



生物多样性是生物与其环境形成的生态复合体以及与此相关的各种生态过程的总和, 包括数以百万计的动物、植物、微生物和它们所拥有的基因及其生存环境形成的复杂生态系统, 是生命系统的基本特征(马克平和钱迎倩, 1998)。昆虫物种多样性作为生物多样性的重要组成部分, 对于整个生态系统有着至关重要的作用(Basset et al, 2012)。蝴蝶隶属昆虫纲鳞翅目, 其个体较大, 世代较短, 易于观测和鉴定(刘立伟等, 2015)。有研究表明, 蝴蝶多样性与植物多样性呈正相关(Soga et al, 2015), 与森林覆盖面积和生境连通性关系密切(Topp & Loos, 2019), 对环境和气候因子敏感, 在一定范围内, 其多样性会随温度和湿度的增加而增加(Beirão et al, 2021)。另外, 生境类型和海拔梯度对蝴蝶物种分布有重要影响(Beirão et al, 2021), 不同季节的降水量也对蝴蝶群落有一定影响(洪雪萌等, 2018)。因此, 蝴蝶被认为是生态系统、生境结构以及气候变化的指示生物之一, 其多样性观测数据已作为对各类型生态系统进行评价的依据(Kremen, 1992; Caldas & Robert, 2003; Kumar et al, 2009; 张立微和张红玉, 2016)。
20世纪末, 美国、日本、澳大利亚和欧洲部分国家就已完成了对本国蝴蝶资源的调查, 并且在蝴蝶群落动态变化、多样性组成、生态环境监测、栖息地恢复和濒危物种保护等方面的研究取得了一定的成果, 为保护蝴蝶和可持续利用蝴蝶资源提供了科学依据(Bried et al, 2014; Iserhard et al, 2017; Wepprich et al, 2019)。近十几年来, 我国蝴蝶多样性调查和保护工作发展迅速, 在蝴蝶多样性丰富的自然保护区(顾欣和任国栋, 2011; 苑彩霞等, 2012; 梅杰等, 2015; 周光益等, 2016)、国家森林公园(蒲正宇等, 2012, 2013; 徐静静和王国秀, 2016; 刘小龙等, 2018)和部分生物多样性热点地区(张承德等, 2017; 钱宏革等, 2018; 林中阳等, 2019)相继开展了蝴蝶群落多样性、物种资源和区系分布等调查工作, 涉及城市(吴云鹤等, 2016)、森林(蒲正宇等, 2012)、草原(戈昕宇等, 2018)、农田(马琦等, 2012)等多种生态系统, 取得了诸多成果, 为我国蝴蝶多样性保护、蝴蝶资源调查和生态环境监测提供了强有力的数据支持。但区域性质的蝴蝶多样性研究并不能完全满足环境大数据的需求, 2016年由环境保护部南京环境科学研究所牵头, 组织国内53家相关科研机构组建了“全国蝴蝶多样性观测网络”, 并出台了《蝴蝶生物多样性观测技术导则》, 该导则规定了蝴蝶多样性观测的目标、原则和方法(马方舟等, 2018), 开启了全国尺度下的蝴蝶多样性研究工作。
秦岭山脉位于亚热带和暖温带的分界线, 在动物区系上处于古北界和东洋界的分界线, 地理位置特殊、气候条件复杂、地形地貌多样, 有着丰富而宝贵的动植物资源。秦岭山脉可分为西、中、东三段, 本文所研究的区域为秦岭西段, 该地区涵盖了落叶阔叶林、针阔混交林、常绿针叶林、农田、落叶灌木丛和高山草甸等多种生境类型, 物种资源丰富, 是研究中国动植物区系特征、起源、演化的热点和关键区域之一(杨星科, 2005)。本研究通过查阅历史文献、书籍(蔡继增等, 2011; 杨星科, 2018), 整理出分布于秦岭西段地区的蝴蝶种类有5科181属440种, 但这些调查均以蝴蝶物种普查为主, 缺乏系统性的多样性调查与评估。本研究根据秦岭西段地区现有生态系统, 基于实际调查数据, 阐明秦岭西段地区蝴蝶群落物种多样性现状, 分析蝴蝶物种多样性随生境和季节的变化规律, 评估蝴蝶群落多样性对环境因子的响应, 以期为该地区蝴蝶物种多样性保护提供基础数据并提出相应的保护建议。
1 研究区域和研究方法
1.1 研究区域概况
1.2 调查方法
综合考虑地域、自然保护区和国家森林公园分布情况, 在秦岭西段地区设立15个10 km × 10 km的调查样区(图1, 附录1), 每个样区中设置9条调查样线, 共计135条样线。为避免边缘效应, 样线间距在300 m以上, 每条样线长1 km, 用“两步路” app进行距离测量和定位。依据不同的植被类型, 将调查区域划分为落叶灌木丛、针阔混交林、落叶阔叶林、常绿针叶林和农田5种生境, 分别于2020年9-10月(秋季)、2021年5月(春季)和2021年7-8月(夏季)对以上135条样线进行蝴蝶多样性观测, 每条样线用时约1 h, 保持匀速前进, 记录样线左右2.5 m、上方5 m、前方5 m范围内见到的所有蝴蝶种类和数量, 对于不易识别的蝴蝶采集标本带回实验室进行鉴定。
图1
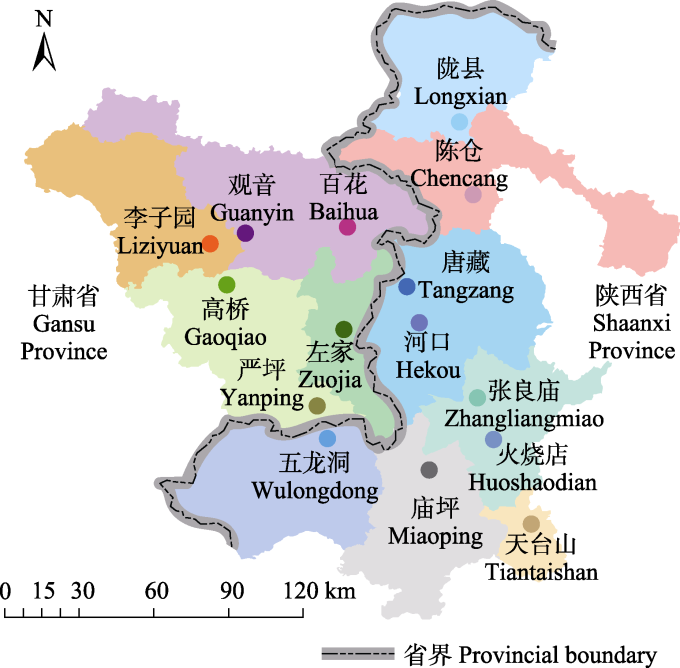
调查时详细记录采集日志, 包括样线编号、经纬度、采集人和采集日期, 并用风速仪记录温度、湿度和风速。在夏季采集中, 根据每条样线中实际的植物类型, 随机设置4个植物样方, 在野外观测记录样方内植物种类、盖度、密度, 测量高度, 收集样方内地表枯枝落叶并称重, 在实验室进行数据录入和整理。
1.3 蝴蝶群落的α多样性和β多样性分析
采用Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数、Simpson指数及Berger- Parker优势度指数来反映物种多样性和群落结构特征, 各指数计算公式如下:
Pielou均匀度指数(J) (Whittaker, 1972):
Margalef丰富度指数(R) (Margalef, 1958):
Simpson指数(D) (Pielou, 1975):
Berger-Parker优势度指数(W) (马克平和刘玉明, 1994):
式中, Ni为第i种的个体数量, N表示全部个体总数, S为物种数, Pi为第i种个体数的比例。定义W ≥ 25%为极优势物种; 5%-25% (不包含25%)之间为优势物种; 0.5%-5% (包含0.5%)之间为常见物种; 0.1%-0.5%为稀有物种; 小于0.1%为极稀有物种(Caruso et al, 2007)。
采用SPSS 22.0对不同季节和不同生境下蝴蝶群落组成、多样性指数进行单因素方差分析(one-way ANOVA), 采用最小显著性差异法(least significance difference, LSD)检验差异显著性, 观察在5%水平上是否具有统计学显著性, 使用Origin 22.0绘图。
利用R 4.1.2软件中的iNEXT程序包(Chao et al, 2014)分析不同生境和季节之间物种多样性的稀疏性并外推(通过现有趋势将观察到的物种数量进行合理的外推, 可以获得理论物种数)。iNEXT程序包只计算希尔系数(Hill numbers)的3种度量: 当参数q = 0时计算得到物种丰富度, 即物种数量; 当参数q = 1时计算得到Shannon-Wiener多样性, 即群落中常见物种的数量; 当参数q = 2时计算得到Simpson多样性, 即群落中优势种的数量(Hsieh et al, 2016)。对于每个多样性度量, 使用观察到的丰度来计算稀疏样本和外推样本的多样性数值, 绘制稀疏和外推(rarefaction/extrapolated)曲线(Colwell et al, 2012)。使用vegan程序包(Oksanen et al, 2015)对不同生境和不同季节的蝴蝶群落组成的β多样性进行非度量多维尺度分析(non-metric multidimensional scaling, NMDS) (Borcard et al, 2014)。使用vegan程序包(Oksanen et al, 2015)、ggdendro程序包(De Vries, 2012)、gridExtra程序包(Auguie, 2015)和ggplot2程序包(Wickham, 2009)对不同生境和季节间的蝶类群落进行非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)聚类, 并绘制聚类图和条形图。
1.4 蝴蝶群落多样性与环境因子相关性分析
使用R 4.1.2软件的mgcv程序包(Wood, 2022)对不同样线蝴蝶群落的Shannon-Wiener多样性指数、Simpson指数、Margalef丰富度指数和Pielou均匀度指数与主要环境因子进行广义加性模型(generalized additive model, GAM)拟合, 绘制响应曲线。
2 结果
2.1 蝴蝶群落组成
共记录蝴蝶8,898头, 隶属于5科84属169种(附录2)。其中粉蝶科、蛱蝶科和灰蝶科的个体数分别占总个体数的41.26%、27.70%和20.30%, 为极优势类群; 凤蝶科占7.04%, 为优势类群; 弄蝶科在0.5%-5%之间, 为常见类群(表1)。蛱蝶科物种数最多, 有37属80种; 其次为弄蝶科和灰蝶科, 均为16属23种; 粉蝶科为9属28种; 凤蝶科最少, 仅6属15种。
表1 秦岭西段蝴蝶群落组成与个体数量
Table 1
| 科 Families | 属 Genera | 种 Species | 个体数 Individuals | 百分比 Percent (%) |
|---|---|---|---|---|
| 蛱蝶科 Nymphalidae | 37 | 80 | 2,465 | 27.70 |
| 粉蝶科 Pieridae | 9 | 28 | 3,671 | 41.26 |
| 凤蝶科 Papilionidae | 6 | 15 | 626 | 7.04 |
| 弄蝶科 Hesperiidae | 16 | 23 | 329 | 3.70 |
| 灰蝶科 Lycaenidae | 16 | 23 | 1,807 | 20.30 |
| 合计 Total | 84 | 169 | 8,898 | 100 |
2.2 蝴蝶群落α多样性
2.2.1 不同生境蝴蝶群落α多样性
秦岭西段地区不同生境间蝴蝶的物种数、Shannon-Wiener多样性指数和Margalef丰富度指数存在显著差异, 个体数、Simpson指数和Pielou均匀度指数间无显著差异。个体数排序为落叶阔叶林 >落叶灌木丛 > 针阔混交林 > 农田 > 常绿针叶林,物种数排序为落叶阔叶林 > 针阔混交林 > 落叶灌木丛 > 常绿针叶林 > 农田, Shannon-Wiener指数和Simpson指数排序为落叶阔叶林 > 针阔混交林 > 常绿针叶林 > 落叶灌木丛 > 农田, Margalef丰富度指数排序为落叶阔叶林 > 针阔混交林 > 落叶灌木丛 > 常绿针叶林 > 农田, Pielou均匀度指数排序为常绿针叶林 > 落叶阔叶林 > 针阔混交林 > 农田 > 落叶灌木丛(图2)。
图2
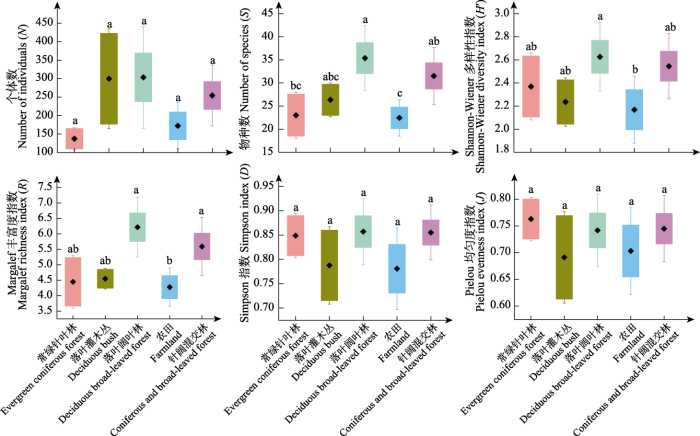
图2
秦岭西段不同生境蝴蝶物种多样性。
不同小写字母表示不同生境多样性指数差异显著。
Fig.2
Species diversity of butterflies under different habitats in the western region of Qinling Mountains.
Different lowercase letters indicate significant differences in different habitats for diversity index.
基于实际观测到的蝴蝶数量进行稀疏外推分析, 结果显示, 针阔混交林中的Shannon-Wiener多样性和Simpson多样性最高, 落叶阔叶林略小, 常绿针叶林居中, 农田生境小于常绿针叶林, 落叶灌木丛最小; 物种丰富度为针阔混交林最高, 落叶阔叶林略小, 农田居中, 常绿针叶林小于农田生境, 落叶灌木丛最小(图3)。
图3
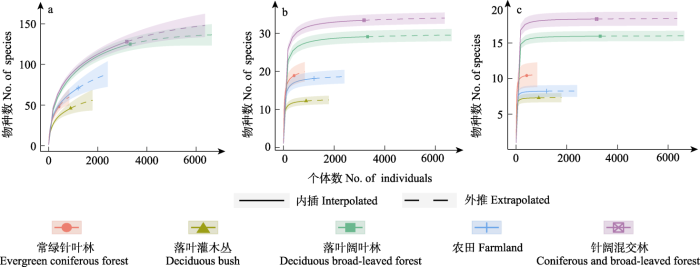
图3
秦岭西段不同生境蝴蝶物种多样性稀疏外推分析结果。
(a)物种丰富度; (b) Shannon-Wiener多样性; (c) Simpson多样性。
Fig. 3
Species diversity extrapolated analysis results of butterflies under different habitats in the western region of Qinling Mountains.
(a) Species richness; (b) Shannon-Wiener diversity; (c) Simpson diversity.
2.2.2 不同季节蝴蝶群落α多样性
不同季节间蝴蝶群落多样性指数差异较大, 除Pielou均匀度指数外, 其余各多样性指数排序为夏季 > 春季 > 秋季, 夏季、春季的蝴蝶群落个体数和Simpson指数同秋季间存在显著差异, 春、夏、秋季的蝴蝶群落物种数、Margalef丰富度指数和Shannon-Wiener多样性指数均存在显著差异; Pielou均匀度指数排序为秋季 > 夏季 > 春季, 且3个季节间均存在显著差异(图4)。
图4
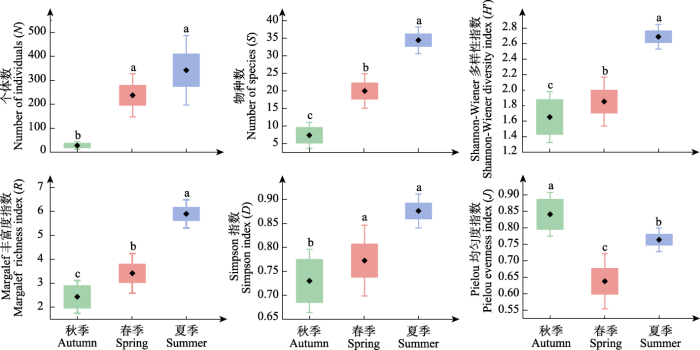
图4
秦岭西段不同季节蝴蝶物种多样性。
不同小写字母表示不同季节多样性指数差异显著。
Fig. 4
Species diversity of butterflies under different seasons in the western region of Qinling Mountains.
Different lowercase letters indicate significant differences in different seasons for diversity index.
由稀疏外推曲线可知(图5), 不同季节的物种丰富度和Shannon-Wiener多样性以夏季最高, 秋季次之, 春季略低于秋季; Simpson多样性是夏季最高, 春季次之, 秋季最低。
图5
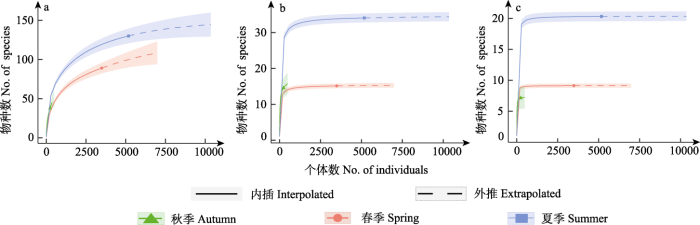
图5
秦岭西段不同季节蝴蝶物种多样性稀疏外推分析结果。
(a)物种丰富度; (b) Shannon-Wiener多样性; (c) Simpson多样性。
Fig. 5
Species diversity extrapolated analysis results of butterflies under different seasons in the western region of Qinling Mountains.
(a) Species richness; (b) Shannon-Wiener diversity; (c) Simpson diversity.
2.3 蝴蝶群落β多样性
2.3.1 不同生境的NMDS和UPGMA聚类结果
β多样性空间分布图显示(图6a), 落叶阔叶林、针阔混交林和农田中物种分布比较分散, 落叶灌木丛和常绿针叶林中较为集中。落叶阔叶林和针阔混交林的蝴蝶物种重叠度高, 农田的物种与针阔混交林、落叶阔叶林重叠度较低。
图6
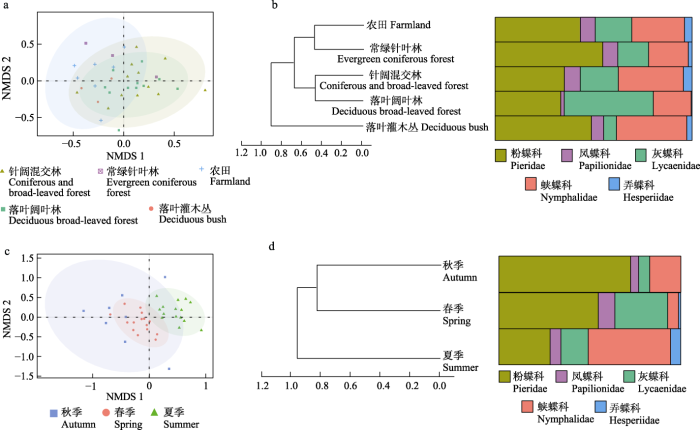
图6
蝴蝶群落β多样性。
(a)不同生境的蝴蝶群落NMDS分析结果; (b)不同生境的蝴蝶群落UPGMA聚类分析结果; (c)不同季节的蝴蝶群落NMDS分析结果; (d)不同季节的蝴蝶群落UPGMA聚类分析结果。
Fig. 6
β diversity analysis of butterfly community.
(a) NMDS analysis of butterflies in different habitats; (b) UPGMA cluster of butterflies in different habitats; (c) NMDS analysis of butterflies in different seasons; (d) UPGMA cluster of butterflies in different seasons.
不同生境间的多样性聚类分析结果显示(图6b), 当类间距离大于0.5时, 针阔混交林和落叶阔叶林聚为一支, 农田和常绿针叶林聚为一支; 当类间距离大于0.7时, 落叶灌木丛为独立生境类型, 与其他生境的多样性差异较大, 其他4种生境聚为一支。落叶阔叶林中, 灰蝶科个体数最多; 针阔混交林中粉蝶科和蛱蝶科个体数相近, 大于其他3科; 农田、常绿针叶林和落叶灌木丛中, 粉蝶科的个体数最多。
2.3.2 不同季节的NMDS和UPGMA聚类结果
β多样性空间分布格局显示, 春季和夏季物种分布较为集中, 秋季较为分散, 秋季各点和春季间距离较近, 同夏季距离较远, 但3个季节明显分离, 少有重叠(图6c)。
不同季节的多样性聚类分析结果显示(图6d), 当类间距离大于0.8时, 春季和秋季聚为一支, 夏季为单独一支。但3个季节间蝴蝶群落未见明显相似性。春季和秋季粉蝶科个体数最多, 夏季蛱蝶科个体数最多。
2.4 蝴蝶群落多样性与环境因子相关性
图7
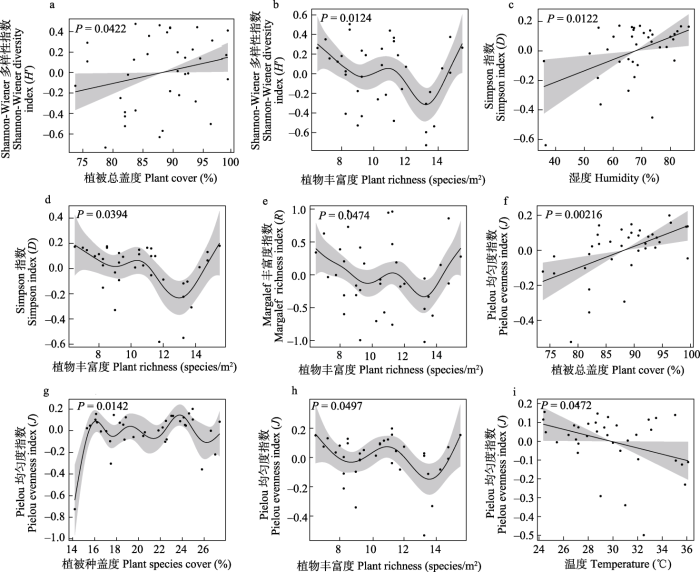
图7
蝴蝶群落多样性对环境因子的广义加性模型拟合曲线
Fig. 7
Fitting curve of generalized additive model of butterfly community diversity to environmental factors
Margalef丰富度指数对相关环境因子响应曲线表明, 植物丰富度对蝴蝶群落Margalef丰富度指数的影响具有统计学意义(P ≤ 0.05)。随着植物丰富度的增大, 蝴蝶群落的Margalef丰富度指数先减小, 在植物丰富度为13种/m²时达到最小值, 而后递增(图7e)。
3 讨论
3.1 秦岭西段地区蝴蝶群落组成
3.2 不同生境蝴蝶群落多样性
基于观测值水平的α多样性计算结果显示, 落叶阔叶林生境中蝴蝶群落的物种数、Margalef丰富度指数、Shannon-Wiener多样性指数和Simpson指数均为最高, 针阔混交林其次, 落叶灌木丛和常绿针叶林居中, 农田处于较低水平。但稀疏外推曲线结果表明: 若继续加大观测值, 针阔混交林的α多样性将最高, 落叶阔叶林次之, 农田和常绿针叶林居中, 落叶灌木丛数值最低。这是因为针阔混交林植物种类最为丰富, 可以满足绝大多种蝴蝶群落取食和繁衍的需求, 为其提供了较为稳定的生存环境。Baguette等(2011)在研究比利时东南部地区蛱蝶分布与生境质量之间的关系后也得到了类似的结论, 生境内植物长势较差或者植物种类减少后, 蝴蝶群落的分布率会降低, 迁移率增加。本次研究中落叶灌木丛的α多样性最低, 但韩艺茹等(2022)关于燕山地区访花昆虫多样性的研究结果表明落叶灌木丛生境α多样性最高, 这可能是因为秦岭西段地区落叶灌木丛均处于较低海拔, 周围人为活动明显, 干扰强度较大, 破坏了蝴蝶群落的生存环境, 但具体原因仍有待进一步深入研究。
由不同生境间的β多样性分析可以看出, 秦岭西段地区针阔混交林和落叶阔叶林的蝴蝶群落相似性较高, 同农田、常绿针叶林和落叶灌木丛的差异较为显著, 这是因为不同蝴蝶群落对蜜源植物有不同的偏向性, 例如, 粉蝶科和蛱蝶科蝴蝶偏爱访问菊科植物, 尤其是飞蓬和大蓟类植物(任炳忠等, 2012), 针阔混交林和落叶阔叶林植被较为相似, 与另外3种生境差异性较大, 从而导致了蝴蝶群落分布的不同。
3.3 不同季节蝴蝶群落多样性
蝴蝶群落组成随季节变化明显, 不同地区所呈现的变化模式不同, 这可能与研究地区的气温和植物多样性有关(Manwar & Wankhade, 2014; 黄敦元等, 2020)。气温显著影响蝴蝶群落的多样性, 适宜的温度有助于蝴蝶的生存和发育(张立微和张红玉, 2016; Gezon et al, 2018)。有研究表明, 气温的升高可缩短蝴蝶幼虫和蛹的发育时长, 使羽化提前, 气温降低会延迟蝴蝶的繁育(Jia et al, 2010; 李密, 2011), 寄主植物的多样性直接影响了区域内蝴蝶群落的分布(Matter et al, 2009)。秦岭西段地区在夏季(7-8月)温度最为适宜, 寄主植物丰富, 可为蝴蝶群落提供较多的取食、栖息和繁衍的场所, 基于观测值水平下的α多样性计算结果和α多样性物种丰富度外推曲线均表明, 夏季(7-8月)蝴蝶群落的个体数、物种数、Margalef丰富度指数、Shannon-Wiener多样性指数和Simpson指数均远高于春季(5月)和秋季(9-10月)。但在秋季(9-10月)蝴蝶群落的Pielou均匀度指数最高, 其原因是在夏季(7-8月)蝴蝶群落虽最为丰富, 但也存在明显的优势种群, 在秋季(9-10月)秦岭西段地区气温较低, 雨水偏多, 植被长势较差, 优势种群的生态结构遭到破坏, 不能体现出其优势性, 故而群落中蝴蝶物种和个体数分配均匀, 均匀度指数提高。
不同季节的β多样性空间分布格局表明, 3个季节间蝴蝶群落差异显著, 并不存在显著相似性, 说明蝴蝶群落结构随季节的变化具有显著的时序特征; 秋季各点较为分散, 春季和夏季较为集中, 说明在秋季不同的样区中蝴蝶种类差异较大, 在春季和夏季不同样区中的蝴蝶种类差异较小。有研究表明, 蝴蝶群落对气候、寄主植物和食物的适应性不同, 也就导致了蝴蝶羽化时间和成虫寿命的差异(Beck, 2008), 秦岭西段地区春、夏、秋季温差明显, 植物种类和丰富度差异较大, 从而导致了如此明显的时序特征和样区间差异。
3.4 环境因子对秦岭西段地区蝴蝶群落多样性的影响
气候因子和植被因子对物种分布有重要影响(Tews et al, 2004; Driscoll et al, 2013)。有研究表明气候因子影响蝴蝶在空间和时间上的分布, 在一定范围内, 蝴蝶的多样性会随着温度和湿度的增大而增大(Acharya & Vijayan, 2015; Beirão et al, 2021)。植被因子对蝴蝶群落多样性的影响较为复杂, 植被盖度、植物丰富度等均起到一定作用。有研究表明, 植被盖度相对较大, 植物相对丰富的次生林中蝴蝶多样性高于植物物种相对单一的人工林, 植物物种更为简单的农田中的蝴蝶多样性远远小于植物物种复杂的森林生境(Vu & Quang, 2011, Öckinger et al, 2012; Vu et al, 2015)。
本研究通过GAM模型将多样性指数与环境因子进行拟合后发现, 秦岭西段地区蝴蝶群落的多样性会随着植被盖度和湿度的增大而增大, 这与Acharya和Vijayan (2015)与Beirão等(2021)的研究结果相似。Kitahara等(2008)通过研究日本富士山西北坡蝴蝶群落和植被的关系, 发现蝴蝶物种多样性和植物丰富度之间有强相关性, 其中蜜源植物的丰富度是支撑蝴蝶群落多样性的重要因素。秦岭西段地区蝴蝶群落多样性随着植物丰富度的增大先减小随后再增大, 这可能是在植物丰富度达到一定数值之前, 植被环境的异质性较低, 蜜源植物较少, 仅可以满足少数几种蝴蝶种群的取食、繁衍和栖息的需求, 这样会出现几个优势种群, 总体蝴蝶群落的多样性反而降低了, 随着植物丰富度的增加植被环境的异质性也增大, 蝴蝶的食物来源增多, 群落多样性指数出现回升。Hamer等(2006)的研究结果表明, 温度会直接或者间接地影响蝴蝶群落组成和分布, 本研究中均匀度指数和温度拟合曲线显示, 随着温度的升高, 蝴蝶群落的均匀度指数反而降低, 这可能是因为在一定范围内, 温度的升高导致蝴蝶群落出现明显的优势类群, 从而使得总体蝴蝶群落的均匀度指数有所减小。
3.5 秦岭西段地区蝴蝶群落多样性保护建议
秦岭西段地区生物资源丰富, 具有很大的开发利用潜力, 经过全面系统的研究该地区蝴蝶群落多样性, 发现调查样区内受到不同程度的干扰, 对蝴蝶多样性造成一定影响。其中, 农林活动、放牧和旅游开发建设对蝴蝶群落多样性影响较大, 为了维护秦岭西段地区生态平衡稳定, 针对当地生态服务功能和资源特点提出以下几点建议: (1)持续开展多样性监测工作, 实时掌握蝴蝶物种和种群的动态变化和生存情况, 为制定合理的保护措施提供数据支持, 但要对采集时间、数量、方法等进行严格控制, 避免过度采集和毁灭性采集; (2)做好生物多样性保护的宣传工作, 加强宣传和引导, 强化人们的生态多样性保护意识, 明确生态多样性保护与林业可持续发展的关系和重要性; (3)充分发挥保护区管理部门的职能和作用, 与当地政府相配合, 考虑制定相关政策, 规范农牧秩序, 减少日常生活耕作放牧等活动对保护区内生物多样性的破坏, 合理发展生态旅游, 在提高生态效益的基础上提高当地的经济水平; (4)营造多样的植物群落和生境, 人工造林时要注重乔木、灌木和草本植物的合理组合, 形成一定规模和面积并加强管护, 促进植被恢复向着自然演替的方向健康、有序地进行。
附录 Supplementary Material
附录1 调查样区基本信息
Appendix 1 Basic information of the survey sample
附录2 秦岭西段地区蝴蝶名录
Appendix 2 Checklist of butterfly in the Western Region of Qinling Mountains
参考文献
Butterfly diversity along the elevation gradient of Eastern Himalaya
DOI:10.1007/s11284-015-1292-0 URL [本文引用: 2]
Metapopulation dynamics of the bog fritillary butterfly: Experimental changes in habitat quality induced negative density- dependent dispersal
DOI:10.1111/j.1600-0587.2010.06212.x URL [本文引用: 1]
Arthropod diversity in a tropical forest
DOI:10.1126/science.1226727
PMID:23239740
[本文引用: 1]

Most eukaryotic organisms are arthropods. Yet, their diversity in rich terrestrial ecosystems is still unknown. Here we produce tangible estimates of the total species richness of arthropods in a tropical rainforest. Using a comprehensive range of structured protocols, we sampled the phylogenetic breadth of arthropod taxa from the soil to the forest canopy in the San Lorenzo forest, Panama. We collected 6144 arthropod species from 0.48 hectare and extrapolated total species richness to larger areas on the basis of competing models. The whole 6000-hectare forest reserve most likely sustains 25,000 arthropod species. Notably, just 1 hectare of rainforest yields >60% of the arthropod biodiversity held in the wider landscape. Models based on plant diversity fitted the accumulated species richness of both herbivore and nonherbivore taxa exceptionally well. This lends credence to global estimates of arthropod biodiversity developed from plant models.
Phylogenetic and ecological correlates with male adult life span of rainforest butterflies
DOI:10.1007/s10682-007-9179-3 URL [本文引用: 1]
Climate and plant structure determine the spatiotemporal butterfly distribution on a tropical mountain
DOI:10.1111/btp.12860 URL [本文引用: 4]
A framework to integrate habitat monitoring and restoration with endangered insect recovery
DOI:10.1007/s00267-014-0351-6
PMID:25108660
[本文引用: 1]
Monitoring is essential to track the long-term recovery of endangered species. Greater emphasis on habitat monitoring is especially important for taxa whose populations may be difficult to quantify (e.g., insects) or when true recovery (delisting) requires continuous species-specific habitat management. In this paper, we outline and implement a standardized framework to facilitate the integration of habitat monitoring with species recovery efforts. The framework has five parts: (1) identify appropriate sample units, (2) select measurable indicators of habitat requirements, (3) determine rating categories for these indicators, (4) design and implement appropriate data collection protocols, and (5) synthesize the ratings into an overall measure of habitat potential. Following these steps, we developed a set of recovery criteria to estimate habitat potential and initially assess restoration activities in the context of recovering an endangered insect, the Karner blue butterfly (Lycaeides melissa samuelis). We recommend basing the habitat potential grading scheme on recovery plan criteria, the latest information on species biology, and working hypotheses as needed. The habitat-based assessment framework helps to identify which recovery areas and habitat patches are worth investing in and what type of site-specific restoration work is needed. We propose that the transparency and decision-making process in endangered insect recovery efforts could be improved through adaptive management that explicitly identifies and tracks progress toward habitat objectives and ultimate population recovery.
Modified Pollard transects for assessing tropical butterfly abundance and diversity
DOI:10.1016/S0006-3207(02)00190-8 URL [本文引用: 1]
The Berger-Parker index as an effective tool for monitoring the biodiversity of disturbed soils: A case study on Mediterranean oribatid (Acari: Oribatida) assemblages
DOI:10.1007/s10531-006-9137-3 URL [本文引用: 1]
Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies
DOI:10.1890/13-0133.1 URL [本文引用: 1]
Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages
DOI:10.1093/jpe/rtr044 URL [本文引用: 1]
Butterfly nectaring flowers: Butterfly morphology and flower form
DOI:10.1046/j.1570-7458.2000.00708.x URL [本文引用: 1]
Host plants and butterfly biology. Do host-plant strategies drive butterfly status?
Conceptual domain of the matrix in fragmented landscapes
Butterfly community structure and diversity in Qinling National Botanical Garden, China
DOI:10.17520/biods.2020041 URL [本文引用: 1]
秦岭国家植物园蝴蝶群落结构与多样性
The species diversity of butterfly community in Saihanwula Nature Reserve of Inner Mongolia
内蒙古赛罕乌拉自然保护区蝶类群落多样性
Drifting phenologies cause reduced seasonality of butterflies in response to increasing temperatures
DOI:10.3390/insects9040174 URL [本文引用: 1]
Study on community diversity of butterfly in Xiaowutai Mountain State Nature Reserves
河北小五台山自然保护区蝶类群落多样性
Diversity and ecology of carrion- and fruit-feeding butterflies in Bornean rain forest
DOI:10.1017/S0266467405002750 URL [本文引用: 1]
Diversity and influencing factors of flower-visiting insects in the Yanshan area
DOI:10.17520/biods.2021448
[本文引用: 1]
<p id="p00005"><strong>Aim</strong> Our aim is to clarify the relationship between diversity and community structure of flower-visiting insects and their habitat type, disturbance degree, and altitude in the Yanshan area.</p><p id="p00010"><strong>Methods</strong> Throughout July-August of 2019 and 2020, we collected flower-visiting insects in five habitats, including wetland, forest, shrub, grassland and farmland, with different gradients of 0-1,200 m above the sea level by using the method of transects and light lures.</p><p id="p00015"><strong>Result</strong> A total of 1,306 flower-visiting insects were collected, belonging to 153 species, 44 families, and 7 orders, among which Lepidoptera had the largest number of species and Hemiptera had the largest number of individuals. The diversity of flower-visiting insects is highest in the shrub cluster. The Shannon-Wiener, Margalef, and Simpson indices are the highest in the middle and low altitudes of 200-400 m. The results of bivariate regression indicate that the Shannon-Wiener and Margalef indices are positively correlated with precipitation in the warmest season and annual precipitation respectively (<i>P</i>< 0.05). Redundancy analysis (RDA) demonstrated that environmental factors significantly affected the diversity of flower-visiting insects, but there was some variation among different measures. Temperature and humidity are positively correlated with the Pielou index and negatively correlated with the Shannon-Wiener, Margalef, and Simpson indices. The results further indicate that scientific management of husbandry activities is the key to protecting the diversity of flower-visiting insects.</p><p id="p00020"><strong>Conclusion</strong> Scientific management of husbandry activities is the key to protecting the diversity of flower-visiting insects.</p>
燕山地区访花昆虫多样性及其影响因子
DOI:10.17520/biods.2021448
[本文引用: 1]
为了解燕山地区访花昆虫的群落结构及与其生境类型、干扰程度、海拔之间的关系, 本文采用样线法和灯诱法于2019年、2020年每年的7-8月对该地区湿地、森林、灌丛、草地、农田5种生境, 不同海拔梯度(0-1,200 m)的访花昆虫进行了采集。共采集访花昆虫1,306头, 隶属7目44科153种, 其中鳞翅目昆虫物种数最多, 半翅目昆虫个体数最多。灌丛生境的访花昆虫多样性最高。在中低海拔200-400 m段, Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数均最高。双变量回归结果表明, Shannon-Wiener多样性指数和Margalef丰富度指数分别与最暖季降水量和年降水量显著正相关(P < 0.05)。冗余分析(redundancy analysis)结果表明, 环境因子显著影响访花昆虫多样性, 但不同测度之间存在一定差异。温度和湿度均与Pielou均匀度指数呈正相关, 与Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数呈负相关; 弱干扰和中干扰程度对访花昆虫多样性影响最小, 科学管理农牧活动是保护访花昆虫多样性的关键。
Butterfly diversity and its influencing factors in Saihanwula Nature Reserve
DOI:10.17520/biods.2018025
[本文引用: 1]
To explore the factors explaining butterfly diversity in a landscape mosaic of different habitats, we conducted a series of field surveys in the Saihawla National Nature Reserve. This reserve comprised of seven habitats: typical grassland, wetland, mountain valley meadow, degraded grassland, farmland, mountain xerophytic shrub and mountainous shrub. From May to September 2017, we recorded a total of 2,290 butterfly individuals belonging to 63 species, 42 genera, and 5 families. Nymphalidae, with the most species (34) and the most individuals (991), was the dominant family in the reserve. Five species dominated the community: Papilio Xuthus, Pontia daplidice, Aporia crataegi, Aglais urticae, Speyeria aglaja. The species-abundance analysis showed a normal distribution model, suggesting a relatively stable community with the range of butterfly activity extending to different habitats due to the continuity of vegetation types. However, we also found that butterfly distribution in Saihanwula was closely related to the distribution and composition of vegetation among habitats. The Mountainous shrub had the highest biodiversity index and the lowest dominance index, whereas degraded grassland had the lowest biodiversity index and species richness, but the highest dominance index. The Mountain valley meadow had the most number of families, genera, species, and individuals, the degraded grassland had the lowest number of families, genera, species, and individuals. Faunal distribution analysis show that widely distributed species occupied 63.49% and Palaearctic species occupied 36.51%. Furthermore, butterfly diversity in the different habitats of the reserve varied significantly between months. Finally, we found that numbers of butterfly individuals and species increased with higher temperatures, while rainfall had no significant correlation. In conclusion, We believe that appropriate interference is conducive to the development of butterfly diversity, strong human disturbance seriously destroyed grassland environment, affected butterfly survival and reproduction, and reduced butterfly diversity.
赛罕乌拉自然保护区蝶类多样性及其影响因素
DOI:10.17520/biods.2018025
[本文引用: 1]
为了解赛罕乌拉自然保护区蝶类多样性及其影响因素, 我们于2017年5-9月分别对保护区的典型草原、湿地、山地沟谷草甸、退化草原、农田、山地旱生灌丛、山地中生灌丛等7种生境中的蝴蝶进行观测调查。共记录和采集蝴蝶2,290只, 隶属5科42属63种。蛱蝶科的种类数(34种)和个体数(991只)最多。柑橘凤蝶(Papilio xuthus)、云粉蝶(Pontia daplidice)、绢粉蝶(Aporia crataegi)、荨麻蛱蝶(Aglais urticae)、银斑豹蛱蝶(Speyeria aglaja)等5种为保护区的优势种。保护区蝶类群落的种-多度曲线呈正态分布模式, 表明保护区生态环境良好, 生境相互重叠, 蝶类活动范围在不同生境中可以延伸。不同生境中蝶类群落种类及数量存在一定差异, 其中山地中生灌丛蝶类群落的多样性指数最高, 优势度指数最低; 退化草原的多样性指数和物种丰富度指数均为最低, 优势度指数最高; 山地沟谷草甸的科、属、种、个体数最多; 退化草原的科、属、种、个体数都是最少。区系组成分析表明广布种占63.49%, 古北种占36.51%。保护区不同生境中蝶类群落多样性特征指数在各月份间有明显差异, 蝴蝶种类及个体数与温度之间呈显著正相关, 与降雨量无显著相关性。综上, 我们认为适当的干扰有利于蝶类多样性发展, 而强烈的人为干扰会严重破坏草场环境, 影响蝶类生存和繁衍, 降低蝶类多样性。
iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers)
DOI:10.1111/2041-210X.12613 URL [本文引用: 1]
Diversity of butterfly communities in the Qiyunshan National Nature Reserve
DOI:10.17520/biods.2019420 URL [本文引用: 1]
齐云山国家级自然保护区蝴蝶群落多样性
Monitoring temporal variation to assess changes in the structure of subtropical Atlantic forest butterfly communities
DOI:10.1093/ee/nvx115
PMID:28881951
[本文引用: 1]
The study of fauna through long-term surveys is important in unveiling how temporal patterns shape the structure of communities in tropical habitats. The butterfly assemblage of the subtropical Atlantic Forest may be considered highly diverse and shows changes in diversity and composition over time, highlighting the importance of long-term inventories. This work assessed temporal diversity patterns in the distribution and composition of butterfly assemblages in an Atlantic Forest site in southern Brazil using combined data from three years of standardized sampling with entomological nets, increasing the knowledge on this group in the Neotropics for monitoring and conservation. The butterfly fauna was analyzed in terms of richness, abundance, and composition. The inventories reached 401 species, with 14,442 butterfly individuals sampled. All the diversity parameters evaluated show significant differences between the first year of sampling compared to the second and third years. The latter had higher values of richness and abundance, followed by the first and second years. Hesperiidae was the richest family, followed by Nymphalidae and Lycaenidae, indicating a good representation of the assemblage as a whole. The results of this work are important for developing conservation programs in the Atlantic Forest and other forested environments in the neotropics, especially concerning reliable diversity assessments for the monitoring and management of protected areas. Decision making and public policy might also benefit from knowledge on temporal patterns of diversity regarding the maintenance of native habitats and integrity of biomes and their associated fauna.© The Authors 2017. Published by Oxford University Press on behalf of Entomological Society of America. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Effects of temperature and rainfall on composition and diversity of butterflies in broad-leaved and Korean pine mixed forests in the Changbai Mountains, China
DOI:10.3724/SP.J.1145.2010.00007 URL [本文引用: 1]
Relationship of butterfly diversity with nectar plant species richness in and around the Aokigahara primary woodland of Mount Fuji, central Japan
DOI:10.1007/s10531-007-9265-4 URL [本文引用: 1]
Assessing the indicator properties of species assemblages for natural areas monitoring
DOI:10.2307/1941776 URL [本文引用: 1]
Effects of spatial heterogeneity on butterfly species richness in Rocky Mountain National Park, CO, USA
DOI:10.1007/s10531-008-9536-8 URL [本文引用: 1]
Studies on Conservation Biology of Butterfly in Wuyunjie National Nature Reserve
乌云界国家级自然保护区蝴蝶保护生物学研究
Climatic elements and their variation characteristics of Xiaolongshan forest region in western Qinling Mountains
秦岭西段小陇山林区气候要素及其变化特征研究
Study on the diversity of butterflies in Xichou County, Yunnan Province
云南省西畴县蝴蝶群落结构与多样性
Resources and fauna of butterflies in Jiulongshan Mountain National Nature Reserve
九龙山国家级自然保护区蝶类资源与区系
Diversity and faunal analysis of butterfly species in Xishan National Forest Park in Kunming
昆明西山国家森林公园蝶类多样性及区系分析
Progress in construction of China butterfly diversity observation network (China BON-Butterflies)
全国蝴蝶多样性观测网络(China BON-Butterflies)建设进展
Measurement of biotic community diversity I. α diversity (Part 1)
生物群落多样性的测度方法I. α多样性的测度方法(上)
Measurement of biotic community diversity I. α diversity (Part 2)
生物群落多样性的测度方法I. α多样性的测度方法(下)
Biodiversity conservation and its research progress
生物多样性保护及其研究进展
The analysis of grade diversity indices of butterfly community in the Three Gorges Reservoir Area of Yangtze River
DOI:10.5846/stxb201101190102 URL [本文引用: 1]
长江三峡库区蝶类群落的等级多样性指数
Seasonal variation in diversity and abundance of butterfly at Sawanga Vithoba Lake area district Amravati, Maharashtra India
DOI:10.3923/jbs.2014.485.493 URL [本文引用: 1]
Interactions between habitat quality and connectivity affect immigration but not abundance or population growth of the butterfly, Parnassius smintheus
DOI:10.1111/j.1600-0706.2009.17438.x URL [本文引用: 2]
Species diversity of butterflies in Fanjing Mountain National Nature Reserve of Guizhou
贵州梵净山国家级自然保护区蝴蝶多样性
Landscape matrix modifies richness of plants and insects in grassland fragments
DOI:10.1111/j.1600-0587.2011.06870.x URL [本文引用: 1]
Butterfly diversity dynamics in different seasons at the Jindian National Forest Park, Kunming, Yunnan
昆明金殿国家森林公园蝶类多样性季节性变化研究
Butterfly diversity of different types of habitats in Jindian National Forest Park, Kunming
昆明金殿国家森林公园不同生境类型蝶类多样性
The community diversity analysis of butterflies in Wula Mountain region from Inner Mongolia
内蒙古乌拉山地区蝴蝶群落多样性分析
Relationship between floral scent of the major nectar source plants and their flower visitors in Changbai Mountain region
长白山地区优势蜜源植物花的气味与访花昆虫种类的关系
Landscape versus local factors shaping butterfly communities in fragmented landscapes: Does host plant diversity matter?
DOI:10.1007/s10841-015-9799-9 URL [本文引用: 1]
Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures
DOI:10.1046/j.0305-0270.2003.00994.x URL [本文引用: 1]
Local and landscape level variables influence butterfly diversity in critically endangered South African renosterveld
DOI:10.1007/s10841-018-0104-6 URL [本文引用: 1]
Flower preferences of woodland butterflies in the UK: Nectaring specialists are species of conservation concern
DOI:10.1016/j.biocon.2004.01.002 URL [本文引用: 1]
Butterfly diversity and habitat variation in a disturbed forest in northern Vietnam
DOI:10.3956/2014-91.1.029 URL [本文引用: 1]
Diversity pattern of butterfly communities (Lepidoptera, Papilionoidae) in different habitat types in a tropical rain forest of southern Vietnam
Butterfly abundance declines over 20 years of systematic monitoring in Ohio, USA
Evolution and measurement of species diversity
DOI:10.2307/1218190 URL [本文引用: 1]
The influence of urbanization on butterfly diversity in Hefei, Anhui Province
城市化对合肥蝶类多样性的影响
Investigation on butterfly resources in Taizi Mountain National Forest Park
太子山国家森林公园蝶类资源调查
Faunal analysis of butterflies in Pingheliang Nature Reserve of Shaanxi Province
陕西省平河梁自然保护区蝶类资源调查及区系研究
Study on species diversity and host plant of butterfly in Qufu City
曲阜地区蝴蝶物种多样性及其寄主植物调查研究
Research progress in butterfly as indicators for habitat change
蝶类对生境的指示作用研究进展
Diversity and fauna of butterflies in Nanling National Nature Reserve
南岭国家级自然保护区蝴蝶多样性与区系研究

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}