



生物多样性对生态系统的影响一直是生态学领域研究的核心问题(Hooper et al, 2005; Maureaud et al, 2019)。以往对生物多样性的测度主要集中在物种多样性方面, 但是不同物种的生理、形态特征等方面都存在巨大差异(Tilman, 2001), 当两个群落的物种种数和相对丰度相同时, 物种多样性就无法区分两个群落多样性的差异。物种的功能特征是联系群落结构和功能的关键因素(陈又清, 2017; Huang et al, 2019)。功能多样性是指特定生态系统中所有物种功能特征的数值和范围(Esler & Rebelo, 2014)。它关注的是与生态系统密切相关的物种的功能特征, 能够反映群落内物种间资源互补的程度(Dı́az & Cabido, 2001; Esler & Rebelo, 2014)。功能多样性可以通过物种的不同功能特征来解释群落差异, 能够充分反映各个物种在生态系统中所起的作用(Lepš et al, 2001; Teichert et al, 2018)。目前功能多样性较多应用在植物和微生物方面, 在鱼类方面应用较少(冯慧芳等, 2021; 盘远方等, 2021)。
本研究于2019年和2020年对三峡库区库首秭归、库中云阳、库尾巴南及库首支流香溪河下游峡口、库中支流小江下游高阳、库尾支流嘉陵江下游合川等江段的鱼类进行了调查, 分析了鱼类群落结构和物种多样性, 从摄食、运动和繁殖3个方面探讨了鱼类功能多样性空间格局, 旨在了解三峡库区及主要支流鱼类群落结构和功能多样性空间格局, 探讨功能多样性和物种多样性之间的关系, 以期为三峡库区鱼类多样性保护和管理提供科学依据, 为长江“十年禁渔”效果评估提供本底资料。
1 材料与方法
1.1 鱼类调查
2019年8-11月和2020年5-8月对三峡库区及其主要支流不同江段的鱼类群落进行了野外调查采样, 在三峡库区库首、库中和库尾分别设置秭归(ZG)、云阳(YY)和巴南(BN)等3个调查站点, 在库首支流香溪河下游、库中支流小江下游和库尾支流嘉陵江下游分别设置峡口(XK)、高阳(GY)和合川(HC)等3个调查站点(图1)。调查网具使用单层多网目复合刺网和地笼。复合刺网由1 cm、1.6 cm、2 cm、2.5 cm、3.1 cm、4 cm、4.8 cm、6 cm、7.5 cm、8.5 cm、11 cm、12.5 cm等12种不同网目的刺网拼接而成, 每个网目的刺网长2.5 m, 高2 m, 复合刺网总长30 m。地笼网目为1.2 cm。采样期间, 每天下午6点下网, 次日早晨6点收网。现场鉴定并记录渔获物种类, 测量鱼类的全长、体长和体重, 全长和体长精确到1 mm, 体重精确到1 g。对于疑难物种, 生物学性状测量后取肌肉用95%酒精保存, 另用10%福尔马林溶液保存鱼类标本, 带回实验室进行分子鉴定和进一步的物种形态鉴定。每个站点根据鱼类多少情况调查7-10 d。
图1
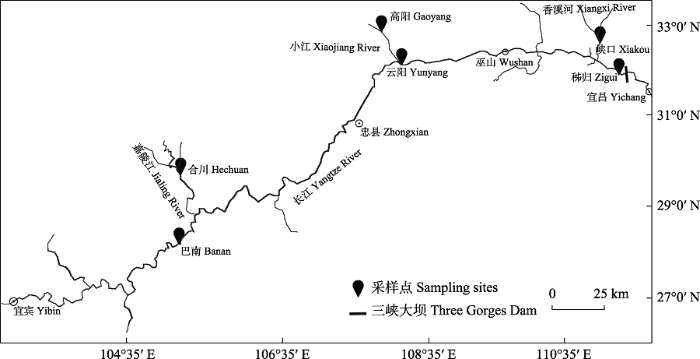
1.2 群落结构分析
基于采样点和物种个体数计算鱼类群落Bray- Curtis (BC)相异性指数, 通过非度量多维尺度排序(non-metric multidimensional scale, NMDS)分析各江段群落组成的差异性。用胁迫系数(stress)衡量NMDS图的优劣, stress < 0.2时, 表示排序结果合理。
式中, Si为江段i的物种个体数之和, Sj为江段j的物种个体数之和, Cij为江段i和江段j共有种中个体数目较少者之和。BC指数介于0-1之间, 值越高表示两群落结构差异越大。
1.3 物种多样性分析
物种多样性指数能够反映群落结构稳定性, 从物种数和物种个体数的均匀性两方面衡量群落结构, 本文选取Margalef丰富度指数(D)、Shannon多样性指数(H′)和Pielou均匀度指数(E)来描述物种多样性(Ludwig & Reynolds, 1988)。计算公式如下:
式中, S是群落中总的物种数, N是总个体数, Wi是群落中第i个物种的个体数占群落中总个体数的比例。
1.4 功能多样性分析
1.4.1 鱼类功能性状
根据Villéger等(2017)的方法, 测量有关摄食、运动和繁殖3个维度的功能性状。其中摄食维度包括食性、营养级、口位等8个功能性状, 运动维度包括最大体长(cm)和体型等6个功能性状, 繁殖维度包括平均绝对繁殖力、平均相对繁殖力和繁殖季节等6个功能性状(附录1)。通过查阅文献和世界鱼类资料库Fishbase (
1.4.2 功能多样性指数
式中, SFic指群落中所有物种所占据的生态位, Rc指特征值的绝对值。
功能均匀度(FEve)衡量物种性状的相对丰度在功能空间分布的均匀程度, 包含物种的相对丰度, 反映了物种对整体资源的利用情况(Villéger et al, 2008), 计算公式如下:
式中, EWl为均匀度权重, dist(i, j)为物种i和j的欧式距离, Wj为物种j的相对丰度, l为分支长, PEWl为分支长权重。
功能离散度(FDiv)表示物种功能性状的相对丰度在功能空间分布的离散程度, 同样包含物种的相对丰度, 反映了资源分异度、物种之间的竞争程度等(Villéger et al, 2008)。
式中, Xik为物种i性状k值, gk为性状k的重心, T为性状数, Δ|d|为物种i与重心的平均距离, Δd为以相对丰度为权重的离散度。
同时选取群落特征加权平均数指数(community-weighted mean, CWM)作为功能多样性的补充。群落特征加权平均数指数为群落内物种功能性状的加权平均值, 能够以物种丰度为权重来量化群落性状(Lavorel et al, 2008)。基于各江段物种的相对丰度和3个维度的功能性状分别计算各江段摄食功能的CWM (F)、运动功能的CWM (L)和繁殖功能的CWM (R)。
式中, traiti为物种i的性状值。
1.5 数据分析
运用Excel 2019对鱼类丰度和性状数据进行收集和整理。考虑到同时具有分类型变量和连续型变量, 基于Gower距离计算功能多样性指数。应用R 4.1.0软件的“vegan”和“FD”包计算不同江段的Bray-Curtis相异性指数、物种多样性指数和功能多样性指数。对所有的物种多样性指数和功能多样性指数进行Pearson相关分析。
2 结果
2.1 鱼类群落结构
2019-2020年在三峡库区及主要支流采集鱼类18,154尾, 鉴定出鱼类78种, 隶属于6目15科56属。其中, 秭归江段采集到鱼类42种, 云阳江段47种, 巴南江段52种, 香溪河下游峡口江段24种, 小江下游高阳江段21种, 嘉陵江下游合川江段18种。鲤形目种数最多, 包括3科41属55种, 占总种数的70.5%。其次是鲇形目, 包括4科7属14种, 占总种数的17.9%。鲈形目包括5科5属6种, 占总种数的7.6%。其余的鲱形目、颌针鱼目和鲑形目各1科1属1种, 占总种数的1.2%。
各江段渔获物中数量百分比大于2%的鱼类共有19种(表1)。库首秭归江段短颌鲚(Coilia brachygnathus)占绝对优势, 相对丰度高达77.9%。库中云阳江段贝氏䱗(Hemiculter bleekeri)、蛇鮈(Saurogobio dabryi)和光泽黄颡鱼(Pelteobagrus nitidus)等占优势, 相对丰度分别为28.5%、16.5%和11.7%。库尾巴南江段蛇鮈和鲫(Carassius auratus)占优势, 相对丰度分别为31.3%和10.6%。香溪河下游峡口江段贝氏䱗、似鳊(Pseudobrama simoni)和短颌鲚占优势, 相对丰度分别为32.1%、20.0%和19.7%。小江下游高阳江段贝氏䱗和似鳊占优势, 相对丰度分别为53.2%和15.1%。嘉陵江下游合川江段光泽黄颡鱼和蛇鮈占优势, 相对丰度分别为42.1%和34.6%。总体上, 库首秭归和支流香溪河下游峡口、小江下游高阳江段的短颌鲚和贝氏䱗等静水性鱼类的相对丰度较高, 库中云阳、库尾巴南和支流嘉陵江下游合川江段的蛇鮈和光泽黄颡鱼等广适性鱼类相对丰度较高。在库首、库中及支流相对丰度较高的短颌鲚(相对丰度5.8%-77.9%), 在库尾巴南及支流嘉陵江下游合川江段并未出现。
表1 三峡库区各江段渔获物数量百分比
Table 1
| 种类 Species | 秭归 Zigui | 云阳 Yunyang | 巴南 Banan | 峡口 Xiakou | 高阳 Gaoyang | 合川 Hechuan |
|---|---|---|---|---|---|---|
| 短颌鲚 Coilia brachygnathus | 77.9 | 7.0 | - | 19.7 | 5.8 | - |
| 似鳊 Pseudobrama simoni | 4.4 | 8.4 | - | 20.0 | 15.1 | 6.1 |
| 贝氏䱗 Hemiculter bleekeri | 3.5 | 28.5 | 5.0 | 32.1 | 53.2 | 2.2 |
| 银鮈 Squalidus argentatus | 2.8 | 3.9 | 1.9 | 3.3 | 0.1 | 0.2 |
| 蛇鮈 Saurogobio dabryi | 2.5 | 16.5 | 31.3 | 1.8 | 5.7 | 34.6 |
| 子陵吻鰕虎鱼 Rhinogobius giurinus | 2.3 | 0.4 | 2.0 | 0.7 | - | - |
| 瓦氏黄颡鱼 Pelteobagrus vachelli | 0.5 | 1.3 | 3.4 | 0.7 | 2.9 | 0.8 |
| 长须黄颡鱼 Pelteobagrus eupogon | 0.4 | - | 2.0 | - | - | - |
| 鲤 Cyprinus carpio | 0.3 | 2.8 | 5.7 | - | 0.1 | - |
| 䱗 Hemiculter leucisculus | 0.1 | 2.9 | 2.9 | 2.0 | - | 1.0 |
| 鲫 Carassius auratus | 0.04 | 2.3 | 10.6 | - | 1.4 | 0.6 |
| 飘鱼 Pseudolaubuca sinensis | 0.02 | - | 0.09 | 0.3 | - | 4.0 |
| 鲇 Silurus asotus | 0.01 | 2.1 | 1.5 | 0.07 | 0.1 | - |
| 张氏䱗 Hemiculter tchangi | - | - | 3.8 | 2.0 | 6.6 | 0.4 |
| 黄颡鱼 Pelteobagrus fulvidraco | - | 0.1 | 3.2 | - | 0.1 | - |
| 光泽黄颡鱼 Pelteobagrus nitidus | - | 11.7 | 5.8 | 3.2 | 1.2 | 42.1 |
| 唇䱻 Hemibarbus labeo | - | 0.03 | 3.8 | - | - | - |
| 大鳞副泥鳅 Paramisgurnus dabryanus | - | - | 2.6 | - | - | - |
| 黑尾近红鲌 Ancherythroculter nigrocauda | - | 0.5 | - | - | - | 5.9 |
| 其他 Others | 5.23 | 11.57 | 14.41 | 14.13 | 7.7 | 2.1 |
三峡库区及主要支流各江段以广适性和静水性鱼类为主, 占鱼类总种数的76.6% ± 8.7%, 占鱼类总数量的93.5% ± 3.94% (图2)。库首秭归和支流香溪河峡口、小江高阳江段以静水性鱼类占优势, 种类百分比分别为26.2%、33.3%、33.3%, 数量百分比分别为82.6%、58.0%、70.9%。库中云阳、库尾巴南和嘉陵江合川江段均以广适性鱼类占优势, 种类百分比分别为42.6%、50.0%和44.4%, 数量百分比分别为53.6%、75.3%和88.5%。静水性鱼类的种类百分比和数量百分比在干流江段为: 库首 > 库中 > 库尾, 在支流江段为: 香溪河下游峡口 > 小江高阳 > 嘉陵江合川, 总体上从库首到库尾呈现降低的趋势。而流水性鱼类的种类百分比和数量百分比呈现相反的趋势。
图2
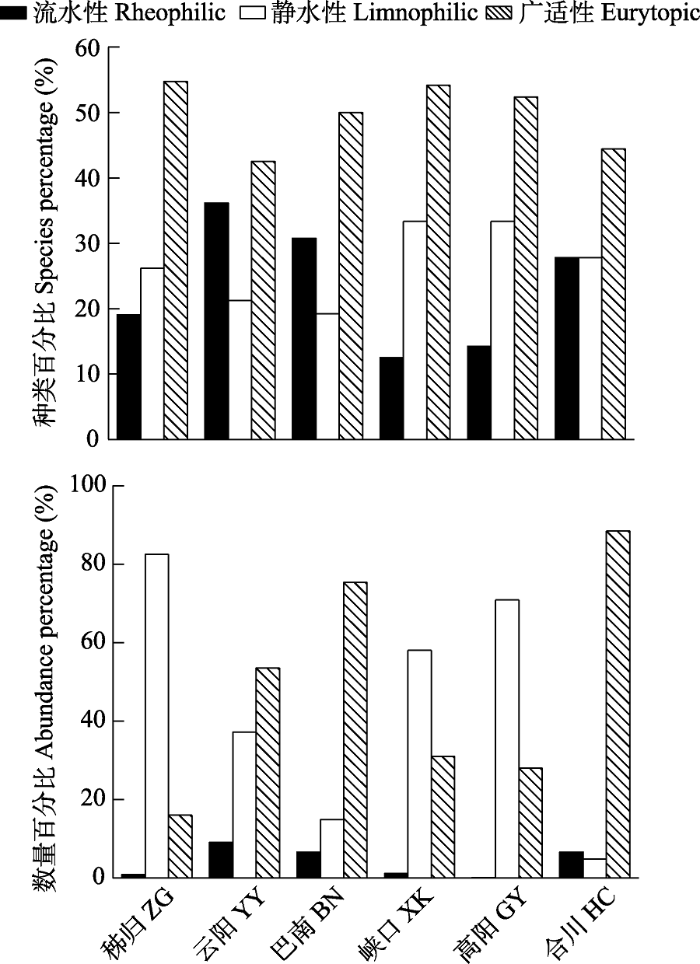
图2
三峡库区不同江段各鱼类生态类群的种类和数量百分比
Fig. 2
Percentages in species and abundance of each fish ecological group in different river section of the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
图3

图3
三峡库区鱼类群落非度量多维尺度排序图
Fig. 3
Non-metric multidimensional scale ranking of fish communities in the Three Gorges Reservoir
表2 三峡库区各江段鱼类群落Bray-Curtis相异性指数
Table 2
| 秭归 Zigui | 云阳 Yunyang | 巴南 Banan | 峡口 Xiakou | 高阳 Gaoyang | |
|---|---|---|---|---|---|
| 云阳 Yunyang | 0.77 | ||||
| 巴南 Banan | 0.88 | 0.59 | |||
| 峡口 Xiakou | 0.79 | 0.49 | 0.83 | ||
| 高阳 Gaoyang | 0.84 | 0.40 | 0.80 | 0.36 | |
| 合川 Hechuan | 0.94 | 0.73 | 0.79 | 0.85 | 0.84 |
2.2 物种多样性
三峡库区各江段鱼类群落Margalef指数的变化范围为2.72-6.25, Shannon指数为1.08-2.68, Pielou均匀度指数为0.29-0.68。3个物种多样性指数在干流江段为: 秭归 < 云阳 < 巴南, 表明物种多样性从库首到库尾呈升高趋势。在支流江段为: 香溪河下游峡口 > 小江下游高阳 > 嘉陵江下游合川, 嘉陵江下游合川的物种多样性最低(图4)。
图4
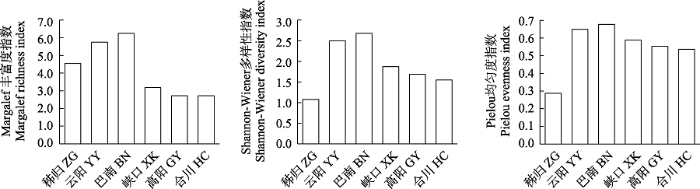
图4
三峡库区不同江段鱼类群落物种多样性空间变化
Fig. 4
Spatial variation of species diversity of fish communities in different river sections of the the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
2.3 功能多样性
2.3.1 摄食功能多样性
三峡库区各江段鱼类群落摄食功能丰富度的变化范围为0.01-0.10, 摄食功能均匀度为0.35-0.58, 摄食功能离散度为0.81-0.94 (图5)。3个摄食功能指数在干流江段为: 秭归 > 云阳 > 巴南。摄食功能丰富度和摄食功能均匀度在支流江段为: 香溪河下游峡口 > 小江下游高阳 > 嘉陵江下游合川, 而摄食功能离散度相差不大, 表明整体上摄食功能多样性从库首到库尾表现出降低的趋势。其中库首秭归江段的3个摄食功能指数均较高(0.10、0.53、0.94), 表明库首秭归江段的摄食功能多样性最高, 嘉陵江下游合川江段的3个摄食功能指数均最低(0.01、0.35、0.83), 表明嘉陵江合川江段的摄食功能多样性最低。
图5
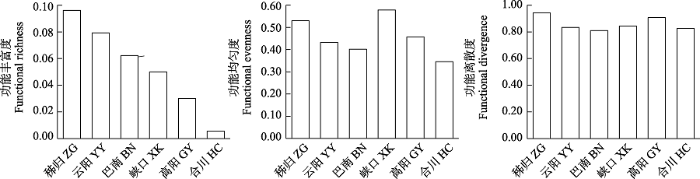
图5
三峡库区鱼类群落摄食功能多样性空间变化
Fig. 5
Spatial variation of food acquisition function diversity of fish communities in the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
CWM (F)指数反映了三峡库区各江段优势种摄食功能相关性状的变化趋势(附录2)。库首秭归江段的优势种以头长/吻长较大(4.47)、营养级较高(3.36)为主, 食性多为肉食性, 口位为下位口。库中云阳和库尾巴南江段的优势种都是以头长/吻长较小(3.32、2.85)、营养级中等(2.92、3.07)为主, 食性多为杂食性, 口位为下位口。香溪河下游峡口和嘉陵江下游合川江段都以头长/吻长中等(3.91、2.81)、营养级中等(2.78、3.34)为主, 食性多为杂食性, 口位为下位口为主。小江下游高阳江段的优势种以头长/吻长较小(3.71)、营养级较小(2.53)为主, 食性多为杂食性, 口位为端位口。其他摄食功能性状的CWM (F)指数见附录2。
2.3.2 运动功能多样性
三峡库区各江段鱼类群落运动功能丰富度的变化范围为0.02-0.07, 运动功能均匀度为0.40-0.56, 运动功能离散度为0.78-0.94 (图6)。运动功能丰富度在干流江段为库首秭归最高(0.07), 在支流江段为嘉陵江下游合川最低(0.02)。库中云阳江段的运动功能均匀度最高(0.56), 其他江段间相差不大。香溪河下游峡口江段的运动功能离散度最小(0.78), 其他江段相差不大。整体上库首秭归江段的运动功能多样性高, 嘉陵江下游合川江段的运动功能多样性最低。
图6
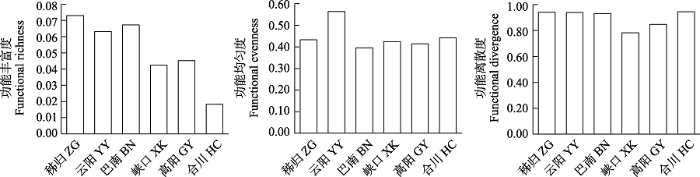
图6
三峡库区鱼类群落运动功能多样性空间变化
Fig. 6
Spatial variation of locomotion functional diversity of fish communities in the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
CWM (L)指数反映了三峡库区各江段优势种运动功能相关性状的变化趋势(附录3)。库首秭归江段的优势种以最大体长较大(30.63)、体长/尾柄高较小(2.47)为主, 生态类型为静水性, 体型多为侧扁形。库尾巴南江段的优势种以最大体长中等(27.75)、体长/尾柄高较大(6.30)为主, 生态类型为广适性, 体型多为圆筒形。库中云阳和香溪河下游峡口江段的优势种都是以最大体长中等(25.92、21.45)、体长/尾柄高较大(5.86、5.19)为主, 生态类型为静水性, 体型多为侧扁形。小江下游高阳江段的优势种以最大体长中等(20.80)、体长/尾柄高较大(5.19)为主, 生态类型为静水性, 体型多为侧扁形。嘉陵江下游合川江段的优势种以最大体长(17.74)较小、体长/尾柄较大(5.82)为主, 生态类型为广适性, 体型多为纺锤形。其他运动功能性状的CWM (L)指数见附录3。
2.3.3 繁殖功能多样性
三峡库区各江段鱼类群落繁殖功能丰富度的变化范围为0.01-0.12, 繁殖功能均匀度为0.43-0.51, 繁殖功能离散度为0.84-0.98 (图7)。繁殖功能丰富度和繁殖功能离散度指数在干流江段为: 秭归 > 云阳 > 巴南, 繁殖功能均匀度在干流江段为: 云阳 > 秭归 > 巴南, 表明库首秭归江段的繁殖功能多样性最高。支流嘉陵江下游合川江段的3个繁殖功能指数均较低(0.04、0.49、0.84), 其他江段的繁殖功能均匀度和繁殖功能离散度均相差不大, 表明嘉陵江合川江段的繁殖功能多样性最低。
图7
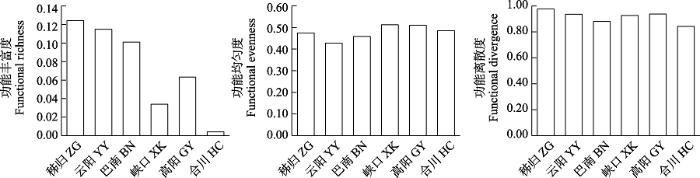
图7
三峡库区鱼类群落繁殖功能多样性空间变化
Fig. 7
Spatial variation of reproduction functional diversity of fish communities in the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
CWM (R)指数反映了三峡库区各江段优势种繁殖功能相关性状的变化趋势(附录4)。库首秭归、库中云阳和库尾巴南江段的优势种都以平均相对繁殖力较高(1,087.00、1,401.00、1,333.00)、初次性成熟体长较大(15.00、13.00、13.00)为主, 繁殖季节多为春季。不同的是产卵类型, 分别是沉性卵、漂流性卵和粘性卵。香溪河下游峡口和小江下游高阳江段优势种都以平均相对繁殖力较小(723.00、439.00)、初次性成熟体长较小(11.00、11.00)为主, 繁殖季节多为春季, 产漂流性卵。嘉陵江下游合川江段的优势种以相对繁殖力较小(283.00)、初次性成熟体长较小(9.00)为主, 繁殖季节多为夏季, 产粘性卵。其他繁殖功能性状的CWM (R)指数见附录4。
2.3.4 总功能多样性
三峡库区各江段鱼类群落总功能丰富度的变化范围为0.006-0.024, 总功能均匀度为0.35-0.49, 总功能离散度为0.80-0.92 (图8)。整体上看, 总功能多样性在干流江段为: 秭归 < 云阳 < 巴南, 在支流江段为: 香溪河下游峡口 > 小江下游高阳 > 嘉陵江下游合川。库尾巴南的总功能丰富度(0.02)和总功能离散度(0.83)均较高, 但是总功能均匀度(0.35)较低。嘉陵江下游合川江段的总功能丰富度极低(0.01), 功能均匀度(0.46)和功能离散度较高(0.92)。
图8
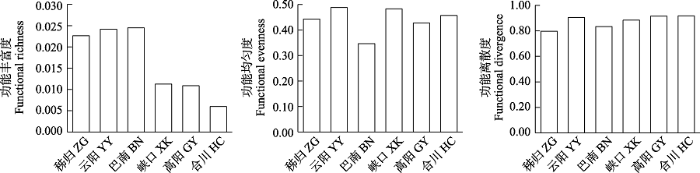
图8
三峡库区鱼类群落总功能多样性的空间变化
Fig. 8
Spatial variation of total functional diversity of fish communities in the Three Gorges Reservoir. ZG, Zigui; YY, Yunyang; BN, Banan; XK, Xiakou; GY, Gaoyang; HC, Hechuan.
2.4 功能多样性和物种多样性的关系
三峡库区各江段鱼类群落功能丰富度之间都呈显著正相关(n = 6, P < 0.05) (表3), 但是其他功能指数之间并无显著关系, 表明3种功能指数之间是相互独立的。总功能丰富度和Margalef丰富度指数呈极显著正相关(n = 6, P < 0.01), 摄食功能离散度和Pielou均匀度指数呈显著负相关(n = 6, P < 0.05), 繁殖功能均匀度和Margalef丰富度指数呈显著负相关(n = 6, P < 0.05), 其他指数之间并无显著性关系。总体上可以看出总功能多样性、摄食和繁殖相关的功能多样性和物种多样性之间存在一定的相关关系, 而运动相关的功能多样性和物种多样性之间相关关系不显著。
表3 不同多样性指数间的Pearson相关系数
Table 3
| Margalef | Shannon | Pielou | T-FRic | T-FEve | T-FDiv | F-FRic | F-FEve | F-FDiv | L-FRic | L-FEve | L-FDiv | R-FRic | R-FEve | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shannon | 0.66 | |||||||||||||
| Pielou | 0.28 | 0.90* | ||||||||||||
| T-FRic | 0.94** | 0.44 | 0.02 | |||||||||||
| T-FEve | -0.41 | -0.33 | -0.19 | -0.33 | ||||||||||
| T-FDiv | -0.51 | 0.18 | 0.52 | -0.65 | 0.44 | |||||||||
| F-FRic | 0.71 | 0.09 | -0.31 | 0.89* | -0.01 | -0.73 | ||||||||
| F-FEve | -0.13 | -0.32 | -0.38 | 0.10 | 0.34 | -0.37 | 0.49 | |||||||
| F-FDiv | -0.29 | -0.76 | -0.84* | 0.04 | 0.13 | -0.37 | 0.35 | 0.51 | ||||||
| L-FRic | 0.79 | 0.24 | -0.17 | 0.94** | -0.33 | -0.74 | 0.93** | 0.35 | 0.32 | |||||
| L-FEve | 0.33 | 0.28 | 0.19 | 0.32 | 0.62 | 0.33 | 0.31 | -0.13 | -0.16 | 0.14 | ||||
| L-FDiv | 0.52 | 0.07 | -0.17 | 0.45 | -0.25 | -0.27 | 0.23 | -0.66 | -0.07 | 0.25 | 0.33 | |||
| R-FRic | 0.79 | 0.22 | -0.19 | 0.95** | -0.26 | -0.64 | 0.91* | 0.20 | 0.34 | 0.97** | 0.30 | 0.41 | ||
| R-FEve | -0.85* | -0.55 | -0.24 | -0.76 | 0.07 | 0.19 | -0.54 | 0.39 | 0.33 | -0.53 | -0.70 | -0.75 | -0.64 | |
| R-FDiv | 0.12 | -0.31 | -0.51 | 0.43 | 0.28 | -0.41 | 0.73 | 0.76 | 0.77 | 0.66 | 0.19 | -0.23 | 0.64 | 0.01 |
*、**分别表示0.05和0.01水平上差异显著。FRic: 功能丰富度; FEve: 功能均匀度; FDiv: 功能离散度。T表示total, F表示food acquisition, L表示locomotion, R表示reproduction。
* and ** represent significant differences at the level of 0.05 and 0.01, respectively. FRic, Functional richness; FEve, Functional evenness; FDiv, Functional divergence. T is total, F is food acquisition, L is locomotion, R is reproduction.
3 讨论
3.1 鱼类群落结构
三峡库区蓄水前为自然流水生境, 流水性鱼类为各江段优势种。蓄水后库区生境变为兼有流水、缓流水和静水的复合栖息生境(邹家祥和翟红娟, 2016), 各江段鱼类生态类群组成存在差异。与库区第二次蓄水后的群落结构相比(Gao et al, 2010), 本次调查库中江段流水性鱼类数量百分比从1%增加到9%, 静水性鱼类从94%减少到37%, 广适性鱼类从5%增加到54%。库尾江段流水性鱼类数量百分比从85%减少到7%, 静水性鱼类从12%增加到15%, 广适性鱼类从3%增加到75%, 广适性鱼类由于适应能力强, 能够在生境转换中稳定存在, 因此在库区各江段中占有较大比例(杨志等, 2012)。蓄水后库区流水生境面积减少导致流水性鱼类种类和数量显著下降, 逐渐向上游的流水环境转移(魏念等, 2021)。库首江段由于离大坝最近, 水体为近似湖泊状, 营养物质的积累使得静水性的鱼类逐渐占据主导地位, 如库首秭归江段短颌鲚的数量百分比高达77.9%, 香溪河下游峡口江段贝氏䱗的数量百分比为32.1%。库中江段在三峡库区正常运行时为静水生境, 静水性鱼类比较多, 如库中云阳江段贝氏䱗的数量百分比为28.5%, 小江下游高阳江段为53.2%。库尾江段在三峡库区低水位运行时为流水生境, 能为流水性和产漂流性卵的鱼类提供适宜的生存环境(杨志等, 2015), 因此流水性的鱼类相对较多。
3.2 物种多样性
建坝前, 20世纪80年代, 三峡库区所在江段有127种鱼(丁瑞华, 1994)。建坝后, 2005-2006年在三峡库区调查到108种鱼(吴强等, 2007), 本次在三峡库区共调查到78种鱼类。中华鲟(Acipenser sinensis)、乐山小鳔鮈(Microphysogobio kiatingensi)、中华细鲫(Aphyocypris chinensis)、昆明裂腹鱼(Schizothorax grahami)等过去库区存在的鱼类此次并未采集到, 过去数量较多的胭脂鱼(Myxocyprinus asiaticus)、铜鱼(Coreius heterodon)、圆口铜鱼(C. guichenoti)等流水性鱼类数量锐减, 表明库区鱼类种数在下降, 这与大坝截流导致库区河湖分区引起的生境条件改变有关(李婷等, 2020)。Perera等(2014)表明2010-2011年三峡库区鱼类群落Shannon-Wiener指数为2.76-3.15, 高于本研究的1.08-2.68, 说明库区的Shannon-Wiener指数呈现下降的趋势。这可能是因为在三峡库区蓄水初期由于群落结构不稳定, 空缺的生态位空间和资源为外来物种入侵提供了条件, 使得物种多样性有所增加(魏念等, 2021)。但随着时间的增加, 入侵物种会影响物种组成和促进区域生物类群的同质化, 从而导致物种多样性降低(张登成和郑娇莉, 2019)。
本研究干流江段中物种多样性从库首到库尾逐渐升高, 这和Perera等(2014)的结果一致, 即在库区干流江段, 距离大坝越远鱼类群落物种多样性越高。库尾巴南江段距离三峡大坝634.4 km, 在库区低水位运行时为流水生境(杨志等, 2015)。库首秭归江段距离三峡大坝仅1.4 km, 为静水生境。有研究表明, 大坝修建前河相段的鱼类会向现存河相段聚集, 导致现存河相段鱼类种类增加(李婷等, 2020)。支流江段中香溪河下游峡口和小江下游高阳江段鱼类物种多样性较高可能是因为调查江段和库区干流之间是连通的, 群落之间的信息交流使得物种更为多样。而嘉陵江下游合川江段物种多样性最低是因为在合川江段下游建有草街水电站, 调查江段位于草街库区的库首, 水体为近似湖泊状, 较为单一, 因此物种多样性较低(刘扬扬等, 2014; 曾燏等, 2014)。
3.3 功能多样性
库首秭归江段的摄食、运动和繁殖功能多样性都最高, 表明群落有较高的稳定性(Sankaran & McNaughton, 1999)。CWM指数表明库首秭归江段群落的优势种为肉食性、静水性和平均相对繁殖力高的鱼类, 如短颌鲚等。功能多样性是由物种功能特征值和物种丰富度共同决定的(Lepš et al, 2006; Petchey & Gaston, 2006), 库首秭归江段的物种数较高, 表明这些鱼类能够占据各自的生态位空间, 充分利用群落中的资源, 保持群落的稳定(Villéger et al, 2008)。嘉陵江下游合川江段的摄食、运动和繁殖功能多样性都最低, 表明群落稳定性较低。嘉陵江下游合川江段物种数最少, 表明功能性状的范围比较小, 进而功能空间小、功能多样性低。CWM指数表明嘉陵江下游合川江段群落的优势种为杂食性、广适性和相对平均繁殖力低的鱼类, 如蛇鮈等。这些小型鱼类占据较小的生态位空间, 对群落资源利用率较低, 进而导致群落结构稳定性较低。总功能多样性在干流江段从库首到库尾逐渐升高, 这和Arantes等(2019)的研究结果类似, 鱼类群落遭受外界干扰的程度越深, 功能多样性受影响越深。自然流量的改变促使鱼类物种数减少, 可能会导致功能冗余增加, 降低功能多样性。
3.4 功能多样性和物种多样性关系
三峡库区鱼类群落各维度功能多样性指数间无显著相关关系, 表明各功能指数之间是相互独立的, 这和很多学者的研究一致(Mason et al, 2005; Parker et al, 2018), 功能多样性和物种多样性之间的关系在不同生态系统中存在差异。一般情况下, 功能丰富度和物种丰富度呈正相关关系(薛倩妮等, 2015), 如本研究中总功能丰富度和Margalef物种丰富度指数呈极显著正相关, 这是因为不存在具有完全相同功能特征的物种, 随着物种数量的增加, 功能特征的范围会增加, 功能空间也会增大(冯晨等, 2019)。而吴昊等(2020)研究表明在有强烈扰动的群落中, 物种功能属性趋于一致, 功能冗余增加导致功能丰富度和物种多样性呈负相关。陈静等(2018)的研究表明功能离散度和物种均匀度呈现正相关关系, 但本研究中摄食功能离散度和Pielou均匀度指数呈显著负相关, 这可能是由于物种均匀度会在一定程度上影响性状分布的均匀程度, 从而影响功能离散度。功能均匀度是衡量性状分布的均匀程度, 不受物种丰富度影响, 而本文中繁殖功能均匀度和Margalef物种丰富度指数呈显著负相关, 这可能是由于新增加的物种和原有的物种有着相似的功能性状, 导致群落的功能冗余过高, 使得群落物种性状分布不均匀(Luck et al, 2013; Parker et al, 2018)。运动相关的功能多样性和物种多样性之间没有显著相关关系可能是因为库区静水环境增多, 即使物种数增加, 也多为运动功能性状相似的物种, 因此对运动功能多样性影响不显著(赵小娜等, 2017)。
附录 Supplementary Material
附录1 三峡库区鱼类功能性状
Appendix 1 Functional traits of fishes in the Three Gorges Reservoir
附录2 三峡库区鱼类群落摄食功能群落特征加权平均数指数(CWMF)空间变化
Appendix 2 Spatial variation of community weighted mean index of the food acquisition function diversity of fish communities in the Three Gorges Reservoir
附录3 三峡库区鱼类群落运动功能群落特征加权平均数指数(CWML)空间变化
Appendix 3 Spatial variation of community weighted mean index of the locomotion functional diversity of fish communities in the Three Gorges Reservoir
附录4 三峡库区鱼类群落繁殖功能群落特征加权平均数指数(CWMR)空间变化
Appendix 4 Spatial variation of community weighted mean index of the reproduction functional diversity of fish communities in the Three Gorges Reservoir
参考文献
Impacts of hydroelectric dams on fishes and fisheries in tropical rivers through the lens of functional traits
Study of macroinvertebrate species and functional diversity in the New Xue River, Shandong Province, China
新薛河底栖动物物种多样性与功能多样性研究
Functional diversity: A new view point in the relationship between biodiversity and ecosystem functioning research
功能多样性——生物多样性与生态系统功能关系研究的新视角
Vive la différence: Plant functional diversity matters to ecosystem processes
DOI:10.1016/S0169-5347(01)02283-2 URL [本文引用: 1]
Fish resource status and biodiversity conservation in the main channel of Three Gorges Reservoir
三峡库区干流鱼类资源现状与物种多样性保护
Quantifying functional biodiversity
Functional diversity of fishes in the Minjiang Estuary, Southeast China
DOI:10.13287/j.1001-9332.201910.040
[本文引用: 1]

Based on samples collected by bottom trawl in May, August, November and December 2016 in the Minjiang Estuary, species richness and functional diversity of fishes, as well as their spatial-temporal variations, were quantified by a trait-based approach. The functional diversity varied with the changes of species richness. The species richness averaged across all stations were (17.8±5.1) species in spring, (22.4±5.6) in summer, (17.7±6.3) in autumn, and (12.1±2.3) in winter. Functional diversity also showed seasonal variation, with the highest in summer as (33.94±28.70), followed by (9.93±8.83) in winter, (11.30±7.55)<sup> </sup>in spring and (19.05±19.32)<sup> </sup>in autumn. The key functional species varied seasonally, as <i>Setipinna taty</i> contributing to 26% of the total functional value in spring, <i>Polydactylus sextarius </i>at 26% in summer, <i>Harpadon nehereus</i> climbing to 69% in autumn, and <i>Collichthys lucidus</i> at 38% in winter. The seasonal variations of fish species function were determined by life history and its interaction with other species.
闽江口鱼类功能多样性
DOI:10.13287/j.1001-9332.201910.040
[本文引用: 1]
根据2016年春季(5月)、夏季(8月)、秋季(11月)和冬季(2月)在闽江口海域进行底拖网作业的渔业资源调查样品,应用鱼类形态功能特征点数据分析了福建闽江口海域的鱼类功能多样性现状及时空变异.结果表明: 功能多样性与物种丰度具有相同的变化趋势,各站位鱼类物种丰度分别为:春季(17.8±5.1);夏季(22.4±5.6);秋季(17.7±6.3);冬季(12.1±2.3).鱼类集群功能多样性指数夏季最高,为(33.94±28.70),冬季最低(9.93±8.83),春季为(11.30±7.55),秋季为(19.05±19.32).群落内功能种呈现显著的季节演替,春季为太的黄鲫(占比为26%),夏季为六指马鲅(26%),秋季为龙头鱼,占比高达69%,显著高于其他物种的总和,冬季为棘头梅童鱼(38%).鱼类功能的季节性变动主要受其生活史及与其他物种之间的种间关系影响.
Interactive effects of nitrogen and phosphorus additions and different stand densities on soil microbial functional diversity of Acacia auriculiformis stands
氮磷添加和栽植密度对大叶相思林土壤微生物群落功能多样性的影响
Immediate impacts of the second impoundment on fish communities in the Three Gorges Reservoir
DOI:10.1007/s10641-009-9577-1 URL [本文引用: 1]
Effects of biodiversity on ecosystem functioning: A consensus of current knowledge
DOI:10.1890/04-0922 URL [本文引用: 1]
Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest
DOI:10.1038/s41598-019-43475-1
[本文引用: 1]
It is essential to understand how the loss of biodiversity impacts both ecosystem function (EF) and multifunctionality (EMF). Previous studies have mostly focused on predicting how species richness (SR) impacts EMF, while the effect of functional diversity (FD) on EMF remains unclear. Specifically, we know little about the primary functional drivers impacting EMF compared with SR. Therefore, we analysed 8 ecosystem functions within 58 natural secondary forest plots to investigate the effect of FD on both individual EF and EMF. Our results suggest that SR and FD had very significant positive effects on plant phosphorus, soil available phosphorus, and soil total nitrogen. FD explained significantly more variations in these functional responses than SR for individual ecosystem functioning. We also used a multiple threshold approach to test the effect of SR and FD on EMF. We found that FD and SR were positively related to EMF regardless of whether low-level function or high-level function was desired, but FD had a larger effect than SR. Based on the averaging approach, OLS regression, multivariate linear regression model and random forest analysis, we found that SR and FD were both drivers of EMF but that FD had a stronger effect and could explain more variation. As such, we conclude that FD drives ecosystem multifunctionality more than SR.
Assessing functional diversity in the field-methodology matters
Separating the chance effect from other diversity effects in the functioning of plant communities
DOI:10.1034/j.1600-0706.2001.920115.x URL [本文引用: 1]
Quantifying and interpreting functional diversity of natural communities: Practical considerations matter
Distribution characteristics and ecological types changes in fish communities under hydropower development from Xiluodu to Xiangjiaba reach
水电开发对鱼类种群分布及生态类型变化的影响——以溪洛渡至向家坝河段为例
Fish assemblage structure and age and growth of the primary fish populations in the Three Gorges Reservoir
三峡水库鱼类群落结构及主要鱼类种群年龄与生长
Biodiversity decline of fish assemblages after the impoundment of the Three Gorges Dam in the Yangtze River Basin, China
DOI:10.1007/s11160-019-09548-0 [本文引用: 1]
Impacts of hydropower development on hydrological regime in mainstream of mid-lower Jialing River
嘉陵江中下游干流水电开发对水文情势的影响
Changes in bird functional diversity across multiple land uses: Interpretations of functional redundancy depend on functional group identity
DOI:10.1371/journal.pone.0063671 URL [本文引用: 1]
Functional richness, functional evenness and functional divergence: The primary components of functional diversity
DOI:10.1111/oik.2005.111.issue-1 URL [本文引用: 1]
Biodiversity-ecosystem functioning relationships in fish communities:Biomass is related to evenness and the environment, not to species richness
Changes in plant functional diversity and environmental factors of Cyclobalanopsis glauca community in response to slope gradient in Karst hills, Guilin
桂林岩溶石山青冈群落植物功能多样性和环境因子与坡向的关联研究
Large river fish functional diversity responses to improved water quality over a 28 year period
DOI:10.1016/j.ecolind.2018.01.035 URL [本文引用: 2]
Effect of the distance from the dam on river fish community structure and compositional trends, with reference to the Three Gorges Dam. Yangtze River, China
Functional diversity: Back to basics and looking forward
DOI:10.1111/j.1461-0248.2006.00924.x
PMID:16706917
[本文引用: 1]
Functional diversity is a component of biodiversity that generally concerns the range of things that organisms do in communities and ecosystems. Here, we review how functional diversity can explain and predict the impact of organisms on ecosystems and thereby provide a mechanistic link between the two. Critical points in developing predictive measures of functional diversity are the choice of functional traits with which organisms are distinguished, how the diversity of that trait information is summarized into a measure of functional diversity, and that the measures of functional diversity are validated through quantitative analyses and experimental tests. There is a vast amount of trait information available for plant species and a substantial amount for animals. Choosing which traits to include in a particular measure of functional diversity will depend on the specific aims of a particular study. Quantitative methods for choosing traits and for assigning weighting to traits are being developed, but need much more work before we can be confident about trait choice. The number of ways of measuring functional diversity is growing rapidly. We divide them into four main groups. The first, the number of functional groups or types, has significant problems and researchers are more frequently using measures that do not require species to be grouped. Of these, some measure diversity by summarizing distances between species in trait space, some by estimating the size of the dendrogram required to describe the difference, and some include information about species' abundances. We show some new and important differences between these, as well as what they indicate about the responses of assemblages to loss of individuals. There is good experimental and analytical evidence that functional diversity can provide a link between organisms and ecosystems but greater validation of measures is required. We suggest that non-significant results have a range of alternate explanations that do not necessarily contradict positive effects of functional diversity. Finally, we suggest areas for development of techniques used to measure functional diversity, highlight some exciting questions that are being addressed using ideas about functional diversity, and suggest some directions for novel research.
Determinants of biodiversity regulate compositional stability of communities
DOI:10.1038/44368 URL [本文引用: 1]
Functional diversity of freshwater fishes and methods of measurement
淡水鱼类功能多样性及其研究方法
Beyond classic ecological assessment: The use of functional indices to indicate fish assemblages sensitivity to human disturbance in estuaries
DOI:10.1016/j.scitotenv.2018.05.179 URL [本文引用: 1]
Functional diversity
DOI:10.1016/B0-12-226865-2/00132-2 URL [本文引用: 1]
Functional ecology of fish: Current approaches and future challenges
DOI:10.1007/s00027-017-0546-z URL [本文引用: 1]
New multidimensional functional diversity indices for a multifaceted framework in functional ecology
DOI:10.1890/07-1206.1
PMID:18724739
[本文引用: 4]
Functional diversity is increasingly identified as an important driver of ecosystem functioning. Various indices have been proposed to measure the functional diversity of a community, but there is still no consensus on which are most suitable. Indeed, none of the existing indices meets all the criteria required for general use. The main criteria are that they must be designed to deal with several traits, take into account abundances, and measure all the facets of functional diversity. Here we propose three indices to quantify each facet of functional diversity for a community with species distributed in a multidimensional functional space: functional richness (volume of the functional space occupied by the community), functional evenness (regularity of the distribution of abundance in this volume), and functional divergence (divergence in the distribution of abundance in this volume). Functional richness is estimated using the existing convex hull volume index. The new functional evenness index is based on the minimum spanning tree which links all the species in the multidimensional functional space. Then this new index quantifies the regularity with which species abundances are distributed along the spanning tree. Functional divergence is measured using a novel index which quantifies how species diverge in their distances (weighted by their abundance) from the center of gravity in the functional space. We show that none of the indices meets all the criteria required for a functional diversity index, but instead we show that the set of three complementary indices meets these criteria. Through simulations of artificial data sets, we demonstrate that functional divergence and functional evenness are independent of species richness and that the three functional diversity indices are independent of each other. Overall, our study suggests that decomposition of functional diversity into its three primary components provides a meaningful framework for its quantification and for the classification of existing functional diversity indices. This decomposition has the potential to shed light on the role of biodiversity on ecosystem functioning and on the influence of biotic and abiotic filters on the structure of species communities. Finally, we propose a general framework for applying these three functional diversity indices.
Current status and changes in fish assemblages in the Three Gorges Reservoir
三峡库区鱼类群落结构现状及变化
Relationships between functional diversity and species diversity of pine-oak mixed forest in Qinling Mountains and their environmental explanations
秦岭松栎林功能多样性与物种多样性和环境异质性的耦合关系
Studies on fishery resources in the Three Gorges Reservoir of the Yangtze River
长江三峡库区蓄水后鱼类资源现状
Functional diversity research of tree and shrub layers in forest communities of the Wulu Mountains Nature Reserve in Shanxi, China
山西五鹿山森林群落木本植物功能多样性
The current situation of fish resources in the Daning River after the impoundment of the Three Gorges Reservoir
三峡库区蓄水后大宁河鱼类资源现状研究
The spatial-temporal distribution characteristics of the endemic fish in the upper reaches of the Yangtze River under the normal operation of the Three Gorges Reservoir
正常运行条件下三峡库区干流长江上游特有鱼类时空分布特征研究
Spatiotemporal patterns of fish community structures in the Three Gorges Reservoir and its upstream during the 175-m-deep impoundment
三峡水库175 m试验性蓄水期库区及其上游江段鱼类群落结构时空分布格局
Research on fish resources variation and protection in reservoir area of TGP after its operation
三峡水库运行后库区鱼类资源变化及保护研究
Utilization and protection status of fish resources in Jialing River
嘉陵江鱼类资源利用与保护现状
Preliminary study on invasion of alien fish species after construction of hydropower projects
水电工程建设前后外来鱼类入侵问题初步研究
Diversity of forest communities in the upstream and middle reaches of the Wenyu River watershed, Shanxi
山西文峪河上中游森林群落多样性

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}