



气候变化包括全球变暖、极端天气频发等气候变动, 是地球生物正在面临的重大挑战之一(IPCC, 2019)。生物如何应对正在发生的气候变化, 是当前生态学和进化生物学面临的重大课题(Bjorkman et al, 2017)。植物的迁移能力有限, 其种群能否在气候变化情形下生存, 很大程度上取决于它们的适应性和表型可塑性(Parmesan, 2006)。本地适应性(local adaptation)是指自然选择作用于一个种群, 经过数个世代的时间, 该种群产生与当地环境条件相适应的表型和遗传变异, 即在当地环境种群适合度最高(Kawecki & Ebert, 2004)。当环境发生变化时, 已形成本地适应性的种群会变得不适应(maladaptation)。表型可塑性是指同一植物因生长环境变化而表现出不同表型的现象, 它可使植物在较短时间(一个世代)内对环境变化做出响应, 反映了植物快速响应环境变化的能力(Shaw & Etterson, 2012)。开展本地适应性和表型可塑性研究, 可帮助我们预测植物如何应对全球气候变化(Franks et al, 2014)。
普通野生稻(Oryza rufipogon)是亚洲栽培稻(O. sativa)的祖先种(Khush, 1997), 是水稻育种最重要的野生基因库之一, 是国家粮食安全的战略性资源(Song et al, 2005)。温度是限制普通野生稻地理分布的关键气候因子(Huang et al, 2012), 普通野生稻最北分布到中国长江以南的亚热带地区。这些北缘种群已经适应当地较低温环境, 即具有本地适应性(Zhao et al, 2013; Zhou et al, 2013)。对于已适应较低温环境的普通野生稻北缘种群, 表型可塑性将决定其能否迅速响应气候变化情形下的环境变化, 从而维持种群生存。尽管已有研究检测了普通野生稻对土壤理化性质和水分条件变化的表型响应( 喻崎雯 (2012) 普通野生稻对不同pH和水分的生理生态响应. 硕士学位论文, 广西师范大学, 桂林.), 或探讨了普通野生稻种群的本地适应性(Zhao et al, 2013; Zhou et al, 2013; 赵耀, 2014; Xu et al, 2020), 但我们对普通野生稻的表型可塑性仍然知之甚少。
同质园实验(common garden experiment)是检测表型可塑性的有效手段(Chevin et al, 2010)。本研究聚焦普通野生稻对未来气候变化的表型响应, 以“空间代替时间”的策略, 将纬度梯度分布且具有明显遗传分化的3个种群(北缘种群、中部种群、南部种群)分别进行南移(气候变暖)或北移(气候变冷)的同质园实验, 以探讨普通野生稻的响应环境变化的可塑性能力。我们侧重检测与普通野生稻适合度相关的繁殖性状, 从而: (1)通过比较同一种群在不同同质园的表型变化, 来确定表型可塑性能力与本地适应性的具体表现; (2)通过比较不同种群在不同同质园的表现来明确种群间的可塑性和本地适应性的差异, 并在普通野生稻物种层面探讨表型可塑性与本地适应性之间的关系。这些研究结果将促进我们对普通野生稻的表型可塑性和适应性的理解, 并对气候变化情形下普通野生稻的保护提供指导。
1 材料和方法
1.1 材料来源与同质园构建
本研究根据普通野生稻在我国的分布状况, 沿纬度梯度, 选取北缘种群(江西省东乡县, 28°05′ N) (Npop)、中部种群(广东省高州市, 21°56′ N) (Mpop)和南部种群(海南省万宁市, 18°43′ N) (Spop)作为研究对象。这些种群的生境的气候条件存在明显差异, 包括生长季的温度、日照长度、降水等。这些种群间存在明显与温度相关的遗传及表型分化, Npop表现出本地适应性(Zhao et al, 2013)。我们从每个种群中随机选取30株个体(相互间隔5 m), 移栽到复旦大学温室, 在温室中培养一年以上以减少母本效应, 然后通过无性扩繁(分蔸)来产生实验所需材料。
分别在上海市(SH-com)、江西省南昌市(JX-com)和海南省三亚市(HN-com)构建同质园, 不同同质园的气候环境存在明显差异(附录1)。SH- com位于上海市复旦大学江湾校区(31°20′ N), 属于亚热带季风气候, 日照充分, 四季分明, 雨量充沛, 年均温15-22℃, 实验期间平均温度21.6℃。JX-com位于江西省南昌县(28°30′ N), 属于亚热带季风湿润气候, 年均温16-23℃, 实验期间平均温度23.1℃; HN-com同质园位于海南省三亚市(18°21′ N), 属于热带季风气候, 长夏无冬, 年均温25-31℃, 实验期间平均温度26.8℃。JX-com同质园与Npop原生境邻近, 气候环境相似; HN-com同质园与Spop原生境相距较近, 气候环境相似; SH-com纬度上超出普通野生稻现有分布区的北界。HN-com同质园环境相对于Npop和Mpop两个种群来说是南移, 其气候条件可用于模拟未来气候变暖情形下的普通野生稻种群面临的环境条件。JX-com同质园对于Mpop和Spop两个种群是北移, SH-com同质园位置则对所有种群都是北移, 它们可用于检测普通野生稻对气候变冷的响应。
移栽前挖取稻蔸, 分成带完整根系的单个健康分蘖, 每个同质园中每份材料包括3个克隆(分蘖), 于2019年5月1日同时移栽, 实验周期约为7个月, 于2019年12月29日收获所有植株。每个同质园分成3个小区, 每个种群为一个小区, 小区之间间隔1.5 m, 小区内每份材料3个重复随机种植, 间距0.5 m, 小区周边种植隔离行。每个同质园总计种植270株普通野生稻。3个同质园的田间管理(栽种之前施肥、灌溉以及人工除草等)保持同步, 生长过程中保持约20 cm左右的水位。
1.2 表型检测
记录每株材料从移栽到开花的天数, 即花期(days to flower, FD)。在繁殖期, 用通风透水的网袋对未开花的稻穗进行套网袋处理, 以保证自花授粉并收获全部种子, 每株套袋3穗。待50%种子的种皮变为褐色(种子成熟)时, 测定主蘖高度作为株高(plant height, HP), 统计分蘖数(number of total tillers, TN)及有效分蘖数(number of effective tillers, ET); 然后剪取套袋稻穗, 带回实验室统计每穗颖花数(spikelet number per panicle, FN)、每穗实粒数(seed number per panicle, SN), 称量每穗实粒重(seed weight per panicle, SW), 计算单株实粒数(grain number per individual, GN = SN × ET)、单株实粒重(grain weight per individual, GW = SW × ET)、以及结实率(seed set, SS = SN / FN × 100%)。另外, 统计了上海同质园中普通野生稻的越冬存活率。
1.3 数据分析
采用SPSS 19.0中广义线性模型(general linear model, GLM)检验种群、栽培地点、以及两者交互作用对各个表型参数的效应, 设置区块为协变量。采用可塑性指数(index of plasticity)来表征种群的表型可塑性能力, 计算方法为: 特定性状参数值在不同条件下的极值的差的绝对值与最大值的比值(|Max-Min| / Max) (Valladares et al, 2007)。该算法得到的可塑性指数无需再次标准化处理, 可直接用于不同实验组之间的比较, 不受计量单位影响。运用Bonferroni校正的成对t检验来比较种群间表型性状和可塑性指数的差异。
2 结果
2.1 普通野生稻的表型变异
GLM分析结果显示, 普通野生稻所有指标均没有受区块效应影响(P > 0.05)。仅有花期(FD) (F = 6.038, P < 0.05)和每穗颖花数(FN) (F = 10.648, P < 0.05)两个指标在种群间差异显著, 其他指标在种群间没有检测到差异。除了每穗颖花数(FN) (F = 1.661, P = 0.436)、每穗实粒重(SW) (F = 0.097, P = 0.953)和单株实粒重(GW) (F = 0.772, P = 0.68)不受栽培地点影响外, 其余指标均因栽培地点不同而存在明显变化(P < 0.05) (表1)。
表1 种群来源和栽培地点及其相互作用、区块对普通野生稻的表型性状的效应的广义线性模型分析结果(* < 0.05, ** < 0.01, *** < 0.001)。
Table 1
| 变异来源 Source | df | 花期 FD | 株高 HP | 分蘖数 TN | 有效分蘖数 ET | 每穗颖花数 FN | 每穗实粒数 SN | 每穗实粒重 SW | 单株实粒数 GN | 单株实粒重 GW | 结实率 SS |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 种群 Population | 2 | 6.038* | 1.832 | 0.159 | 0.666 | 10.648* | 4.025 | 0.217 | 4.675 | 0.039 | 5.182 |
| 地点 Site | 2 | 14.906* | 6.387* | 17.029** | 8.525* | 1.661 | 17.338*** | 0.097 | 18.283*** | 0.772 | 33.757*** |
| 区块 Block | 1 | 2.067 | 0.256 | 0.444 | 0.094 | 1.696 | 1.306 | 0 | 0.307 | 0.001 | 1.459 |
| 种群 × 地点的互作 Population × Site | 4 | 16.041** | 2.577 | 1.779 | 1.982 | 5.081 | 7.256 | 4.277 | 0.387 | 3.194 | 2.653 |
与南移或北移相比(JX-com vs. HN-com/SH- com), 北缘种群(Npop)的株高(HP)、每穗实粒数(SN)、单株实粒数(GN)、单株实粒重(GW)以及结实率(SS)指标都表现出一定的本地优势。北移后(JX-com vs. SH-com), Npop除SN、SW和SS略有上升外, 其他指标都呈下降趋势, 其中HP显著降低(LSD test, P < 0.05); 南移后(JX-com vs. HN-com)则总体上呈相反趋势, 其中HP、有效分蘖数(ET)、SN和SW均显著下降(P < 0.05), 总分蘖数(TN)显著增加(P < 0.05) (图1, 附录1)。中部种群(Mpop)北移到JX-com与南移到HN-com (JX-com vs. HN-com)相比, 在HP、FN、SN、SW、GN、GW、SS都显著增加(P < 0.05); 再往北移到SH-com (SH-com vs. JX-com), 则花期(FD)显著提前(P < 0.05), SN、SW显著提高(P < 0.05), 其他指标呈下降趋势(图1, 附录2)。南部种群(Spop)北移到JX-com (JX-com vs. HN-com)后, HP和SS显著增加(P < 0.05), ET显著减少(P < 0.05); 进一步北移(即SH-com vs. HN-com), 除SN、SW、GN、GW、SS仍上升外, 其他指标都下降, 特别是花期(FD)明显提前(P < 0.05); 随着北移距离增加(即SH-com vs. JX-com), 3个普通野生稻种群都表现出ET降低(P = 0.0001), SS升高(P < 0.0001), 其他指标没有检测到显著差异(图1, 附录2)。
图1

图1
3个普通野生稻种群在不同同质园的表型性状变化(均值 ± 标准误)。误差棒表示95%的置信区间。Npop: 北缘种群; Mpop, 中部种群; Spop: 南部种群。SH-com: 上海同质园; JX-com: 江西同质园; HN-com: 海南同质园。
Fig. 1
Variations of phenotypic traits of three Oryza rufipogon populations in different common gardens. Error bars indicate 95% confidence intervals. Npop, Northern population; Mpop, Middle population; Spop, Southern population. SH-com, Shanghai common garden; JX-com, Jiangxi common garden; HN-com, Hainan common garden.
上海同质园对普通野生稻来说是超过其分布北缘的相对寒冷生境, 越冬率的结果可以反映植株耐受寒冷环境的能力。越冬率的结果显示, Npop种群有60% (18/30)的样本能在上海越冬存活, Mpop和Spop种群均无越冬存活样本。
2.2 可塑性指数
可塑性指数估算结果显示, 各种群种子繁殖相关性状SN、SW、GW、GN以及SS的表型可塑性指数都大于0.7, 且这些指标的可塑性指数在种群间没有显著差异。FD、TN和ET等指标的可塑性指数在种群间存在明显差异, 其中中间种群Mpop的FD可塑性最大, 南部种群Spop的TN可塑性最小(图2, 附录3, 附录4)。
图2
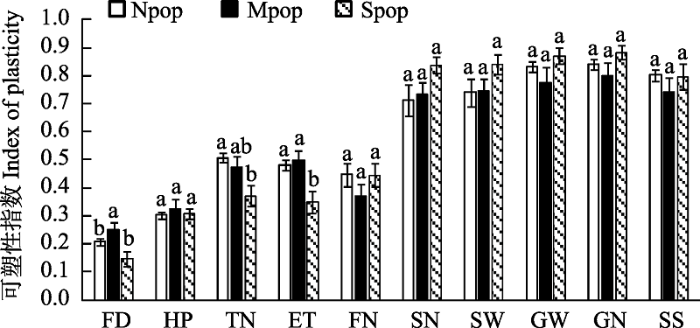
图2
3个普通野生稻种群的可塑性指数(= |Max-Min| / Max)。Max: 最大值; Min: 最小值。Npop: 北缘种群; Mpop, 中部种群; Spop: 南部种群。FD: 花期; HP: 株高; TN: 分蘖数; ET: 有效分蘖数; FN: 每穗颖花数; SN: 每穗实粒数; SW: 每穗实粒重; GN: 单株实粒数; GW: 单株实粒重; SS: 结实率。
Fig. 2
Index of plasticity (= |Max-Min| / Max) of three Oryza rufipofon populations. Max, Maximum value; Min, Minimum value. Npop, Northern population; Mpop, Middle population; Spop, Southern population. FD, Days to flower; HP, Plant height; TN, Number of total tillers; ET, Number of effective tillers; FN, Spikelet number per panicle; SN, Number per panicle; SW, Seed weight per panicle; GN, Grain number per individual; GW, Grain weight per individual; SS, Seed set.
3 讨论
多地同质园实验结果显示各种群具有较强表型可塑性, 3个普通野生稻种群几乎所有检测的表型指标都因生长地点不同而存在明显变化(表1)。特别是种子繁殖性状结实率、种子数量和重量相关性状指标(SN、SW、GW、GN)表现出较强可塑性(图2)。Zhou等(2013)也发现普通野生稻在繁殖相关性状, 如种子产量、旗叶长/宽度、穗长、平均每穗可育/不育颖花等指标上表现出明显的可塑性。这些结果, 一方面说明普通野生稻具有较强的表型可塑性, 能够对环境变化做出快速响应; 另一方面也支持草本植物的某些性状的可塑性与种群来源无关, 而与物种属性有关的观点(Hamann et al, 2017)。可塑性指数估算结果还显示, 普通野生稻的花期(FD)和分蘖数(TN和ET)等指标在种群间表现出明显差异, 中部种群(Mpop)的花期变化明显大于另外两个种群, 而南部种群(Spop)的分蘖变化最小(图2), 表明这些性状的可塑性在种群间出现了分化, 暗示了不同种群对环境变化的响应差异。变化趋势(反应规范)的分析结果也显示, 各种群南移或北移后, 除了株高和结实率的变化趋势类似之外, 其余性状的反应规范都存在不同程度的差异(图1), 进一步说明种群间表型可塑性存在差异(Bjorkman et al, 2017)。种群间可塑性差异与种群遗传分化有关(Albecker et al, 2022), 这暗示种群发生了适应性分化(Simón-Porcar et al, 2021)。
另外, 各种群在本地适应性上表现出明显差异。本地适应性假说认为, 种群在当地环境下的表型表现明显优于其他环境(Kawecki & Ebert, 2004; Hereford, 2017)。JX-com与北缘种群(Npop)原生境邻近, HN-com与南部种群(Spop)原生境邻近, 因此本研究可以检验Npop或Spop是否存在明显的本地适应性。Npop的HP、SN和GW在当地(JX-com)的表现明显优于南移(HN-com)或北移(SH-com) (图1), 说明该种群具有本地适应性。Npop的花期虽然不是在JX-com最短, 但总体呈现随着纬度升高而逐渐提前, 中部种群(Mpop)和南部种群(Spop)则无此变化趋势(图1), 表明Npop会因短日照出现而开花, 即具有感光性。感光性可保障普通野生稻北缘种群在较短生长季中完成生活史, 已被证实是本地适应性表现之一(Zhao et al, 2013)。此外, 所有种群北移之后, 仅有Npop种群保持越冬成活, 表现出很强的耐低温能力, 适应北缘生境的低温环境。普通野生稻北缘种群的低温适应性的分子机制已被部分解析, Xu等(2020)的移植实验发现普通野生稻边缘种群(江西省茶陵种群)北移能够安全越冬, 指示其具有超过边缘限制的扩散能力。相反, Spop仅有表型指标SW在当地(HN-com)表现较好(图1), 没有表现出明显的本地适应性。本地适应性是当地环境条件作为选择压力长期作用于种群的结果(Kawecki & Ebert, 2004)。北缘种群长期生活在温度相对低、短日照出现较早以及生长季较短的环境下, 因此形成了耐低温、感光性等本地适应性。这些结果总体上显示, 普通野生稻不同种群的本地适应性存在明显差异, 种群间存在明显的适应性分化(Zhao et al, 2013); 这也间接支持了边缘-中心理论(Lee-Yaw et al, 2016)。
一般认为, 种群的本地适应性在某种程度上限制表型可塑性(Simón-Porcar et al, 2021), 而本研究普通野生稻的相关结果中并没有发现本地适应性对表型可塑性的限制。本研究结果显示, 尽管北缘种群有较明显的本地适应性, 但各项检测指标均未表现出明显的表型可塑性下降; 相反, 南部种群却在分蘖性状(TN、ET)上可塑性明显较低(图2)。这种现象可能与同质园的地理位置有关。3个同质园的温度差异为Npop提供了较低-适宜-较高的完整温度梯度, 为Mpop提供了较低-较高的温度梯度, 却只为Spop提供了较低-适宜的温度梯度, 因而能够较完整反映Npop和Mpop对于温度的表型变化, 但不能完整反映Spop对温度的表型可塑性。这一结果暗示Spop也具有某种程度的本地适应性(适宜温暖环境), 进一步说明普通野生稻种群存在明显的沿温度(纬度)梯度的适应性分化(Zhao et al, 2013; Zhou et al, 2013)。设置更详细的温度梯度的栽培实验, 将会全面检测来自热带地区的普通野生稻种群的表型可塑性以及本地适应性。
气候变化影响种群生存, 本地适应性降低了种群的耐受性, 使种群对环境变化敏感(Sexton et al, 2011), 表型可塑性则使种群对环境变化做出迅速响应, 为适应新环境提供了可能性(Parmesan & Hanley, 2015)。边缘种群由于其独特的地理位置, 被认为是物种能否适应气候变化的关键(Bontrager & Angert, 2019)。本研究以普通野生稻的北缘种群为核心对象, 以中部和南部种群为对照, 进行移植实验, 以“空间换时间”的策略来模拟未来气候变化情形下普通野生稻种群的表型表现。根据IPCC (2019)报告估计, 北缘种群2080年的环境温度约上升1.5-3℃, 在这种情况下, 南部同质园当前环境温度相当于气候变暖情形下2080年中部种群Mpop面临的环境温度, 比Npop的当前气温高4-6℃, 对于Npop来说属于极端的升温情形。北移实验则相当于模拟了极端低温情形(比如, 明朝小冰河时期(竺可桢, 1972))对普通野生稻种群的影响。Npop南移或北移后的种子量和株高都不同程度下降(图1), 据此推测气候变暖或变冷对北缘种群的繁殖和营养生长都有一定的影响。然而, 无论是北移还是南移, Npop都能够正常结实、完成生活史, 说明普通野生稻北缘种群在未来气候变暖, 即使是极端变暖情形下也能生存。Mpop和Spop种群在JX-com和HN-com同质园中都能正常结实, 甚至北移后繁殖量都有不同程度的提高(图1), 但在SH-com同质园都不能越冬存活。普通野生稻为多年生物种, 越冬存活是种群生存的重要基础。因此, 超出普通野生稻现有分布区的北缘即极端低温环境, 低纬度种群(Mpop、Spop)生存受影响较大。
不同植物对于气候环境变化的响应模式存在差异。多年生草本植物泉卷耳(Cerastium fontanum)在开花物候上表现出对温度变化的表型可塑和逆梯度的局部适应, 即环境温度升高开花相对延后(Valdés et al, 2019)。麒麟菊(Liatris ligulistylis)北移有相对更好的繁殖表现(花蕾数量和结实率), 南移则表现出劣势, 体现了其对低温环境的适应。美耳草属植物Houstonia longifolia南移和北移都表现出适应不良, 表型可塑性能力较弱(Wang et al, 2019)。拟南芥(Arabidopsis thaliana, Wilczek et al, 2014)和美国花旗松(Pseudotsuga menziesii, Bansal et al, 2015)的多地同质园实验则揭示来自温暖地区的种群更适应变暖的环境。本研究尽管所选种群数量较少, 且只有一年的结果, 但仍然可以清晰显示普通野生稻种群与温度相关的适应性和可塑性。特别是, 无论北缘种群(Npop)还是分布中心的种群(Mpop、Spop)都具有较强的表型可塑性, 暗示了普通野生稻响应气候变化的潜力。未来的研究应在不同气候区同时比较不同地理梯度上多个种群的表现, 以进一步探究本地适应性与表型可塑性的关系及其对普通野生稻适应气候变化的影响。
附录 Supplementary Material
附录1 实验期间3个同质园的气候条件
Appendix 1 Climatic conditions of the three common gardens during the experiment
附录2 3个普通野生稻种群在不同同质园的表型差异的LSD检验结果(P值)
Appendix 2 LSD results of phenotypic trait of three Oryza rufipogon populations in different common gardens (P value)
附录3 3个普通野生稻种群在不同同质园的表型可塑性(index of plasticity)的统计结果
Appendix 3 Statistical results of index of plasticity of three Oryza rufipogon populations in different common gardens
附录4 3个普通野生稻种群在不同同质园的表型可塑性的LSD检验结果(P值)
Appendix 4 LSD results of phenotypic plasticity of three Oryza rufipogon populations in different common gardens (P value)
参考文献
A novel analytical framework to quantify co-gradient and countergradient variation
DOI:10.1111/ele.v25.6 URL [本文引用: 1]
Climate-related genetic variation in drought-resistance of Douglas-fir (Pseudotsuga menziesii)
DOI:10.1111/gcb.12719
PMID:25156589
[本文引用: 1]

There is a general assumption that intraspecific populations originating from relatively arid climates will be better adapted to cope with the expected increase in drought from climate change. For ecologically and economically important species, more comprehensive, genecological studies that utilize large distributions of populations and direct measures of traits associated with drought-resistance are needed to empirically support this assumption because of the implications for the natural or assisted regeneration of species. We conducted a space-for-time substitution, common garden experiment with 35 populations of coast Douglas-fir (Pseudotsuga menziesii var. menziesii) growing at three test sites with distinct summer temperature and precipitation (referred to as 'cool/moist', 'moderate', or 'warm/dry') to test the hypotheses that (i) there is large genetic variation among populations and regions in traits associated with drought-resistance, (ii) the patterns of genetic variation are related to the native source-climate of each population, in particular with summer temperature and precipitation, (iii) the differences among populations and relationships with climate are stronger at the warm/dry test site owing to greater expression of drought-resistance traits (i.e., a genotype × environment interaction). During midsummer 2012, we measured the rate of water loss after stomatal closure (transpiration(min)), water deficit (% below turgid saturation), and specific leaf area (SLA, cm(2) g(-1)) on new growth of sapling branches. There was significant genetic variation in all plant traits, with populations originating from warmer and drier climates having greater drought-resistance (i.e., lower transpiration(min), water deficit and SLA), but these trends were most clearly expressed only at the warm/dry test site. Contrary to expectations, populations from cooler climates also had greater drought-resistance across all test sites. Multiple regression analysis indicated that Douglas-fir populations from regions with relatively cool winters and arid summers may be most adapted to cope with drought conditions that are expected in the future.Published 2014. This article is a U.S. Government work and is in the public domain in the USA.
Climate adaptation is not enough: Warming does not facilitate success of southern tundra plant populations in the high Arctic
DOI:10.1111/gcb.13417
PMID:27391174
[本文引用: 2]
Rapidly rising temperatures are expected to cause latitudinal and elevational range shifts as species track their optimal climate north and upward. However, a lack of adaptation to environmental conditions other than climate - for example photoperiod, biotic interactions, or edaphic conditions - might limit the success of immigrants in a new location despite hospitable climatic conditions. Here, we present one of the first direct experimental tests of the hypothesis that warmer temperatures at northern latitudes will confer a fitness advantage to southern immigrants relative to native populations. As rates of warming in the Arctic are more than double the global average, understanding the impacts of warming in Arctic ecosystems is especially urgent. We established experimentally warmed and nonwarmed common garden plots at Alexandra Fiord, Ellesmere Island in the Canadian High Arctic with seeds of two forb species (Oxyria digyna and Papaver radicatum) originating from three to five populations at different latitudes across the Arctic. We found that plants from the local populations generally had higher survival and obtained a greater maximum size than foreign individuals, regardless of warming treatment. Phenological traits varied with latitude of the source population, such that southern populations demonstrated substantially delayed leaf-out and senescence relative to northern populations. Our results suggest that environmental conditions other than temperature may influence the ability of foreign populations and species to establish at more northerly latitudes as the climate warms, potentially leading to lags in northward range shifts for some species.© 2016 John Wiley & Sons Ltd.
Gene flow improves fitness at a range edge under climate change
DOI:10.1002/evl3.91
PMID:30788142
Populations at the margins of a species' geographic range are often thought to be poorly adapted to their environment. According to theoretical predictions, gene flow can inhibit these range edge populations if it disrupts adaptation to local conditions. Alternatively, if range edge populations are small or isolated, gene flow can provide beneficial genetic variation and may facilitate adaptation to environmental change. We tested these competing predictions in the annual wildflower using greenhouse crosses to simulate gene flow from sources across the geographic range into two populations at the northern range margin. We planted these between-population hybrids in common gardens at the range edge and evaluated how genetic differentiation and climatic differences between edge populations and gene flow sources affected lifetime fitness. During an anomalously warm study year, gene flow from populations occupying historically warm sites improved fitness at the range edge and plants with one or both parents from warm populations performed best. The effects of the temperature provenance of gene flow sources were most apparent at early life history stages, but precipitation provenance also affected reproduction. We also found benefits of gene flow that were independent of climate: after climate was controlled for, plants with parents from different populations performed better at later lifestages than those with parents from the same population, indicating that gene flow may improve fitness via relieving homozygosity. Further supporting this result, we found that increasing genetic differentiation of parental populations had positive effects on fitness of hybrid seeds. Gene flow from warmer populations, when it occurs, is likely to contribute adaptive genetic variation to populations at the northern range edge as the climate warms. On heterogeneous landscapes, climate of origin may be a better predictor of gene flow effects than geographic proximity.
Evolutionary and plastic responses to climate change in terrestrial plant populations
DOI:10.1111/eva.12112
PMID:24454552
[本文引用: 1]
As climate change progresses, we are observing widespread changes in phenotypes in many plant populations. Whether these phenotypic changes are directly caused by climate change, and whether they result from phenotypic plasticity or evolution, are active areas of investigation. Here, we review terrestrial plant studies addressing these questions. Plastic and evolutionary responses to climate change are clearly occurring. Of the 38 studies that met our criteria for inclusion, all found plastic or evolutionary responses, with 26 studies showing both. These responses, however, may be insufficient to keep pace with climate change, as indicated by eight of 12 studies that examined this directly. There is also mixed evidence for whether evolutionary responses are adaptive, and whether they are directly caused by contemporary climatic changes. We discuss factors that will likely influence the extent of plastic and evolutionary responses, including patterns of environmental changes, species' life history characteristics including generation time and breeding system, and degree and direction of gene flow. Future studies with standardized methodologies, especially those that use direct approaches assessing responses to climate change over time, and sharing of data through public databases, will facilitate better predictions of the capacity for plant populations to respond to rapid climate change.
Plant responses to simulated warming and drought: A comparative study of functional plasticity between congeneric mid and high elevation species
DOI:10.1093/jpe/rtx023 URL [本文引用: 1]
Genetic divergence for physiological response to temperature between populations of a C3-C 4 intermediate annual
DOI:10.1086/691695 URL [本文引用: 1]
Phylogeography of Asian wild rice, Oryza rufipogon: A genome-wide view
DOI:10.1111/j.1365-294X.2012.05625.x
PMID:22646149
[本文引用: 1]
Asian wild rice (Oryza rufipogon) that ranges widely across the eastern and southern part of Asia is recognized as the direct ancestor of cultivated Asian rice (O. sativa). Studies of the geographic structure of O. rufipogon, based on chloroplast and low-copy nuclear markers, reveal a possible phylogeographic signal of subdivision in O. rufipogon. However, this signal of geographic differentiation is not consistently observed among different markers and studies, with often conflicting results. To more precisely characterize the phylogeography of O. rufipogon populations, a genome-wide survey of unlinked markers, intensively sampled from across the entire range of O. rufipogon is critical. In this study, we surveyed sequence variation at 42 genome-wide sequence tagged sites (STS) in 108 O. rufipogon accessions from throughout the native range of the species. Using Bayesian clustering, principal component analysis and amova, we conclude that there are two genetically distinct O. rufipogon groups, Ruf-I and Ruf-II. The two groups exhibit a clinal variation pattern generally from north-east to south-west. Different from many earlier studies, Ruf-I, which is found mainly in China and the Indochinese Peninsula, shows genetic similarity with one major cultivated rice variety, O. satvia indica, whereas Ruf-II, mainly from South Asia and the Indochinese Peninsula, is not found to be closely related to cultivated rice varieties. The other major cultivated rice variety, O. sativa japonica, is not found to be similar to either O. rufipogon groups. Our results support the hypothesis of a single origin of the domesticated O. sativa in China. The possible role of palaeoclimate, introgression and migration-drift balance in creating this clinal variation pattern is also discussed.© 2012 Blackwell Publishing Ltd.
2019 refinement to the 2006 IPCC guidelines for national greenhouse gas inventories
In: Key Concepts Unchanged from the 2006 IPCC Guidelines (eds Calvo BE, Tanabe K, Kranjc A, Baasansuren J, Fukuda M, Ngarize S, Osako A, Pyrozhenko Y, Shermanau P, Federici S), pp. 45. IPCC, Interlaken.
Conceptual issues in local adaptation
DOI:10.1111/ele.2004.7.issue-12 URL [本文引用: 3]
Origin, dispersal, cultivation and variation of rice
There are two cultivated and twenty-one wild species of genus Oryza. O. sativa, the Asian cultivated rice is grown all over the world. The African cultivated rice, O. glaberrima is grown on a small scale in West Africa. The genus Oryza probably originated about 130 million years ago in Gondwanaland and different species got distributed into different continents with the breakup of Gondwanaland. The cultivated species originated from a common ancestor with AA genome. Perennial and annual ancestors of O. sativa are O. rufipogon and O. nivara and those of O. glaberrima are O. longistaminata, O. breviligulata and O. glaberrima probably domesticated in Niger river delta. Varieties of O. sativa are classified into six groups on the basis of genetic affinity. Widely known indica rices correspond to group I and japonicas to group VI. The so called javanica rices also belong to group VI and are designated as tropical japonicas in contrast to temperate japonicas grown in temperate climate. Indica and japonica rices had a polyphyletic origin. Indicas were probably domesticated in the foothills of Himalayas in Eastern India and japonicas somewhere in South China. The indica rices dispersed throughout the tropics and subtropics from India. The japonica rices moved northward from South China and became the temperate ecotype. They also moved southward to Southeast Asia and from there to West Africa and Brazil and became tropical ecotype. Rice is now grown between 55 degrees N and 36 degrees S latitudes. It is grown under diverse growing conditions such as irrigated, rainfed lowland, rainfed upland and floodprone ecosystems. Human selection and adaptation to diverse environments has resulted in numerous cultivars. It is estimated that about 120,000 varieties of rice exist in the world. After the establishment of International Rice Research Institute in 1960, rice varietal improvement was intensified and high yielding varieties were developed. These varieties are now planted to 70% of world's riceland. Rice production doubled between 1966 and 1990 due to large scale adoption of these improved varieties. Rice production must increase by 60% by 2025 to feed the additional rice consumers. New tools of molecular and cellular biology such as anther culture, molecular marker aided selection and genetic engineering will play increasing role in rice improvement.
A synthesis of transplant experiments and ecological niche models suggests that range limits are often niche limits
DOI:10.1111/ele.12604
PMID:27111656
[本文引用: 1]
Global change has made it important to understand the factors that shape species' distributions. Central to this area of research is the question of whether species' range limits primarily reflect the distribution of suitable habitat (i.e. niche limits) or arise as a result of dispersal limitation. Over-the-edge transplant experiments and ecological niche models are commonly used to address this question, yet few studies have taken advantage of a combined approach for inferring the causes of range limits. Here, we synthesise results from existing transplant experiments with new information on the predicted suitability of sites based on niche models. We found that individual performance and habitat suitability independently decline beyond range limits across multiple species. Furthermore, inferences from transplant experiments and niche models were generally concordant within species, with 31 out of 40 cases fully supporting the hypothesis that range limits are niche limits. These results suggest that range limits are often niche limits and that the factors constraining species' ranges operate at scales detectable by both transplant experiments and niche models. In light of these findings, we outline an integrative framework for addressing the causes of range limits in individual species. © 2016 John Wiley & Sons Ltd/CNRS.
Ecological and evolutionary responses to recent climate change
DOI:10.1146/ecolsys.2006.37.issue-1 URL [本文引用: 1]
Plants and climate change: Complexities and surprises
DOI:10.1093/aob/mcv169
PMID:26555281
[本文引用: 1]
Anthropogenic climate change (ACC) will influence all aspects of plant biology over coming decades. Many changes in wild species have already been well-documented as a result of increased atmospheric CO2 concentrations, warming climate and changing precipitation regimes. A wealth of available data has allowed the use of meta-analyses to examine plant-climate interactions on more sophisticated levels than before. These analyses have revealed major differences in plant response among groups, e.g. with respect to functional traits, taxonomy, life-history and provenance. Interestingly, these meta-analyses have also exposed unexpected mismatches between theory, experimental, and observational studies.We reviewed the literature on species' responses to ACC, finding ∼42 % of 4000 species studied globally are plants (primarily terrestrial). We review impacts on phenology, distributions, ecophysiology, regeneration biology, plant-plant and plant-herbivore interactions, and the roles of plasticity and evolution. We focused on apparent deviations from expectation, and highlighted cases where more sophisticated analyses revealed that unexpected changes were, in fact, responses to ACC.We found that conventionally expected responses are generally well-understood, and that it is the aberrant responses that are now yielding greater insight into current and possible future impacts of ACC. We argue that inconclusive, unexpected, or counter-intuitive results should be embraced in order to understand apparent disconnects between theory, prediction, and observation. We highlight prime examples from the collection of papers in this Special Issue, as well as general literature. We found use of plant functional groupings/traits had mixed success, but that some underutilized approaches, such as Grime's C/S/R strategies, when incorporated, have improved understanding of observed responses. Despite inherent difficulties, we highlight the need for ecologists to conduct community-level experiments in systems that replicate multiple aspects of ACC. Specifically, we call for development of coordinating experiments across networks of field sites, both natural and man-made.© The Author 2015. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Gene flow increases fitness at the warm edge of a species’ range
Rapid climate change and the rate of adaptation: Insight from experimental quantitative genetics
DOI:10.1111/j.1469-8137.2012.04230.x
PMID:22816320
[本文引用: 1]
Evolution proceeds unceasingly in all biological populations. It is clear that climate-driven evolution has molded plants in deep time and within extant populations. However, it is less certain whether adaptive evolution can proceed sufficiently rapidly to maintain the fitness and demographic stability of populations subjected to exceptionally rapid contemporary climate change. Here, we consider this question, drawing on current evidence on the rate of plant range shifts and the potential for an adaptive evolutionary response. We emphasize advances in understanding based on theoretical studies that model interacting evolutionary processes, and we provide an overview of quantitative genetic approaches that can parameterize these models to provide more meaningful predictions of the dynamic interplay between genetics, demography and evolution. We outline further research that can clarify both the adaptive potential of plant populations as climate continues to change and the role played by ongoing adaptation in their persistence.© 2012 The Authors. New Phytologist © 2012 New Phytologist Trust.
Rapid local adaptation in both sexual and asexual invasive populations of monkeyflowers (Mimulus spp.)
DOI:10.1093/aob/mcab004
PMID:33604608
[本文引用: 2]
Traditionally, local adaptation has been seen as the outcome of a long evolutionary history, particularly with regard to sexual lineages. By contrast, phenotypic plasticity has been thought to be most important during the initial stages of population establishment and in asexual species. We evaluated the roles of adaptive evolution and phenotypic plasticity in the invasive success of two closely related species of invasive monkeyflowers (Mimulus) in the UK that have contrasting reproductive strategies: M. guttatus combines sexual (seeds) and asexual (clonal growth) reproduction while M. × robertsii is entirely asexual.We compared the clonality (number of stolons), floral and vegetative phenotype, and phenotypic plasticity of native (M. guttatus) and invasive (M. guttatus and M. × robertsii) populations grown in controlled environment chambers under the environmental conditions at each latitudinal extreme of the UK. The goal was to discern the roles of temperature and photoperiod on the expression of phenotypic traits. Next, we tested the existence of local adaptation in the two species within the invasive range with a reciprocal transplant experiment at two field sites in the latitudinal extremes of the UK, and analysed which phenotypic traits underlie potential local fitness advantages in each species.Populations of M. guttatus in the UK showed local adaptation through sexual function (fruit production), while M. × robertsii showed local adaptation via asexual function (stolon production). Phenotypic selection analyses revealed that different traits are associated with fitness in each species. Invasive and native populations of M. guttatus had similar phenotypic plasticity and clonality. M. × robertsii presents greater plasticity and clonality than native M. guttatus, but most populations have restricted clonality under the warm conditions of the south of the UK.This study provides experimental evidence of local adaptation in a strictly asexual invasive species with high clonality and phenotypic plasticity. This indicates that even asexual taxa can rapidly (<200 years) adapt to novel environmental conditions in which alternative strategies may not ensure the persistence of populations.© The Author(s) 2021. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Genetic diversity and conservation of common wild rice (Oryza rufipogon) in China
DOI:10.1111/psb.2005.20.issue-2 URL [本文引用: 1]
A natural heating experiment: Phenotypic and genotypic responses of plant phenology to geothermal soil warming
DOI:10.1111/gcb.14525
PMID:30430704
Under global warming, the survival of many populations of sedentary organisms in seasonal environments will largely depend on their ability to cope with warming in situ by means of phenotypic plasticity or adaptive evolution. This is particularly true in high-latitude environments, where current growing seasons are short, and expected temperature increases large. In such short-growing season environments, the timing of growth and reproduction is critical to survival. Here, we use the unique setting provided by a natural geothermal soil warming gradient (Hengill geothermal area, Iceland) to study the response of Cerastium fontanum flowering phenology to temperature. We hypothesized that trait expression and phenotypic selection on flowering phenology are related to soil temperature, and tested the hypothesis that temperature-driven differences in selection on phenology have resulted in genetic differentiation using a common garden experiment. In the field, phenology was related to soil temperature, with plants in warmer microsites flowering earlier than plants at colder microsites. In the common garden, plants responded to spring warming in a counter-gradient fashion; plants originating from warmer microsites flowered relatively later than those originating from colder microsites. A likely explanation for this pattern is that plants from colder microsites have been selected to compensate for the shorter growing season by starting development at lower temperatures. However, in our study we did not find evidence of variation in phenotypic selection on phenology in relation to temperature, but selection consistently favoured early flowering. Our results show that soil temperature influences trait expression and suggest the existence of genetically based variation in flowering phenology leading to counter-gradient local adaptation along a gradient of soil temperatures. An important implication of our results is that observed phenotypic responses of phenology to global warming might often be a combination of short-term plastic responses and long-term evolutionary responses, acting in different directions.© 2018 John Wiley & Sons Ltd.
Ecological limits to plant phenotypic plasticity
DOI:10.1111/j.1469-8137.2007.02275.x
PMID:17997761
[本文引用: 1]
Phenotypic plasticity is considered the major means by which plants cope with environmental heterogeneity. Although ubiquitous in nature, actual phenotypic plasticity is far from being maximal. This has been explained by the existence of internal limits to its expression. However, phenotypic plasticity takes place within an ecological context and plants are generally exposed to multifactor environments and to simultaneous interactions with many species. These external, ecological factors may limit phenotypic plasticity or curtail its adaptive value, but seldom have they been considered because limits to plasticity have typically addressed factors internal to the plant. We show that plastic responses to abiotic factors are reduced under situations of conservative resource use in stressful and unpredictable habitats, and that extreme levels in a given abiotic factor can negatively influence plastic responses to another factor. We illustrate how herbivory may limit plant phenotypic plasticity because damaged plants can only rarely attain the optimal phenotype in the challenging environment. Finally, it is examined how phenotypic changes involved in trait-mediated interactions can entail costs for the plant in further interactions with other species in the community. Ecological limits to plasticity must be included in any realistic approach to understand the evolution of plasticity in complex environments and to predict plant responses to global change.
Experimental test of assisted migration for conservation of locally range-restricted plants in Alberta, Canada
Studies on the cold tolerance of ratoon ‘Chaling’ common wild rice
DOI:10.1186/s40659-020-00276-5
[本文引用: 2]
Rice is the staple food of many people around the world. However, most rice varieties, especially widely grown indica varieties and hybrids, are sensitive to cold stress. In order to provide a basis for the utilization of a common wild rice (CWR, Oryza rufipogon Griff.) named ‘Chaling’ CWR in cold-tolerant rice breeding and deepen the understanding of rice cold tolerance, the cold tolerance of ratoon ‘Chaling’ CWR was studied under the stress of the natural low temperature in winter in Changsha, Hunan province, China, especially under the stress of abnormal natural low temperature in Changsha in 2008, taking other ratoon CWR accessions and ratoon cultivated rice phenotypes as control.
The Mechanism Shaping Distribution Pattern of Population Variations in Oryza rufipogon Griff
普通野生稻种群变异的分布格局及其形成机制
Geographic variation and local adaptation in Oryza rufipogon across its climatic range in China
DOI:10.1111/1365-2745.12143 URL [本文引用: 4]

{kind=link}
{kind=link}
{kind=link}
{kind=link}