


进入21世纪以来, 以全球变暖为代表的气候变化问题成为研究者们关注的焦点(陈嘉哲等, 2020), 而常年被冰雪覆盖、处于极端环境中的北极地区对全球气候变化响应更为敏感, 其平均气温的升高幅度是全球的两倍之多(Serreze & Francis, 2006; Holmes et al, 2012; NSIDC, 2018)。气候变暖引起的海冰覆盖面积减少和海水温度升高等问题正悄悄地改变着北极海洋食物网的结构、功能以及物种间的营养关系, 进而影响整个北极海洋生态系统的稳定性(Whitehouse et al, 2017; 陈建芳等, 2018; 王慧等, 2020)。白令海与楚科奇海是连接太平洋与北冰洋的唯一通道, 其生态系统对气候变化的响应与反馈同样十分剧烈, 是北极地区受气候变化影响的真实写照(NSIDC, 2018; 余兴光等, 2018), 也是研究气候-环境-生物相互影响、相互作用的绝佳场所。
营养级表征了生物在生态系统中的营养位置, 揭示了生态系统的营养结构组成(韩东燕等, 2016; 宁加佳等, 2016), 是食物网营养结构研究中不可分割的一部分(高世科等, 2021)。营养生态位是生态环境中生物体之间营养相互作用的体现, 由生物间的捕食和竞争关系形成(Leibold, 1995)。鱼类作为海洋食物网的重要组成部分, 是上下水层营养物质转移和能量流通的重要枢纽, 也是中上层群落与底栖群落连接的重要节点, 具有复杂的捕食-被捕食以及竞争关系。充分了解和掌握北极鱼类在食物网中的营养位置和营养关系, 有利于揭示北极海洋生态系统或群落的营养格局和结构组成特征, 进而对于解析北极海洋生态系统对气候变化的响应具有重要意义。
本研究运用稳定同位素技术分析了北极地区白令海与楚科奇海海域主要鱼类的碳氮稳定同位素特征, 计算其在食物网中所处的营养位置, 比较了种间营养生态位的差异, 以期科学评估该海域主要鱼类在食物网中所处的营养位置以及种间营养关系, 研究结果可为今后北极海洋生态系统研究提供营养动力学基础。
1 材料与方法
1.1 样品采集与鉴定
鱼类调查数据和样品来源于我国第6次(2014年)和第8-11次(2017-2020年)北极科学考察, 调查海域为白令海与楚科奇海(57°-79° N, -155°至-179° E), 共布设了56个调查站点(图1)。北极鱼类调查采样网具为三角底拖网(宽2.2 m, 高0.65 m, 长6.5 m, 囊网网目20 mm), 拖网船速约为3 kn, 拖网时间为10-33 min。样品捕获后立即在-20℃冷冻保存, 带回实验室进行鉴定和测量。
图1
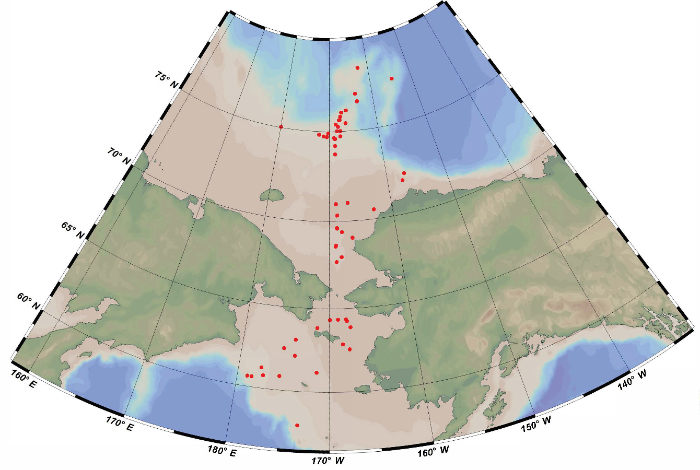
图1
北极科学考察鱼类调查站位图
Fig. 1
Surveyed stations of fishes in the Chinese Arctic Research Expedition
鱼类鉴定参考Fishes of Alaska (Mecklenburg et al, 2002)、Pacific Arctic Marine Fishes (Mecklenburg et al, 2016), 系统分类与命名主要依据Fishes of the World, 5th edn. (Nelson et al, 2016) 和《拉汉世界鱼类系统名典》(伍汉霖等, 2017)。共捕获鱼类8科21属29种(表1), 测定碳、氮稳定同位素样品401个, 其中鳕科共4种129尾, 鲽科共3种64尾, 杜父鱼科共9种118尾, 绵鳚科共8种51尾, 线鳚科共2种28尾, 八角鱼科共1种3尾, 狮子鱼科共1种5尾, 玉筋鱼科共1种3尾。
表1 白令海与楚科奇海鱼类营养级
Table 1
| 科 Family | 种类 Species | 样品数 Number of samples | 营养级X (最小值-最大值) Trophic level X (Min-Max) | Fishbase营养级 Trophic level of Fishbase |
|---|---|---|---|---|
| 鳕科 Gadidae | 白令海北鳕 Boreogadus saida | 80 | 3.76 (3.56-3.97) | 2.9-4.2 |
| 北极鳕 Arctogadus glacialis | 17 | 4.31 (4.08-4.58) | 3.2 | |
| 黄线狭鳕 Gadus chalcogrammus | 29 | 3.11 (2.91-3.31) | 3.0-4.5 | |
| 细身宽突鳕 Eleginus gracilis | 3 | 3.45 (2.71-4.20) | 3.0-4.1 | |
| 鲽科 Pleuronectidae | 糙黄盖鲽 Limanda aspera | 2 | 3.96 (2.00-4.02) | 3.1-3.7 |
| 黄腹鲽 Pleuronectes quadrituberculatus | 2 | 3.67 (2.00-3.76) | 3.1-3.9 | |
| 太平洋拟庸鲽 Hippoglossoides elassodon | 60 | 3.57 (3.37-3.79) | 3.4-3.8 | |
| 杜父鱼科 Cottidae | 粗糙钩杜父鱼 Artediellus scaber | 2 | 3.98 (2.01-4.03) | 3.4 |
| 大西洋钩杜父鱼 Artediellus atlanticus | 23 | 4.59 (4.37-4.86) | 3.6 | |
| 短角床杜父鱼 Myoxocephalus scorpius | 9 | 3.40 (3.07-3.78) | 2.1-4.4 | |
| 横带杂鳞杜父鱼 Hemilepidotus papilio | 3 | 3.05 (2.01-4.64) | 3.3 | |
| 锯棘冰杜父鱼 Icelus spiniger | 3 | 3.16 (2.00-4.28) | 3.6 | |
| 尼氏鮄杜父鱼 Triglops nybelini | 4 | 3.84 (3.17-4.57) | 3.3 | |
| 强棘杜父鱼 Enophrys diceraus | 2 | 4.06 (2.01-4.09) | 3.8 | |
| 三叉裸棘杜父鱼 Gymnocanthus tricuspis | 62 | 3.43 (3.25-3.62) | 3.2-3.3 | |
| 匙形冰杜父鱼 Icelus spatula | 10 | 3.68 (3.42-3.93) | 3.9 | |
| 绵鳚科 Zoarcidae | 阿氏狼绵鳚 Lycodes adolfi | 8 | 4.26 (3.73-4.86) | 3.3 |
| 半花裸鳚 Gymnelus hemifasciatus | 4 | 3.33 (2.09-4.58) | 3.1 | |
| 半裸狼绵鳚 Lycodes seminudus | 6 | 4.53 (4.14-4.97) | 3.2-4.0 | |
| 北极狼绵鳚 Lycodes polaris | 2 | 4.29 (2.00-4.35) | 3.4 | |
| 苍色狼绵鳚 Lycodes pallidus | 10 | 4.44 (4.18-4.71) | 3.3-4.1 | |
| 短鳍狼绵鳚 Lycodes brevipes | 12 | 3.44 (3.19-3.68) | 3.5-4.2 | |
| 砂栖狼绵鳚 Lycodes palearis | 6 | 3.71 (3.34-4.09) | 3.5 | |
| 网纹狼绵鳚 Lycodes reticulatus | 3 | 4.62 (4.07-5.17) | 3.5 | |
| 线鳚科 Stichaeidae | 斑鳍北鳚 Lumpenus fabricii | 9 | 3.40 (3.09-3.72) | 3.3 |
| 中间弧线鳚 Anisarchus medius | 19 | 3.85 (3.63-4.11) | 3.2 | |
| 八角鱼科 Agonidae | 北极胶八角鱼 Aspidophoroides olrikii | 3 | 3.01 (2.01-3.85) | 2.1-3.9 |
| 狮子鱼科 Liparidae | 费氏狮子鱼 Liparis fabricii | 5 | 4.17 (3.56-4.75) | 3.3 |
| 玉筋鱼科 Ammodytidae | 六斑玉筋鱼 Ammodytes hexapterus | 3 | 2.42 (2.28-2.60) | 2.7-3.3 |
X代指营养级的值, 取95%置信区间。
X stands for trophic level value, with 95% confidence interval.
1.2 稳定同位素的测定
将获取的401个鱼类样品解冻后取背部侧线上方肌肉, 取样量以冻干后能顺利研磨为标准。以上所有样品置于-50℃冻干机中冻干至恒重, 经球磨仪研磨后用60目筛绢过滤以保证样品的均匀性, 保存于干燥箱中待测。所有样品在自然资源部第三海洋研究所信息测试与保障中心进行检测, 使用德国Elementar公司的稳定同位素质谱仪联机(Vario ISOPOTE Cube-Isoprime)测定碳、氮稳定同位素比率。稳定同位素的自然丰度表示为(Peterson & Fry, 1987):
式中, X为13C或15N, R表示13C/12C或15N/14N的比值; 碳、氮稳定同位素测定的标准物质分别为美洲拟箭石和大气中的氮气。样品δ13C和δ15N的分析精度分别为0.2‰和0.3‰。
1.3 营养级的计算
营养级可用来描述物种在食物网中所处的营养水平以及所发挥的生态作用, 对分析食物网结构和揭示生态系统的营养格局具有重要意义(Post, 2002)。本文以Quezada-Romegialli等(2018)提出的基于贝叶斯概率统计的贝叶斯模型(tRophicPosition)来计算鱼类营养级, 通过tRophicPosition包在R 4.1.2中实现。贝叶斯模型将马尔可夫链蒙特卡洛模拟(Markov Chain Monte Carlo, MCMC)计算方法与R软件中统计和图形分析相结合, 同时可包括1-2个基线, 其明确地考虑了个体差异性和抽样误差的传递效应(如同位素富集度、基线生物和消费者的测量值) (Quezada-Romegialli et al, 2018)。该模型对自适应阶段进行了40,000次迭代, 前20,000次丢弃(从后验估计开始时丢弃), 后20,000次保留, 通过JAGS 4.3.0吉布斯取样器, 使用了5个平行的马尔可夫链蒙特卡洛(MCMC)模拟。
1.4 营养生态位宽幅和重叠
基于R语言构图需要和保证研究结果的准确性, 选取本研究中样本数量大于5的16种鱼类(表2)作为研究对象进行营养生态位作图分析。根据测得的16种鱼类样品的稳定同位素比值, 运用R软件中的SIBER包构建δ13C-δ15N稳定同位素营养生态位, 参考Layman等(2007)提出的一套度量指标来定量分析其中的坐标点信息。Layman营养多样性度量指标有4项, 包括: (1) δ15N范围(δ15N range, NR): 所有样品中δ15N的最大值与最小值之间的差值大小, 可量化食物网的营养长度信息; (2) δ13C范围(δ13C range, CR): 所有样品中δ13C的最大值与最小值之间的差值大小, 可表征物种的食源多样性; (3)不规则图形面积(total area of the convex hull, TA): 由物种所有坐标点形成的凸多边形面积, 可指示物种所占据的营养生态位空间, Jackson等(2011)引入稳定同位素贝叶斯椭圆模型(stable isotope Bayesian ellipses in R, SIBER), 通过计算标准椭圆面积(standard ellipse area, SEA)代替TA作为同位素生态位空间边界, SEAc (corrected standard ellipse area)是对SEA的一种校正, 目前该指标已被广泛用作估算物种的营养生态位重叠(Parnell et al, 2013); (4)质心的平均距离(mean distance to centroid, CD): 物种的每一个坐标点到凸边形质心的平均欧氏距离, 可代表物种的平均营养多样性信息。Layman群落营养冗余度量指标有2项, 包括: (1)最短均值(mean nearest neighbour distance, MNND): 每个物种坐标点与其最近的相邻物种坐标点的平均最邻近距离, 可衡量物种在群落内堆积的密度和聚集性, 数值越小意味着营养冗余越大; (2)最短标准差(standard deviation of nearest neighbour distance, SDNND): 每个物种与其最近相邻物种的欧式距离标准偏差, 可度量物种聚集的均匀性, 较低的SDNND意味着物种分布会更均匀。
表2 白令海与楚科奇海鱼类的营养生态位参数值
Table 2
| 种名 Species | 不规则 图形面 积(TA) | 标准椭 圆面积(SEA) | 校正后 的标准 椭圆面积(SEAc) |
|---|---|---|---|
| 白令海北鳕 Boreogadus saida | 23.95 | 5.44 | 5.51 |
| 黄线狭鳕 Gadus chalcogrammus | 5.86 | 1.73 | 1.80 |
| 北极鳕 Arctogadus glacialis | 2.41 | 0.84 | 0.90 |
| 费氏狮子鱼 Liparis fabricii | 3.28 | 3.10 | 4.13 |
| 三叉裸棘杜父鱼 Gymnocanthus tricuspis | 11.62 | 2.86 | 2.91 |
| 短角床杜父鱼 Myoxocephalus scorpius | 1.35 | 0.81 | 0.92 |
| 大西洋钩杜父鱼 Artediellus atlanticus | 2.44 | 0.81 | 0.85 |
| 匙形冰杜父鱼 Icelus spatula | 1.78 | 0.91 | 1.02 |
| 斑鳍北鳚 Lumpenus fabricii | 4.96 | 3.15 | 3.61 |
| 中间弧线鳚 Anisarchus medius | 3.91 | 1.64 | 1.73 |
| 太平洋拟庸鲽 Hippoglossoides elassodon | 12.34 | 3.30 | 3.36 |
| 短鳍狼绵鳚 Lycodes brevipes | 1.77 | 0.79 | 0.87 |
| 苍色狼绵鳚 Lycodes pallidus | 2.28 | 1.13 | 1.27 |
| 阿氏狼绵鳚 Lycodes adolfi | 6.46 | 4.05 | 4.72 |
| 砂栖狼绵鳚 Lycodes palearis | 1.46 | 1.02 | 1.29 |
| 半裸狼绵鳚 Lycodes seminudus | 5.02 | 4.03 | 5.04 |
TA, Total area of the convex hull; SEA, Standard ellipse area; SEAc, Corrected standard ellipse area.
根据Swanson等(2015)对物种间生态位重叠的概率计算, 本研究中定义O(A/B)代表物种A和物种B的生态位重叠比例, 即物种A同位素生态位(OA)和物种B同位素生态位(OB)的贝叶斯椭圆相互重叠面积(OO)占物种A和物种B减去重叠部分后的贝叶斯椭圆面积的百分比:
2 结果
2.1 碳、氮稳定同位素比值特征和营养级
本研究全部鱼类样品δ13C范围为-23.88‰至-15.78‰, 总跨度为8.10‰, 均值为-19.36‰; δ15N的范围为11.80‰-20.37‰, 总跨度为8.57‰, 均值为16.16‰。δ13C均值最大的为短鳍狼绵鳚(Lycodes brevipes, -17.13‰), 最小的为尼氏鮄杜父鱼(Triglops nybelini, -21.89‰); δ15N均值最大的为网纹狼绵鳚(Lycodes reticulatus, 19.26‰), 最小的为六斑玉筋鱼(Ammodytes hexapterus, 11.84‰)。
如表1所示, 所有鱼类样品总营养级范围为2.42-4.62, 均值为3.74, 其中六斑玉筋鱼的营养级最低, 小于3.00; 网纹狼绵鳚、大西洋钩杜父鱼和半裸狼绵鳚(Lycodes seminudus)的营养级均大于4.50。位于2.0-3.0的鱼类只有六斑玉筋鱼1种, 占总种类数的3.44%; 位于3.0-4.0营养级的有19种, 占总种类数的65.51%; 位于4.0-4.5营养级的有6种, 占总种类数的20.69%; 营养级大于4.5的有3种, 占总种类数的13.79%。
2.2 白令海与楚科奇海鱼类营养生态位宽幅与重叠
白令海与楚科奇海16种鱼类在食物网中所占据的营养生态位宽幅大小情况如表2所示: 白令海北鳕、费氏狮子鱼(Liparis fabricii)、阿氏狼绵鳚(Lycodes adolfi)和半裸狼绵鳚的营养生态位宽幅相对较大, 对应的SEAc大于4.00, 其中白令海北鳕的营养生态位宽幅最大(TA = 23.95, SEA = 5.44, SEAc = 5.51, CR = 5.94‰, NR = 6.29‰); 北极鳕(Arctogadus glacialis)、短角床杜父鱼(Myoxocephalus scorpius)、短鳍狼绵鳚、大西洋钩杜父鱼的营养生态位相对较小, 对应的SEAc小于1.00, 其中大西洋钩杜父鱼的营养生态位宽幅最小(TA = 2.44, SEA = 0.81, SEAc = 0.85, CR = 3.42‰, NR = 4.70‰)。
表3和图2显示了16种鱼类在食物网中所占据的营养生态位重叠情况以及重叠百分比, 基本每种鱼类的标准椭圆都与其他几种鱼类存在一定程度的重叠, 其中大西洋钩杜父鱼、短鳍狼绵鳚的标准椭圆分别完全位于半裸狼绵鳚、三叉裸棘杜父鱼(Gymnocanthus tricuspis)之内, 除此之外其他多数物种间并未完全重叠, 保持一定的独特性; 线鳚科(斑鳍北鳚Lumpenus fabricii、中间弧线鳚Anisarchus medius)与其他鱼类种间重叠严重, 其中斑鳍北鳚与太平洋拟庸鲽(Hippoglossoides elassodon)之间营养生态位重叠百分比的值最大, 几乎达到50%; 白令海北鳕的SEAc虽然最大(表2), 但是与其他鱼类重叠程度较低。
表3 白令海与楚科奇海鱼类的营养生态位重叠百分比。A: 白令海北鳕; B: 黄线狭鳕; C: 北极鳕; D: 费氏狮子鱼; E: 三叉裸棘杜父鱼; F: 短角床杜父鱼; G: 大西洋钩杜父鱼; H: 匙形冰杜父鱼; I: 斑鳍北鳚; J: 中间弧线鳚; K: 太平洋拟庸鲽; L: 短鳍狼绵鳚; M: 苍色狼绵鳚; N: 阿氏狼绵鳚; O: 砂栖狼绵鳚; P: 半裸狼绵鳚。
Table 3
| 物种 Species | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 1.21 | 1.67 | 19.51 | - | - | - | - | - | - | - | - | - | - | - | - |
| B | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| C | 21.35 | - | - | - | - | - | - | - | - | - | - | - | - | ||
| D | - | - | - | - | - | - | - | - | 12.01 | 9.49 | - | 5.31 | |||
| E | 8.14 | - | 23.95 | 24.77 | 6.31 | 19.31 | 29.90 | - | - | 11.79 | - | ||||
| F | - | - | 23.30 | 9.31 | 21.66 | - | - | - | 13.84 | - | |||||
| G | - | - | - | - | - | 25.46 | - | - | 16.88 | ||||||
| H | 7.96 | 7.41 | 3.58 | 8.73 | - | - | 23.38 | - | |||||||
| I | 7.93 | 48.98 | 1.43 | - | - | 13.72 | - | ||||||||
| J | 15.96 | - | - | 1.50 | 17.49 | - | |||||||||
| K | - | - | 9.76 | 13.52 | - | ||||||||||
| L | - | - | - | - | |||||||||||
| M | 24.59 | - | 20.68 | ||||||||||||
| N | - | 25.74 | |||||||||||||
| O | - |
- 没有重叠
No overlap
图2
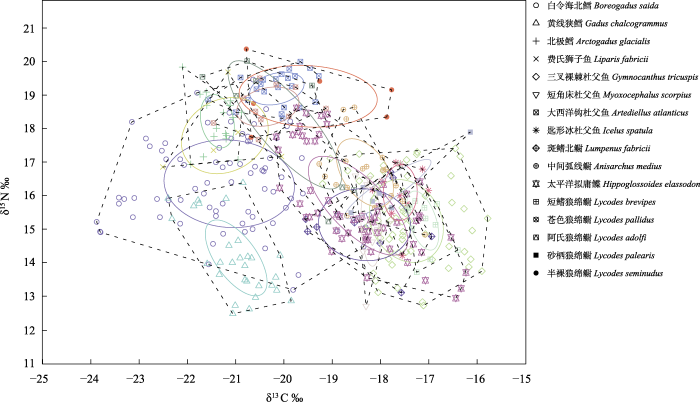
图2
白令海与楚科奇海鱼类的营养生态位宽幅及重叠。凸多边形表示物种所占据的营养生态位空间, 椭圆表示物种的核心同位素生态位空间。
Fig. 2
Trophic niche breadth and overlap of fishes from Bering Sea-Chukchi Sea. The convex polygon represents the trophic niche space occupied by species. The ellipse represents the core isotopic niche space of a species.
3 讨论
3.1 不同水域稳定同位素比值特征分析
Marsh等(2017)研究得出了16种楚科奇海鱼类的稳定同位素比值, 本研究与其相同种类达到11种(附录1); 在邻近水域波弗特海(Beaufort Sea)和阿蒙森特湾(Amundsen Gulf), Stasko等(2017)研究了28种鱼类的稳定同位素δ13C值和δ15N值(附录1), 本研究与其相同种类有13种。相比之下, Marsh等(2017)与Stasko等(2017)研究的稳定同位素δ13C、δ15N特征比本研究稍显贫化。究其原因, 一是海域范围不同, 本研究范围不仅包括楚科奇海还涵盖了白令海北部海域, 本地区营养水平较高, 营养盐充足, 利于鱼类饵料生物的生长发育(Sigler et al, 2017; Whitehouse et al, 2017), 使得本地区的消费者食物来源多元化程度高, 饮食选择更多样(Walkusz et al, 2011); 而波弗特海处于更北方的北极海域, 开冰季节短, 缺乏充足的太阳照射, 初级生产力较低(Carey, 1991; Gray et al, 2016), 进而影响了鱼类的食源多样性, 因此海域环境及饵料生物的不同影响了鱼类的稳定同位素组成(纪炜炜等, 2013); 二是种类范围略有差异, 本研究除两者相同种类以外还包括诸如强棘杜父鱼(Enophrys diceraus)、短鳍狼绵鳚等较高营养水平鱼类, 食物链更长, 15N更富集(Layman et al, 2007)。
除此之外, 当将本研究与我国近海海域鱼类稳定同位素结果相比较时可以发现, 随着纬度升高, 鱼类群落中的最负δ13C值自南向北出现贫化趋势。孔业富等(2020)研究调查了南海中西部海域22种鱼类, 其δ13C最小为-19.86‰; 纪炜炜等(2013)研究调查了东海中北部及黄海南部海域19种鱼类, 其δ13C最小为-21.37‰; 麻秋云等(2015)研究调查了胶州湾海域34种鱼类, 其δ13C最小为-22.49‰; 本研究调查了白令海与楚科奇海29种鱼类, 其δ13C最小为-23.88‰。这与蔡德陵等(2001)对9个不同生态系统中δ13C值变动趋势的研究结果一致, 最北的格陵兰东北海域δ13C值最负, 温带地区的胶州湾海域δ13C值居中, 最南的马来西亚海域δ13C值最高。这一趋势可能与不同纬度的光照水平有关, 日照水平较高的环境中初级生产者体内的碳更为富集(Grice et al, 1996)。
3.2 白令海与楚科奇海鱼类营养级分析
白令海与楚科奇海鱼类营养级范围为2.42- 4.62, 均值为3.74。如表1所示, 本研究鱼类营养级结果与Fishbase相比有11种鱼类偏高, 4种偏低, 其余种类相似。其中偏高的大多是食源多样性较高、捕食随机性较大的鱼类, 例如北极鳕、三叉裸棘杜父鱼等, 其食物组成中可能含有δ15N值较高的鱼虾以及底栖无脊椎动物(Walkusz et al, 2011; Christiansen et al, 2012; Gray et al, 2016, 2017), 最终导致营养级结果偏大。而4种偏低的鱼类营养级大小差值范围为0.22-0.44, 属于误差控制范围0.5以内(蔡德陵等, 2005)。在与Marsh等(2017)的研究可比较的11种鱼类中(附录1), 8种营养级结果高于后者, 其中白令海北鳕、粗糙钩杜父鱼(Artediellus scaber)与中间弧线鳚差值较大, 分别高出0.76、0.63、0.50, 另外5种差值较小, 范围为0.22-0.29; 3种鱼类的营养级结果低于Marsh等(2017)的研究, 差值范围为0.07-0.35。总体来看, 两项研究在同样基于稳定同位素技术的前提下, 共同种类的营养级除白令海北鳕以外相差不大, 后者可能是由于主要食物的来源不同造成的。Marsh等(2017)的研究海域主要是受白令海与楚科奇海夏季水(Bering-Chukchi summer water, BCSW)的影响(Danielson et al, 2017; Marsh et al, 2017), 而本研究中纬度超过72° N的部分海域(图1)主要受海冰融水/楚科奇海冬季水(melt water, MW/Chukchi winter water, CWW)的影响, 在以BCSW为主导的海域中, 白令海北鳕主要以浮游生物为食, 而在MW/CWW为主导的海域中则更多以底栖生物为食(Marsh et al, 2017), 且底栖无脊椎动物的δ15N值范围普遍高于浮游生物(Walkusz et al, 2011; Gray et al, 2016), 因此这可能是本研究中白令海北鳕的营养级结果偏高的原因之一。另外, 白令海北鳕是一种杂食性鱼类, 营养生态位较宽(Cui et al, 2012), 在底层海域也能以高营养级别的鱼类为食(Rand et al, 2013), 所以营养级受其所食种类的不同变化较大, 波动性较强, 与Fishbase上营养级大小范围为2.9-4.2 (表1)相符。
3.3 白令海与楚科奇海鱼类营养生态位
Layman等(2007)提出了6种衡量营养生态位的度量参数, 其中TA、CR、NR、CD反映了群落的营养多样性与食源多样性水平, MNND和SDNND则反映了群落的营养冗余程度大小。根据营养生态位分布图计算的6个稳定同位素生态位指标结果表明(表4), 本研究6个稳定同位素生态位指标皆大于Marsh等(2017)的研究。由此可得, 与Marsh等(2017)对楚科奇海鱼类群落研究的结果相比, 本研究鱼类群落营养多样性与食源多样性水平相对较高(TA、CR、NR、CD值更高), 但营养冗余程度相对较低(MNND、SDNND较高)。因为衡量营养生态位的度量参数会因为水体的不同而异(Marsh et al, 2017), 本研究调查区域除楚科奇海以外还包含白令海海域, 鱼类群落的食物来源比Marsh等(2017)研究更多样(CR: 4.25 > 3.3), 导致不规则图形面积(TA)更大(同位素生态位空间), 与质心的平均距离(CD)更大(营养多样性更高), 最短均值(MNND)更大(营养冗余度更低)。与近海海域相比(表4), 除CR外, 本研究的TA、NR、CD、MNND、SDNND等皆大于纪炜炜等(2015)的研究, 虽然CR小于纪炜炜等(2015)对东海北部鱼类群落的调查结果, 但NR高出后者更多, 表明东海北部鱼类饵料资源更丰富, 食源多样性更高, 鱼类群落中处于相同生态位的种数更多, 导致TA略小于本研究, 而营养冗余程度高于本研究(MNND与SDNND值较低)。
表4 白令海与楚科奇海鱼类群落的稳定同位素参数值对比
Table 4
| 不规则图形面积 Total area of the convex hull (TA) | δ15N范围 δ15N range (NR) | δ13C范围 δ13C range (CR) | 质心的平均距离 Mean distance to centroid (CD) | 最短均值 Mean nearest neighbour distance (MNND) | 最短标准差 Standard deviation of nearest neighbour distance (SDNND) | |
|---|---|---|---|---|---|---|
| 本研究 This study | 13.08 | 5.06 | 4.25 | 2.04 | 0.61 | 0.53 |
| 纪炜炜等, 2015 | 12.05 | 3.59 | 4.83 | 1.51 | 0.41 | 0.22 |
| Marsh et al, 2017 | 8.8 | 4.4 | 3.3 | 1.28 | 0.50 | 0.32 |
生态位可以表征生物对环境的适应能力, 而营养生态位则反映了生物在食物网中的营养位置以及生物之间的营养关系, 物种在生态系统中占据的营养生态位宽幅代表了其利用所有可利用资源状态(杨效文和马继盛, 1992), 宽幅越大, 说明其对食物资源的利用以及竞争能力越强(Chen et al, 2011; 姜亚洲等, 2015)。本研究中白令海北鳕的营养生态位宽幅最大(SEAc为5.51), 根据以往文献研究结果(Walkusz et al, 2011; Cui et al, 2012; Rand et al, 2013; Gray et al, 2016), 白令海北鳕摄食范围广泛, 对食物资源有很强的利用能力, 这种特性是保证白令海北鳕在北极地区长期处于优势地位的基础。
Kondoh (2003)研究发现具有较大营养生态位宽幅的物种一般属于机会主义摄食者, 抵御外界因素的侵入以及适应环境变化的能力较强, 在资源有限的环境中通过避免激烈的种间竞争与其他物种达到稳定共存的状态。本研究中白令海北鳕虽然具有最大宽幅的营养生态位, 但是与其他物种重叠程度相对较低, 表明白令海北鳕可能根据饵料的可获得性而调整其饵料生物组成, 从而避免与其他物种出现激烈的种间竞争, 这种较强的营养弹性使其在营养生态位竞争中一直保持着较好的优势。而杜父鱼科、绵鳚科与线鳚科等鱼类物种之间营养生态位重叠严重, 因为其中绝大多数物种活动范围较窄, 底栖生物是它们最主要的食物来源, 在有限的空间以及有限的资源内产生了激烈的种间竞争。
附录 Supplementary Material
附录1 Marsh等(2017)和Stasko等(2017)研究中鱼类营养级与稳定同位素特征
参考文献
Preliminary studies on trophic structure of tidal zone in the Laoshan Bay by using carbon stable isotopes
崂山湾潮间带食物网结构的碳稳定同位素初步研究
Establishment of continuous nutrient spectrum of food web in Yellow Sea and East China Sea ecosystem: Results from stable isotope method of carbon and nitrogen
黄东海生态系统食物网连续营养谱的建立: 来自碳氮稳定同位素方法的结果
Ecology of North American Arctic continental shelf benthos: A review
DOI:10.1016/0278-4343(91)90083-I URL [本文引用: 1]
Isotopic niche overlap of two planktivorous fish in southern China
DOI:10.1007/s10201-010-0332-2 URL [本文引用: 1]
Marine ecological and environmental responses to the Arctic rapid change
北极快速变化的生态环境响应
Changes in vegetation growing season and active accumulated temperature in circum-Arctic region under 21st century climate change scenario
21世纪气候变化情景下环北极地区植被生长季与活动积温变化
Trophic ecology of sympatric Arctic gadoids, Arctogadus glacialis (Peters, 1872) and Boreogadus saida (Lepechin, 1774), in NE Greenland
DOI:10.1007/s00300-012-1170-y URL [本文引用: 1]
Feeding ecology of dominant groundfish in the northern Bering Sea
DOI:10.1007/s00300-012-1180-9 URL [本文引用: 2]
A comparison between late summer 2012 and 2013 water masses, macronutrients, and phytoplankton standing crops in the northern Bering and Chukchi Seas
The potential carbon source and trophic level analysis of main organisms in coastal water of Lüsi fishing ground, based on carbon and nitrogen stable isotope analysis
基于稳定同位素方法的吕泗渔场近岸海域主要生物潜在碳源及营养级分析
Variability in the summer diets of juvenile polar cod (Boreogadus saida) in the northeastern Chukchi and western Beaufort Seas
DOI:10.1007/s00300-015-1796-7 URL [本文引用: 4]
Food habits of Arctic staghorn sculpin (Gymnocanthus tricuspis) and shorthorn sculpin (Myoxocephalus scorpius) in the northeastern Chukchi and western Beaufort Seas
DOI:10.1016/j.dsr2.2016.05.013 URL [本文引用: 1]
Light intensity and the interactions between physiology, morphology and stable isotope ratios in five species of seagrass
DOI:10.1016/0022-0981(95)00096-8 URL [本文引用: 1]
Feeding habits of Amblychaeturichthys hexanema in Jiaozhou Bay based on carbon and nitrogen stable isotope analysis
应用碳、氮稳定同位素技术分析胶州湾六丝钝尾虾虎鱼的摄食习性
Climatic Change and Global Warming of Inland Waters: Climate Change Impacts on the Hydrology and Biogeochemistry of Arctic Rivers
Comparing isotopic niche widths among and within communities: SIBER—Stable isotope Bayesian ellipses in R
DOI:10.1111/j.1365-2656.2011.01806.x
PMID:21401589
[本文引用: 1]

1. The use of stable isotope data to infer characteristics of community structure and niche width of community members has become increasingly common. Although these developments have provided ecologists with new perspectives, their full impact has been hampered by an inability to statistically compare individual communities using descriptive metrics. 2. We solve these issues by reformulating the metrics in a Bayesian framework. This reformulation takes account of uncertainty in the sampled data and naturally incorporates error arising from the sampling process, propagating it through to the derived metrics. 3. Furthermore, we develop novel multivariate ellipse-based metrics as an alternative to the currently employed Convex Hull methods when applied to single community members. We show that unlike Convex Hulls, the ellipses are unbiased with respect to sample size, and their estimation via Bayesian inference allows robust comparison to be made among data sets comprising different sample sizes. 4. These new metrics, which we call SIBER (Stable Isotope Bayesian Ellipses in R), open up more avenues for direct comparison of isotopic niches across communities. The computational code to calculate the new metrics is implemented in the free-to-download package Stable Isotope Analysis for the R statistical environment.© 2011 The Authors. Journal of Animal Ecology © 2011 British Ecological Society.
Long-term trends in the trophic structure of the North Sea fish community: Evidence from stable-isotope analysis, size-spectra and community metrics
DOI:10.1007/s00227-002-0905-7 URL [本文引用: 1]
Stable isotope analysis on the feeding character of representative fishes during spring in central and northern East China Sea and south Yellow Sea
基于稳定同位素方法分析东海中北部及黄海南部春季主要鱼类的食性特征
Variation in trophic structure of nekton organisms from the northern East China Sea and adjacent waters based on stable isotope values
基于稳定同位素方法的东海北部及其邻近水域主要游泳动物营养结构变化
Trophic niches of nine gobiid fishes in Xiangshan Bay determined by carbon and nitrogen stable isotope analysis
基于碳、氮稳定同位素技术研究象山港虾虎鱼类营养生态位
以象山港水域9种虾虎鱼为研究对象,利用稳定同位素技术分析虾虎鱼类的营养生态位和种间竞争关系。结果表明:所测样品δ<sup>13</sup>C值和δ<sup>15</sup>N值的变化范围分别为-20.4‰~-12.7‰和6.7‰~14.5‰,不同种类样品的δ<sup>15</sup>N值和δ<sup>13</sup>C值均存在显著差异;象山港虾虎鱼类营养层次整体较低,多数种类在该生态系统中起到次级消费者的作用;舌虾虎鱼(Glossogobius giuris)、矛尾虾虎鱼(Chaeturichthys stigmatias)和六丝钝尾虾虎鱼(Amblychaeturichthys hexanema)的营养生态位宽幅相对较大,而小头栉孔虾虎鱼(Ctenotrypauchen microcephalus)、绿斑缰虾虎鱼(Amoya chlorostigmatoides)和纹缟虾虎鱼(Tridentiger trigonocephalus)等营养生态位宽幅相对较小;象山港虾虎鱼类营养生态位重叠严重,小头栉孔虾虎鱼-孔虾虎鱼(Trypauchen vagina)、髭缟虾虎鱼(Tridentiger barbatus)纹缟虾虎鱼等之间存在激烈的营养生态位竞争。
Foraging adaptation and the relationship between food-web complexity and stability
Ecological theory suggests that complex food webs should not persist because of their inherent instability. "Real" ecosystems often support a large number of interacting species. A mathematical model shows that fluctuating short-term selection on trophic links, arising from a consumer's adaptive food choice, is a key to the long-term stability of complex communities. Without adaptive foragers, food-web complexity destabilizes community composition; whereas in their presence, complexity may enhance community persistence through facilitation of dynamical food-web reconstruction that buffers environmental fluctuations. The model predicts a linkage pattern consistent with field observations.
Trophic structure of pelagic fishery organism assemblage in the central and western South China Sea in spring revealed by carbon and nitrogen stable isotope analysis
基于碳氮稳定同位素的南海中西部海域春季中上层渔业生物群落营养结构
DOI:10.13287/j.1001-9332.202010.038
[本文引用: 1]
根据2018年春季于南海中西部海域进行的灯光罩网渔业生物调查,本研究采用碳氮稳定同位素技术,分析了南海中西部海域中上层渔业生物的碳氮稳定同位素基本特征,构建了连续营养谱,划分了营养功能群,并比较了不同营养功能群间的营养结构特征。结果表明: 采集的23种渔业生物δ<sup>13</sup>C均值最大为鲯鳅,(-17.58±0.21)‰,最小为六带线纹鱼,(-19.86±0.33)‰;δ<sup>15</sup>N均值的变化范围为8.31‰(琉璃玉鲳)~(12.46±0.74)‰(宝刀鱼)。连续营养谱分析表明,采集的中上层渔业生物营养级介于3.01~4.23,其中19种渔业生物(占总渔获种类的83%)的营养级位于3.0~4.0。23种渔业生物可划分成浮游生物食性、游泳生物食性和混合生物食性3个营养功能群。稳定同位素标准椭圆面积(SEA)分析显示,浮游生物食性功能群占据的营养生态位宽幅最大(SEA=1.56‰<sup>2</sup>),其次为混合生物食性功能群(SEA=0.99‰<sup>2</sup>)和游泳生物食性功能群(SEA=0.31‰<sup>2</sup>)。混合生物食性功能群分别与游泳生物食性功能群和浮游生物食性功能群存在17%和26%的营养生态位重叠度,而游泳生物食性功能群与浮游生物食性功能群不存在生态位重叠。
Can stable isotope ratios provide for community-wide measures of trophic structure?
DOI:10.1890/0012-9658(2007)88[42:csirpf]2.0.co;2
PMID:17489452
[本文引用: 3]
Stable isotope ratios (typically of carbon and nitrogen) provide one representation of an organism's trophic niche and are widely used to examine aspects of food web structure. Yet stable isotopes have not been applied to quantitatively characterize community-wide aspects of trophic structure (i.e., at the level of an entire food web). We propose quantitative metrics that can be used to this end, drawing on similar approaches from ecomorphology research. For example, the convex hull area occupied by species in delta13C-delta15N niche space is a representation of the total extent of trophic diversity within a food web, whereas mean nearest neighbor distance among all species pairs is a measure of species packing within trophic niche space. To facilitate discussion of opportunities and limitations of the metrics, we provide empirical and conceptual examples drawn from Bahamian tidal creek food webs. These examples illustrate how this methodology can be used to quantify trophic diversity and trophic redundancy in food webs, as well as to link individual species to characteristics of the food web in which they are embedded. Building from extensive applications of stable isotope ratios by ecologists, the community-wide metrics may provide a new perspective on food web structure, function, and dynamics.
The niche concept revisited: Mechanistic models and community context
DOI:10.2307/1938141 URL [本文引用: 1]
Construction of a continuous trophic spectrum for the food web in Jiaozhou Bay using stable isotope analyses
应用稳定同位素技术构建胶州湾食物网的连续营养谱
Ontogenetic, spatial and temporal variation in trophic level and diet of Chukchi Sea fishes
DOI:10.1016/j.dsr2.2016.07.010 URL [本文引用: 14]
Pacific Arctic Marine Fishes
Fishes of the World, 5th edn
Dietary composition and trophic position of blue swimmer crab (Portunus pelagicus) in Honghai Bay
红海湾远海梭子蟹 Portunus pelagicus的食物组成及营养位置分析
Bayesian stable isotope mixing models
Stable isotopes in ecosystem studies
DOI:10.1146/ecolsys.1987.18.issue-1 URL [本文引用: 1]
Using stable isotopes to estimate trophic position: Models, methods, and assumptions
DOI:10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2 URL [本文引用: 2]
tRophicPosition, an R package for the Bayesian estimation of trophic position from consumer stable isotope ratios
DOI:10.1111/mee3.2018.9.issue-6 URL [本文引用: 2]
The diets of polar cod (Boreogadus saida) from August 2008 in the US Beaufort Sea
The Arctic amplification debate
DOI:10.1007/s10584-005-9017-y URL [本文引用: 1]
Late summer zoogeography of the northern Bering and Chukchi seas
DOI:10.1016/j.dsr2.2016.03.005 URL [本文引用: 1]
Stable isotope data (δ15N, δ13C) for marine fishes and invertebrates from the Beaufort Regional Environmental Assessment Marine Fishes Project, August-September 2012 and 2013
A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap
Considerable progress has been made in the development of statistical tools to quantify trophic relationships using stable isotope ratios, including tools that address size and overlap of isotopic niches. We build upon recent progress and propose a new probabilistic method for determining niche region and pairwise niche overlap that can be extended beyond two dimensions, provides directional estimates of niche overlap, accounts for species-specific distributions in niche space, and, unlike geometric methods, produces consistent and unique bivariate projections of multivariate data. We define the niche region (NR) as a given 95% (or user-defined a) probability region in multivariate space. Overlap is calculated as the probability that an individual from species A is found in the N(R) of species B. Uncertainty is accounted for in a Bayesian framework, and is the only aspect of the methodology that depends on sample size. Application is illustrated with three-dimensional stable isotope data, but practitioners could use any continuous indicator of ecological niche in any number of dimensions. We suggest that this represents an advance in our ability to quantify and compare ecological niches in a way that is more consistent with Hutchinson's concept of an "n-dimensional hypervolume".
Distribution and diet of larval and juvenile Arctic cod (Boreogadus saida) in the shallow Canadian Beaufort Sea
DOI:10.1016/j.jmarsys.2010.09.001 URL [本文引用: 4]
The ocean and cryosphere in a changing climate: Changes, impacts and risks
气候变化中海洋和冰冻圈的变化、影响及风险
Diet compositions and trophic guild structure of the eastern Chukchi Sea demersal fish community
DOI:10.1016/j.dsr2.2016.03.010 URL [本文引用: 2]
A review on some terms related to niche and their measurements
生态位有关术语的定义及计算公式评述

{kind=link}
{kind=link}
{kind=link}
{kind=link}