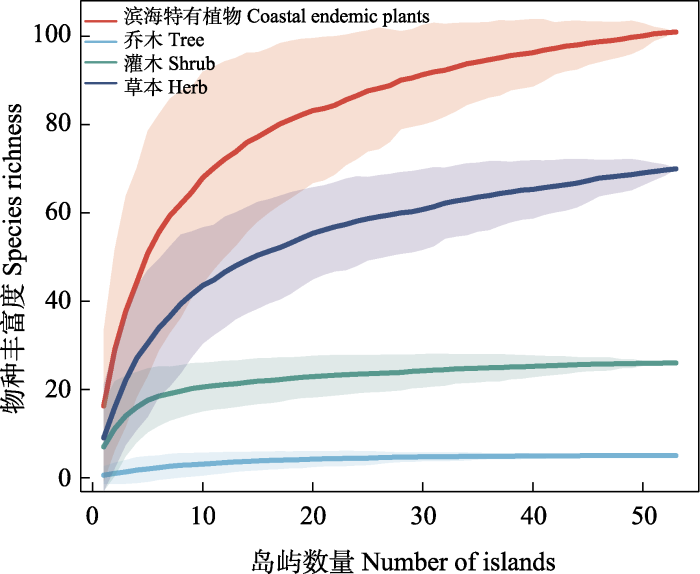
图2
福建滨海特有植物及不同生活型植物的物种累计曲线
Fig. 2
Species accumulation curve of coastal endemic plants and different growth forms of plants in Fujian
2.2 种-面积关系
滨海特有植物及乔木、灌木和草本的种-面积关系拟合均为正相关, 滨海特有植物物种丰富度表现出随着面积增加而增加的趋势(图3)。面积对滨海特有植物丰富度的解释率分别为: 滨海特有植物45%、乔木88%、灌木23%、草本32%。种-面积关系的斜率(z值)分别为滨海特有植物0.16、乔木0.15、灌木0.15、草本0.14。
图3
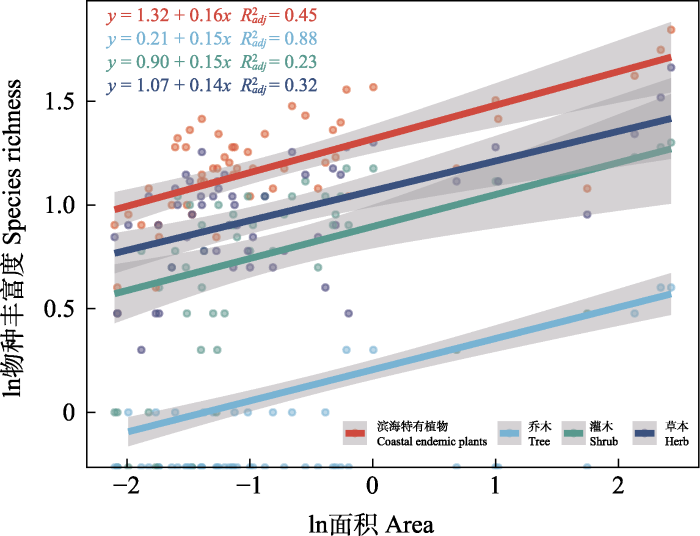
图3
福建滨海特有植物和不同生活型植物的种-面积关系
Fig. 3
The species-area relationship about coastal endemic plants and different growth forms of plants in Fujian
2.3 滨海特有植物与海岛环境因子
根据海岛环境因子之间的相关性分析可知(图4), 面积与海拔(r = 0.82, P < 0.001)、形状指数(r =0.30, P < 0.05)、建筑与农田面积占比(r = 0.50, P < 0.001)呈显著正相关, 而与周长面积比(r = -0.40, P < 0.01)、年均风速(r = -0.45, P < 0.001)呈极显著负相关。周长面积比与海拔呈显著负相关(r = -0.63, P < 0.001)。距大陆距离与其他环境因子之间的相关性最弱, 仅与年均风速具有极显著的正相关关系(r = -0.46, P < 0.001)。Global Moran’s I分析表明, 距大陆距离(Moran’s I = 0.90, P < 0.001)具有极显著的空间自相关。然而, Mantel检验表明, 距大陆距离对滨海特有植物及不同生活型植物的物种丰富度均未产生显著影响(P > 0.05)。
图4
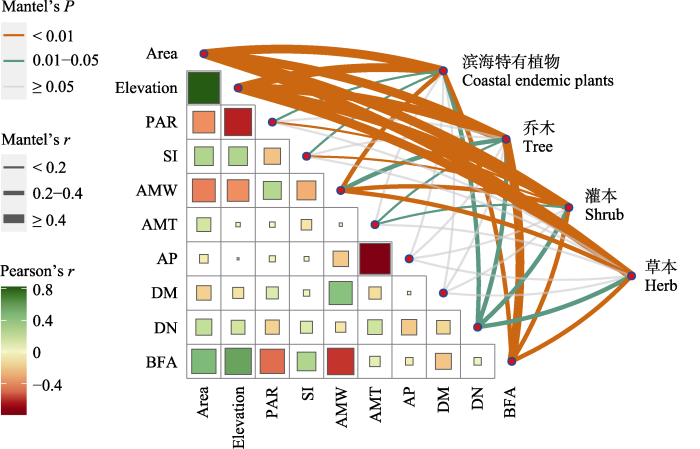
图4
环境因子之间的Pearson相关性分析和Mantel检验。Area: 面积; Elevation: 海拔; PAR: 周长面积比; SI: 形状指数; AMW: 年均风速; AMT: 年均温; AP: 年降水量; DM: 距大陆距离; DN: 距最近大岛距离; BFA: 建筑与农田面积占比。* P < 0.05; ** P < 0.01; *** P < 0.001。
Fig. 4
Pearson correlation and Mantel test between environmental factors. Area, Area; Elevation, Elevation; PAR, Perimeter area ratio; SI, Shape index; AMW, Annual mean wind; AMT, Annual mean temperature; AP, Annual precipitation; DM, Distance to the mainland; DN, Distance to the nearest big island; BFA, Proportion of buildings to farmland area. * P < 0.05; ** P < 0.01; *** P < 0.001.
基于全子集回归筛选出的最佳模型(图5)表明, 景观中的面积、形状指数和周长面积比是影响滨海特有植物和不同生活型植物物种丰富度最主要的因子。面积和形状指数对滨海特有植物及乔木、灌木和草本的物种丰富度均有显著的正向影响, 而滨海特有植物以及灌木和草本的物种丰富度则是随着周长面积比的增加而减少。隔离对滨海特有植物及不同生活型植物的物种丰富度影响较弱, 乔木的物种丰富度随着距最近大岛距离的增加而增加, 但灌木的物种丰富度随着距大陆距离的增加而降低。人为干扰中建筑与农田面积占比对滨海特有植物及不同生活型植物的物种丰富度均无显著影响。而对于气候方面而言, 除了乔木物种丰富度外, 年均风速对滨海特有植物以及灌木和草本的物种丰富度均有显著的正向影响。
图5
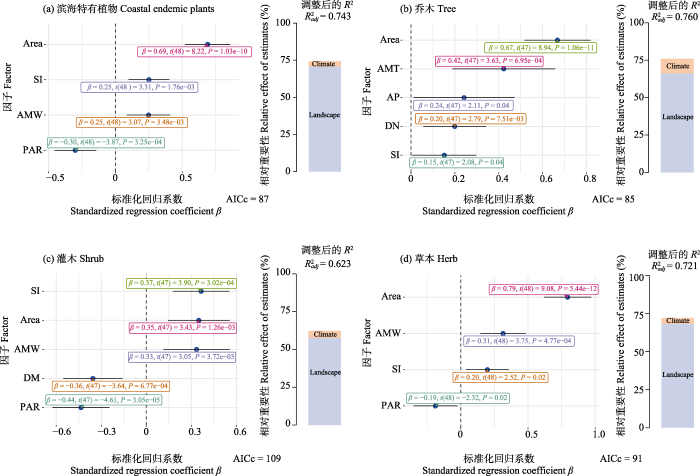
图5
滨海特有植物以及不同生活型物种丰富度的最佳模型选择和层次分割分析。图中森林图为使用全子集回归, 并根据最小AICc值筛选得出的最优模型, 森林图中虚线左侧为负相关, 右侧为正相关。柱形图为基于最优模型的层次分割分析。Area: 面积; Elevation: 海拔; PAR: 周长面积比; SI: 形状指数; AMW: 年均风速; AMT: 年均温; AP: 年降水量; DM: 距大陆距离; DN: 距最近大岛距离; BFA: 建筑与农田面积占比。
Fig. 5
The best model selection and hierarchical partitioning analysis of coastal endemic plants and different growth forms of species richness. The forest plot in the figure shows the optimal model obtained by using full subset regression and screening based on the minimum AICc value. The left side of the dashed line in the forest plot shows a negative correlation, while the right side shows a positive correlation. The column chart is a hierarchical segmentation analysis based on the optimal model. Area, Area; Elevation, Elevation; PAR, Perimeter area ratio; SI, Shape index; AMW, Annual mean wind; AMT, Annual mean temperature; AP, Annual precipitation; DM, Distance to the mainland; DN, Distance to the nearest big island; BFA, Proportion of buildings to farmland area.
层次分割结果表明, 对于滨海特有植物和不同生活型植物的物种丰富度而言, 景观因子对比气候和人为干扰方面贡献了更多的解释率(滨海特有植物70.63%、乔木66.15%、灌木57.59%、草本68.17%) (图5)。气候因子也对滨海特有植物(3.70%)、乔木(9.82%)、灌木(4.67%)、草本(3.92%)贡献了一部分的解释率。
3 讨论
本研究发现滨海特有植物及不同生活型植物的物种丰富度呈现随着面积的扩大而增加的现象。在种-面积关系的研究中, z值范围通常在0.20和0.35之间(Panitsa et al, 2006; Santos et al, 2010)。本研究中滨海特有植物及不同生活型植物种-面积关系的z值(斜率)为0.14-0.16。这表明随着福建海岛面积的变化, 滨海特有植物及不同生活型植物的物种丰富度变化速率相对较慢(Yu et al, 2012)。此外, 与其他海岛生态系统相比, 福建海岛滨海特有植物及不同生活型植物物种丰富度对海岛生境片段化的敏感性较弱。这可能是由于福建海岛以木麻黄(Casuarina equisetifolia)、台湾相思(Acacia confusa)、黑松(Pinus thunbergii)等优势群落为主, 生境类型较为单一, 能提供的空余生态位有限, 因而导致滨海特有物种随海岛面积增加的速率减慢(Rosenzweig, 1995)。另一种可能是海岛长期被海水包围, 与大陆隔离, 而仅分布在海岛或滨海地区的滨海特有植物或许早已适应了片段化的海岛生境, 从而导致其具有较低的z值(Hu et al, 2011)。
在岛屿生物地理学的研究中, 许多结果都强调了生物因素和非生物因素对海岛物种丰富度的影响(Steinbauer et al, 2017; Rojas-Sandoval et al, 2020)。但对于气候变化与人类活动影响敏感的海岛滨海特有植物而言, 其作用程度尚不明晰。在通过层次分割分析所有海岛环境因子的解释率时, 发现景观对滨海特有植物和不同生活型植物物种丰富度的贡献更大, 其中面积是决定滨海特有植物及不同生活型植物物种丰富度的主导因素, 这一结果支持岛屿生物地理学研究中面积是决定物种丰富度的主要因素的假设(Kalmar & Currie, 2006)。生境多样性假说认为, 面积较大的海岛比小型海岛具有更多的生境数量, 能提供更多的生态位(Chen et al, 2020), 支撑更多的物种和更大的种群(Kohn & Walsh, 1994), 有助于滨海特有植物的增加。与此同时, 海洋环境(大风、盐雾、风暴潮)作为环境过滤器(Mologni et al, 2021)也同样会影响到海岛上物种的分布, 随着海岛面积的增加, 海洋与海岛内部的距离增加, 植物受海洋环境的影响可能减少, 从而也将促进物种数的增加(Neufeld et al, 2017)。
同时, 表征海岛形状复杂程度的形状指数和表征海岛边缘生境相对于内部生境数量的周长面积比也是与滨海特有植物密切相关的因素。在海岛环境因子间的相关性分析(图4)中, 海岛面积和形状指数之间的正相关关系表明面积较大的海岛具有更复杂的形状; 海岛面积和周长面积比的负相关关系表明海岛面积越大, 形状越复杂, 海岛的相对内部生境则更多。因此当综合考虑面积因素后, 具有更复杂形状(高形状指数)和更多的岛屿内部生境(低周长面积比)的海岛可以支持更多的物种分布, 这也侧面反映了面积、形状指数和周长面积比与生境异质性的正相关关系(Yu et al, 2012)。这与前人研究得出的结论相一致, 即随着生境异质性的增加, 物种多样性会随着面积的增加而增加(MacArthur & Wilson, 1963; Lomolino, 2000)。在对全球700个海岛的研究中也发现代表生境异质性的环境因子才是影响物种丰富度的决定因素(Schrader et al, 2020)。
在岛屿生物地理学研究中, 隔离被广泛认为是影响物种丰富度的一个主要因素(Chiarucci et al, 2011; Cabral et al, 2014)。然而本研究发现福建海岛滨海特有植物与隔离度并没有像理论预测的那样密切相关。本研究发现隔离度对滨海特有植物物种丰富度的重要性较低, 距最近大岛距离仅对乔木物种丰富度产生了正向影响, 距大陆距离仅对灌木物种丰富度产生了微弱的作用(图5), 这可能是由于本研究中海岛与距大陆、大岛之间的距离相对较短(距离大陆和最近大岛的最远距离分别为26.50 km和40.97 km), 这种较小的隔离度不足以限制大多数滨海特有物种的扩散, 因此隔离度并不是限制滨海特有植物分布的主要障碍。对千岛湖生境岛屿的研究也发现, 隔离不影响千岛湖植物的物种丰富度(Hu et al, 2011)。
不同生活型的滨海特有植物对气候的响应存在差异。具体而言, 本研究中风速对灌木以及草本物种丰富度具有正向影响, 但乔木物种丰富度却对风速的响应不明显。这可能是由于海岛以强风、高日晒、高蒸发率等恶劣的生境条件为主(Murcia, 1995), 草本和灌木由于低矮的植株高度和较小的植株密度, 其适应强风环境的能力高于乔木, 因而可以占据更多的生态位。其次, 风速对不同生活型植物的扩散方式也产生不同的影响, 草本的种子以风力传播为主, 而乔木则主要依靠动物扩散种子, 其扩散范围受到动物数量和分布的限制。因此, 相比乔木, 具有更强扩散能力和更高迁移率的草本植物(Tamme et al, 2014)更有利于在风速较高的海岛进行繁殖。
相比于景观因子, 气候和人为干扰因子对滨海特有植物及不同生活型植物物种丰富度的解释度低。一方面可能与本研究使用的气候因子(年均风速、年均温、年降水)和人为干扰因子(建筑与农田面积占比)有关, 它们只描述了影响特有植物分布的气候、干扰的某些方面, 可能忽视了环境因子的季节性或者年际变化(Parepa et al, 2013)。同时, 对于具有复杂地形和生境异质性丰富的海岛而言, 这些环境因子可能在小范围内发生剧烈变化, 这是本研究环境因子数据的不足之处。因此后续研究中可引入更加具体细化的气候、人为干扰、土壤理化性质等因素, 或将有助于更全面深入地理解福建海岛乃至亚热带海岛滨海特有植物物种丰富度的分布格局。
本研究探讨了景观、气候、人为干扰3个方面10个环境因子对福建海岛滨海特有植物的驱动作用, 促进了对福建海岛滨海特有植物及不同生活型植物物种丰富度分布格局的理解。研究结果表明, 福建海岛滨海特有植物及不同生活型植物存在着明显的种-面积关系, 符合岛屿生物地理学理论。滨海特有植物及不同生活型植物的物种丰富度受景观、气候和人为干扰等环境因子的影响程度不同, 景观是影响滨海特有植物物种丰富度的主要方面。具体而言, 面积、形状指数和周长面积比是福建海岛滨海特有植物和不同生活型植物物种丰富度的3个最重要的决定因素, 这些环境因子都与生境异质性密切相关。此外, 不同生活型植物对环境因子的响应存在差异, 除景观因素外, 气候方面的环境因子对也乔木产生了一定影响。探明海岛环境因子对滨海特有植物及不同生活型植物物种丰富度的影响对于后续开展海岛滨海特有植物多样性保护及合理科学利用滨海特有植物资源具有重要意义。例如面积大、形状复杂、周长面积比小、生境异质性高的海岛可以作为今后选定滨海特有植物保护区的参考依据。
附录 Supplementary Material
附录1 本研究选取的福建53个海岛概况
Appendix 1 An overview of 53 islands in Fujian selected for this study
参考文献
Time, area and isolation: Factors driving the diversification of Azorean arthropods
DOI:10.1111/jbi.2008.36.issue-1 URL [本文引用: 1]
Habitat fragmentation and species loss across three interacting trophic levels: Effects of life-history and food-web traits
DOI:10.1111/j.1523-1739.2009.01214.x
PMID:19765035
[本文引用: 1]

Not all species are likely to be equally affected by habitat fragmentation; thus, we evaluated the effects of size of forest remnants on trophically linked communities of plants, leaf-mining insects, and their parasitoids. We explored the possibility of differential vulnerability to habitat area reduction in relation to species-specific and food-web traits by comparing species-area regression slopes. Moreover, we searched for a synergistic effect of these traits and of trophic level. We collected mined leaves and recorded plant, leaf miner, and parasitoid species interactions in five 100-m2 transects in 19 Chaco Serrano woodland remnants in central Argentina. Species were classified into extreme categories according to body size, natural abundance, trophic breadth, and trophic level. Species-area slopes differed between groups with extreme values of natural abundance or trophic specialization. Nevertheless, synergistic effects of life-history and food-web traits were only found for trophic level and trophic breadth: area-related species loss was highest for specialist parasitoids. It has been suggested that species position within interaction webs could determine their vulnerability to extinction. Our results provide evidence that food-web parameters, such as trophic level and trophic breadth, affect species sensitivity to habitat fragmentation.
The role of habitat diversity in generating the small-island effect
DOI:10.1111/ecog.2020.v43.i8 URL [本文引用: 1]
A study on the flora of the islands of Zhejiang Province
浙江海岛植物区系的研究
Biogeographical determinants of pteridophytes and spermatophytes on oceanic archipelagos
DOI:10.1080/14772000.2011.603381 URL [本文引用: 1]
Confounding factors in the detection of species responses to habitat fragmentation
DOI:10.1017/S1464793105006949
PMID:16318651
[本文引用: 1]
Habitat loss has pervasive and disruptive impacts on biodiversity in habitat remnants. The magnitude of the ecological impacts of habitat loss can be exacerbated by the spatial arrangement -- or fragmentation -- of remaining habitat. Fragmentation per se is a landscape-level phenomenon in which species that survive in habitat remnants are confronted with a modified environment of reduced area, increased isolation and novel ecological boundaries. The implications of this for individual organisms are many and varied, because species with differing life history strategies are differentially affected by habitat fragmentation. Here, we review the extensive literature on species responses to habitat fragmentation, and detail the numerous ways in which confounding factors have either masked the detection, or prevented the manifestation, of predicted fragmentation effects. Large numbers of empirical studies continue to document changes in species richness with decreasing habitat area, with positive, negative and no relationships regularly reported. The debate surrounding such widely contrasting results is beginning to be resolved by findings that the expected positive species-area relationship can be masked by matrix-derived spatial subsidies of resources to fragment-dwelling species and by the invasion of matrix-dwelling species into habitat edges. Significant advances have been made recently in our understanding of how species interactions are altered at habitat edges as a result of these changes. Interestingly, changes in biotic and abiotic parameters at edges also make ecological processes more variable than in habitat interiors. Individuals are more likely to encounter habitat edges in fragments with convoluted shapes, leading to increased turnover and variability in population size than in fragments that are compact in shape. Habitat isolation in both space and time disrupts species distribution patterns, with consequent effects on metapopulation dynamics and the genetic structure of fragment-dwelling populations. Again, the matrix habitat is a strong determinant of fragmentation effects within remnants because of its role in regulating dispersal and dispersal-related mortality, the provision of spatial subsidies and the potential mediation of edge-related microclimatic gradients. We show that confounding factors can mask many fragmentation effects. For instance, there are multiple ways in which species traits like trophic level, dispersal ability and degree of habitat specialisation influence species-level responses. The temporal scale of investigation may have a strong influence on the results of a study, with short-term crowding effects eventually giving way to long-term extinction debts. Moreover, many fragmentation effects like changes in genetic, morphological or behavioural traits of species require time to appear. By contrast, synergistic interactions of fragmentation with climate change, human-altered disturbance regimes, species interactions and other drivers of population decline may magnify the impacts of fragmentation. To conclude, we emphasise that anthropogenic fragmentation is a recent phenomenon in evolutionary time and suggest that the final, long-term impacts of habitat fragmentation may not yet have shown themselves.
Rethinking patch size and isolation effects: The habitat amount hypothesis
DOI:10.1111/jbi.12130 URL [本文引用: 1]
Methods and protocols for plant community inventory
DOI:10.3724/SP.J.1003.2009.09253
[本文引用: 1]
A plant community is an assemblage of plant populations that live in certain area, and interact with and adapt to one another in the context of long-term environmental changes. Plant communities maintain global ecosystem functions, and provide food and habitats for animals and other organisms. Plant communities also provide primary resources for human survival and development, and are therefore indispensable to human societies. China is among the countries with the most diverse plant communities in the world. However, no systematic national inventory has been conducted for Chinese plant communities. This fact obstructs exploitation and protection of China’s plant resources, and also hampers the development of the fields of Chinese ecology and geography. There is an urgent need to survey Chinese plant communities using consis-tent methods and protocols. In this paper, we review major concepts in plant community ecology, and pro-pose a framework for developing plant community inventories based on recent progress in community ecol-ogy and our own experience with long-term field surveys. Our framework provides protocols for site selec-tion and plot design, items to be measured in a plot, and measurements of functional traits of dominant spe-cies. We also review protocols for field surveys of large, long-term plots. The protocols proposed in this pa-per are expected to be a base for standardizing methodology for inventory of Chinese plant communities.
植物群落清查的主要内容、方法和技术规范
DOI:10.3724/SP.J.1003.2009.09253
[本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合。它提供着人类赖以生存的主要物质资源, 维系着地球生态系统的健康和功能, 也为各种动物和其他生物提供食物来源和栖息地, 是人类生存和发展不可或缺的物质基础, 具有不可替代的作用。我国植物群落类型多样, 在世界上首屈一指, 但我国至今尚没有一次全面和系统的植物群落清查, 不仅影响了人们对我国植物资源的了解、利用和保护, 也不利于我国生态学、环境科学和地理学等相关学科的发展。采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行, 并具有紧迫性。本文基于作者长期的野外工作实践和国内外的群落调查方法, 首先简要定义了与植物群落清查有关的重要概念, 在此基础上, 论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范, 并介绍了大样地调查的主要步骤。通过本文的介绍、归纳和总结, 试图为制定我国植物群落清查的技术规范提供基础材料和技术储备。
Plant genetic diversity in the Canary Islands: A conservation perspective
The Canary Islands are an Atlantic volcanic archipelago with a rich flora of ∼570 endemic species. The endemics represent ∼40% of the native flora of the islands, and ∼20% of the endemics are in the E (endangered) category of the International Union for Conservation of Nature. A review of allozyme variation in 69 endemic species belonging to 18 genera and eight families is presented. The average species-level genetic diversity (H(T)) at allozyme loci is 0.186, which is twice as high as the mean reported for endemics of Pacific archipelagos. Possible factors contributing to this higher diversity are discussed, but the reasons remain obscure. An average of 28% of the allozyme diversity within species resides among populations, indicating a high level of interpopulational differentiation. Studies of reproductive biology indicate that many of the endemic species are outcrossers. The high total diversity within species, the relatively high differentiation among populations, and the outcrossing breeding systems have implications for species conservation. Decreased population sizes in outcrossing species would promote biparental inbreeding and increase inbreeding depression. The relatively high proportion of allozyme diversity among populations indicates that the most effective strategy for preserving genetic variation in species is to conserve as many populations as possible. The genetic diversity in many Canary Island endemics is endangered by: (1) overgrazing by introduced animals, such as barbary sheep, goats, mouflons, rabbits, and sheep; (2) interspecific hybridization following habitat disturbance or planting of endemics along roadsides or in public gardens; (3) competition with alien plant species; and (4) decline of population size because of urban development and farming.
Factors affecting the number of species of plants on islands of the Great Barrier Reef, Australia
DOI:10.2307/2845294 URL [本文引用: 1]
Ecological and evolutionary consequences of biological invasion and habitat fragmentation
DOI:10.1007/s10021-003-0138-8 URL [本文引用: 1]
Satellite based land use and landscape complexity indices as predictors for regional plant species diversity
DOI:10.1016/S0169-2046(02)00194-9 URL [本文引用: 1]
Determinants of plant species richness and patterns of nestedness in fragmented landscapes: Evidence from land-bridge islands
DOI:10.1007/s10980-011-9662-7 URL [本文引用: 3]
Climate controls plant life-form patterns on a high-elevation oceanic island
DOI:10.1111/jbi.v47.10 URL [本文引用: 1]
Human impact, climate and dispersal strategies determine plant invasion on islands
DOI:10.1111/jbi.v48.8 URL [本文引用: 1]
A global model of island biogeography
DOI:10.1111/geb.2006.15.issue-1 URL [本文引用: 2]
Plant species richness—The effect of island size and habitat diversity
DOI:10.2307/2261304 URL [本文引用: 1]
Global diversity of island floras from a macroecological perspective
DOI:10.1111/j.1461-0248.2007.01129.x
PMID:18036182
[本文引用: 1]
Islands harbour a significant portion of all plant species worldwide. Their biota are often characterized by narrow distributions and are particularly susceptible to biological invasions and climate change. To date, the global richness pattern of islands is only poorly documented and factors causing differences in species numbers remain controversial. Here, we present the first global analysis of 488 island and 970 mainland floras. We test the relationship between island characteristics (area, isolation, topography, climate and geology) and species richness using traditional and spatial models. Area is the strongest determinant of island species numbers (R(2) = 0.66) but a weaker predictor for mainlands (R(2) = 0.25). Multivariate analyses reveal that all investigated variables significantly contribute to insular species richness with area being the strongest followed by isolation, temperature and precipitation with about equally strong effects. Elevation and island geology show relatively weak yet significant effects. Together these variables account for 85% of the global variation in species richness.
Role of climate and geohistorical factors in driving plant richness patterns and endemicity on the east Asian continental islands
DOI:10.1111/ecog.2015.v38.i6 URL [本文引用: 2]
High beta diversity among small islands is due to environmental heterogeneity rather than ecological drift
DOI:10.1111/jbi.2018.45.issue-10 URL [本文引用: 1]
A species-based theory of insular zoogeography
DOI:10.1046/j.1365-2699.2000.00188.x URL [本文引用: 1]
An equilibrium theory of insular zoogeography
DOI:10.2307/2407089 URL [本文引用: 1]
Similar yet distinct distributional patterns characterize native and exotic plant species richness across northern New Zealand islands
DOI:10.1111/jbi.v48.7 URL [本文引用: 1]
Edge effects in fragmented forests: Implications for conservation
DOI:10.1016/S0169-5347(00)88977-6 URL [本文引用: 1]
Disturbance and diversity in a continental archipelago: A mechanistic framework linking area, height, and exposure
DOI:10.1002/ecs2.1957 URL [本文引用: 1]
Patterns of species richness on very small islands: The plants of the Aegean archipelago
DOI:10.1111/jbi.2006.33.issue-7 URL [本文引用: 1]
Environmental variability promotes plant invasion
DOI:10.1038/ncomms2632
PMID:23511469
[本文引用: 1]
Global environmental change not only entails changes in mean environmental conditions but also in their variability. Changes in climate variability are often associated with altered disturbance regimes and temporal patterns of resource availability. Here we show that increased variability of soil nutrients strongly promotes another key process of global change, plant invasion. In experimental plant communities, the success of one of the world's most invasive plants, Japanese knotweed, is two-to four-fold increased if extra nutrients are not supplied uniformly, but in a single large pulse, or in multiple pulses of different magnitudes. The superior ability to take advantage of variable environments may be a key mechanism of knotweed dominance, and possibly many other plant invaders. Our study demonstrates that increased nutrient variability can promote plant invasion, and that changes in environmental variability may interact with other global change processes and thereby substantially accelerate ecological change.
Island biogeography of marine organisms
DOI:10.1038/nature23680 URL [本文引用: 1]
Community diversity: Relative roles of local and regional processes
The species richness (diversity) of local plant and animal assemblages-biological communities-balances regional processes of species formation and geographic dispersal, which add species to communities, against processes of predation, competitive exclusion, adaptation, and stochastic variation, which may promote local extinction. During the past three decades, ecologists have sought to explain differences in local diversity by the influence of the physical environment on local interactions among species, interactions that are generally believed to limit the number of coexisting species. But diversity of the biological community often fails to converge under similar physical conditions, and local diversity bears a demonstrable dependence upon regional diversity. These observations suggest that regional and historical processes, as well as unique events and circumstances, profoundly influence local community structure. Ecologists must broaden their concepts of community processes and incorporate data from systematics, biogeography, and paleontology into analyses of ecological patterns and tests of community theory.
Island biogeography of native and alien plant species: Contrasting drivers of diversity across the Lesser Antilles
DOI:10.1111/ddi.v26.11 URL [本文引用: 2]
Are species-area relationships from entire archipelagos congruent with those of their constituent islands?
Species-area relationships on small islands differ among plant growth forms
DOI:10.1111/geb.v29.5 URL [本文引用: 2]
Habitat diversity, ecological requirements of species and the small island effect
DOI:10.1111/ddi.2009.15.issue-1 URL [本文引用: 1]
Flora of the Taizhou islands
台州列岛植物区系的研究
Plant invasion and speciation along elevational gradients on the oceanic island La Palma, Canary Islands
DOI:10.1002/ece3.2640
PMID:28116071
[本文引用: 1]
Ecosystems that provide environmental opportunities but are poor in species and functional richness generally support speciation as well as invasion processes. These processes are expected not to be equally effective along elevational gradients due to specific ecological, spatial, and anthropogenic filters, thus controlling the dispersal and establishment of species. Here, we investigate speciation and invasion processes along elevational gradients. We assess the vascular plant species richness as well as the number and percentage of endemic species and non-native species systematically along three elevational gradients covering large parts of the climatic range of La Palma, Canary Islands. Species richness was negatively correlated with elevation, while the percentage of Canary endemic species showed a positive relationship. However, the percentage of Canary-Madeira endemics did not show a relationship with elevation. Non-native species richness (indicating invasion) peaked at 500 m elevation and showed a consistent decline until about 1,200 m elevation. Above that limit, no non-native species were present in the studied elevational gradients. Ecological, anthropogenic, and spatial filters control richness, diversification, and invasion with elevation. With increase in elevation, richness decreases due to species-area relationships. Ecological limitations of native ruderal species related to anthropogenic pressure are in line with the absence of non-native species from high elevations indicating directional ecological filtering. Increase in ecological isolation with elevation drives diversification and thus increased percentages of Canary endemics. The best preserved eastern transect, including mature laurel forests, is an exception. The high percentage of Canary-Madeira endemics indicates the cloud forest's environmental uniqueness-and thus ecological isolation-beyond the Macaronesian islands.
Predicting species’ maximum dispersal distances from simple plant traits
Many studies have shown plant species' dispersal distances to be strongly related to life-history traits, but how well different traits can predict dispersal distances is not yet known. We used cross-validation techniques and a global data set (576 plant species) to measure the predictive power of simple plant traits to estimate species' maximum dispersal distances. Including dispersal syndrome (wind, animal, ant, ballistic, and no special syndrome), growth form (tree, shrub, herb), seed mass, seed release height, and terminal velocity in different combinations as explanatory variables we constructed models to explain variation in measured maximum dispersal distances and evaluated their power to predict maximum dispersal distances. Predictions are more accurate, but also limited to a particular set of species, if data on more specific traits, such as terminal velocity, are available. The best model (R2 = 0.60) included dispersal syndrome, growth form, and terminal velocity as fixed effects. Reasonable predictions of maximum dispersal distance (R2 = 0.53) are also possible when using only the simplest and most commonly measured traits; dispersal syndrome and growth form together with species taxonomy data. We provide a function (dispeRsal) to be run in the software package R. This enables researchers to estimate maximum dispersal distances with confidence intervals for plant species using measured traits as predictors. Easily obtainable trait data, such as dispersal syndrome (inferred from seed morphology) and growth form, enable predictions to be made for a large number of species.
A model for the species-area-habitat relationship
DOI:10.1046/j.1365-2699.2003.00805.x URL [本文引用: 1]
Determinants of alpha and beta vascular plant diversity in Mediterranean island systems: The Ionian islands, Greece
DOI:10.1111/njb.2019.v37.i1 URL [本文引用: 1]
Factors affecting species richness and beta diversity of vascular plants on small islands in the Wenzhou region of eastern China
温州沿海小型海岛植物丰富度和β多样性及其影响因子
Bryophyte diversity on a tropical continental island (Hainan, China): Potential vulnerable species and environmental indicators
DOI:10.1080/03736687.2019.1653557 URL [本文引用: 1]
A general dynamic theory of oceanic island biogeography
DOI:10.1111/j.1365-2699.2008.01892.x URL [本文引用: 1]
Effects of changing scale on landscape pattern analysis: Scaling relations
DOI:10.1023/B:LAND.0000021711.40074.ae URL [本文引用: 1]
Determinants of bryophyte species richness on the Zhoushan Archipelago, China
DOI:10.1016/j.baae.2019.05.002 URL [本文引用: 1]
Richness and composition of plants and birds on land-bridge islands: Effects of island attributes and differential responses of species groups
DOI:10.1111/jbi.2012.39.issue-6 URL [本文引用: 5]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}