



中国幅员辽阔, 河流众多, 是世界上河流最为丰富的国家之一。然而, 我们对河流底栖动物多样性的研究仍然滞后于世界同期水平。国内已有的研究多集中在长江、黄河、珠江和澜沧江等流域的中下游江段和支流, 而对中西部高原河流的调查仍然不足(王昱等, 2021)。雅鲁藏布江发源于西藏自治区境内, 是中国最长的高原跨境河流, 也是世界上海拔最高的大河(刘天仇, 1999)。该河流为我国西藏地区和下游南亚地区提供了主要的淡水资源, 在全球具有重要的地位(聂宁等, 2012)。由于具有海拔高、地表物理性质复杂等特点, 栖居于此的生物在全球气候变化中具有其自身特殊的演化规律(Jiang et al, 2014; Li et al, 2019a)。相比低海拔河流, 雅鲁藏布江流域的生态环境更为脆弱, 其生物有着明显的地理区系特点, 与其他区系重叠较少, 具有丰富而独特的基因、物种及生态系统多样性, 是全球生物多样性的热点区域(Guo et al, 2010; 拉琼等, 2014; Li et al, 2021)。
关于雅鲁藏布江流域的植物和脊椎动物已有许多报道, 然而对该河流底栖动物多样性的研究尚不充分, 主要体现在: (1)研究的空间尺度较小。关于雅鲁藏布江流域底栖动物的调查多只聚焦在局部河段(马宝珊等, 2012; 徐梦珍等, 2012; Jiang et al, 2014), 而缺乏囊括干流和重要支流的大范围、多位点的样品采集; (2)调查的时间尺度明显不足。多数研究仅基于单次采样, 缺乏连续性或季节性的调查, 所得结论不能充分反映该流域底栖动物种类及分布的真实状况(刘海平等, 2014)。此外, 由于雅鲁藏布江中下游水系复杂、生境多样, 底栖动物资源极为丰富, 以往的研究缺乏系统性, 物种需要进一步复核。在全球气候变化和河流水电开发的背景下, 开展雅鲁藏布江流域大范围底栖动物多样性调查, 摸清其家底, 是科学评价河流生态系统健康状况, 实现生物多样性保护和资源可持续开发利用的重要基础。有鉴于此, 本文于2015-2016年开展了雅鲁藏布江中下游干流(朗县至墨脱段)和19条支流底栖动物的调查工作, 力图揭示这一江段底栖动物的物种多样性, 刻画底栖动物的群落结构和时空分布格局, 并探讨影响群落结构的关键环境要素, 以期为合理利用和保护该流域的底栖动物多样性资源提供科学依据。
1 材料与方法
1.1 研究区域
雅鲁藏布江中下游地处藏东南(26°52′- 30°40′ N, 92°09′-98°47′ E), 位于地球第三极的较低位置(图1)。不同于上游地区的高寒少雨, 该区域气候温暖湿润, 年均气温8.7℃, 年均降水量650 mm左右。雅鲁藏布江中下游地区横跨多个气候带, 从中游高原温带半干旱气候向下游山地亚热带和热带湿润气候过渡。该江段河道落差极大, 蕴含丰富的水能资源, 尤其在流经雅鲁藏布江大峡谷后, 海拔落差达到2,000 m。大峡谷的存在使得上下游形成截然不同的气候、生境和植被类型, 是区域内生物多样性形成的重要基础, 亦是阻碍生物扩散和分布的天然屏障。独特的气候特征、地形地貌和多样的生态系统、丰富的生物资源, 使雅鲁藏布江中下游成为亚洲乃至北半球气候变化的调节器, 成为维系高原生态系统、生物多样性及周边地区生态平衡的重要缓冲区。
图1
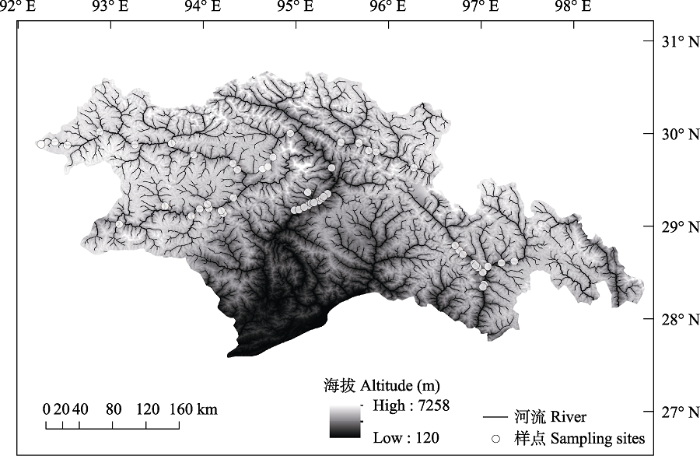
图1
雅鲁藏布江流域采样点分布图
Fig. 1
Geographical locations of the sampling sites in the Yarlung Zangbo River, China
1.2 样品采集和处理
本文于2015年10月和2016年3月对雅鲁藏布江干流(朗县-墨脱)和19条支流进行底栖动物样品的采集。采集的19条支流包括大峡谷上游的10条(拉多、金东曲、比布曲、里龙、当嘎小溪、扎绕、玉松普曲、南伊沟、尼洋河和帕隆藏布)和大峡谷下游的9条(察隅河、金珠藏布、西莫河、哈果河、西贡河、果果糖支流、甲嘎支流、白马西路河和多雄河)。共设样点75个, 其中干流样点17个, 大峡谷上游支流样点30个, 大峡谷下游支流样点28个(图1, 附录1)。
在每个样点选取流速、水深和底质组成均有代表性的地点进行定量采集, 各样点均采集3个重复样。依据样点的生境状况, 在鹅卵石-砾石底质为主的可涉水区域采用Surber网(筛网孔径420 μm, 采样面积0.09 m2)进行采集, 在水位较深的不可涉水样点用Peterson采泥器(1/16 m2)采集。同时, 使用手抄网或D型网对各样点底栖动物进行定性采集, 目的是对定量采集进行补充和完善。
在采集底栖动物样品的同时, 测定该样点的生境指标。使用美国麦哲伦手持式GPS (探险家210型)对海拔和经纬度进行测定, 使用优利德激光测距仪(LM600型)对河宽进行测量。使用美国黄泉仪器公司(Yellow Springs Instrument)的多参数水质分析仪(YSI6600)对pH值、水温、溶解氧和电导率进行测定。流速(LJD-10A型打印式流速流量仪, 北京戴美克科技有限公司)和水深(用标尺)则需要在样点所在的河段内随机选取5-10个断面测量, 并取平均值。参考Barbour等(1999)的标准, 将底质组成划分为四大类: (1)沙; (2)砾石; (3)鹅卵石; (4)大石, 通过目测估算每一种底质类型的百分比。
1.3 数据分析
基于以下公式计算群落多样性指数:
(1) Margalef丰富度指数(Margalef, 1958):
D = (S - 1)/lnN
(2) Shannon-Wiener多样性指数(Shannon, 1948):
H' = –
(3) Simpson多样性指数(Simpson, 1949):
D = 1–
(4) Pielou均匀度指数(Pielou, 1966):
J = H'/lnS
其中, S为样本中总分类单元数; Pi为物种i在样本中的相对丰度; N为样本中总个体数。
采用单因素方差分析(one-way ANOVA)比较干流、大峡谷上游支流及下游支流密度、生物量和多样性指数间的差异。如果方差分析的组间差异显著, 就进行多重比较; 如果方差齐, 就进行HSD (Tukey’s honestly significant difference)检验; 如果方差不齐, 就选用Games-Howell检验(Beckmann et al, 2005)。
使用相似性分析(analysis of similarities, ANOSIM)检测底栖动物群落结构在时间(春季和秋季)和空间(干流、峡谷上游支流、峡谷下游支流)尺度上差异的显著性, 并采用主坐标典范分析(canonical analysis of principal coordinates, CAP)对不同组间的群落差异进行可视化。为了优化分析, 在CAP分析时, 每组生物数据都进行lg(x + 1)转换, 以减少优势种类的优势度(Clarke & Warwick, 2001)。转换后的数据经Bray-Curtis不相似性测定, 建立所有样点间的等级相似矩阵。在不同相似矩阵的基础上建立CAP平面图。多样性指数的计算、 ANOSIM和CAP分析均在PRIMER 6.0中运行(Anderson, 2008)。
采用束缚型排序(constrained ordination)解析两个季度底栖动物群落结构与环境因子之间的关系。环境参数作为环境数据源, 底栖动物物种组成作为生物数据源, 构成环境因子与生物矩阵。对底栖动物群落的除趋势对应分析(detrended correspondence analysis, DCA)显示, 非线性模式(gradient length > 4 standard units)更为适合动物群落与环境因子关系的分析。因此采用典范对应分析(canonical correspondence analysis, CCA)。为了优化分析, 底栖动物群落数据进行lg(x + 1)转换, 具有较高偏相关系数的环境因子(r > 0.80)、膨胀因子(inflation factor)大于20的环境因子均被剔除, 并在分析中降低了稀有种的权重(downweighting option)。用前选(forward selection)和Monte Carlo转置法确定对底栖动物群落分布具有重要且独立作用的最小变量组合, 该最小变量组用于最终的CCA模型中。数据分析采用Canoco for Windows 4.5软件完成(ter Braak & Šmilauer, 2002)。
2 结果
2.1 群落结构
对采集的标本进行鉴定, 共发现底栖动物270种, 隶属于5门8纲20目92科(附录2)。底栖动物的优势类群为水生昆虫(246种, 占总物种数的91.11%); 此外还发现寡毛纲14种(占5.56%), 腹足纲4种、软甲纲2种, 涡虫纲、线虫纲、蛭纲、蛛形纲各1种(图2)。水生昆虫包括蜉蝣目9科24种, 襀翅目9科22种, 毛翅目20科43种, 双翅目19科123种(摇蚊科85种), 蜻蜓目5科14种, 鞘翅目11种, 半翅目5种, 鳞翅目2种以及广翅目和脉翅目各1种。
图2
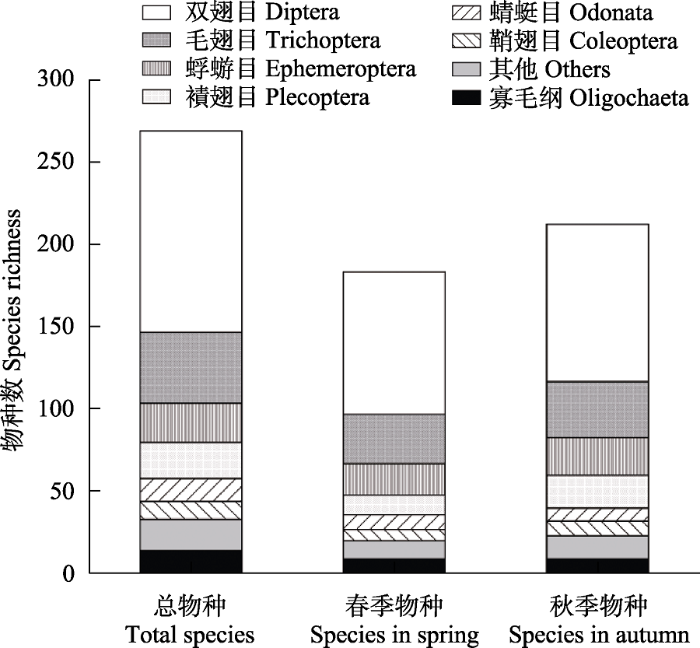
图2
雅鲁藏布江流域大型底栖动物的物种组成
Fig. 2
Species composition of benthic macroinvertebrates in the Yarlung Zangbo River
春季和秋季采集到的底栖动物组成上存在一定的差异(ANOSIM, Global R = 0.22, P < 0.001)。其中, 春季共采集到184种, 包括寡毛类9种, 软体动物2种, 水生昆虫171种和其他动物2种。优势种为四节蜉属一种(Baetis sp., 相对丰度百分比为10.00%)、蚋属一种(Simulium sp., 7.60%)、小突摇蚊属一种(Micropsetra sp., 7.50%)、短石蛾属一种(Brachycentrus sp., 5.06%)、东方帕摇蚊(Pagastia orientalis, 3.90%)以及弯握蜉属一种(Drunella sp., 3.20%)。秋季共采集到214种, 包括寡毛类9种, 软体动物3种, 软甲纲2种, 水生昆虫196种和其他动物4种(图2)。优势种为四节蜉属一种(18.65%)、花翅蜉属一种(Baetiella sp., 6.50%)、溪颏蜉属一种(Rhithrogena sp., 4.03%)、小突摇蚊属一种(3.65%)以及纽多聚石蛾属一种(Neureclipsis sp., 3.48%)。干支流各样点底栖动物类群组成如附录3所示, 虽然各断面的物种数和类群组成在两个季度略有差异, 但总体来说, 水生昆虫在春秋两季均为绝对优势的类群, 而软体动物、寡毛类以及其他底栖动物数量极少。
ANOSIM分析显示, 干流、大峡谷上游和下游支流的底栖动物群落结构在春秋两季均存在明显的空间差异(P < 0.001)。由CAP排序图可以看出, 大峡谷上游和下游支流的群落可以比较明显地区分开, 而干流群落的区分度则较差(图3)。
图3
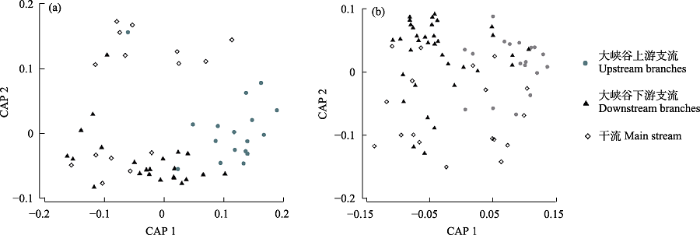
图3
雅鲁藏布江各区域底栖动物群落差异的CAP排序图。(a)春季; (b)秋季。
Fig. 3
Canonical analysis of principal coordinates (CAP) ordination diagrams of benthic macroinvertebrates in different reaches of the Yarlung Zangbo River. (a) Spring; (b) Autumn.
2.2 密度和生物量
全流域底栖动物的平均密度为939.1 ind./m2,平均生物量为5.44 g/m2。密度和生物量在各断面的差异均较大。春季干流各样点的密度为44-1,494 ind./m2,均值为477.5 ind./m2。生物量方面, 除朗县干流(14.29 g/m2)外, 其余各断面均处于较低的水平, 均值为2.12 g/m2。方差分析显示, 大峡谷上游支流(平均密度1,402.2 ind./m2;402.2 ind./m2;生物量7.05 g/m2)及大峡谷下游支流(1,075.7 ind./m2;8.24 g/m2)的密度及生物量均显著高于干流(表1)。
表1 雅鲁藏布江干支流断面底栖动物密度与生物量的比较
Table 1
| 时期 Periods | 干流 Main stream | 大峡谷上游支流 Upstream branches | 大峡谷下游支流 Downstream branches | F | P | |
|---|---|---|---|---|---|---|
| 密度 Density (ind./m2) | 春季 Spring | 477.5 ± 465.6a | 1,402.2 ± 1,080.6b | 1,075.7 ± 1,319.4b | 6.741 | 0.005 |
| 秋季 Autumn | 327.1 ± 301.7a | 1,219.2 ± 1,008.3b | 1,321.9 ± 864.6b | 5.945 | 0.007 | |
| 生物量 Biomass (g/m2) | 春季 Spring | 2.12 ± 3.69a | 7.05 ± 4.36ab | 8.24 ± 7.28b | 4.114 | 0.028 |
| 秋季 Autumn | 2.89 ± 4.68 | 5.71 ± 7.50 | 3.37 ± 2.31 | 0.872 | 0.428 |
不同字母表示存在显著差异(P < 0.05)
Different letters indicate significant differences (P < 0.05)
秋季干流底栖动物密度为56-1,217 ind./m2,均值为327.1 ind./m2;干流生物量为0.02-16.81 g/m2,干流生物量为0.02-16.81 g/m2,均值为2.89 g/m2。方差分析显示, 大峡谷上游支流(1,219.2 ind./m2)和下游支流(1,321.9 ind./m2)的密度均显著高于干流(P < 0.05), 而生物量在各组间并未表现出显著差异(表1)。
2.3 物种多样性
春季底栖动物的Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数分别在0.68-5.08 (均值为1.83)、0-2.36 (1.65)、0-0.91 (0.73)和0-1.00 (0.75)之间。支流的Margalef丰富度指数和Shannon- Wiener多样性指数均显著高于干流(P < 0.05, 图4), 而Pielou均匀度指数和Simpson多样性指数则无明显差异。
图4
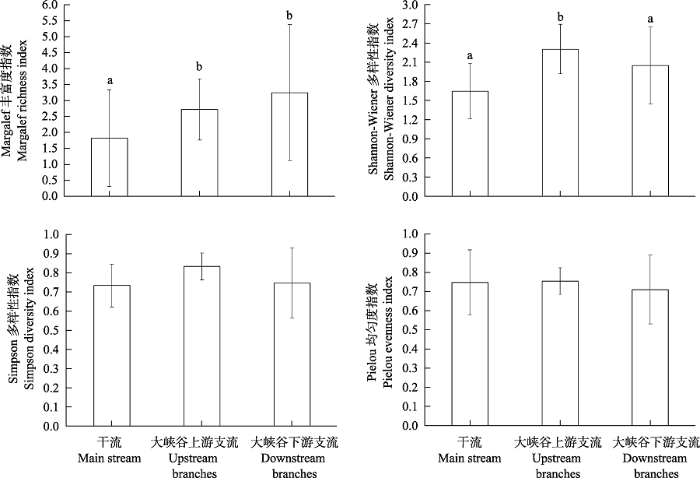
图4
雅鲁藏布江春季干支流河段底栖动物多样性指数的差异。不同字母表示存在显著差异(P < 0.05)。
Fig. 4
Comparation of diversity index of benthic macroinvertebrates in different reaches of the Yarlung Zangbo River in spring. Different letters indicate significant differences (P < 0.05).
秋季底栖动物的Margalef丰富度指数、Shannon- Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数分别在0.54-2.82 (均值为1.37)、0.75-2.20 (1.49)、0-2.36 (1.65)和0.57-0.83 (0.74)之间。除Pielou均匀度指数无显著差异外, 支流底栖动物的多样性指数均显著高于干流(P < 0.05, 图5)。
图5
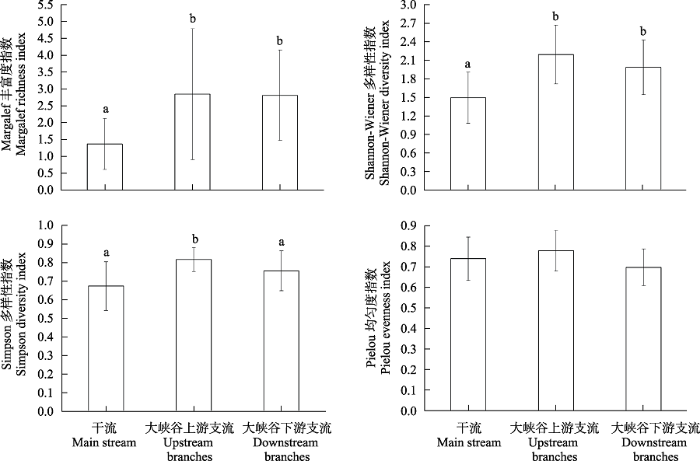
图5
雅鲁藏布江秋季干支流河段底栖动物多样性指数的差异。不同字母表示存在显著差异(P < 0.05)。
Fig. 5
Comparation of diversity index of macroinvertebrates in different regions of the Yarlung Zangbo River in autumn. Different letters indicate significant differences (P < 0.05).
2.4 关键驱动力
CCA分析结果表明, 环境因子共解释了春季底栖动物群落结构变异的19.08%。其中, 第一轴和第二轴分别解释了6.91%和4.49%。海拔、河宽、pH值和流速是影响春季底栖动物分布的关键环境因子(图6a, 表2)。寡角摇蚊属一种(Diamesa sp.)、原石蛾属一种(Rhyacophila sp.)和微动蜉属一种(Cinygmula sp.)等种类主要分布在高海拔的样点; 而脊突摇蚊属一种(Cyphomella sp.)、四节蜉属一种等种类则表现出对较高流速生境的偏好(图6a)。对于秋季来讲, 环境因子共解释了21.71%底栖动物群落结构的变异。其中, 第一轴和第二轴分别解释了4.92%和4.21%。海拔、底质(沙%)、pH值、溶解氧、水深和河宽是影响秋季雅鲁藏布江底栖动物分布的关键环境因子(图6b, 表2)。
图6
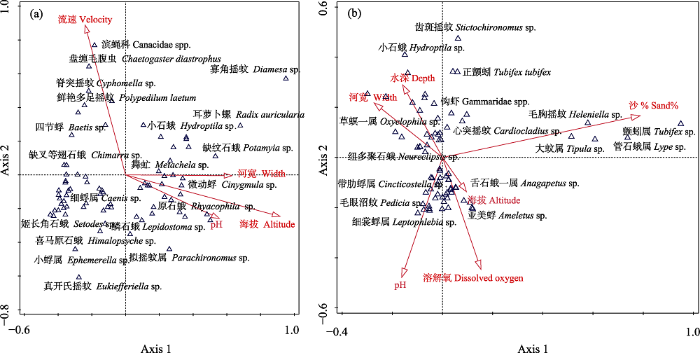
图6
雅鲁藏布江春季(a)和秋季(b)底栖动物物种与环境因子的典范对应分析排序图
Fig. 6
Canonical correspondence analysis ordination diagram of benthic macroinvertebrate communities with significant environmental variables in spring (a) and autumn (b)
表2 雅鲁藏布江底栖动物群落结构与环境因子关系的典范对应分析结果汇总表
Table 2
| 获取的关键因子 Key factors selected | 与排序轴的相关系数 Correlation coefficients with the axes | ||||||
|---|---|---|---|---|---|---|---|
| F | P | Axis 1 | Axis 2 | Axis 3 | Axis 4 | ||
| 春季 Spring | 海拔 Altitude | 1.60 | 0.001 | 0.762 | -0.183 | -0.408 | -0.182 |
| 河宽 Depth | 1.93 | 0.001 | 0.643 | 0.019 | 0.121 | 0.164 | |
| pH | 1.23 | 0.064 | 0.485 | -0.405 | 0.292 | 0.097 | |
| 流速 Velocity | 1.25 | 0.080 | -0.325 | 0.175 | 0.454 | -0.606 | |
| 秋季 Autumn | 海拔 Altitude | 1.78 | 0.001 | 0.070 | 0.266 | 0.612 | -0.521 |
| 沙% Sand% | 1.89 | 0.004 | 0.682 | 0.243 | -0.339 | -0.015 | |
| pH | 1.61 | 0.004 | -0.165 | -0.205 | -0.226 | -0.478 | |
| 溶解氧 Dissolved oxygen | 1.67 | 0.004 | -0.406 | 0.453 | -0.168 | 0.226 | |
| 水深 Depth | 1.46 | 0.052 | 0.289 | 0.123 | 0.551 | 0.244 | |
| 河宽 Width | 1.48 | 0.021 | 0.203 | -0.268 | 0.194 | -0.047 | |
3 讨论
3.1 物种组成和多样性
有关雅鲁藏布江底栖动物的调查可追溯至20世纪90年代, 武云飞和吴翠珍(1992)在《青藏高原鱼类》中记载了寡毛纲、腹足纲和昆虫纲中的一些种类。之后, 赵伟华和刘学勤(2010)报道了日喀则市雄村河段及其附近支流的底栖动物, 共计20科42属47种。徐梦珍等(2012)对位于日喀则至林芝的干流河段和部分支流进行了调查, 共记录底栖动物57科102属110种。马宝珊等(2012)报道了雅鲁藏布江中游谢通门江段底栖动物26种。Jiang等(2014)对雅鲁藏布江支流尼洋河进行了调查, 共记录38科78属94种; 刘海平等(2014)随后对尼洋河底栖动物与环境的关系进行了初步研究。简东等(2015)记录了拉萨河中下游底栖动物22科30属31种。本次调查涵盖雅鲁藏布江干流和支流样点75个, 共记录动物270种, 是近年来该流域底栖动物调查最为全面的一次。由于区域内人类活动较少, 河流基本保持原始状态, 各样点的底栖动物多以EPT类群(蜉蝣目、襀翅目和毛翅目)及其他水生昆虫的敏感种类为主。此外, 由于样点分布在海拔500-4,500 m范围内, 区域内气候、地貌、植被以及其他环境条件差异极大, 造就了丰富且复杂的底栖动物区系: 既分布着寡角摇蚊属一种(Diamesa sp.)、帕摇蚊属一种(Pagastia sp.)、同襀属一种(Isoperla sp.)、短石蛾属一种(Brachycentrus sp.)等诸多适应高海拔低温水体的种类, 又有东方春蜓属一种(Orientogomphus sp.)、缅春蜓属一种(Burmagomphus sp.)等喜好分布在中低海拔的种类。
水生昆虫(246种)是雅鲁藏布江中下游底栖动物中的优势类群, 而寡毛类、软体动物和软甲类的物种数较少。就昆虫而言, 双翅目和毛翅目的物种多样性最高, 分别记录了19科96属和20科37属(附录2)。一般认为, 双翅目和毛翅目是进化上较为高等的昆虫(Morse et al, 1994), 这些动物可通过多种生存对策(如全变态、复杂的生活史、较强的迁移能力、较高的繁殖力等)来适应不同的环境(Vieira et al, 2006)。蜉蝣目和襀翅目则是较为低等的昆虫类群, 它们一般为原变态和半变态发育, 具有相对较弱的迁移能力(一般几公里)和繁殖能力(Li et al, 2018)。但蜉蝣目(9科25属)和襀翅目(9科22属)在雅鲁藏布江水系的分化亦较为强烈, 可能与这些物种对低温和清洁水体的偏好有关。值得注意的是, 蜻蜓目主要分布在大峡谷下游, 尚未突破大峡谷的天然屏障。尽管这类动物有着较强的主动迁移能力, 但大峡谷上游的极端环境可能不适宜其生存和繁衍。与西江(软体动物70种; 苏炳之等, 1989)、赣江(65种; 邢圆等, 2019)和淮河(26种; 丁建华等, 2017)等低海拔河流不同, 软体动物在雅鲁藏布江水系的物种多样性极低, 且出现的多为机会主义者和广布种, 如腹足纲的萝卜螺属一种(Radix sp.)。一般认为软体动物的物种起源是在低海拔地区(如海洋), 随着时间的推移, 逐渐向高海拔地区扩散和物种分化(王军, 2018)。它们的主要迁移方式为通过水体的被动扩散(Li et al, 2019a), 移动能力极为有限, 难以突破大峡谷的阻隔。此外, 高海拔地区恶劣的环境(如极端低温和食物匮乏)也使这些物种难以生存和定居(Maiolini & Lencioni, 2001)。
3.2 底栖动物时空分布差异的影响因素
由CCA分析结果可以看出, 海拔均为春秋两季最为关键的环境因子(图6, 表2)。本次调查所采样点涵盖约4,000 m的海拔范围, 如此大的海拔梯度引起的一系列环境变化是导致样点间底栖动物物种组成及多样性差异的重要因素。海拔决定了一个地区的水温、溶解氧、光照、植被覆盖率等环境条件, 因而从根本上决定了底栖动物的群落结构(Jacobsen et al, 2003; Wang et al, 2010; Laursen et al, 2015)。许多研究表明, 高海拔河流中较低的水温和溶解氧, 以及稀少的岸带植被和沉水植物(造成食物来源较少)是造成底栖动物种类数和现存量低的主要原因(Milner et al, 2001; Chaves et al, 2008)。这些恶劣的环境条件对底栖动物的扩散、定居和繁衍提出了巨大的挑战(Lencioni et al, 2007; Madsen et al, 2015)。相反, 在气候温暖湿润的低海拔区域, 适宜的生境条件和岸带植被为底栖动物提供了更多的栖息地和食物来源。因此, 大峡谷下游断面(察隅和墨脱地区)的物种多样性和现存量均较高。
河流底栖动物的种类组成很大程度上取决于生境的多样性和稳定性(Heino, 2013; Milesi et al, 2016)。底质是构成局域生境异质性的主体, 也是底栖动物的生长、繁殖等一切生命活动的必备条件。底质的颗粒大小、稳定程度、表面构造和营养成分等都对底栖动物有很大的影响(Beisel et al, 2000; 段学花等, 2007)。一般来说, 底栖动物的物种多样性随底质的异质性和稳定性的增加而增加(Jiang et al, 2014; 李正飞等, 2016)。例如, 卵石等大粒径的河床底质结构稳定且异质性高, 其中存在的大量间隙能够保护动物免受外界的干扰(如洪水、天敌等), 因此底栖动物的种类组成丰富而稳定; 反之, 在底质较为细密的河床(如泥沙), 密实的底质限制了生存空间的大小, 同时也降低了氧气的可获取性(Allan, 1995), EPT等好氧类群往往较少, 而摇蚊和寡毛类等适应此环境的物种则会有较高的密度(Milesi et al, 2016; Zhang et al, 2018)。干流许多断面的底质主要为泥沙, 其生境异质性较低, 也无法为底栖动物的生存提供充足的空间, 因此物种数和现存量较低。
河宽是影响流域内底栖动物群落结构的重要因素。河宽在一定程度上能够代表河流级别(Tonkin et al, 2018)。河流的局域环境(如底质类型、营养物质和水深)会随河流级别发生一系列变化, 相应地, 群落内物种组成和多样性亦会发生明显的改变(Li et al, 2019b)。通常来讲, 河流的源头多是浅水激流, 河道狭窄, 底质类型以卵石和砾石为主, 河岸带植被凋落物丰富。底栖动物的功能摄食类群主要以凋落物和粗有机颗粒为食的撕食者(shredders)为主, 如毛翅目的沼石蛾科和襀翅目的卷襀科等。随着河流级别的增加, 植被凋落物不断被降解利用, 以细小颗粒为食的收集者(collectors)逐渐占据优势(Vannote et al, 1980), 如蜉蝣目的蜉蝣科及寡毛类等。
流速、水深和pH值对流域内底栖动物的物种组成亦起着重要作用。流速一方面塑造着局域群落底栖动物的身体外形, 使之在形态上适应流水生境, 避免被水流冲走, 同时还影响着溶解氧与营养物质的获取(Gallardo et al, 2009; Jiang et al, 2014)。此外, 一定的流速也加快了水体更新, 保证了水质的清洁, 有利于敏感物种生存①(①许晨 (2017) 长白山源头溪流大型底栖动物群落特征初步研究. 硕士学位论文, 东北师范大学, 长春.)。但流速过快会降低底栖动物的密度和多样性, 成为影响底栖动物定居和繁殖的重要限制因素(Li et al, 2019b)。水深的变化会引起一些重要环境指标(如溶解氧、水压等)的改变, 从而间接影响底栖动物的群落结构(Meng et al, 2016)。通常来讲, 底栖动物的密度、生物量和多样性均会随着水深增加而不断递减(赵永晶等, 2011; 李正飞等, 2016)。多数底栖动物生存于略偏碱性的水体中, 而酸性水体不利于底栖动物生存和繁衍。有研究表明, 当pH值在7-8之间时摇蚊幼虫生存状况最好, pH过高或过低均会影响摇蚊幼虫的正常生长(谢建春, 2001)。然而在本次调查中, 各样点的pH值多呈弱碱性(6.8-9.0之间; 均值7.9), 因此不太可能成为限制底栖动物分布的因素。推测某个重要却未被测量的环境因子可能与pH值存在较强的共线性, 因而被CCA模型筛选出来。
本文对雅鲁藏布江中下游地区进行重点调查, 初步摸清了这一区域底栖动物的家底, 可为后续生物多样性评估和生态环境监测积累重要的原始资料。总体来看, 雅鲁藏布江水系维持了极为丰富的底栖动物多样性, 但其群落结构相对简单, 主要以喜清洁和冷水的水生昆虫为主, 对抗外界胁迫和环境变化的能力相对较弱。可喜的是, 目前该区域人类活动还比较少, 基本保持了水域环境和生物群落的自然状态。然而, 全球气候变暖及流域内人类活动(如大规模的建坝行为、城镇化、外来种入侵等)的增加极易破坏包括底栖动物在内的河流生物群落的结构、功能和生物多样性, 导致不可逆转的影响。需要说明的是, 由于我国水生昆虫的分类体系尚不完善, 对雅鲁藏布江的昆虫标本主要鉴定至属级水平。相信随着我国无脊椎动物分类学知识的不断积累和完善, 我们对该流域水生昆虫物种多样性的认识水平会逐渐提高。值得注意的是, 本文仅对雅鲁藏布江中下游底栖动物与环境因子的关系进行了初步探讨。受限于野外工作条件, 多元分析中涉及的环境因子仅为现场测量的物理因子, 而水体化学指标(如总磷、总氮等)等可能影响底栖动物群落的因素则未能测量。但鉴于流域内人类干扰较弱, 生态环境保存较好, 水体的营养盐浓度通常不会成为底栖动物分布的限制因素。后续的研究可考虑更多的环境因子, 如河道形态、水体有机质含量和植被盖度等, 以便对雅鲁藏布江流域底栖动物多样性格局做出更合理的解释。
致谢
感谢中国科学院水生生物研究所的曹亮、刘飞、林鹏程和巩政博士在采样过程中的帮助和关心。感谢西藏大学农牧学院的刘海平教授在项目执行期间提供的无私帮助。
附录 Supplementary Material
附录1 雅鲁藏布江中下游大型底栖动物采样断面
Appendix 1 Sampling sections of benthic macroinvertebrates in the middle and lower reaches of the Yarlung Zangbo River
附录2 雅鲁藏布江中下游大型底栖动物物种名录
Appendix 2 Species list of benthic macroinvertebrates in the middle and lower reaches of the Yarlung Zangbo River
附录3 雅鲁藏布江流域主要干支流样点春秋两季大型底栖动物物种组成
Appendix 3 Species composition in the main stream and tributaries of the Yarlung Zangbo River basin in spring and autumn
参考文献
Animal-sediment relationships re-visited: Characterising species’ distributions along an environmental gradient using canonical analysis and quantile regression splines
DOI:10.1016/j.jembe.2008.07.006 URL [本文引用: 1]
Rapid Bioassessment for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd edn
Haplotype sharing analysis using mantel statistics
The spatial heterogeneity of a river bottom, a key factor determining macroinvertebrate communities
DOI:10.1023/A:1017094606335 URL [本文引用: 1]
Towards stressor-specific macroinvertebrate indices: Which traits and taxonomic groups are associated with vulnerable and tolerant taxa?
DOI:10.1016/j.scitotenv.2017.11.022 URL [本文引用: 1]
Guide to the freshwater aquatic microdrile oligochaetes on North America
Macroinvertebrate communities of non-glacial high altitude intermittent streams
A further biodiversity index Applicable to species lists: Variation in taxonomic distinctness
DOI:10.3354/meps216265 URL [本文引用: 1]
Community structure of macrozoobenthos and biological evaluation of water quality in the mainstream of Huai River
淮河干流大型底栖动物群落结构及水质生物学评价
Benthic macroinvertebrates communities and biodiversity in various stream substrata
典型河床底质组成中底栖动物群落及多样性
Identification Manual for the Larval Chironomidae (Diptera) of North and South Carolina
Testing the response of macroinvertebrate functional structure and biodiversity to flooding and confinement
DOI:10.4081/jlimnol.2009.315 URL [本文引用: 1]
Assessment of the genetic diversity among Glyptosternum maculatum, an endemic fish of Yarlung Zangbo River, Tibet, China using SSR markers
DOI:10.1016/j.bse.2010.12.008 URL [本文引用: 1]
Environmental heterogeneity, dispersal mode, and co-occurrence in stream macroinvertebrates
DOI:10.1002/ece3.470
PMID:23467653
[本文引用: 1]

Both environmental heterogeneity and mode of dispersal may affect species co-occurrence in metacommunities. Aquatic invertebrates were sampled in 20-30 streams in each of three drainage basins, differing considerably in environmental heterogeneity. Each drainage basin was further divided into two equally sized sets of sites, again differing profoundly in environmental heterogeneity. Benthic invertebrate data were divided into three groups of taxa based on overland dispersal modes: passive dispersers with aquatic adults, passive dispersers with terrestrial winged adults, and active dispersers with terrestrial winged adults. The co-occurrence of taxa in each dispersal mode group, drainage basin, and heterogeneity site subset was measured using the C-score and its standardized effect size. The probability of finding high levels of species segregation tended to increase with environmental heterogeneity across the drainage basins. These patterns were, however, contingent on both dispersal mode and drainage basin. It thus appears that environmental heterogeneity and dispersal mode interact in affecting co-occurrence in metacommunities, with passive dispersers with aquatic adults showing random patterns irrespective of environmental heterogeneity, and active dispersers with terrestrial winged adults showing increasing segregation with increasing environmental heterogeneity.
Are macroinvertebrates in high altitude streams affected by oxygen deficiency?
DOI:10.1046/j.1365-2427.2003.01140.x URL [本文引用: 1]
Zoobenthos community structure in the middle and lower reaches of Lhasa River
拉萨河中下游底栖动物群落结构特征分析
Longitudinal patterns of macroinvertebrate communities in relation to environmental factors in a Tibetan-Plateau River system
DOI:10.1016/j.quaint.2013.02.034 URL [本文引用: 5]
Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet, China
DOI:10.3724/SP.J.1003.2014.13186 URL [本文引用: 1]
雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释
DOI:10.3724/SP.J.1003.2014.13186
[本文引用: 1]
沿环境梯度物种多样性的分布格局及其环境影响因子的研究是生态学研究的重点内容之一。雅鲁藏布江是我国海拔最高的大河, 目前还没有针对其流域的河岸植物群落物种丰富度分布格局的系统研究。作者沿海拔梯度从雅鲁藏布江上游源头区至下游共设置了15个海拔样带, 在每个海拔样带随机取6个5 m×5 m样方, 共对90个样方的河岸植物群落物种组成和植被类型进行了调查, 共记录到238种维管束植物。DCA排序结果表明物种组成变化主要受海拔和经度控制。CCA排序区分出5种大的植被类型, 即高寒草甸或高寒草原、高山草原、两种亚高山灌丛草原和林缘灌丛植被。各植被类型的分布和变化主要受海拔和经度影响。雅鲁藏布江河岸植物物种丰富度沿海拔和经度梯度呈浅“U”形分布格局, 物种丰富度与年均降水量和植被盖度成显著正相关。
Diversity and composition of macroinvertebrate assemblages in high-altitude Tibetan streams
DOI:10.5268/IW-5.3.818 URL [本文引用: 1]
Macroinvertebrate assemblages in glacial stream systems, a comparison of linear multivariate methods with artificial neural networks
DOI:10.1016/j.ecolmodel.2006.04.028 URL [本文引用: 1]
Community structure and diversity distribution pattern of sandy plants in the middle and upper reaches of the Yarlung Zangbo River
Dispersal capacity and broad-scale landscape structure shape benthic invertebrate communities along stream networks
DOI:10.1016/j.limno.2018.06.003 URL [本文引用: 1]
Multiple facets of stream macroinvertebrate alpha diversity are driven by different ecological factors across an extensive altitudinal gradient
DOI:10.1002/ece3.4841 URL [本文引用: 3]
Disentangling the effects of dispersal mode on the assembly of macroinvertebrate assemblages in a heterogeneous highland region
DOI:10.1086/701755 URL [本文引用: 2]
Relationship between zoobenthos biodiversity and environmental factors in Nanla River
南腊河底栖动物多样性与环境因子的关系
Macrozoobenthos
DOI:10.18307/2014.0119 URL [本文引用: 2]
西藏尼洋河水生生物群落时空动态及与环境因子关系. 3. 大型底栖动物
Hydrological characteristics of Yarlung Zangbo River
雅鲁藏布江水文特征
DOI:10.11821/xb1999S1021
[本文引用: 1]
雅鲁藏布江是我国重要的国际河流,是世界上海拔最高的大江,河川径流丰沛含沙量低,水质优良,水温低,拉孜以下河段结冻。
A preliminary study on periphyton and zoobenthos in the Xaitongmoin reach of the Yarlung Zangbo River
雅鲁藏布江谢通门江段着生生物和底栖动物资源初步研究
Altitudinal distribution limits of aquatic macroinvertebrates: An experimental test in a tropical alpine stream
DOI:10.1111/een.12232 URL [本文引用: 1]
Longitudinal distribution of macroinvertebrate assemblages in a glacially influenced stream system in the Italian Alps
DOI:10.1046/j.1365-2427.2001.00849.x URL [本文引用: 1]
Mediated spatio-temporal patterns of macroinvertebrate assemblage associated with key environmental factors in the Qinghai Lake area, China
DOI:10.1016/j.limno.2015.11.005 URL [本文引用: 1]
Substrate heterogeneity influences the trait composition of stream insect communities: An experimental in situ study
DOI:10.1086/688706 URL [本文引用: 2]
Trends of macroinvertebrate community structure in glacier-fed rivers in relation to environmental conditions: A synthesis
DOI:10.1046/j.1365-2427.2001.00861.x URL [本文引用: 1]
Spatial and temporal climate variations from 1978 to 2009 and their trend projection over the Yarlung Zangbo River Basin
雅鲁藏布江流域1978-2009年气候时空变化及未来趋势研究
The measurement of diversity in different types of biological collections
DOI:10.1016/0022-5193(66)90013-0 URL [本文引用: 1]
A mathematical theory of communication
DOI:10.1002/j.1538-7305.1948.tb01338.x URL [本文引用: 1]
A survey on benthic macroinvertebrates in the Pearl River basin (Guangdong reach)
珠江水系(广东江段)底栖动物调查
CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5)
The role of dispersal in river network metacommunities: Patterns, processes, and pathways
DOI:10.1111/fwb.13037 URL [本文引用: 1]
The river continuum concept
DOI:10.1139/f80-017 URL [本文引用: 1]
A database of lotic invertebrate traits for North America
The role of macroinvertebrates in stream ecosystem function
This review focuses on some of the roles of macroinvertebrate functional groups, i.e. grazers, shredders, gatherers, filterers, and predators, in stream-ecosystem processes. Many stream-dwelling insects exploit the physical characteristics of streams to obtain their foods. As consumers at intermediate trophic levels, macroinvertebrates are influenced by both bottom-up and top-down forces in streams and serve as the conduits by which these effects are propagated. Macroinvertebrates can have can important influence on nutrient cycles, primary productivity, decomposition, and translocation of materials. Interactions among macroinvertebrates and their food resources vary among functional groups. Macroinvertebrates constitute an important source of food for numerous fish, and unless outside energy subsidies are greater than in-stream food resources for fish, effective fisheries management must account for fish-invertebrate linkages and macroinvertebrate linkages with resources and habitats. Macroinvertebrates also serve as valuable indicators of stream degradation. The many roles performed by stream-dwelling macroinvertebrates underscore the importance of their conservation.
Macroinvertebrate Diversity and Its Application in Health Bioassessment in the Chishui River
赤水河大型无脊椎动物多样性与河流生态系统健康评价
Contrasting patterns in elevational diversity between microorganisms and macroorganisms
DOI:10.1111/j.1365-2699.2010.02423.x URL [本文引用: 1]
Assessment of macrozoobenthos habitat suitability in the upper and middle reaches of the Heihe River
黑河中上游大型底栖动物栖息地适宜度评估
Water pollution and aquatic animal
水体污染与水生动物
Assessment of macrobenthos biodiversity and potential human-induced stressors in the Ganjiang River system
DOI:10.17520/biods.2018296
[本文引用: 1]
The Ganjiang River is the seventh-largest first-level tributary of the Yangtze River, and knowledge is limited about the river’s macrobenthos assemblages. Here, we carried out a comprehensive assessment of macrobenthos species diversity in the river system, and then identified the potential drivers of the observed community patterns based on combined datasets of available historical records and field investigations from 2016-2017. A total of 330 species have been recorded to date, including 138 from the historical record and an additional 267 from the 2016-2017 investigations. In particular, this river network harbors a high array of mollusk diversity, with 17 gastropods and 31 bivalves endemic to China while 32 molluscs endemic to China were recorded in the 2016-2017 investigations. The dominant species in the Ganjiang River can tolerate pollutants. The density, biomass and richness index of branches of midstream were all higher than those of main stream, branches of the upstream and branches of the downstream. The canonical correspondence analysis (CCA) showed that five environmental factors (substrate, sand-excavating, altitude, turbidity, velocity) and four spatial factors (PCNM1, PCNM11, PCNM12, PCNM15) were the key drivers structuring macrobenthos community variation. The variation partitioning analysis indicated that the environmental factors had a stronger effect on macrobenthos communities than the spatial factors. This study provides useful information to enhance the conservation of benthic biodiversity in the Ganjiang River.
赣江水系大型底栖动物多样性与受胁因子初探
DOI:10.17520/biods.2018296
[本文引用: 1]
赣江是长江的第七大支流, 孕育了极为丰富的大型底栖动物多样性, 而相关的研究明显不足。基于文献调研和2016-2017年现场调查, 本研究系统评估了赣江水系大型底栖动物多样性及其受胁因素。共记录底栖动物5门10纲27目95科204属330种(历史记录138种, 2016-2017年记录267种)。历史记录中国特有软体动物计48种(腹足类17种, 双壳类31种), 目前记录32种。优势种主要是一些耐污种和广布种。中游支流的密度、生物量和丰富度指数要高于赣江干流、上游支流和下游支流。典范对应分析结果表明, 底栖动物的分布主要受海拔、基质、流速、浊度、挖沙等环境因子以及不同尺度空间因子的驱动。偏CCA结果显示, 环境过滤对群落结构的影响高于空间过程。本研究结果可为赣江流域水生生物的保护和管理提供科学依据。
Research on assemblage characteristics of macroinvertebrates in the Yalu Tsangpo River Basin
DOI:10.5846/stxb201102220200 URL [本文引用: 2]
雅鲁藏布江流域底栖动物多样性及生态评价
Community structure and environmental determinants of macroinvertebrates in Ili River
新疆伊犁河不同生境大型底栖动物群落及其影响因素
Substrate degradation and nutrient enrichment structuring macroinvertebrate assemblages in agriculturally dominated Lake Chaohu Basins, China
DOI:10.1016/j.scitotenv.2018.01.232 URL [本文引用: 1]
Preliminary study on macrozoobenthos in Yarlung Zangbo River and its branches around Xiongcun, Tibet, China
西藏雅鲁藏布江雄村河段及其支流底栖动物初步研究
Community structure of benthic macroinvertebrates in Lake Ulungur in Xinjiang
DOI:10.18307/2011.0622 URL [本文引用: 1]
新疆乌伦古湖大型底栖无脊椎动物的群落结构

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}