



荒漠猫(Felis bieti)又名中国山猫、草猫、草猞猁, 属脊椎动物门哺乳动物纲食肉目猫科猫属(Felis), 是世界猫科动物中唯一的中国特有类群。荒漠猫定名于19世纪末(Felis bieti Milne-Edwards, 1892), 其分类学地位长期存在争议, 传统形态学和生物地理学的证据支持荒漠猫作为独立物种。而近期的分子遗传学(Driscoll et al, 2007)和基因组学研究(Yu et al, 2021)则表明荒漠猫应当与分布在欧洲地区的欧洲野猫(Felis silvestris silvestris)、分布在北非及近东地区的非洲野猫(F. s. lybica)、分布在里海东部中亚地区的亚洲野猫(F. s. ornata)以及分布在非洲南部的南非野猫(F. s. cafra)一同被划分为野猫(Felis silvestris)的亚种。
图1

图1
青藏高原特有猫科动物荒漠猫(Felis bieti)正模标本(法国巴黎国家自然历史博物馆馆藏ZM-MO-1891-391; 图Cecile Callou/MNHN)
Fig. 1
The holotype specimen of the Chinese mountain cat (Felis bieti) endemic to the east of the Qinghai-Tibet Plateau (ZM-MO-1891-391, National Museum of Natural History, Paris, France; Photo by Cecile Callou/ MNHN)
荒漠猫种群曾被认为广泛存在于中国西部地区, 包括甘肃、四川、内蒙、青海、宁夏、西藏和新疆等省区(Jacobi, 1923; Allen, 1938; Pocock, 1951; 高耀亭等, 1987; Wang, 1990)。最新评估结果显示其明确的分布区域仅限于青藏高原东部, 包括青海东部、四川西北部、甘肃西南部以及紧邻青海的西藏东南部部分地区(He et al, 2004; Luo et al, 2022)。在此范围, 荒漠猫和家猫的杂交现象偶有报道(Yin et al, 2007; Sanderson et al, 2010)。荒漠猫等中国本土野猫类群是否与家猫存在基因交流甚至参与家猫在亚洲本土的驯化, 一直是世界家猫遗传领域悬而未决的问题。
根据荒漠猫分布面积和种群密度的评估结果, 其种群数量不超过1万只, 已被IUCN濒危物种红色名录列为“易危”等级(Vulnerable, VU), 并是CITES附录Ⅱ中的保护动物。2021年, 我国将荒漠猫提升为国家一级重点保护野生动物(
1 荒漠猫的野外发现和分布
根据记载, 荒漠猫在中国西部包括甘肃、四川、内蒙古、青海、宁夏、西藏和新疆等省区均有分布(Jacobi, 1923; Allen, 1938; Pocock, 1951; 高耀亭等, 1987; Wang, 1990)。然而, 研究者通过对文献信息的整理以及对标本或活体动物的检验和核对发现, 许多荒漠猫的记录属于鉴定错误或者来源不可靠(He et al, 2004)。来自新疆、宁夏和内蒙古的荒漠猫记录属于其他野猫谱系(亚洲野猫)或家猫, 或者来源不可靠无法证实(Haltenorth, 1953; Guggisberg, 1975; Groves, 1980; Sunquist & Sunquist, 2002; Smith & Xie, 2008)。通过对相关荒漠猫的分布资料, 尤其是过去十多年来自然观察记录及红外相机调查结果的核实, 其县级分布区如表1所示(Luo et al, 2022)。一个县只要满足下述条件之一, 即可认为该县为荒漠猫的分布区: (1)该县通过红外相机捕捉或直接拍摄到过荒漠猫的活体或者尸体; (2)该县为荒漠猫剥制标本等样品的确切地理来源; (3)该县虽然近期没有确切的荒漠猫野外记录, 但这可能是由于野外调查的匮乏, 根据可靠的信息仍可判定该县为荒漠猫分布区的可能性较高。以县级行政单位的地理坐标作为分布点, 通过核密度估计(kernel density estimation)计算其分布范围, 其中位于格尔木市的记录仅来自于西宁动物园的动物来源记载(廖炎发, 1988), 而无更确切的分布信息, 因此将该点排除在分布区域之外, 最终得到荒漠猫分布面积为941,620 km2 (图2)。综上所述, 荒漠猫明确的分布区域确定为青藏高原东部, 包括青海东部、四川西北部、甘肃西南部以及紧邻青海的西藏东南部的部分地区。
图2
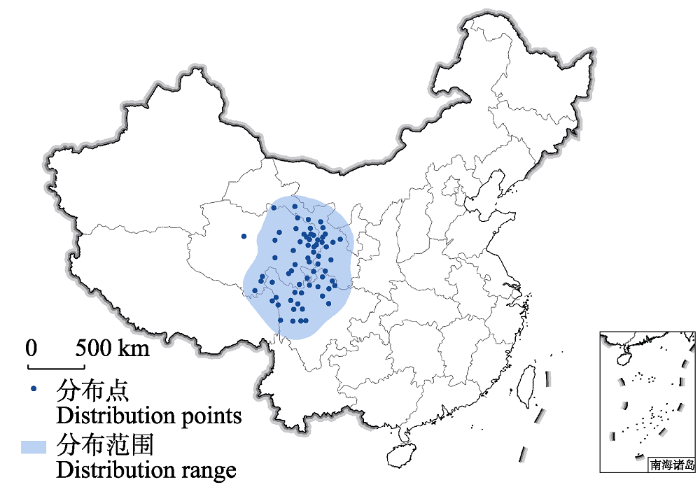
表1 中国荒漠猫县级分布地区
Table 1
| 省/自治区 Province | 州/市 Prefecture/City | 县/区 County/District | 参考文献 Reference | 其他来源(个人通讯) Other sources (personal communications) |
|---|---|---|---|---|
| 四川省 Sichuan Province | 甘孜藏族自治州 Ganzi Tibetan Autonomous Prefecture | 甘孜县, 新龙县, 雅江县, 石渠县, 康定市, 巴塘县, 道孚县, 炉霍县, 德格县, 理塘县#, 白玉县#, 色达县# Ganzi, Xinlong, Yajiang, Shiqu, Kangding, Batang, Daofu, Luhuo, Dege, Litang#, Baiyu# and Seda# | Milne-Edwards, 1892; de Pousargues, 1898; Allen, 1938; Pocock, 1951; Haltenorth, 1953; Guggisberg, 1975; Groves, 1980; He et al, 2004; Chen et al, 2005; Wang et al, 2021 | 李晟, 宋大昭, 张铭, CatSG数据库 Sheng Li, Dazhao Song, Ming Zhang; CatSG database |
| 阿坝藏族羌族自治州 Aba Tibetan and Qiang Autonomous Prefecture | 阿坝县, 红原县, 若尔盖县, 九寨沟县#, 松潘县, 壤塘县, 黑水县#, 汶川县# Aba, Hongyuan, Ruoergai, Jiuzhaigou#, Songpan, Rangtang, Heishui# and Wenchuan# | Jacobi, 1923; Weigold, 1923; Allen, 1938; Pocock, 1951; Haltenorth, 1953; Guggisberg, 1975; Groves, 1980; He et al, 2004; Yin et al, 2007; Webb et al, 2016 | 李晟, 罗述金, 宋大昭, CatSG数据库 Sheng Li, Shu-Jin Luo, Dazhao Song; CatSG database | |
| 绵阳市 Mianyang City | 平武县 Pingwu | Li et al, 2020 | ||
| 甘肃省 Gansu Province | 兰州市 Lanzhou City | 永登县, 皋兰县# Yongdeng and Gaolan# | Sanderson et al, 2010; Wan et al, 2020 | 孙悦华, 方昀, CatSG数据库 Yuehua Sun, Yun Fang; CatSG database |
| 白银市 Baiyin City | 靖远县# Jingyuan# | 罗述金 Shu-Jin Luo | ||
| 临夏回族自治州 Linxia Hui Autonomous Prefecture | 永靖县, 康乐县 Yongjing and Kangle | Wan et al, 2020 | 孙悦华, 方昀 Yuehua Sun and Yun Fang | |
| 定西市 Dingxi City | 渭源县 Weiyuan | 孙悦华, 方昀 Yuehua Sun and Yun Fang | ||
| 甘南藏族自治州 Gannan Tibetan Autonomous Prefecture | 碌曲县, 玛曲县, 临潭县, 卓尼县, 夏河县, 迭部县 Luqu, Maqu, Lintan, Zhuoni, Xiahe and Diebu | Lonnberg, 1925; Pocock, 1951, Haltenorth, 1953; Guggisberg, 1975; Groves, 1980; Sanderson et al, 2010 | 孙悦华, 方昀, 刘炎林, 罗述金, 张铭, 善觉, CatSG数据库 Yuehua Sun, Yun Fang, Yanlin Liu, Shu-Jin Luo, Ming Zhang, Jue Shan; CatSG database | |
| 武威市 Wuwei City | 古浪县, 凉州区, 天祝藏族自治县 Gulang, Liangzhou and Tianzhu Zangzu Autonomous County | 罗述金 Shu-Jin Luo | ||
| 张掖市 Zhangye City | 山丹县, 肃南裕固族自治县 Shandan and Yugur Autonomous County of Sunan | 何兵, 马堆芳 Bing He and Duifang Ma | ||
| 青海省 Qinghai Province | 西宁市 Xining City | 大通回族土族自治县, 湟中区#, 湟源县# Datong Hui and Tu Autonomous County, Huangzhong# and Huangyuan# | Büchner, 1892; Allen, 1938; Pocock, 1951; Guggisberg, 1975; Groves, 1980; Liao, 1988 | 刘炎林 Yanlin Liu |
| 海北藏族自治州 Haibei Tibetan Autonomous Prefecture | 祁连县, 门源回族自治县, 刚察县, 海晏县 Qilian, Menyuan Hui Autonomous County, Gangcha and Haiyan | Liao, 1988; He et al, 2004 | 刘炎林 Yanlin Liu | |
| 海南藏族自治州 Hainan Tibetan Autonomous Prefecture | 共和县, 兴海县#, 贵德县 Gonghe, Xinghai# and Guide | Matschie, 1908; Allen, 1938; Pocock, 1951; Guggisberg, 1975; Groves, 1980; Liao, 1988; He et al, 2004 | ||
| 海东市 Haidong City | 互助县, 民和县, 乐都区#, 化隆回族自治县# Huzhu, Minhe, Ledu# and Hualong# | Liao, 1988; He et al, 2004 | 罗述金 Shu-Jin Luo | |
| 黄南藏族自治州 Huangnan Tibetan Autonomous Prefecture | 河南蒙族自治区, 尖扎县#, 泽库县#, 同仁市# Mongolian Autonomous County of Henan, Jianzha#, Zeku# and Tongren# | Liao, 1988 | 刘炎林 Yanlin Liu | |
| 果洛藏族自治州 Guoluo Tibetan Autonomous Prefecture | 久治县, 玛沁县, 玛多县#, 甘德县#, 达日县#, 班玛县# Jiuzhi, Maqin, Maduo#, Gande#, Dari# and Banma# | Liao, 1988; Sanderson et al, 2010 | 阿旺, 刘炎林, 罗述金, 宋大昭, 张铭 Wang A, Yanlin Liu, Shu-Jin Luo, Dazhao Song and Ming Zhang | |
| 玉树藏族自治州 Yushu Tibetan Autonomous Prefecture | 称多县, 囊谦县#, 玉树市# Chengduo, Nangqian# and Yushu# | Liao, 1988; Han et al, 2020 | ||
| 省/自治区 Province | 州/市 Prefecture/City | 县/区 County/District | 参考文献 Reference | 其他来源(个人通讯) Other sources (personal communications) |
| 海西蒙古族藏族自治州 Haixi Mongol and Tibetan Autonomous Prefecture | 天峻县, 都兰县#, 乌兰县#, 格尔木市# Tianjun, Dulan#, Wulan# and Golmud# | Liao, 1988 | 刘炎林 Yanlin Liu | |
| 西藏自治区 Tibet Autonomous Region | 昌都市 Changdu City | 江达县# Jiangda# | 宋大昭, CatSG数据库 Dazhao Song; CatSG database |
# 该县为荒漠猫分布区的可能性较高
# There is a likely presence of the Chinese mountain cat in the county
2 荒漠猫的演化历史
作为青藏高原的特有类群, 荒漠猫的遗传溯源、基因交流和演化历史, 尤其是荒漠猫等中国本土野猫类群是否和家猫之间存在基因交流, 是否参与了家猫在中国本土的驯化, 都是世界家猫遗传研究领域的热点。2021年, Yu等从中国全境尤其是在家猫-荒漠猫同域分布地区采集了27只荒漠猫、4只亚洲野猫和239只家猫的组织、血液、粪便、毛发或博物馆标本样品, 综合系统发生和种群动态历史研究分析, 发现荒漠猫与亚洲野猫等其他野猫类群的分歧时间相近(Yu et al, 2021)。家猫-非洲野猫支系和荒漠猫-亚洲野猫支系大约在187万年前开始分化, 而荒漠猫与亚洲野猫两支系大约在127万年前开始分化(Yu et al, 2021)。种群动态历史分析结果显示, 荒漠猫在约200万年至100万年前曾经历了一个较小的种群增长, 随后从约100万年前开始至数万年前, 其有效种群数量持续下降(Yu et al, 2021)。
基因交流是演化生物学界备受关注的问题。越来越多的基因组学证据表明, 杂交是自然界中普遍存在的现象, 并在生物演化过程中起着重要的作用(Taylor & Larson, 2019)。种间或者亚种间的杂交在猫科动物中十分普遍, 如在豹支系、豹猫支系、猞猁支系等多个类群中都有发现(Trigo et al, 2013; Li et al, 2016; Figueiró et al, 2017)。基于最新的系统发生学和种群历史分析结果, 荒漠猫、亚洲野猫和家猫3个支系间也存在多种杂交事件(Yu et al, 2021)。荒漠猫与亚洲野猫在历史上存在着密切的基因交流(图3), 结合线粒体数据和基因组数据的分析结果, 可以推断这一历史杂交事件大概率发生于雌性亚洲野猫和雄性荒漠猫之间, 并且其杂交后代进入到了荒漠猫种群中。而对于荒漠猫和中国家猫, 该研究发现, 尽管包括荒漠猫在内的中国本土野猫没有参与中国家猫的驯化, 但荒漠猫与其同域家猫之间广泛存在近期的基因渗透。该遗传交流信号普遍存在于荒漠猫分布区内的家猫中, 时间尺度对应于20世纪中叶, 可能与藏区社会经济发展以及家猫种群在当地的增长相关(Yu et al, 2021)。此外值得注意的是, 种群遗传多样性的历史分析的确检测到从家猫到荒漠猫的基因渗透, 不过该基因流的强度在基于不同荒漠猫个体进行的分析中结果差异很大。这一发现也意味着青藏高原家猫种群对同域分布的荒漠猫可能产生的遗传影响将是亟需关注的问题。
图3
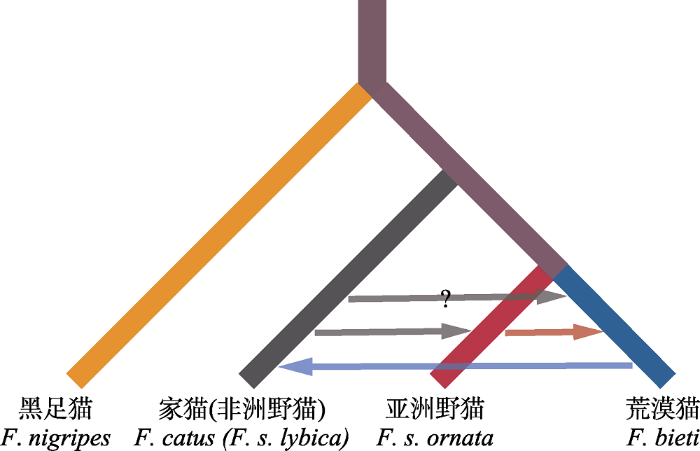
图3
野猫类群和家猫系统发生关系和基因流信号示意图
Fig. 3
Phylogenetic relationships and introgression signals among wildcat lineages and domestic cats
3 荒漠猫的分类学研究进展
1892年, 法国动物学家Milne-Edwards对荒漠猫进行了首次记录, 并将其命名为“Felis bieti” (Milne-Edwards, 1892)。1917年, 俄国科学家Birula将采集自“Goizso from Gobi of Southern Mongolia” (实际上是宁夏西北部)的标本命名为一个新物种“Felis chutuchta” (Birula, 1917; 高耀亭等, 1987; He et al, 2004), 之后该物种被认定为是荒漠猫的一个亚种(Allen, 1938; Pocock, 1943, 1951), 即宁夏亚种。1943年, 英国动物学家Pocock将采集于“Yulin, Shaanxi” (陕西榆林)的标本, 命名为荒漠猫的另一亚种“Felis bieti vellerosa” (Pocock, 1943; 高耀亭等, 1987), 即陕西亚种。但Haltenorth在1953年以及Groves在1980年分别指出上述所提及的宁夏亚种事实上应为亚洲野猫(Haltenorth, 1953; Groves, 1980; He et al, 2004)。此外, Groves等还在1980年指出陕西亚种实际上是家猫(Groves, 1980; He et al, 2004)。因此, 荒漠猫被认为是单型种。
尽管荒漠猫传统意义上被认为是一个独立的物种, 但它与其他野猫类群的分类学关系在学界尚未达成一致意见。Driscoll等(2007)基于分子遗传学证据, 提出荒漠猫、亚洲野猫、欧洲野猫、非洲野猫和南非野猫分属5个分歧明显但又关系紧密的演化支系, 且野猫支系和家猫间能普遍发生基因交流, 表明各支系间并不存在生殖隔离, 因此荒漠猫及其他4类野猫应当属于野猫的不同亚种。然而, Kitchener和Rees (2009)基于气候变化对野猫在末次冰盛期以来的地理分布进行了重建, 结果表明荒漠猫和亚洲野猫的分布区域可能存在交叠。结合形态上的差异, 他们推测二者之间存在生殖隔离, 因此认为荒漠猫应当被列为一个独立的物种。IUCN猫科动物专家组(IUCN SSC Cat Specialist Group)在发表分类名录时也采用了上述观点, 将荒漠猫列为独立的物种(Kitchener et al, 2017)。Yu等(2021)通过所采集的荒漠猫、亚洲野猫和家猫共计270只个体的线粒体和Y染色体多位点测序以及51只个体的全基因组测序数据, 基于系统发生、种群遗传结构、基因交流和种群动态历史研究分析, 证明了荒漠猫和亚洲野猫之间的演化遗传距离相当, 依据分类学原则应当将荒漠猫归为野猫亚种。不过由于荒漠猫、亚洲野猫和家猫的分歧时间已有100万年以上, 该研究也为解决野猫类群的分类学争议提供了另一种可能, 即将所有野猫亚种提升为独立的物种, 从而维持荒漠猫的物种地位。但这一分类观点的可行性还需要获得所有野猫类群的基因组数据, 在全基因组层面进行深入地分析(Yu et al, 2021)。
4 荒漠猫的致危因素和保护
目前尚没有关于荒漠猫种群数量的研究和报道, 通过其分布区面积和种群密度可做粗略估计(Luo et al, 2022)。荒漠猫的分布面积(图2)大约941,620 km2, 以其近缘野猫种群密度为参考, 荒漠猫成体数目保守估计不到1万只(Luo et al, 2022)。基于现有的荒漠猫野外记录, 可以推测其种群实际栖息地仅占分布区的部分区域。假设荒漠猫的实际栖息地面积仅为分布区域的50%或25%, 而密度范围为1-10只/100 km2, 则荒漠猫个体数预计在2,354-47,081只之间。假设其中50%的荒漠猫为成体, 则其成体数目在1,177-23,540只之间。根据这一估算结果, 并考虑到荒漠猫种群正在面临的威胁以及潜在威胁包括鼠药施用造成的二次毒害、毛皮贸易、西部开发造成的栖息地的丧失和碎片化以及与当地家猫种群之间的遗传杂交现象(He et al, 2004; Sanderson et al, 2010; Yu et al, 2021), 荒漠猫成体数目保守估计不到1万只。且这些威胁因素的持续存在可能会导致荒漠猫种群数量继续下降。
鼠药施用造成的二次毒害和非法毛皮交易可能仍在威胁着荒漠猫种群(Sanderson et al, 2010; Riordan et al, 2015)。鼠兔被认为会和家畜争夺牧草, 1958-1978年, 青海、四川和甘肃等地广泛使用灭鼠剂控制鼠兔数量, 但同时造成大量以鼠兔为捕食对象的食肉动物受到毒害(Sanderson et al, 2010)。虽然现在已经禁止大范围使用有毒化学试剂控制小型啮齿动物数量, 但仍存在一些小规模的灭鼠活动。此外, 与毛皮贸易相关的捕猎活动也曾在部分地区广泛存在(Chen et al, 2005)。但由于荒漠猫毛皮的经济价值不高, 市场和贸易规模较小, 毛皮贸易目前对荒漠猫可能并不是主要威胁(Luo et al, 2022)。
此外, 当地家猫种群对荒漠猫种群可能存在的基因渗透影响不容忽视。长期以来对欧洲野猫的研究已经证明, 来自家猫的基因渗透严重影响了这一类群的遗传多样性和环境适应能力(Allendorf et al, 2001; Randi et al, 2001; Randi, 2008)。近期的基因组学研究同样显示荒漠猫与其同域家猫间存在普遍的基因渗透(Yu et al, 2021)。虽然目前发现的交流信号为从荒漠猫到家猫, 但考虑到该研究收集的样品以家猫为主, 荒漠猫检测数量较少, 我们不能忽视荒漠猫中同样存在来自家猫的基因渗入的可能性。因此, 对荒漠猫种群进行进一步广泛的遗传评估, 全面深入地在荒漠猫基因组中检测是否存在来自当地家猫的基因渗透, 对于了解野生荒漠猫种群的现状和制定有效的保护措施至关重要。
总体而言, 为保护中国唯一的地方性猫科动物, 当务之急是评估现有荒漠猫种群保护措施的有效性, 包括现有保护区管理的有效性。
5 总结与展望
荒漠猫是世界猫科动物中唯一的中国特有类群, 也是世界上最鲜为人知的猫科动物之一。随着分子遗传学和基因组学的发展, 有关荒漠猫遗传演化和分类的研究取得了重要的进展。最新的基因组学研究为荒漠猫分类学地位的划分提供了新的遗传学观点和证据, 并系统地阐释了荒漠猫自更新世中期开始的演化历程, 以及荒漠猫与亚洲野猫、家猫之间的基因交流图景, 为荒漠猫和亚洲野猫等珍稀濒危野生猫科动物的保护提供了科学依据(Yu et al, 2021)。
基于荒漠猫的栖息地面积及其近缘野猫类群的密度估计, 荒漠猫成熟个体数保守估计在1万只以内, 且种群数量目前仍处于下降趋势(Luo et al, 2022)。基于现有研究, 荒漠猫种群正面临的一系列威胁, 不仅包括鼠药施用造成的二次毒害、毛皮贸易(Sanderson et al, 2010), 还包括经济建设开发造成的栖息地的丧失和片断化(He et al, 2004)以及路撞等人为因素。此外, 当地家猫种群与荒漠猫之间可能的基因渗透也应当被重视(Yu et al, 2021)。这可能会导致荒漠猫种群未来面临遗传多态性和环境适应能力丧失的风险。因此, 与荒漠猫相关的适应性演化机制以及荒漠猫是否面临着来自青藏高原同域家猫遗传渗透的威胁以及威胁程度仍需进一步地探索和研究。
此外, 最近十余年来, 研究人员和自然爱好者通过红外相机监测和自然观察项目, 在西北分布区域逐步发现了更多关于荒漠猫的活动记录(Luo et al, 2022)。然而, 为进一步评估栖息地等相关因素对荒漠猫种群数量的影响, 以及制定出一套有效的保护行动计划, 我们仍迫切地需要对荒漠猫进行更多的野外监测和研究, 开展更为全面的系统调查, 并评估当前保护管理措施的有效性, 以此来保护我国唯一特有的猫科动物。
致谢
感谢宋大昭、李晟、刘炎林、何兵、马堆芳、张铭、阿旺、孙悦华、方昀、齐新章、蒋可威、玛才让加、陈怀庆、蒙皓、闻丞、图登华旦、尹玉峰、耿栋、赵建友、刘永萍、李文靖、巫嘉伟、Nobuyuki Yamaguchi等在确定荒漠猫分布信息时提供的帮助。
参考文献
The problems with hybrids: Setting conservation guidelines
DOI:10.1016/S0169-5347(01)02290-X URL [本文引用: 1]
De Felibus asiaticus duabus novis
Über eine neue Katzen-Art (Felis pallida n. sp.) aus China
Status of the Chinese mountain cat in Sichuan Province (China)
Sur I’identite specifique du Felis bieti (A. M.-E.) et du Felis pallida (Buchn.)
The Near Eastern origin of cat domestication
The world's domestic cats carry patterns of sequence variation in their genome that reflect a history of domestication and breed development. A genetic assessment of 979 domestic cats and their wild progenitors-Felis silvestris silvestris (European wildcat), F. s. lybica (Near Eastern wildcat), F. s. ornata (central Asian wildcat), F. s. cafra (southern African wildcat), and F. s. bieti (Chinese desert cat)-indicated that each wild group represents a distinctive subspecies of Felis silvestris. Further analysis revealed that cats were domesticated in the Near East, probably coincident with agricultural village development in the Fertile Crescent. Domestic cats derive from at least five founders from across this region, whose descendants were transported across the world by human assistance.
Checklist of Palaearctic and Indian Mammals 1758 to 1946. British Museum (Natural History) Trustees
Genome-wide signatures of complex introgression and adaptive evolution in the big cats
Die Wildkatzen der Alten Welt
Discovery of first active breeding den of Chinese mountain cat
(Felis bieti).
Distribution and conservation status of the endemic Chinese mountain cat Felis bieti
Zoologische Ergebnisse der Walter Stötznerschen Expeditionen nach Szetschwan, Osttibet und Tschili auf Grund der Sammlungen und Beobachtungen Dr. Hugo Weigolds. 2. Teil, Aves: 4. Fringillidae und Ploceidae
A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/ SSC Cat Specialist Group
Modelling the dynamic biogeography of the wildcat: Implications for taxonomy and conservation
DOI:10.1111/j.1469-7998.2009.00599.x URL [本文引用: 1]
Phylogenomic evidence for ancient hybridization in the genomes of living cats (Felidae)
DOI:10.1101/gr.186668.114
PMID:26518481
[本文引用: 1]

Inter-species hybridization has been recently recognized as potentially common in wild animals, but the extent to which it shapes modern genomes is still poorly understood. Distinguishing historical hybridization events from other processes leading to phylogenetic discordance among different markers requires a well-resolved species tree that considers all modes of inheritance and overcomes systematic problems due to rapid lineage diversification by sampling large genomic character sets. Here, we assessed genome-wide phylogenetic variation across a diverse mammalian family, Felidae (cats). We combined genotypes from a genome-wide SNP array with additional autosomal, X- and Y-linked variants to sample ∼150 kb of nuclear sequence, in addition to complete mitochondrial genomes generated using light-coverage Illumina sequencing. We present the first robust felid time tree that accounts for unique maternal, paternal, and biparental evolutionary histories. Signatures of phylogenetic discordance were abundant in the genomes of modern cats, in many cases indicating hybridization as the most likely cause. Comparison of big cat whole-genome sequences revealed a substantial reduction of X-linked divergence times across several large recombination cold spots, which were highly enriched for signatures of selection-driven post-divergence hybridization between the ancestors of the snow leopard and lion lineages. These results highlight the mosaic origin of modern felid genomes and the influence of sex chromosomes and sex-biased dispersal in post-speciation gene flow. A complete resolution of the tree of life will require comprehensive genomic sampling of biparental and sex-limited genetic variation to identify and control for phylogenetic conflict caused by ancient admixture and sex-biased differences in genomic transmission. © 2016 Li et al.; Published by Cold Spring Harbor Laboratory Press.
Construction progress of the camera-trapping network for the mountains of Southwest China
DOI:10.17520/biods.2020038 URL [本文引用: 1]
西南山地红外相机监测网络建设进展
Some biological informations of desert cat in Qinghai
The Chinese desert cat (<i>Fells bieti</i>,MiIne-Edwards) lives in six different habitats in Qinghai province at an elevation of 2,800-4,100m,i.e.alpine meadow, alpine bush,edge of coniferous forest, grassy meadow, desert-semi desert zone and loess hill steppes.The desert cat occurs in Huangzhong and other 18 counties (suburb) in Qinghai, frequently in Huangzhong,Huangyuan,Menyuan, Qilian,Tongren and Dulan.The den of desert cat can be found in natural rocky cave on sunny slope between 3,000-3, 600m.There are breeding dens and temporary ones.The desert cat moves actively in morning,evening and at night.It lives a solitary life except in mating season.In breeding period, activity range of the female is limited and fixed. The main food is rodents, when lacking in winter, it preys on birds.
青海荒漠猫的一些生物学资料
本文对荒漠猫(Felis bieti)的形态、野外栖息环境、洞穴和捕食活动、食性等作了实地调查,同时结合西宁人民公园人工饲养荒漠猫的行为观察,综合整理的生物学初步观察资料。
Notes on some cats from Eastern Asia with description of a new subgenus
Felis bieti
Mammalia
In: #x000DC;ber chinesische Säugetiere, bosonders aus den Sammlungen des Herren Wilhelm Filchner. Wissenschaftliche Ergebnisse der Expedition Filchner nach China und Tibet 1903-1905.
Observations sur les Mammifères du Thibet
Catalogue of the Genus Felis
Detecting hybridization between wild species and their domesticated relatives
DOI:10.1111/j.1365-294X.2007.03417.x
PMID:18173502
[本文引用: 1]
The widespread occurrence of free-ranging domestic or feral carnivores (dogs, cats) or ungulates (pigs, goats), and massive releases of captive-reproduced game stocks (galliforms, waterfowl) is raising fear that introgressive hybridization with wild populations might disrupt local adaptations, leading to population decline and loss of biodiversity. Detecting introgression through hybridization is problematic if the parental populations cannot be sampled (unlike in classical stable hybrid zones), or if hybridization is sporadic. However, the use of hypervariable DNA markers (microsatellites) and new statistical methods (Bayesian models), have dramatically improved the assessment of cryptic population structure, admixture analyses and individual assignment testing. In this paper, I summarize results of projects aimed to identify occurrence and extent of introgressive hybridization in European populations of wolves (Canis lupus), wildcats (Felis silvestris), rock partridges and red-legged partridges (Alectoris graeca and Alectoris rufa), using genetic methods. Results indicate that introgressive hybridization can be locally pervasive, and that conservation plans should be implemented to preserve the integrity of the gene pools of wild populations. Population genetic methods can be fruitfully used to identify introgressed individuals and hybridizing populations, providing data which allow evaluating risks of outbreeding depression. The diffusion in the wild of invasive feral animals, and massive restocking with captive-reproduced game species, should be carefully controlled to avoid loss of genetic diversity and disruption of local adaptations.
Genetic identification of wild and domestic cats (Felis silvestris) and their hybrids using Bayesian clustering methods
Crossbreeding with free-ranging domestic cats is supposed to threaten the genetic integrity of wildcat populations in Europe, although the diagnostic markers to identify "pure" or "admixed" wildcats have never been clearly defined. Here we use mitochondrial (mt) DNA sequences and allelic variation at 12 microsatellite loci to genotype 128 wild and domestic cats sampled in Italy which were preclassified into three separate groups: European wildcats (Felis silvestris silvestris), Sardinian wildcats (Felis silvestris libyca), and domestic cats (Felis silvestris catus), according to their coat color patterns, collection localities, and other phenotypical traits, independently of any genetic information. For comparison, we included some captive-reared hybrids of European wild and domestic cats. Genetic variability was significantly partitioned among the three groups (mtDNA estimate of F(ST) = 0.36; microsatellite estimate of R(ST) = 0.30; P < 0.001), suggesting that morphological diversity reflects the existence of distinct gene pools. Multivariate ordination of individual genotypes and clustering of interindividual genetic distances also showed evidence of distinct cat groups, partially congruent with the morphological classification. Cluster analysis, however, did not enable hybrid cats to be identified from genetic information alone, nor were all individuals assigned to their populations. In contrast, a Bayesian admixture analysis simultaneously assigned the European wildcats, the Sardinian wildcats, and the domestic cats to different clusters, independent of any prior information, and pointed out the admixed gene composition of the hybrids, which were assigned to more than one cluster. Only one putative Sardinian wildcat was assigned to the domestic cat cluster, and one presumed European wildcat showed mixed (hybrid) ancestry in the domestic cat gene pool. Mitochondrial DNA sequences indicated that three additional presumed European wildcats might have hybrid ancestry. These four cats were sampled from the same area in the northernmost edge of the European wildcat distribution in the Italian Apennines. Admixture analyses suggest that wild and domestic cats in Italy are distinct, reproductively isolated gene pools and that introgression of domestic alleles into the wild-living population is very limited and geographically localized.
Felis bieti
Of the only endemic cat species in China: The Chinese mountain cat—Felis bieti
Insights from genomes into the evolutionary importance and prevalence of hybridization in nature
DOI:10.1038/s41559-018-0777-y
PMID:30697003
[本文引用: 1]
Hybridization is an evolutionary phenomenon that has fascinated biologists for centuries. Prior to the advent of whole-genome sequencing, it was clear that hybridization had played a role in the evolutionary history of many extant taxa, particularly plants. The extent to which hybridization has contributed to the evolution of Earth's biodiversity has, however, been the topic of much debate. Analyses of whole genomes are providing further insight into this evolutionary problem. Recent studies have documented ancient hybridization in a diverse array of taxa including mammals, birds, fish, fungi, and insects. Evidence for adaptive introgression is being documented in an increasing number of systems, though demonstrating the adaptive function of introgressed genomic regions remains difficult. And finally, several new homoploid hybrid speciation events have been reported. Here we review the current state of the field and specifically evaluate the additional insights gained from having access to whole-genome data and the challenges that remain with respect to understanding the evolutionary relevance and frequency of ancient hybridization, adaptive introgression, and hybrid speciation in nature.
Molecular data reveal complex hybridization and a cryptic species of neotropical wild cat
DOI:10.1016/j.cub.2013.10.046
PMID:24291091
[本文引用: 1]
Hybridization among animal species has recently become more recognized as an important phenomenon, especially in the context of recent radiations. Here we show that complex hybridization has led to contrasting patterns of genomic composition among closely related species of the Neotropical cat genus Leopardus. We show strong evidence of ancient hybridization and introgression between the pampas cat (L. colocolo) and northeastern populations of tigrina (L. tigrinus), leading to remarkable cytonuclear discordance in the latter. In contrast, southern tigrina populations show recent and continuing hybridization with Geoffroy's cat (L. geoffroyi), leading to extreme levels of interspecific admixture at their contact zone. Finally, we demonstrate that two seemingly continuous Brazilian tigrina populations show no evidence of ongoing gene flow between them, leading us to support their formal recognition as distinct species, namely L. tigrinus in the northeast and L. guttulus in the south.Copyright © 2013 Elsevier Ltd. All rights reserved.
Progress of the China mammal diversity observation network (China BON-Mammal) based on camera-trapping
DOI:10.17520/biods.2020142 URL [本文引用: 2]
基于红外相机的中国哺乳动物多样性观测网络建设进展
Xinlong County of Ganzi, Sichuan, a newly discovered felid hotspot in South-west China
Chinese mountain cat and Pallas’s cat co-existing on the Tibetan Plateau in Sinchuan
First photographs in nature of the Chinese mountain cat
Genomic evidence for the Chinese mountain cat as a wildcat conspecific (Felis silvestris bieti) and its introgression to domestic cats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}