



生物地理区的差异是长久分离或无法来往的许多障碍直接造成的, 广阔的海洋与巨大的温差是陆栖生命形式扩散的最大障碍。——华莱士《马来群岛自然考察记》第39章
生物多样性的空间分布及其决定因素是生物地理学与宏生态学(macroecology)研究的核心内容(Lomolino et al, 2017)。岛屿因具有明确的地理边界, 是检验群落生态学四个核心过程(选择、扩散、漂变与成种)如何共同构建不同尺度生物多样性格局的重要平台(Vellend, 2010, 2016, 2020)。在MacArthur和Wilson (1967)提出的岛屿生物地理学理论中, 岛屿物种丰富度由灭绝率和迁入率的平衡决定, 灭绝率和迁入率与岛屿属性(面积和隔离度)紧密相关。面积大的岛屿可容纳更多的生物类群, 具有较低的灭绝率, 即丰富度与面积正相关; 偏远的岛屿由于物种很难通过大陆或大岛扩散, 具有较低的迁入率, 即丰富度与隔离度负相关(阎恩荣等, 2022)。以往研究表明岛屿面积对植物多样性有强烈影响(Yu et al, 2012; 谢艳秋等, 2023; Xu et al, 2023)。Kreft等(2008)对全球488个岛屿植物丰富度的分析发现, 在不同的生物地理区和气候区, 面积是影响岛屿物种丰富度最主要的因子。隔离度对岛屿植物丰富度的影响与岛屿类型有关, 对大陆性岛屿的影响小于海洋性岛屿(Weigelt & Kreft, 2013)。隔离度对不同植物功能型组(生长型、叶物候型)丰富度的影响也存在差异。例如, Schrader等(2020)在全球17个群岛700个岛屿和Liu等(2023)在浙江温州三垟湿地35个岛屿的研究都发现草本植物比木本植物拥有更强的扩散能力, 更易到达偏远的岛屿, 受隔离度的影响小于木本植物。另外, 岛屿形状越不规则, 可能包含更多的生境种类, 因而包括更高的生物多样性(Honnay et al, 2003; Triantis & Sfenthourakis, 2012)。
环境过滤对岛屿物种丰富度格局也有重要影响(Carvajal-Endara et al, 2017)。当多个物种迁入或扩散到岛屿后, 只有少数能够适应当地环境的物种才能成功定殖(Gillespie et al, 2012)。Carvajal-Endara等(2017)基于加拉帕戈斯群岛本土植物的研究发现环境过滤对植物多样性有强烈的影响, 且强于扩散限制的影响。Liu等(2020)基于浙江千岛湖29个岛屿木本植物的研究发现在控制面积与隔离度的影响后, 环境过滤显著影响木本植物丰富度。温度、降水等气候因子通过限制物种定殖的成功率影响岛屿生物多样性, 已被证实是岛屿生物多样性的关键驱动因素。在全球尺度上, 温度及温度季节性是决定岛屿植物丰富度格局的重要因子(Kreft et al, 2008; Cabral et al, 2014)。在区域尺度上, Kubota等(2015)发现温度是影响日本列岛及琉球群岛植物丰富度的重要因子, Valli等(2019)发现年降水量是影响希腊爱奥尼亚群岛17个岛屿植物丰富度的重要因子。同时, 物种对环境的适应能力与自身的功能性状有关, 环境过滤有助于筛选出具有相似性状的物种在岛上定殖(Schrader et al, 2020)。例如, 草本植物个体小、生命周期短, 比起木本植物可能更易在资源匮乏、环境条件不稳定的岛屿定殖(Schrader et al, 2020); 落叶阔叶树种通过在低温环境下脱落叶片, 减少蒸腾作用与呼吸作用, 与常绿阔叶树种相比更能适应低温环境(Givnish, 2002)。
随着全球交通网络的急速扩张以及城市化导致的人口迁移, 人类活动也正在重塑岛屿生物多样性格局。人类活动影响了物种的选择与扩散等多个过程, 改变生物群落的组成和结构, 愈来愈被认为是影响岛屿物种丰富度的重要因素(Helmus et al, 2014; Matthews & Triantis, 2021; Gleditsch et al, 2023)。随着人类对岛屿的交通建设, 岛屿的连通性增加, 隔离程度降低, 促进了本土和外来物种从大陆扩散到岛屿以及在群岛内的扩散(Helmus et al, 2014)。同时, 人类破坏岛屿原有生境, 造成生境丧失, 对本土物种生存造成巨大威胁(Russell & Kueffer, 2019)。另外, 人类干扰对不同生长型的植物多样性影响可能存在差异。Liu等(2023)在受人类干扰的浙江温州三垟湿地岛屿发现, 道路面积占比是影响本土草本植物丰富度的重要因子, 但对本土木本植物丰富度影响很小。因此, 人类干扰与岛屿的地理位置、面积等多种因素有关, 应该包括在岛屿生物地理学研究中(Helmus et al, 2014)。
舟山群岛位于亚热带季风气候区, 是我国第一大群岛, 具有广泛的面积、隔离度、气候与人类干扰等梯度, 是研究人类世背景下的岛屿生物多样性格局的理想系统。近年来, 基于舟山群岛生物多样性的研究表明, 岛屿面积是影响木本植物、苔藓、蝴蝶、土壤细菌丰富度的重要因素, 隔离度是影响木本植物和土壤细菌丰富度、维管植物β多样性的重要因素(刘翔宇等, 2019; Yu et al, 2019; Chen et al, 2021; Xu et al, 2023)。Xu等(2023)基于舟山37个岛屿木本植物与土壤细菌多样性的研究发现, 岛屿面积与隔离度可以通过影响土地利用强度与松材线虫入侵间接影响木本植物与土壤细菌的多样性。以往关于舟山植物多样性的研究包括的岛屿数量较少, 且多基于样方或样线获得的岛屿尺度的不完整数据, 较少考虑岛屿属性、气候与人类活动对植物多样性的综合影响。舟山群岛属于亚热带常绿阔叶林区, 虽然常绿阔叶林是其地带性植被, 但由于舟山各岛屿存在较强的环境过滤和人类干扰, 常绿阔叶与落叶阔叶植物在各岛屿上呈不同比例的分布。常绿阔叶与落叶阔叶代表不同的功能性状, 对气候因子的响应存在差异(Ge & Xie, 2017)。本文以舟山群岛的92个岛屿为研究对象, 利用岛屿尺度的本土种子植物名录数据, 按生长型划分为乔木、灌木和草本, 按叶物候型分为常绿阔叶和落叶阔叶木本植物, 探究以下3个科学问题: (1)舟山群岛植物丰富度呈现怎样的分布格局? 不同生长型的植物丰富度分布格局是否存在差异? (2)岛屿属性、气候与人类活动等如何影响植物丰富度? (3)常绿阔叶木本植物的丰富度比率主要受哪些环境因子影响?
1 材料与方法
1.1 研究区概况
舟山群岛位于长江口以南、杭州湾外缘的东海海域, 是我国第一大群岛, 在7,000-9,000年前的海侵事件中与大陆分离, 属于大陆性岛屿。舟山群岛包含500 m2以上的岛屿1,339个, 大岛人口较多, 小岛多为无居民海岛(中国海岛志编纂委员会, 2014)。其中, 舟山本岛(515.4 km2)是面积最大的岛屿。舟山群岛处于亚热带季风气候区, 四季分明, 受季风、台风影响显著。植被以针叶林、阔叶林、灌丛和草丛为主。自明末清初起, 舟山群岛开始经历了较大程度的人为活动(中国海岛志编纂委员会, 2014)。近些年来, 舟山群岛密集建设航道、口岸, 实施“连岛工程” “小岛迁大岛建”工程, 经济的发展及政策的实行促使舟山群岛人为干扰愈加严重, 土地利用发生转变。
图1
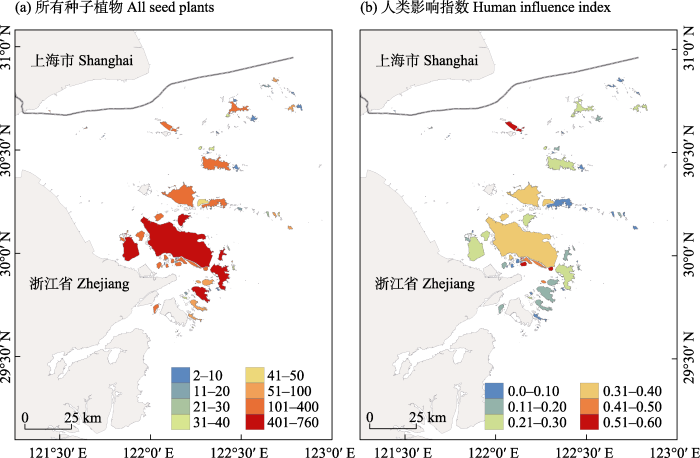
图1
舟山群岛92个岛屿种子植物丰富度(a)和人类影响指数(b)的空间分布
Fig. 1
Spatial distributions of all seed plant richness (a) and human influence index (b) of 92 islands in the Zhoushan Archipelago
1.2 植物名录数据
基于舟山群岛已有研究工作, 我们建立了一个全面的植物分布数据库。数据库主要由实地调查资料和文献资料组成, 标本数据与其他网络公开数据等作为补充。实地调查资料包括2012‒2013年基于样线法、样方法等获得的植物调查记录(毕玉科, 2014①(① 毕玉科 (2014) 舟山岛外来植物及其入侵性分析. 硕士学位论文, 华东师范大学, 上海.); 高浩杰等, 2015)、2014‒2020年基于样方法获得的植物数据(石娇星, 2021②(② 石娇星 (2021) 舟山群岛植被分类与制图. 硕士学位论文, 华东师范大学, 上海.))和舟山市植物资源调查记录(库伟鹏未发表资料)。文献资料包括舟山多个岛屿植物研究中用到的数据(朱弘等, 2015; 魏永杰等, 2016; 王国明和叶波, 2017; 郑俊鸣等, 2017; 高浩杰, 2018; 刘翔宇等, 2019; 朱峻熠等, 2020), 占总数据的2%。标本数据来自中国国家标本资源平台(NSII, 2022)在舟山群岛的6,000余条记录, 标本采集时间为1917‒2018年, 其中90%以上的标本数据为1980年之后采集。网络公开数据包括GBIF (2022)数据(26条)、中国自然标本馆(
物种名标准化使用R包U.Taxonstand (Zhang & Qian, 2023), 基于World Flora Online数据库(WFO, 2023)进行匹配, 并参考iPlant数据库相关资料(
1.3 环境数据
本研究包括岛屿属性、气候和人类干扰3类环境因子(附录2)。岛屿属性包括面积(area)、距大陆距离(distance to mainland, DM)和形状指数(shape index, SI) 3个因子。岛屿面积和距大陆距离参考Wang等(2023)的计算方法, 使用UTM (universal transverse Mercator)投影。岛屿面积、周长使用ESRI ArcMap 10.4计算; 距大陆距离指岛屿到大陆海岸线的最短距离。形状指数表示岛屿形状的复杂程度, 通过岛屿面积和周长计算(SI = C/[2 × (π × A)0.5])。
气候因子包括年均温(mean annual air temperature, MAT)、温度季节性(temperature seasonality, TS)、年降水量(mean annual precipitation, MAP)和降水季节性(precipitation seasonality, PS)。气候数据来自CHELSA (Climatologies at High Resolution for the Earth’s Land Surface Areas)的1981-2010年的30 弧秒气候数据(Karger et al, 2021)。人类影响指数(human influence index, HI)用于衡量人类对岛屿的改造和利用, 本研究使用建筑与耕地面积占岛屿陆地面积的比值来代表。土地利用数据产品来自ESA (European Space Agency)的WorldCover产品(Zanaga et al, 2022), 该产品提供的10 m分辨率的土地利用类型信息可精准覆盖本研究中大部分岛屿(4个小岛的土地利用类型信息进行手工校正)。
1.4 数据分析
采用一般线性回归探究单一环境因子与植物丰富度之间的关系。为提高一般线性回归的拟合优度, 对植物丰富度、面积、距大陆距离进行以10为底的对数转换。为比较不同环境因子对丰富度的影响, 本文采用广义线性模型(伪泊松分布)对丰富度构建多元回归模型; 采用beta回归(Cribari-Neto & Zeileis, 2010)对常绿阔叶木本植物丰富度比率构建多元回归模型。首先, 对面积、距大陆距离进行以10为底的对数转换以提高模型拟合度, 再对所有环境因子进行标准化。然后, 使用Spearman秩相关检验以上8个因子间的相关性(附录3); 对于相关系数大于0.7的因子, 在模型中增加交互效应以减少共线性的影响。同时, 我们也基于方差膨胀因子(variance inflation factor, VIF)检测了环境因子之间的共线性, 8个环境因子的VIF均小于10。最后, 使用R包stats中的glm方程和R包betareg中的betareg方程分别对丰富度和丰富度比率构建全模型。采用全子集回归得到所有可能的模型, 根据AICc (Akaike’s second-order corrected information criterion)值选择最优模型(Burnham & Anderson, 2002)。若ΔAICc > 2, ΔAICc值为0的模型为最优模型; 若ΔAICc ≤ 2, 则对ΔAICc ≤ 2的模型采用模型平均。模型选择和模型平均使用R包MuMIn完成(Bartoń, 2023)。以上数据分析在R 4.2.2中完成(R Core Team, 2022)。
2 结果
2.1 岛屿植物多样性丰富度的生物地理格局
舟山92个岛屿共记录了本土种子植物129科558属1,158种。所有植物在92座岛屿上的丰富度为2‒760, 面积大的岛屿丰富度更高; 沿着靠近大陆的西南到远离大陆的东北方向, 丰富度逐渐减少(图1)。
在92个岛屿中, 共有乔木108种, 灌木318种, 草本732种。乔木以壳斗科(12种)、樟科(7种)和蔷薇科(7种)为主, 灌木以蔷薇科(33种)、豆科(20种)和唇形科(17种)为主, 草本以禾本科(100种)、菊科(84种)和莎草科(79种)为主。3种不同生长型的种子植物丰富度格局与所有植物相似, 随岛屿面积增加而增加、随距大陆距离增加而降低。乔木丰富度随距大陆距离增加而减少的趋势比灌木和草本更加明显(附录4)。
图2
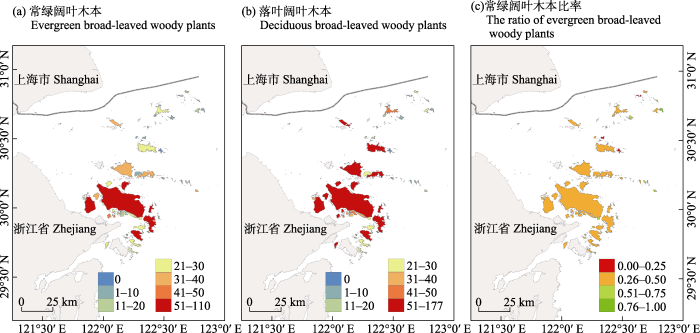
图2
舟山群岛92个岛屿木本植物丰富度的空间分布格局。(a)常绿阔叶木本; (b)落叶阔叶木本; (c)常绿阔叶木本占总阔叶木本植物的比率。
Fig. 2
Spatial distributions of woody plant richness in 92 islands of the Zhoushan Archipelago. (a) Evergreen broad-leaved woody plants; (b) Deciduous broad-leaved woody plants; (c) The ratio of evergreen broad-leaved woody plant richness to all broad-leaved woody plant richness.
2.2 岛屿植物丰富度的影响因素
图3
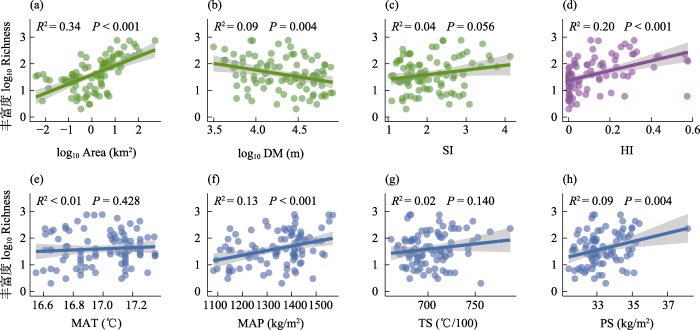
图3
植物丰富度与环境因子的相关性。(a)面积(Area); (b)距大陆距离(DM); (c)岛屿形状指数(SI); (d)人类影响指数(HI); (e)年均温(MAT); (f)年降水量(MAP); (g)温度季节性(TS); (h)降水季节性(PS)。
Fig. 3
Correlations between all seed plant richness and environmental drivers. (a) Area; (b) The distance to mainland (DM); (c) Shape index (SI); (d) Human influence index (HI); (e) Mean annual air temperature (MAT); (f) Mean annual precipitation (MAP); (g) Temperature seasonality (TS); and (h) Precipitation seasonality (PS).
图4
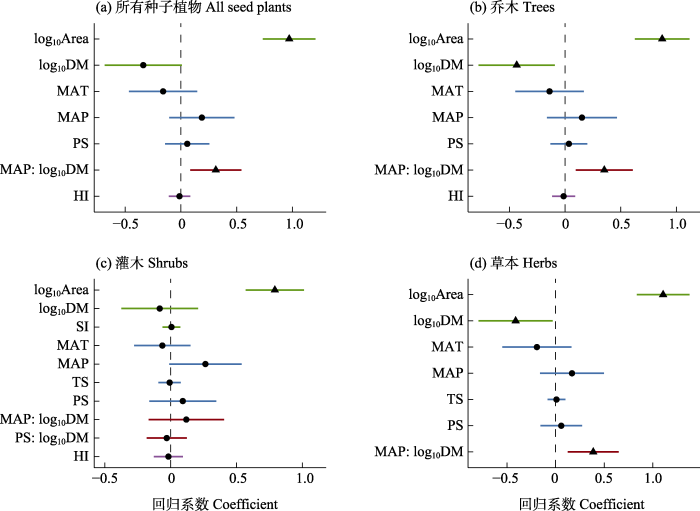
图4
岛屿属性、气候与人类影响对本土植物丰富度的影响。(a)所有种子植物; (b)乔木; (c)灌木; (d)草本。直线代
Fig. 4
Effects of island physical characteristics, climate and human influence on native seed plant richness. (a) All seed plants; (b) Trees; (c) Shrubs; and (d) Herbs. The straight line represents the 95% confidence interval. The left side of the vertical dashed line indicates for negative correlations, while the right for positive correlations. The triangles indicate statistically significant estimates of standardized coefficients, and the dot points show the non-significant ones. Island characteristics, climate, human influence and their interactions are represented in green, blue, purple and red, respectively. Variable abbreviations are the same in
两种木本植物叶物候型的丰富度与环境因子的关系与总丰富度‒环境关系基本一致(附录8, 9), 落叶阔叶木本植物丰富度呈现出与温度季节性更强的关系(P < 0.05), 而常绿阔叶木本比率仅与温度季节性显著负相关(P < 0.001), 与其他环境因子无显著相关(附录10)。多元回归结果表明, 常绿阔叶木本植物丰富度受面积影响最大, 年降水量影响次之; 落叶阔叶木本植物丰富度受面积影响最大, 距大陆距离影响次之; 常绿阔叶木本比率受温度季节性影响显著, 其他因子影响都较小(图5)。
图5
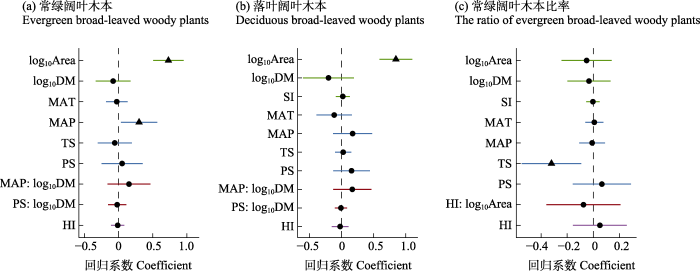
图5
岛屿属性、气候与人类影响对木本植物丰富度的影响。(a)常绿阔叶木本; (b)落叶阔叶木本; (c)常绿阔叶木本占总阔叶木本植物的比率。直线代
Fig. 5
Effects of island physical characteristics, climate and human influence on woody plant richness. (a) Evergreen broad-leaved woody plants; (b) Deciduous broad-leaved woody plants; and (c) The ratio of evergreen broad-leaved woody plant richness to all broad-leaved woody plant richness. The straight line represents the 95% confidence interval. The left side of the vertical dashed line indicates negative correlations, while the right for positive correlations. The triangles indicate statistically significant estimates of standardized coefficients, and the dot points show the non-significant ones. Island characteristics, climate, human influence and their interactions are represented in green, blue, purple and red, respectively. Variable abbreviations are the same in
3 讨论
3.1 岛屿属性对植物丰富度的影响
岛屿生物地理学理论认为物种丰富度与岛屿面积正相关, 与隔离度负相关(MacArthur & Wilson, 1967)。很多研究为这一经典理论提供了数据支持(如: Kreft et al, 2008; Storch et al, 2012; Cabral et al, 2014)。在本研究中, 我们发现在受到人类强烈干扰的岛屿系统中, 岛屿生物地理学理论所预测的格局仍然稳固。本土植物总丰富度随岛屿面积增加而显著增加, 随隔离度增加显著降低。这一结果在舟山群岛未考虑人类影响的鸟类多样性研究中得到支持(Wang et al, 2023)。综合考虑气候和人类影响因子, 面积和隔离度仍是影响最大的因子。此结果在华东地区考虑人类影响的研究中得到支持, 如舟山群岛已有研究(包括木本植物、土壤细菌、苔藓) (Yu et al, 2019; Xu et al, 2023)和浙江温州三垟湿地岛屿植物多样性研究(Liu et al, 2023)。与隔离度相比, 面积对植物丰富度贡献更大。这一结果可能是由于岛屿隔离程度对很多植物扩散的限制较小, 同时舟山群岛较强的人类活动有助于部分植物扩散, 从而削弱了隔离度对植物丰富度的影响。以往研究也证明面积对岛屿植物丰富度的影响强于隔离度(Kreft et al, 2008), 面积还可以通过影响生境异质性间接影响植物丰富度(Walentowitz et al, 2023), 因此, 在舟山群岛应加强对面积较大岛屿的保护, 通过维持多样的生境和减少人类干扰来保护更多的本土物种。
岛屿面积与隔离度对不同生长型、叶物候型植物丰富度都有显著影响, 但是对不同功能型组的影响程度存在差异。在舟山群岛, 隔离度对乔木丰富度的影响强于草本和灌木。基于我国东部海岛维管植物扩散方式的统计表明, 依靠风力与鸟类传播种子的方式在草本植物中的比例显著高于在木本植物中的比例(刘翔宇等, 2019)。相比于草本, 很多乔木的扩散能力更弱, 更易受隔离度影响。另外, 常绿阔叶木本比率受面积、隔离度的影响很小, 主要原因可能与舟山群岛的起源历史有关。舟山群岛在7,000-9,000年前由于海平面上升而形成, 形成时间较短, 很多常绿阔叶植物作为海平面上升前的地带性植被的代表性植物仍分布在很多岛屿上。值得注意的是, 基于小岛屿效应(small-island effect), 面积并非是影响小岛屿上植物丰富度的主要因子, 即当面积低于某个阈值时, 物种数不随岛屿面积的增加而增加(Lomolino, 2000)。小岛屿效应在很多岛屿系统中广泛存在(Wang et al, 2018), 其对舟山群岛植物多样性的影响值得进一步深入研究。
岛屿形状越复杂, 岛屿包含的生境种类和数量可能越多(Yu et al, 2012), 可以支持更多的物种丰富度。在舟山群岛, 岛屿形状指数对种子植物及不同生长型、叶物候型植物丰富度的影响大多较弱。首先, 舟山群岛的岛屿形状指数的变异较小(本研究92个岛屿有51个岛屿的形状指数小于2)。其次, 本研究选取的岛屿形状指数更多代表岛屿的边缘生境的复杂程度, 不能表征岛屿内部的生境的数量和类型(Hortal et al, 2009; Triantis & Sfenthourakis, 2012)。在舟山群岛, 仅草本植物丰富度随岛屿形状指数增加而显著增加, 这可能与很多草本植物适应环境条件恶劣的岛屿边缘生境(如滨海滩地、近海山地)有关。岛屿的边缘生境为滨海植物(如: 滨海前胡(Peucedanum japonicum),提供了生存场所, 这类生境在岛屿开发过程中需得到重视和保护。
3.2 气候对岛屿植物丰富度的影响
气候因子是影响很多岛屿物种多样性的关键因子, 可以通过限制物种多度和定殖成功率限制岛屿物种多样性(Kreft et al, 2008)。本研究表明植物总丰富度与年均温间的关系并不显著, 而与年降水量显著正相关, 这与以往很多研究并不完全一致。例如, Kreft等(2008)基于全球488个岛屿植物多样性的研究表明, 年均温与年降水量对植物丰富度具有较强的影响; Blackburn等(2016)基于全球海洋岛屿植物多样性的研究表明, 年均温与本土植物丰富度有强的正相关。在舟山群岛, 年均温对植物总丰富度影响不显著可能与年均温的范围较小(16.5‒17.4℃)相关。与之相比, 年降水量对植物总丰富度具有强烈影响, 这可能是由于距大陆距离和台风的过境路线造成了明显的降水梯度。
不同生长型的植物丰富度对年降水量的响应存在差异, 乔木、灌木的响应比草本更明显。这与它们的生存策略相关, 很多草本植物采取资源获取型生存策略, 环境生态位较宽, 而很多木本植物采取资源保守型生存策略, 更易在资源稳定环境中生存, 降水量增加明显促进木本植物的生长(Pierce et al, 2017; Šímová et al, 2018)。对于常绿与落叶两种叶物候型, 降水量对常绿阔叶木本植物丰富度的影响强于落叶阔叶木本, 这与亚热带地区常绿阔叶木本与落叶阔叶木本植物在水分利用策略上的差异有关, 落叶树种具有更高的茎水分利用效率, 常绿树种具有更强的抗木质部栓塞能力(Fu et al, 2012)。Ge和Xie (2017)对中国亚热带常绿阔叶林的研究也发现降水量增加可以提高常绿阔叶木本植物在群落中的重要性。
在亚热带地区, 气候存在着明显的季节性变化(宋永昌, 2013)。关于目前亚热带地区岛屿生物多样性的研究, 大多仅考虑年均温和年降水量(Yu et al, 2019; 谢艳秋等, 2023), 忽视了气候季节性变化对多样性的潜在影响(Weigelt et al, 2013; Cabral et al, 2014)。在舟山群岛, 我们发现降水季节性与植物总丰富度显著正相关。这可能由于随着降水季节性增强, 岛屿气候情况更接近大陆的气候情况, 适合更多植物定殖(Weigelt et al, 2013)。气候季节性对植物多样性的影响在叶物候上表现更强。温度季节性对常绿阔叶木本比率的影响最强且显著, 这与Ge等(2019)在亚热带常绿阔叶林的研究结果一致。植物对温度季节性的响应也反映了植物抵御寒冷天气的能力。常绿木本的耐寒性弱于落叶木本(Harrison et al, 2010; Ge & Xie, 2017), 所以落叶阔叶木本植物丰富度随温度季节性增强而增加, 常绿阔叶木本比率随之减小。综上, 不同功能型组植物具有不同的气候需求, 需通过长期监测才能深入了解岛屿植物如何响应与适应气候变化, 进而提出针对性的保护措施。
3.3 人类活动对岛屿植物丰富度的影响
在舟山群岛, 随着人类干扰强度增加, 种子植物及不同生长型、叶物候型植物的丰富度显著增加。然而, 当同时考虑其他因子时, 人类干扰的影响并不突出。首先, 人类干扰与岛屿面积之间存在非常强的非线性正相关(附录3, 附录11)。一般来说, 面积大的岛屿一般交通更便利, 拥有更丰富的淡水与食物等资源, 更易被人类开发利用(Jesse et al, 2018)。当同时考虑不同岛屿属性和气候因子时, 人类活动与丰富度间的关系变弱, 可能受岛屿面积和人类活动共线性关系的影响(附录3)。值得注意的是, 人类影响是一个复杂的因子, 虽然与面积显著相关, 但并非完全依赖于面积。例如, 人们对一些岛屿存在特别需求(如工业发展、旅游发展等)的情况下, 人类对岛屿的利用强度则与面积无关。在舟山群岛, 鲁家峙(2000年开始密集性城市建设)、岙山(1989年开始建造油库)、小洋山(2002年开始设立港口)等面积较小的岛屿存在非常强的人类活动, 很大程度上重塑着这些岛屿以及邻近岛屿的植物组成。其次, 本研究使用的植物名录数据是舟山群岛近几十年来累积的结果, 长时间尺度的植物数据可能会掩盖植物丰富度随时间的变化及对人类活动的响应, 尽管数据主要集中在近20年, 仍会减弱或混淆人类活动与多样性之间的关系。在舟山群岛的人类活动(如: 大岛建、小岛迁、陆岛连等工程)可以在短期内改变植物的分布与数量, 而这样的变化基于目前使用的低时空精度的人类干扰和植物名录数据是无法捕捉到的。另外, 人类活动(以及气候等因子)对植物多样性的影响很大程度上并非影响物种在岛屿上存在与否, 而对物种多度、均匀度等的影响更大。当前考虑人类活动对岛屿生物多样性影响的研究工作多仅评估了物种丰富度(如: Walentowitz et al, 2022; Gleditsch et al, 2023)。因此, 评估人类活动、气候变化、岛屿属性等对岛屿生物多样性的综合影响, 需获取更高时空分辨率的物种分布与人类活动等数据, 全面评估不同生物多样性指标的影响, 最终才能全面理解人类世背景下的岛屿生物多样性的变化规律(Helmus et al, 2014)。
3.4 小结
综上, 基于我国第一大群岛舟山群岛92个岛屿的种子植物数据, 我们发现岛屿面积、年降水量、温度季节性等是影响舟山群岛本土种子植物及其不同功能型组植物丰富度格局的主要因素。但值得注意的是, 舟山群岛包含500 m2以上的岛屿1,339个, 本文尽管包括了目前能够获得的最完整的舟山岛屿植物调查资料, 但这仅占岛屿总数的很小一部分。舟山群岛生物多样性的数据尚不完整, 且缺少长期深入的调查与研究工作, 这也极大限制了对舟山群岛生物地理学的研究。这一问题在我国其他区域的岛屿研究中也都普遍存在。因此, 我国岛屿生物地理学的研究需要长时间、多尺度、多类群的研究积累, 才能最终深入理解我国岛屿生物多样性时空格局的生态与演化机制, 为岛屿生物多样性保护和自然保护地管理提供科学依据。
附录 Supplementary Material
参考文献
Model Selection and Multimodel Inference: A Practical Information-theoretic Approach, 2nd edn
On the island biogeography of aliens: A global analysis of the richness of plant and bird species on oceanic islands
DOI:10.1111/geb.2016.25.issue-7 URL [本文引用: 1]
Habitat filtering not dispersal limitation shapes oceanic island floras: Species assembly of the Galápagos archipelago
DOI:10.1111/ele.12753
PMID:28294532
[本文引用: 2]

Remote locations, such as oceanic islands, typically harbour relatively few species, some of which go on to generate endemic radiations. Species colonising these locations tend to be a non-random subset from source communities, which is thought to reflect dispersal limitation. However, non-random colonisation could also result from habitat filtering, whereby only a few continental species can become established. We evaluate the imprints of these processes on the Galápagos flora by analysing a comprehensive regional phylogeny for ~ 39 000 species alongside information on dispersal strategies and climatic suitability. We found that habitat filtering was more important than dispersal limitation in determining species composition. This finding may help explain why adaptive radiation is common on oceanic archipelagoes - because colonising species can be relatively poor dispersers with specific niche requirements. We suggest that the standard assumption that plant communities in remote locations are primarily shaped by dispersal limitation deserves reconsideration.© 2017 John Wiley & Sons Ltd/CNRS.
Area threshold and trait-environment associations of butterfly assemblages in the Zhoushan Archipelago, China
DOI:10.1111/jbi.v48.4 URL
Beta regression in R
Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest
DOI:10.1093/aob/mcs092 URL [本文引用: 1]
Three newly recorded plants in Zhoushan Islands, Zhejiang
浙江舟山群岛三种新记录植物
Distribution characteristics and species diversity of seed plants in Zhoushan, Zhejiang
舟山市种子植物物种多样性及其分布特征
Climatic seasonality is linked to the occurrence of the mixed evergreen and deciduous broad-leaved forests in China
Geographical and climatic gradients of evergreen versus deciduous broad-leaved tree species in subtropical China: Implications for the definition of the mixed forest
DOI:10.1002/ece3.2967
PMID:28616161
[本文引用: 3]
Understanding climatic influences on the proportion of evergreen versus deciduous broad-leaved tree species in forests is of crucial importance when predicting the impact of climate change on broad-leaved forests. Here, we quantified the geographical distribution of evergreen versus deciduous broad-leaved tree species in subtropical China. The Relative Importance Value index (RIV) was used to examine regional patterns in tree species dominance and was related to three key climatic variables: mean annual temperature (MAT), minimum temperature of the coldest month (MinT), and mean annual precipitation (MAP). We found the RIV of evergreen species to decrease with latitude at a lapse rate of 10% per degree between 23.5 and 25°N, 1% per degree at 25-29.1°N, and 15% per degree at 29.1-34°N. The RIV of evergreen species increased with: MinT at a lapse rate of 10% per °C between -4.5 and 2.5°C and 2% per °C at 2.5-10.5°C; MAP at a lapse rate of 10% per 100 mm between 900 and 1,600 mm and 4% per 100 mm between 1,600 and 2,250 mm. All selected climatic variables cumulatively explained 71% of the geographical variation in dominance of evergreen and deciduous broad-leaved tree species and the climatic variables, ranked in order of decreasing effects were as follows: MinT > MAP > MAT. We further proposed that the latitudinal limit of evergreen and deciduous broad-leaved mixed forests was 29.1-32°N, corresponding with MAT of 11-18.1°C, MinT of -2.5 to 2.51°C, and MAP of 1,000-1,630 mm. This study is the first quantitative assessment of climatic correlates with the evergreenness and deciduousness of broad-leaved forests in subtropical China and underscores that extreme cold temperature is the most important climatic determinant of evergreen and deciduous broad-leaved tree species' distributions, a finding that confirms earlier qualitative studies. Our findings also offer new insight into the definition and distribution of the mixed forest and an accurate assessment of vulnerability of mixed forests to future climate change.
Long-distance dispersal: A framework for hypothesis testing
DOI:10.1016/j.tree.2011.08.009 URL [本文引用: 1]
Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox
Contemporizing island biogeography theory with anthropogenic drivers of species richness
DOI:10.1111/geb.v32.2 URL [本文引用: 2]
Ecophysiological and bioclimatic foundations for a global plant functional classification
DOI:10.1111/jvs.2010.21.issue-2 URL [本文引用: 1]
Island biogeography of the Anthropocene
DOI:10.1038/nature13739 [本文引用: 4]
Satellite based land use and landscape complexity indices as predictors for regional plant species diversity
DOI:10.1016/S0169-2046(02)00194-9 URL [本文引用: 1]
Island species richness increases with habitat diversity
Human land use promotes the abundance and diversity of exotic species on Caribbean Islands
DOI:10.1111/gcb.14334
PMID:29851186
[本文引用: 1]
Human land use causes major changes in species abundance and composition, yet native and exotic species can exhibit different responses to land use change. Native populations generally decline in human-impacted habitats while exotic species often benefit. In this study, we assessed the effects of human land use on exotic and native reptile diversity, including functional diversity, which relates to the range of habitat use strategies in biotic communities. We surveyed 114 reptile communities from localities that varied in habitat structure and human impact level on two Caribbean islands, and calculated species richness, overall abundance, and evenness for every plot. Functional diversity indices were calculated using published trait data, which enabled us to detect signs of trait filtering associated with impacted habitats. Our results show that environmental variation among sampling plots was explained by two Principal Component Analysis (PCA) ordination axes related to habitat structure (i.e., forest or nonforest) and human impact level (i.e., addition of man-made constructions such as roads and buildings). Several diversity indices were significantly correlated with the two PCA axes, but exotic and native species showed opposing responses. Native species reached the highest abundance in forests, while exotic species were absent in this habitat. Human impact was associated with an increase in exotic abundance and species richness, while native species showed no significant associations. Functional diversity was highest in nonforested environments on both islands, and further increased on St. Martin with the establishment of functionally unique exotic species in nonforested habitat. Habitat structure, rather than human impact, proved to be an important agent for environmental filtering of traits, causing divergent functional trait values across forested and nonforested environments. Our results illustrate the importance of considering various elements of land use when studying its impact on species diversity and the establishment and spread of exotic species.© 2018 The Authors. Global Change Biology published by John Wiley & Sons Ltd.
Species cataloging of the seed plants in Zhejiang, East China
DOI:10.17520/biods.2021408
[本文引用: 1]
<p id="p00005"><strong>Aims:</strong> The inventory and classification of biodiversity and biodiversity monitoring are the two core issues of global biodiversity research. Species inventory is the basis for understanding species diversity, and only by mastering the distribution pattern of species and the relationship between species and the environment we can provide a basis for species monitoring and scientific management. <br><strong>Method:</strong> Based on field work, specimen collection, specimen examination, and literature review, we provided an inventory of seed plants in the Zhejiang Province. <br> <strong>Results:</strong> The present checklist records a total of 4,430 species belonging to 1,469 genera in 212 families, including 3,347 wild plants in 1,085 genera of 190 families. There are 7 families with more than 100 species, namely Poaceae (285 species), Cyperaceae (216 species), Asteraceae (186 species), Rosaceae (153 species), Orchidaceae (126 species), Fabaceae (109 species) and Lamiaceae (108 species). There are 15 genera with more than 20 species, including <i>Carex</i> (126 species), <i>Phyllostachys</i> (44 species), <i>Rubus</i> (44 species), <i>Ilex</i> (35 species), <i>Polygonum</i> (34 species), <i>Lysimachia</i> (32 species), <i>Clematis</i> (31 species), <i>Sedum</i> (28 species), <i>Acer</i> (26 species), <i>Viburnum</i> (26 species), <i>Fimbristylis</i> (26 species), <i>Artemisia</i> (25 species), <i>Viola</i> (22 species), <i>Vitis</i> (21 species) and <i>Symplocos</i> (21 species). There are 149 families with less than 20 species (78.42% of total species), which contains 388 genera (35.76%) and 902 species (26.95%). Genera with less than 5 species are 952 genera (87.74%), which includes 1,707 species (51.00%). <br> <strong>Conclusions:</strong> Based on the statistical analysis, the floristic characteristics of Zhejiang are as follows: (1) the region is rich in species, with diverse families and genera; (2) the glacial refugia in East China retained many ancient and relict plants; (3) multiform geographic components reflect the transitional zone from the tropic to the temperate zone; (4) the region is rich in endemic and protected species which need to be protected; (5) there are many alien and invasive plants which need to be prevented early on.</p>
浙江种子植物物种编目
Global daily 1 km land surface precipitation based on cloud cover-informed downscaling
DOI:10.1038/s41597-021-01084-6
PMID:34836980
[本文引用: 1]
High-resolution climatic data are essential to many questions and applications in environmental research and ecology. Here we develop and implement a new semi-mechanistic downscaling approach for daily precipitation estimate that incorporates high resolution (30 arcsec, ≈1 km) satellite-derived cloud frequency. The downscaling algorithm incorporates orographic predictors such as wind fields, valley exposition, and boundary layer height, with a subsequent bias correction. We apply the method to the ERA5 precipitation archive and MODIS monthly cloud cover frequency to develop a daily gridded precipitation time series in 1 km resolution for the years 2003 onward. Comparison of the predictions with existing gridded products and station data from the Global Historical Climate Network indicates an improvement in the spatio-temporal performance of the downscaled data in predicting precipitation. Regional scrutiny of the cloud cover correction from the continental United States further indicates that CHELSA-EarthEnv performs well in comparison to other precipitation products. The CHELSA-EarthEnv daily precipitation product improves the temporal accuracy compared with a large improvement in the spatial accuracy especially in complex terrain.© 2021. The Author(s).
Global diversity of island floras from a macroecological perspective
DOI:10.1111/j.1461-0248.2007.01129.x
PMID:18036182
[本文引用: 6]
Islands harbour a significant portion of all plant species worldwide. Their biota are often characterized by narrow distributions and are particularly susceptible to biological invasions and climate change. To date, the global richness pattern of islands is only poorly documented and factors causing differences in species numbers remain controversial. Here, we present the first global analysis of 488 island and 970 mainland floras. We test the relationship between island characteristics (area, isolation, topography, climate and geology) and species richness using traditional and spatial models. Area is the strongest determinant of island species numbers (R(2) = 0.66) but a weaker predictor for mainlands (R(2) = 0.25). Multivariate analyses reveal that all investigated variables significantly contribute to insular species richness with area being the strongest followed by isolation, temperature and precipitation with about equally strong effects. Elevation and island geology show relatively weak yet significant effects. Together these variables account for 85% of the global variation in species richness.
Role of climate and geohistorical factors in driving plant richness patterns and endemicity on the East Asian continental islands
DOI:10.1111/ecog.2015.v38.i6 URL [本文引用: 1]
Environmental filtering underpins the island species—Area relationship in a subtropical anthropogenic archipelago
DOI:10.1111/jec.v108.2 URL [本文引用: 1]
Plant diversity on islands in the Anthropocene: Integrating the effects of the theory of island biogeography and human activities
DOI:10.1016/j.baae.2023.07.006 URL [本文引用: 3]
Beta diversity of vascular plants and its drivers in sea-islands of eastern China
DOI:10.17520/biods.2018235
[本文引用: 3]
Plant beta diversity describes how species composition shifts along environmental gradients. Islands are characterized by natural boundaries, variation in size and distance from the mainland, and they also experience intensive shifts in environmental properties. To date, plant beta diversity and its association with influencing factors across islands remain poorly understood. This study examined patterns and abiotic drivers of vascular plant beta diversity across 36 sea-islands in eastern China. Beta diversity was quantified using the Jaccard dissimilarity index based on presence/absence data, and patterns and drivers were analyzed by using partial Mantel tests and the variance partitioning approach. In total, there were 1,404 vascular plant species, including 481 woody species, 859 herbaceous species and 64 liana species. Plant beta diversity increased significantly with increase in cross-island distance and distance difference from the mainland. Differences in both island area and climatic factors did not affect plant beta diversity significantly. Cross-island distance accounted for 29.3% of total variation in plant beta diversity. Distance from the mainland explained 2.8%, and island area and climatic differences together explained 0.5%. The pattern of plant beta diversity was consistent between woody and herbaceous plants, but distance-based variables accounted for more beta diversity variation for woody than herbaceous plants (37.5% > 25.3%). Collectively, cross-island distance and the difference between islands in distance from the mainland are the main drivers of plant beta diversity. This suggests that dispersal limitation plays a key role in shaping plant beta diversity in the sea-islands of eastern China.
中国东部海岛维管植物的beta多样性及其驱动因素
DOI:10.17520/biods.2018235
[本文引用: 3]
beta多样性描述群落物种组成如何随环境梯度而变化。海岛具有边界清晰、面积和离岸距离不同以及环境变化剧烈等自然禀赋。目前, 我们对离岸距离、岛间距离和气候因素在海岛植物beta多样性变化格局中的相对贡献仍认识不足。本研究基于中国东部36个海岛的维管植物物种名录, 以Jaccard相异性指数度量beta多样性, 利用Mantel偏相关分析和beta多样性的变异分解, 探究了海岛不同生活型维管植物的beta多样性格局及其非生物影响因素。结果显示: 36个海岛共记录维管植物1,404种, 其中木本植物481种, 草本植物859种, 藤本植物64种。植物beta多样性随岛间距离和离岸距离差的增大而显著增加(P < 0.001); 海岛面积和气候要素对植物beta多样性无显著影响(P > 0.05)。岛间距离单独对beta多样性总变异的解释度为29.3%, 离岸距离独立解释了2.8%, 面积和气候共同解释了0.5%。木本植物与草本植物的beta多样性格局与总体一致, 距离因子对木本植物beta多样性的解释度高于草本植物(37.5% > 25.3%)。综上, 海岛植物beta多样性主要受岛间距离和离岸距离的影响, 反映了扩散限制是塑造中国东部海岛植物beta多样性格局的主要生态过程。
Ecology’s most general, yet protean pattern: The species-area relationship
DOI:10.1046/j.1365-2699.2000.00377.x URL [本文引用: 1]
A global method for calculating plant CSR ecological strategies applied across biomes world-wide
DOI:10.1111/fec.2017.31.issue-2 URL [本文引用: 1]
R: A Language and Environment for Statistical Computing
Island biodiversity in the Anthropocene
DOI:10.1146/annurev-environ-101718-033245
[本文引用: 1]
Biodiversity on marine islands is characterized by unique biogeographic, phylogenetic and functional characteristics. Islands hold a disproportionate amount of the world's biodiversity, and they have also experienced a disproportionate loss of it. Following human contact, island biodiversity has sustained negative human impacts increasing in rate and magnitude as islands transitioned from primary through secondary to tertiary economies. On islands, habitat transformation and invasive non-native species have historically been the major threats to biodiversity, and although these threats will continue in new forms, new impacts such as human-induced climate change and sea-level rise are emerging. Island biodiversity is changing with some species going extinct, others changing in abundance, non-native species becoming a part of many ecosystems, and humans shaping many ecological processes. Islands thus are microcosms for the emerging biodiversity and socioecological landscapes of the Anthropocene. Islands will require new strategies for the protection and restoration of their biodiversity, including maintaining biological and cultural heritage through regenerative practices, mainstreaming biodiversity in cultural and production landscapes, and engaging with the reality of novel ecosystems.
Species-area relationships on small islands differ among plant growth forms
DOI:10.1111/geb.v29.5 URL [本文引用: 3]
Spatial patterns and climate relationships of major plant traits in the New World differ between woody and herbaceous species
DOI:10.1111/jbi.2018.45.issue-4 URL [本文引用: 1]
Universal species-area and endemics-area relationships at continental scales
DOI:10.1038/nature11226 [本文引用: 1]
Island biogeography is not a single-variable discipline: The small island effect debate
DOI:10.1111/ddi.2011.18.issue-1 URL [本文引用: 2]
Determinants of alpha and beta vascular plant diversity in Mediterranean island systems: The Ionian Islands
Greece
Conceptual synthesis in community ecology
DOI:10.1086/652373 URL [本文引用: 1]
Disentangling natural and anthropogenic drivers of native and non-native plant diversity on North Sea Islands
Plant dispersal characteristics shape the relationship of diversity with area and isolation
DOI:10.1111/jbi.v49.9 URL [本文引用: 1]
Nearby large islands diminish biodiversity of the focal island by a negative target effect
DOI:10.1111/jane.v92.2 URL [本文引用: 2]
Floristic composition and diversity of typical plant community in Zhoushan Archipelago, East China
舟山群岛典型植物群落物种组成及多样性
以舟山群岛特有树种普陀樟(Cinnamomum japonicum var. chenii)、舟山新木姜子(Neolitsea sericea)和全缘冬青(Ilex integra)等占优势的植物群落为研究对象,通过典型样地调查,对物种组成及多样性进行了研究。结果表明:7个植物群落共调查到维管束植物125种,隶属于62科100属,其中滨海特有植物有12科14属16种。地理成分多样,热带成分大于温带、亚热带成分。各层次优势种明显,乔木层主要有红楠(Machilus thunbergii)、普陀樟、全缘冬青和舟山新木姜子等,在更新层中普陀樟和红楠最具优势,而灌木层、草本层、藤本层的优势种取决于群落类型及其所处的环境。群落间各层次的物种丰富度指数(R),Shannon指数(H),Pielou指数(J)均有较大波动,一般与群落的演替阶段、所处岛屿的大小、离大陆远近以及人为干扰有关,其中R、H保持一致的变化趋势。舟山海岛最具代表性的普陀樟群落各层次的R、H均低于其他群落。
Distribution of plants and relationship between plants and environment factors in Small Yangshan Island, Zhejiang Shengsi
浙江嵊泗小洋山岛植物分布特点和环境因子分析
Bioclimatic and physical characterization of the world’s islands
Quantifying island isolation—Insights from global patterns of insular plant species richness
DOI:10.1111/ecog.2013.36.issue-4 URL [本文引用: 1]
Determinants of species-area relationship and species richness of coastal endemic plants in the Fujian Islands
DOI:10.17520/biods.2022345
[本文引用: 2]
<p id="p00005"><strong>Aim:</strong> Fujian Province is made up of numerous islands that are rich in endemic plant resources. Conducting research on their distribution patterns and mechanisms of diversification can provide a basis for conservation and protection of coastal endemic plants. Our aim is to investigate the distribution patterns and mechanisms of species richness of coastal endemic plants in Fujian islands. <br><strong>Methods:</strong> This study investigated the species-area relationship of coastal endemic plants in 53 islands of Fujian Province as well as the impact of 10 environmental factors on plant species richness from three sources: landscape, human disturbance, and climate. Differential responses by various growth forms of plants, such as trees, shrubs, and herbs, to environmental factors were also discussed.<br><strong>Results:</strong> The species-area relationship between coastal endemic plants and different growth forms of plants showed a trend of increasing species richness with increasing area. The area, shape index, and perimeter area ratio of environmental factors of the landscape were the main factors that explain the abundance of coastal endemic plants and different growth forms of species on Fujian islands. Both the coastal endemic plants and their differing growth forms increased as the area and shape index increased, while the coastal endemic plants species richness as well as shrub and herb richness decreased with an increase in the perimeter area ratio. Further, the various growth forms exhibited differential species richness responses to island environmental factors. Apart from landscape factors, climate had the highest interpretation rate for trees, reaching 9.82%.<br><strong>Conclusion:</strong> Compared to other island ecosystems, the species richness of coastal endemic plants and different growth forms in the Fujian islands is strongly correlated with island area, but are less sensitive to island habitats. Landscape environmental factors contributed more to the interpretation rate than climate and human disturbance. Previous studies have found that island environmental factors are significantly correlated with habitat heterogeneity. For example, the larger the island, the more irregular its shape, and the lower its perimeter area ratio, the higher the habitat heterogeneity. Therefore, the high level of habitat heterogeneity in this study may be the underlying mechanism that explains the changes in coastal endemic plant abundance with landscape environmental factors.</p>
福建海岛滨海特有植物种-面积关系及物种丰富度决定因素
DOI:10.17520/biods.2022345
[本文引用: 2]
福建海岛具有丰富的滨海特有植物资源, 研究其物种丰富度的分布格局及机制, 能够为滨海特有植物资源保护提供依据。本研究探讨了福建53个海岛滨海特有植物的种-面积关系, 以及景观、人为干扰和气候3个方面10个环境因子对滨海特有植物物种丰富度的影响。同时还探讨了不同生活型(乔木、灌木、草本)对海岛环境因子的响应。结果表明: 滨海特有植物和不同生活型植物的种-面积关系呈现物种丰富度随着面积增加而增加的趋势, 滨海特有植物、乔木、灌木和草本的种-面积关系的斜率(z)分别为0.16、0.15、0.15和0.14。景观环境因子中的面积、形状指数和周长面积比是解释福建海岛滨海特有植物及不同生活型植物物种丰富度的主要决定因素, 滨海特有植物及各生活型植物物种丰富度随着面积和形状指数的增加而增加, 而滨海特有植物、灌木和草本的物种丰富度随着周长面积比的增加而减少。此外, 不同生活型植物的物种丰富度对海岛环境因子的响应存在差异, 除景观方面的因子外, 气候对乔木的解释率最大, 达9.82%。综上所述, 相比于其他海岛生态系统, 福建海岛滨海特有植物及不同生活型植物的物种丰富度与面积密切相关, 其较低的斜率(z)表明滨海特有植物对海岛生境的敏感性较弱。景观方面的环境因子相比气候和人为干扰方面贡献了更多的解释率。以往研究发现岛屿环境因子与生境异质性显著相关, 如岛屿越大、形状越不规则、周长面积比越低, 则生境异质性越高。因此生境异质性的提升可能是解释滨海特有植物丰富度随景观环境因子变化的内在机制。
Island area and remoteness shape plant and soil bacterial diversity through land use and biological invasion
DOI:10.1111/fec.v37.5 URL [本文引用: 4]
Edward O. Wilson and the Theory of Island Biogeography
DOI:10.17520/biods.2022024 [本文引用: 1]
E. O. 威尔逊与岛屿生物地理学理论
Determinants of bryophyte species richness on the Zhoushan Archipelago, China
DOI:10.1016/j.baae.2019.05.002 URL [本文引用: 3]
Richness and composition of plants and birds on land-bridge islands: Effects of island attributes and differential responses of species groups
DOI:10.1111/jbi.2012.39.issue-6 URL [本文引用: 2]
U.Taxonstand: An R package for standardizing scientific names of plants and animals
DOI:10.1016/j.pld.2022.09.001
[本文引用: 1]
The scientific names of organisms are key identifiers of plants and animals. Correctly treating scientific names is a prerequisite for biodiversity research and documentation. Here, we present an R package, ‘U.Taxonstand’, which can standardize and harmonize scientific names in plant and animal species lists at a fast speed and at a high rate of matching success. Unlike most of other similar R packages each of which works with only one taxonomic database, U.Taxonstand can work with all taxonomic databases, as long as they are properly formatted. Multiple databases for plants and animals that can be directly used by U.Taxonstand, which include bryophytes, vascular plants, amphibians, birds, fishes, mammals, and reptiles, are available online. U.Taxonstand can be a very useful tool for botanists, zoologists, ecologists and biogeographers to standardize and harmonize scientific names of organisms.
Vegetation characteristics and plant diversity of Waimalangshan Island, Zhoushan
外马廊山岛植被特性与植物多样性
Seed plant flora of Dongfushan Island in Zhoushan, Zhejiang Province
浙江舟山东福山岛种子植物区系初探
Species components and hazards of alien invasive plants in Putuoshan Island, Zhejiang Province
浙江普陀山外来入侵植物组成及危害现状

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}