


Beta多样性是指区域环境梯度下生物群落物种组成的差异, 通常用于评估群落地带分布及其环境关联性(Whittaker, 1972; Condit et al, 2002)。物种周转表示不同群落间的物种替换, 而物种得失会导致群落间物种丰富度产生差异。物种周转的产生多源于环境过滤、空间和历史事件等, 而物种丰富度差异是由于物种稀疏或其他生态过程, 如人为干扰或物理屏障导致的物种贫乏地区生物群落物种丰富度的不同(Socolar et al, 2016; 斯幸峰等, 2017)。将传统的beta多样性分解为周转和丰富度差异两个组分, 可以更好地阐明造成群落差异的潜在模式(Harrison et al, 1992; Williams, 1996; Lennon, 2001), 更好地理解保护区的群落构建机制, 并使生物多样性保护战略更加有效。当物种周转组分占比较大时, 需要保护多个地点, 而当物种丰富度差异组分占比较大时则可以优先保护物种多样性较高的地点(Gutiérrez-Cánovas et al, 2013)。
自然保护区的科学设置及保护强度有效区划的目的是通过最小的保护面积和投入, 获得最大的生物多样性的整体保护, 其中植物beta多样性是一个重要方面。地理位置与空间距离对植物beta多样性的形成具有决定作用, 这主要由生态位理论的环境过滤和中性理论的扩散限制所驱动(陈圣宾等, 2010)。土壤基质差异越大伴随物种组成差异就越大, 这反映了土壤基质的环境过滤作用(Tuomisto et al, 2003; Kraft et al, 2008; Fernandez-Going et al, 2013)。假设群落内的物种具有相同的竞争能力、出生率和死亡率, 那么由森林复杂垂直结构导致不同植物的扩散限制能够直接影响beta多样性(Hubbell, 2001; Condit et al, 2002; 李新辉等, 2016)。针对保护区设置而言, 合理区分保护位置选择、保护强度设置、土壤基质与林分结构特征对乔、灌、草多层beta多样性的影响, 将有助于最大化保护效果和更加科学地制定保护策略(Socolar et al, 2016; Wang et al, 2021)。
本研究以凉水保护区为研究对象, 分析了保护区植物beta多样性及其分解组分特征, 量化了群落结构、土壤因子、保护强度、地理地形对植物beta多样性及其分解组分的影响。主要探讨以下科学问题: (1)凉水地区乔、灌、草3层的beta多样性及其分解组分的特征如何? 哪种组分对beta多样性起主要作用? (2)凉水地区森林beta多样性及其分解组分的主要驱动因子是什么?
1 材料与方法
1.1 研究区域概况、野外调查与实验设置
凉水保护区(128°47°-128°57° E, 47°6°-47°16° N)位于黑龙江省伊春市, 小兴安岭南部达里带岭支脉东坡, 面积12,133 ha。全境为山地, 平均海拔400 m, 坡度10°-15°。温带大陆性气候, 年均降水量676 mm, 年均温-0.3℃。土壤类型主要为暗棕壤, 森林覆被率为98%。主要树种有白桦(Betula platyphylla)、黑桦(B. dahurica)、椴树(Tilia tuan)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、红皮云杉(Picea koraiensis)等。
2015年7月, 按保护强度由低到高在保护区外、实验区、缓冲区、核心区各设置20个20 m × 20 m的乔木样方(总计80个)进行调查。在每个乔木样方内, 利用网格法将样方均匀划分为16个5 m × 5 m的小样方, 随机选取2个进行灌木调查, 在每个灌木样方中随机选取1个1 m × 1 m的小样方进行草本调查。乔木层调查所有胸径 ≥ 2.5 cm的个体, 记录种类、树高、枝下高、胸径、林分郁闭度; 灌木层记录灌木的种类、株丛、株高、冠幅及地径; 草本层记录植物种名, 同时记录每个种的盖度(样方中该种覆盖面积占样方面积的比)、多度、高度。物种多度测量方法: 分别统计乔、灌、草3层的物种个体数; 对于乔木及多数灌木和草本, 直接计数总株数。对于某些匍匐茎丛生灌木, 如笃斯越桔(Vaccinium uliginosum)和越桔(V. vitis-idaea), 以其直立分株数量代表株数; 对于丛生草本, 如薹草属(Carex), 统计其丛数, 计数每丛草本株数, 通过丛数和株数的乘积估算该种植物总株数。保护强度用赋值法, 按保护区外、实验区、缓冲区、核心区顺序转为数值变量, 分别用1-4表示。
同时记录每个样方的经纬度、海拔、坡度、坡向和坡位等信息。坡向用0°-360°表示(0°表示正北方向), 分析时经过正弦转化; 样地坡位分为6个等级, 分别为平地、下坡、中下坡、中坡、中上坡、上坡, 并按此顺序转变为数值变量, 分别用1-6表示; 样地空间位置信息用每一样地中心位置的经纬度表示。林龄采用生长锥法测定(陆兆苏等, 1993)。在每个乔木样方内按五点取样法, 用锹挖0-20 cm土层土壤, 用100 cm3标准环刀取4环刀, 装袋带回实验室用于土壤理化指标测定。
附录1显示了研究地点的基本信息, 其中坡度最大可达31°, 平均8.44°, 经纬度均值分别为128.88° E、47.17° N, 范围都在0.2°左右。平均林龄115年, 树高14.25 m, 胸径18.11 cm, 灌木高为1.50 m。
1.2 Beta多样性计算与组分分解
其中, A表示两个配对样方中每个物种在两个样方间的最小多度的总和; B表示两个配对样方中样方1的总多度与A的差值; C表示样方2的总多度与A的差值。当数据类型为0, 1 (存在与否)时: A表示两个配对样方中共有物种的数量; B表示两个配对样方中样方1独有的物种数; C表示样方2独有的物种数。 物种多度差异组分和物种周转组分所占比例分别为AbDiff/ Btotal和Repl/ Btotal。beta多样性的分解通过R软件adespatial程序包中的beta.div.comp函数实现(Dray et al, 2017)。
1.3 地理地形、群落结构与土壤理化性质相关指标的获得
(1)地理地形。运用ArcGIS将80个样方的地理坐标转换为笛卡尔坐标, 并采用欧氏距离计算各样点之间的地理距离。基于各样方的笛卡尔坐标, 运用R语言中的主轴邻距法(PCNM)引入空间变量, 选择相关的PCNM特征值作为空间因子指标。在地理地形相关指标的选择上, 我们还同时测定并记录了每个样方的海拔、坡度、坡向、坡位等。
1.4 数据分析
分析乔、灌、草3层的beta多样性及其分解组分的频率分布, 并生成箱线图, 比较平均值、中位数和分布范围等。
采用Mantel检验回答群落中地理地形、土壤因子、保护强度和生物因子(群落结构)是否对植物beta多样性及其分解组分有显著影响。将地理地形、土壤因子、生物因子和保护强度中的所有因子转变为距离矩阵, 分析每个矩阵与植物beta多样性距离矩阵的显著性(显著性水平设定为0.05)。通过R语言中的Vegan程序包完成。
采用Canoco 5.0软件进行冗余分析(redundancy analysis, RDA)和方差分解分析。冗余分析以图示的方式显示地理地形、群落结构、土壤因子和保护强度对凉水保护区植物beta多样性影响的强弱。箭头长度代表某个指标对beta多样性影响程度的高低。箭头连线和beta多样性指标的夹角代表了该指标与多样性的相关性高低, 夹角越小, 相关性越高。方差分解分析中, 响应变量是beta多样性的4个不同的距离矩阵, 解释变量先分为两部分, 第一部分是非生物因子(保护强度、土壤因子、地理地形)变量, 第二部分是生物因子(群落结构)变量。非生物因子可分为三部分, 第一部分是地理地形(空间距离、海拔、坡位、坡度、坡向)变量, 第二部分是土壤因子(有机碳、全氮、碳氮比、pH值、电导率、容重、含水量)变量, 第三部分是保护强度变量; 生物因子分为两部分, 第一部分是乔木层群落结构(郁闭度、林龄、乔木树高、乔木胸径)变量, 第二部分为灌木层(灌木树高、灌木地径)变量。
2 结果
2.1 Beta多样性及其组分特征
由图1可知, 乔木层beta多样性介于0.10-1之间, 均值为0.72, 物种周转组分和物种多度差异组分均介于0-1之间, 其均值分别为0.50和0.22; 灌木层beta多样性介于0.10-1之间, 均值为0.77, 物种周转组分介于0-1之间, 均值为0.49, 物种多度组分介于0-0.94之间, 均值为0.27; 草本层beta多样性介于0.31-1.00之间, 均值为0.81, 物种周转组分介于0.07-0.99之间, 均值为0.59, 物种多度差异组分介于0-0.84之间, 均值为0.22。
图1
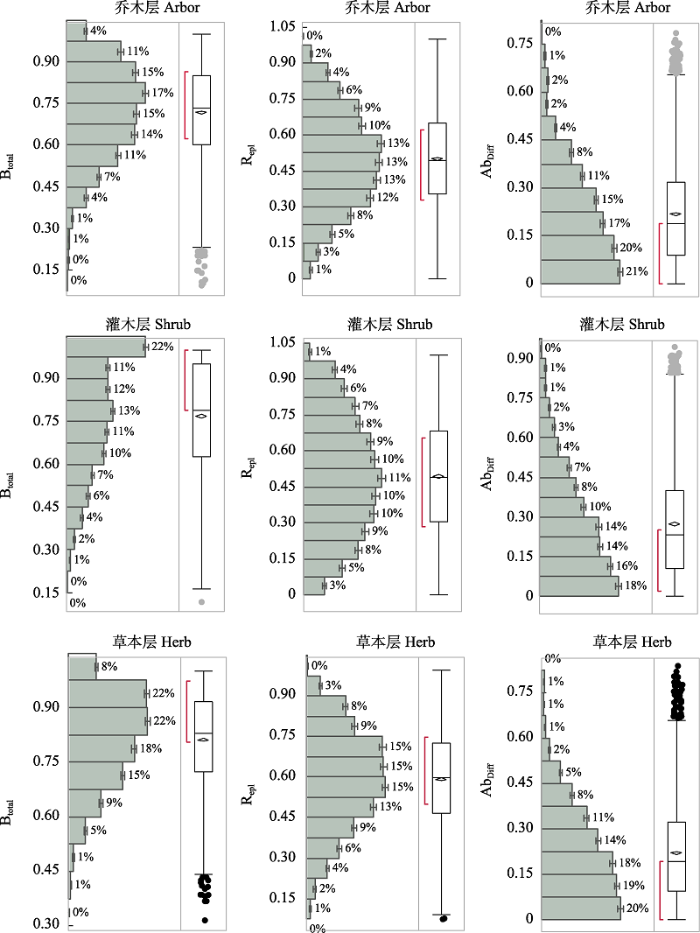
图1
Beta多样性(Btotal)及其物种周转组分(Repl)和物种多度差异组分(AbDiff)特征频率分布箱线图
Fig. 1
The relationship of taxonomic and functional trait beta diversity and its components (turnover and nestedness) of macrobenthic communities with difference in geographic distance
2.2 Beta多样性及其分解组分的相对贡献
结果表明, 在乔、灌、草中对beta多样性的贡献均是物种周转组分占主导地位(图2): 乔木层beta多样性中物种周转组分(Repl = 0.50)占比为69.6%; 灌木层(Repl = 0.49)占比为64.4%; 草本层(Repl = 0.59)占比为72.9%。
图2
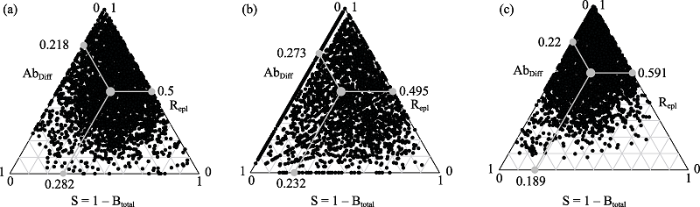
图2
乔木层(a)、灌木层(b)、草本层(c) beta多样性(Btotal)及其物种周转组分(Repl)和物种多度差异组分(AbDiff)三角图。图中每个黑点表示一个样方对, 其位置由物种组成相似性(S = 1 - Btotal)、Repl和AbDiff决定, 三者之和等于1。灰色大点表示平均值。
Fig. 2
Triangle figures for arbor (a), shrub (b), and herb (c) of plant beta diversity (Btotal), and its components of species turnover (Repl) and abundance differences (AbDiff). Each black dot represents a pair of sites. Their positions were determined by a triplet of values from the species composition similarity (S = 1 - Btotal), Repl, and AbDiff; each triplet sums to 1. The larger grey dots represent the mean values.
我们也计算了基于物种组成0, 1数据的beta多样性及其分解组分。乔、灌、草的物种多度差异分别为0.191、0.204和0.126, 物种周转组分分别是0.307、0.460和0.597 (附录2)。据此计算得到的乔、灌、草的物种周转组分贡献分别为64%、69%和83%。与基于多度的趋势基本一致, 即凉水保护区乔木层、灌木层、草本层beta多样性的两个组分中都是物种周转组分占比较大。
2.3 Beta多样性特征影响因素分析
表1 Beta多样性(Btotal)及其物种周转组分(Repl)和物种多度差异组分(AbDiff)与地理地形、保护强度、土壤因子、生物因子相关性的Mantel检验
Table 1
| 乔木层 Arbor layer | 灌木层 Shrub layer | 草本层 Herb layer | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sørensen index | AbDiff | Repl | Sørensen index | AbDiff | Repl | Sørensen index | AbDiff | Repl | |||
| 地理地形 Geography and terrain | 经纬度 Longitude and latitude | 0.079** | 0.01 | 0.06 | 0.05 | 0.02 | 0.02 | 0.06 | -0.06 | 0.1** | |
| 海拔 Altitude | 0.21*** | 0.21* | 0.01 | 0.07 | 0.07 | <0.001 | 0.18*** | 0.06 | 0.07 | ||
| 坡度 Slope | 0.2*** | -0.01 | 0.174** | 0.18** | 0.03 | 0.12* | 0.3*** | 0.05 | 0.16*** | ||
| 坡位 Slope position | 0.35*** | 0.14*** | 0.182*** | 0.18*** | 0.09* | 0.08** | 0.13*** | 0.05 | 0.05* | ||
| 坡向 Aspect | -0.08 | 0.02 | -0.08 | -0.07 | -0.08 | 0.01 | -0.01 | -0.04 | 0.03 | ||
| 保护强度 Protection intensity | 0.14*** | 0.12*** | 0.02 | 0.1** | 0.06 | 0.03 | 0.08** | -0.01 | 0.07** | ||
| 土壤因子 Soil factors | 有机碳 Soil organic carbon | 0.03 | -0.04 | 0.06 | 0.05 | 0.09 | -0.03 | -0.01 | -0.01 | <0.001 | |
| 全氮 Total nitrogen | 0.08 | 0.04 | 0.04 | 0.16** | 0.06 | 0.08 | 0.06 | 0.02 | 0.03 | ||
| C : N | -0.06 | 0.004 | -0.05 | 0.01 | -0.06 | 0.06 | -0.02 | <0.001 | -0.01 | ||
| pH | 0.05 | 0.13* | -0.06 | 0.09 | 0.03 | 0.05 | -0.04 | <0.001 | -0.03 | ||
| 电导率 Electroconductibility | 0.05 | -0.08 | 0.02 | -0.003 | 0.11 | -0.09 | -0.01 | -0.01 | <0.001 | ||
| 容重 Soil bulk density | 0.04 | 0.04 | 0.003 | 0.13* | 0.11 | 0.02 | -0.003 | -0.01 | 0.01 | ||
| 含水量 Soil water content | 0.09* | 0.07 | 0.16 | 0.21** | -0.01 | 0.18* | 0.07 | 0.02 | 0.03 | ||
| 生物因子 Biological factors | 郁闭度 Canopy density | 0.06* | 0.01 | 0.05 | 0.008 | -0.04 | 0.04 | 0.06 | 0.03 | 0.01 | |
| 林龄 Forest age | 0.28*** | 0.42*** | -0.1 | 0.08 | 0.07 | 0.004 | 0.08 | 0.02 | 0.03 | ||
| 乔木树高 Tree height | 0.24*** | -0.01 | 0.21*** | 0.14** | 0.07 | 0.05 | 0.11* | -0.06 | 0.13*** | ||
| 乔木胸径 Tree DBH | 0.18*** | 0.45*** | -0.21 | 0.05 | 0.02 | 0.03 | 0.09* | 0.03 | 0.03 | ||
| 灌木树高 Shrub height | 0.05 | 0.04 | -0.01 | 0.12* | 0.14* | -0.02 | -0.03 | 0.01 | -0.03 | ||
| 灌木地径 Shrub ground diameter | 0.02 | -0.02 | 0.03 | 0.16** | 0.17* | 0.01 | -0.08 | <0.001 | -0.06 | ||
草本层beta多样性及其物种周转组分与坡度、坡位、保护强度和乔木树高呈显著正相关(P < 0.05), 即两块样地间草本组成差异随样地间坡度、坡位、保护强度和乔木树高的差异增大而增大, 物种周转组分随空间距离的增加而增加(P < 0.05); 而草本物种多度差异组分与保护相关因子、群落结构和土壤因子均未达到显著水平(表1)。
2.4 Beta多样性特征影响因素的冗余分析排序与方差分解分析
方差分解分析结果反映了生物因子(乔木层和灌木层)、土壤因子、地理地形、保护强度对于beta多样性的解释作用。保护区整体beta多样性的变异大部分可以由地理地形解释, 达到9.2%, 其次是乔木层(7.2%)和保护强度(1.7%)。乔、灌、草3层间存在明显差异, 其中乔木层由生物因子解释的部分最大(13.1%), 约是土壤因子(1.4%)的9倍、地理地形(8.6%)的1.5倍; 灌木层由土壤因子解释的部分最大(5%), 约是地理地形的2倍; 对于草本层来说, 地理地形具有最大解释力(5.2%), 约是生物因子(1.4%)的3倍(图3)。
图3
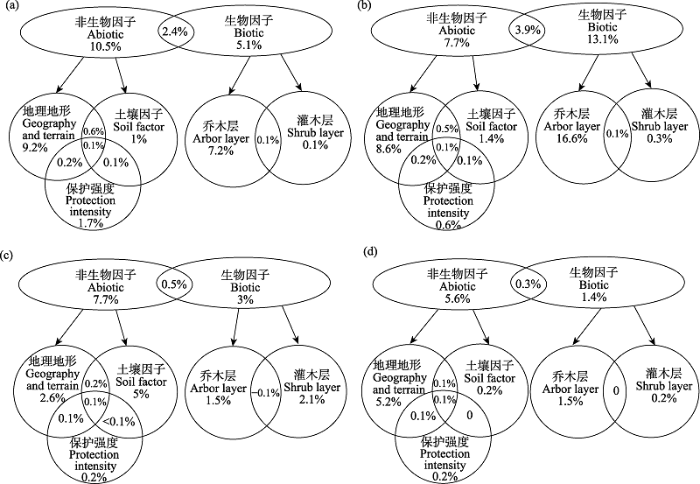
图3
地理地形、保护强度、群落结构与土壤因子对beta多样性特征解释力的方差分解分析。(a)乔、灌、草整体; (b)乔木; (c)灌木; (d)草本。
Fig. 3
Using variation partitioning to analysis the effect of geography and terrain, protection intensity, community characteristics and soil factor on beta diversity characteristics. (a) Arbor-shrub-herb; (b) Arbor; (c) Shrub; (d) Herb.
冗余分析排序结果表明, 地理地形、群落结构、土壤因子和保护强度对植物beta多样性的总解释量为14.8%, 第一轴和第二轴分别解释了多样性变化的7.35%和4.11%。在没有去除共线性的时候(简单效应, simple term effects), 乔木胸径是影响beta多样性变化的最主要因子, 解释量为3.2%, 其次是林龄, 解释量为3.1%, 而坡位和坡度的解释量分别为2.7%和2.6%。以上指标解释量均达到显著(P < 0.05); 当去除共线性影响后(条件效应, conditional term effects), 乔木胸径依然是最大解释因子, 解释量与单独作用相等; 其次是坡度, 解释量为2.5%, 乔木树高和坡位的解释量均为1.6%; 以上指标解释量均达到显著(P < 0.05)但解释量均不高(图4)。
图4
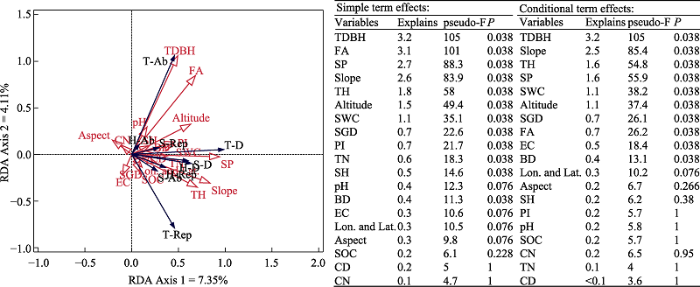
图4
地理地形、保护强度、土壤因子、群落结构与beta多样性复杂关系的冗余分析(RDA)排序分析(左)和最显著解释因子筛选(右)。T-D: 乔木层beta多样性; T-Ab: 乔木层物种多度差异组分; T-Rep: 乔木层物种周转组分; S-D: 灌木层beta多样性; S-Ab: 灌木层物种多度差异组分; S-Rep: 灌木层物种周转组分; H-D: 草本层beta多样性; H-Ab: 草本层物种多度差异组分; H-Rep: 草本层物种周转组分; Lon. and Lat.: 经纬度; Altitude: 海拔; Slope: 坡度; SP: 坡位; Aspect: 坡向; PI: 保护强度; CD: 郁闭度; FA: 林龄; TDBH: 乔木胸径; TH: 乔木树高; SH: 灌木树高; SGD: 灌木地径; SOC: 土壤有机碳; TN: 全氮; CN: 碳氮比; EC: 电导率; BD: 土壤容重; SWC: 土壤含水量。
Fig. 4
Redundancy analysis (RDA) ordination between geography and terrain, protection intensity, soil matrix, and community characteristics and beta diversity indices (left) and the most significant explaining factors (right). Btotal, Beta diversity; AbDiff, component of abundance differences; Repl, component of species turnover; T-D, Arbor Btotal; T-Ab, Arbor AbDiff; T-Rep, Arbor Repl; S-D, Shrub Btotal; S-Ab, Shrub AbDiff; S-Rep, Shrub Repl; H-D, Herb Btotal; H-Ab, Herb AbDiff; H-Rep, Herb Repl; Lon. And Lat., Longitude and Latitude; SP, Slope position; PI, Protection intensity; CD, Crown density; FA, Forest age; TDBH, Tree diameter; TH, Tree height; SH, Shrub height; SGD, Shrub ground diameter; SOC, Soil organic carbon; TN, Total nitrogen; CN, C : N; EC, Electroconductibility; BD, Soil bulk density; SWC, Soil water content.
3 讨论
3.1 凉水保护区beta多样性及其分解组分特征
目前对植物beta多样性的研究报道较多, 但对小兴安岭地区的研究还很少, 本研究是一个重要补充。在凉水保护区乔、灌、草3层beta多样性(0.72-0.81)的两个组分(Repl: 0.50-0.59; AbDiff: 0.22-0.27)中, 物种周转组分始终占比较大(65%-73%), 这表明该区域乔、灌、草beta多样性均由物种周转组分主导, 物种组成差异主要来源于样方间的物种替换, 而物种多度差异的贡献均较低。以往很多研究也得到过类似的结果(Tisseuil et al, 2012; Soininen et al, 2018; Wang et al, 2018), 如姚志良等(2020)发现, 在不同取样尺度下, 哀牢山常绿阔叶林树种beta多样性(0.43-0.75)的两个组分(Repl: 0.30-0.47; AbDiff: 0.13-0.27)中, 物种周转组分均占主导地位(> 60%)。
群落间物种周转通常由环境限制和环境过滤等生态过程共同驱动, 导致物种周转组分较高的因素较多, 如环境异质性、竞争和优先效应等(Leprieur et al, 2011)。凉水保护区针叶林、针阔叶混交林与阔叶林3种林型有乔木28-30种, 灌木22-25种, 草本78-90种; 林分多样性的差异主要由草本层控制, 并受乔木层结构和土壤理化性质的调节(王文杰等, 2019)。凉水保护区土壤理化性质存在较大的林型间差异以及季节变化(刘宏伟等, 2010); 保护区内天然形成的林隙变化对林分树种更新及物种多样性也有显著影响(刘少冲等, 2011), 这些都是影响beta多样性变化的潜在环境筛。相比于前人的研究, 本研究系统探讨了小兴安岭地区乔、灌、草3层的beta多样性及其分解组分。相关数据为小兴安岭地区天然林保护提供了数据支撑。
3.2 保护区beta多样性驱动分析及保护建议
就保护区乔、灌、草整体beta多样性而言, 地理地形对beta多样性变化的贡献最大, 其解释率约是生物因子乔木层的1.2倍。基于保护区乔、灌、草整体beta多样性的保护应该首先着重于地理地形的选择, 同时兼顾群落结构的影响。地形因子对于植物多样性的影响主要是通过影响降水、温度、湿度等环境条件间接实现的, 么旭阳等(2014)发现, 在长白山阔叶红松林中海拔对植物群落的物种多样性影响最大。以往很多研究表明, 群落结构对植物α多样性具有重要影响, 如王文杰等(2019)研究发现灌木地径、乔木胸径是导致该区域针阔混交林多样性特征差异显著的因子。本研究针对群落结构对beta多样性的影响做出了重要补充, 结果表明坡度、坡位、乔木大小、土壤含水量、保护强度与乔、灌、草beta多样性具有显著正相关关系。从本研究结果来看, 对寒温带保护区的整体保护首先要选择地理位置差异较大的区域, 其次兼顾考虑选择树木大小和土壤含水量差异较大的不同生境类型。综合比较保护强度与其他因子的作用可以发现, 保护强度对beta多样性差异的贡献仅排在所测试指标的第9-14位, 解释力远低于林分结构、土壤与某些地形特征指标(图4)。
就乔木层而言, 群落结构对乔木层beta多样性的贡献最大。基于乔木层beta多样性的保护, 在保护区位置选择上应该着重于生物因子(群落结构)差异性选择。关于群落结构影响α多样性的研究已有诸多报道, 如从非林隙到林隙中心, 植物多样性均逐渐升高(刘少冲等, 2011); 在华北落叶松林中, 郁闭度、枝下高、胸径、树高、冠幅对多样性的影响程度逐渐减小(孔令伟等, 2014)。这些研究表明, 郁闭度对多样性的影响主要是通过改变森林光照环境的方式改变物种α多样性。这可能是因为群落郁闭度和乔木树高等因子影响群落下层光照和其他生境因子分布, 郁闭度大的森林群落中光线较暗、直射到地面的阳光较少, 使得幼树的成活率低, 不利于林分更新, 增加了群落间物种组成的不确定性。因此, 对于乔木层beta多样性的保护选择郁闭度和乔木大小差异较大的区域。
就灌木层而言, 其beta多样性主要受土壤因子影响, 其解释率分别约是地理地形、生物因子的2倍。如果保护区设置主要以灌木保护为主, 应优先选择土壤理化性质差异较大的异质性区域。土壤可以提供植物生长发育所需的水分和养分, 而土壤水分可以通过影响养分的可利用性来影响植物分布(Hájek et al, 2012), 进而影响植物的beta多样性。本研究表明, 土壤含水量、容重、全氮含量与灌木层beta多样性显著正相关。提升保护区内的灌木层beta多样性, 应优先考虑具有较大水分生境及土壤全氮含量差异的区域。本研究结果显示坡位、保护强度、灌木大小均与灌木层beta多样性显著正相关, 这说明保护强度的设置对保护区植物多样性起到积极的作用。对于灌木层beta多样性的保护不仅要选择土壤理化性质差异较大的区域, 还要选择坡位差异较大(如山脊和平地)和灌木大小差异较大的区域。
就草本层而言, 其beta多样性受地理地形影响最大(5.2%)。如果保护区以保护草本植物为主, 要提高草本层beta多样性应着重考虑保护位置的选择。以往许多研究也表明, 海拔、坡度、坡位对于物种组成是一个重要的影响因素(唐志尧等, 2004; 朱彪等, 2004)。通常认为海拔、坡度、坡位对植物群落产生的作用是间接的, 它们主要通过气候、土壤、光照等因素对群落产生影响, 一般情况下气温会随着海拔的升高而逐渐下降, 而降水量和相对湿度会随着海拔的升高而增加(谭珊珊等, 2013)。光照影响各生境中植被的生长发育, 而不同坡向与坡位上的光照强弱以及受光时间存在差异。本研究结果表明, 海拔、坡位、坡度、保护强度这些地理位置和保护强度指标均与草本层beta多样性显著正相关。保护强度是人为对保护区的划分, 在不同保护强度的区域, 人为因素对植物群落的影响程度不同, 从而产生了植物多样性的差异。陈晨等(2020)研究表明, 放牧等人为干扰对科尔沁草原植物beta多样性具有重要影响。因此, 对于草本层beta多样性的保护应首先在不同的保护强度中选择海拔、坡度、坡位差异较大的区域进行。其次, 乔木大小也与草本层beta多样性显著正相关, 在考虑保护相关因子的基础上, 可以考虑乔木大小差异较大的样地进行保护。
综合来看, 林分结构、保护强度、地理地形与土壤因素对于乔、灌、草beta多样性的提升往往存在不同的影响; 这为天然林分类的经营管理提供了一个数据基础。我国实施天然林保护工程已20多年, 从本研究结果来看, 天然林保护工程对于植物beta多样性的提升起到了积极的作用。我国现已设置2,750处保护区(http://www.mee.gov.cn/searchnew/?s
earchword=自然保护区名录), 提升植物多样性的保护往往是其他生物多样性保护的一个基础。相比前人的研究, 本研究探讨保护强度对beta多样性影响, 是对该区域天然林保护和物种多样性协同提升理论研究的重要补充。
附录 Supplementary Material
附录1 样方调查区域的地理位置、土壤理化性质和林分群落结构特征
Appendix 1 The geographical information related to protection, soil properties and forest characteristics of studied plots
附录2 乔木层、灌木层、草本层beta多样性(Btotal)及其物种周转组分(Repl)和物种丰富度差异组分(RichDiff)三角图
Appendix 2 Triangle figures for arbor, shrub, and herb of plant beta diversity (Btotal), and its components of species turnover (Repl) and richness differences (RichDiff)
参考文献
Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns
DOI:10.1111/geb.2012.21.issue-7 URL [本文引用: 1]
Species diversity of plant communities and its main influencing factors in Horqin Sandy Land, Inner Mongolia of northern China
科尔沁沙地植物群落物种多样性及其主要影响因素
A review of beta diversity studies
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 1]

<p>Beta diversity is an important component of biological diversity, measuring compositional change in species assemblages across temporal and spatial scales. Beta diversity concerns not only a number of ecological and evolutionary issues, but can also guide the selection of protected areas and help to optimize conservation networks. It has thus become a hot topic in biodiversity research in recent years. Researchers have used various measures and analytical methods to investigate patterns of beta diversity and its underlying mechanisms for various taxa and in different regions. Here, we reviewed literature from the past decade pertaining to the following aspects of beta diversity: metrics, temporal and spatial patterns, determinants and applications in biodiversity conservation. Whittaker introduced the term beta diversity in 1960, but defined it vaguely. As the concept of beta diversity evolved, a high variety of measures were developed to quantify the concept. The comparison of results from different studies may be hindered by the variety of measures used to quantify beta diversity. Presently, the most popular methods for measuring beta diversity are similarity/ dissimilarity coefficients such as Jaccard index and Sørensen index. In the last few years, several methods to quantify beta diversity have emerged, some of which are worth noting. Beta diversity depends on temporal scale, spatial scale and taxonomic scale, and decreases with increasing analytical grain size. There is no consensus among scientists that beta diversity decreases with latitude, i.e. that it is higher in tropics and lower near the poles. Beta diversity is high within mountain ranges and at the interface of biogeographic realms; thus, larger/more reserves are needed in these regions to cover the entire gradient of species turnover. Studies on beta diversity across temporal scales have shown that climatic change has resulted in shifts in species composition through time, and that the migration of species between different continents/regions has led to biotic homogenization. Based on a thorough review of beta diversity literature, we think the following questions might be the focus of future research: (1) the influence of evolutionary history and biological characteristics of different taxonomic groups on their beta diversity; (2) the influence of temporal/spatial scales on beta diversity and its determinants; and (3) the effect of anthropogenic activities on beta diversity.</p>
Beta多样性研究进展
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 1]
Beta多样性度量时空尺度上物种组成的变化, 是生物多样性的重要组成部分, 与许多生态学和进化生物学问题密切相关, 并且其信息可用于保护区选址和布局规划, 因此在最近10年间成为生物多样性研究的热点问题之一。多年来, 学者们利用各种度量方式和分析方法, 在不同地理区域, 对许多生物类群beta多样性的时空格局和形成机制进行了大量研究。本文主要从beta多样性的度量方法、时空格局、形成机制及其在生物多样性保护中的应用等几个方面, 总结了最近10多年来相关研究的进展。Whittaker(1960)最初提出beta多样性概念时就缺乏严格的定义, 随着概念的不断演化, 度量方法也同样呈现出多样化, 而度量手段的多样化非常不利于不同研究之间的比较。目前应用最普遍的度量方法是采用相似性指数, 如Jaccard和Sørensen指数。最近几年, 新的度量方法还在不断出现, 其中一些方法非常值得注意。Beta多样性具有时空尺度和分类尺度依赖性, 一般随分析粒度(grain)的增加而降低。虽然有些研究表明beta多样性随纬度增加而降低, 但学者们并没有达成共识。山区和生物地理区的交界处beta多样性都比较高, 因而需要在这些地区增加保护区的面积或者数量以囊括物种变化梯度。对时间尺度上beta多样性的研究表明, 气候变化确实导致了物种组成在时间上的变化, 并且物种在不同大陆和地区间的迁移导致了生物同质化。扩散过程和生态位过程共同决定了beta多样性, 只是这两个过程的相对重要性依尺度、地理区域和物种类群的不同而有所差异。综上所述, 我们认为未来beta多样性研究的热点问题是:(1)不同生物类群的进化历史和生物学特征对beta多样性的影响; (2)不同的时空尺度对beta多样性及其维持机制的影响; (3)人类活动对beta多样性的影响。
Community composition and diversity of fungi in soils under different types of Pinus koraiensis forests
不同林型红松林土壤真菌群落组成和多样性
Beta-diversity in tropical forest trees
DOI:10.1126/science.1066854 URL [本文引用: 2]
Climate interacts with soil to produce beta diversity in Californian plant communities
Spatially distinct communities can arise through interactions and feedbacks between abiotic and biotic factors. We suggest that, for plants, patches of infertile soils such as serpentine may support more distinct communities from those in the surrounding non-serpentine matrix in regions where the climate is more productive (i.e., warmer and/or wetter). Where both soil fertility and climatic productivity are high, communities may be dominated by plants with fast-growing functional traits, whereas where either soils or climate impose low productivity, species with stress-tolerant functional traits may predominate. As a result, both species and functional composition may show higher dissimilarity between patch and matrix in productive climates. This pattern may be reinforced by positive feedbacks, in which higher plant growth under favorable climate and soil conditions leads to higher soil fertility, further enhancing plant growth. For 96 pairs of sites across a 200-km latitudinal gradient in California, we found that the species and functional dissimilarities between communities on infertile serpentine and fertile non-serpentine soils were higher in more productive (wetter) regions. Woody species had more stress-tolerant functional traits on serpentine than non-serpentine soil, and as rainfall increased, woody species functional composition changed toward fast-growing traits on non-serpentine, but not on serpentine soils. Soil organic matter increased with rainfall, but only on non-serpentine soils, and the difference in organic matter between soils was positively correlated with plant community dissimilarity. These results illustrate a novel mechanism wherein climatic productivity is associated with higher species, functional, and landscape-level dissimilarity (beta diversity).
Contrasting effects of natural and anthropogenic stressors on beta diversity in river organisms
DOI:10.1111/geb.12060 URL [本文引用: 1]
Do we need soil moisture measurements in the vegetation-environment studies in wetlands?
DOI:10.1111/jvs.2012.24.issue-1 URL [本文引用: 1]
Beta diversity on geographic gradients in Britain
DOI:10.2307/5518 URL [本文引用: 1]
Relationships among growth of Larix principis-rupprechtii, herbaceous plants diversity and landform
华北落叶松林木生长、草本植物多样性及地形因子之间的关系
Functional traits and niche-based tree community assembly in an Amazonian forest
DOI:10.1126/science.1160662 URL [本文引用: 1]
Interpreting the replacement and richness difference components of beta diversity
DOI:10.1111/geb.12207 URL [本文引用: 1]
The geographical structure of British bird distributions: Diversity, spatial turnover and scale
DOI:10.1046/j.0021-8790.2001.00563.x URL [本文引用: 1]
Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes
DOI:10.1111/j.1461-0248.2011.01589.x
PMID:21303436
[本文引用: 1]
Here, we employ an additive partitioning framework to disentangle the contribution of spatial turnover and nestedness to beta diversity patterns in the global freshwater fish fauna. We find that spatial turnover and nestedness differ geographically in their contribution to freshwater fish beta diversity, a pattern that results from contrasting influences of Quaternary climate changes. Differences in fish faunas characterized by nestedness are greater in drainage basins that experienced larger amplitudes of Quaternary climate oscillations. Conversely, higher levels of spatial turnover are found in historically unglaciated drainage basins with high topographic relief, these having experienced greater Quaternary climate stability. Such an historical climate signature is not clearly detected when considering the overall level of beta diversity. Quantifying the relative roles of historical and ecological factors in explaining present-day patterns of beta diversity hence requires considering the different processes generating these patterns and not solely the overall level of beta diversity.© 2011 Blackwell Publishing Ltd/CNRS.
Impacts of geographical distances and environmental differences on the beta diversity of plant communities in the dry-hot valley of the Yuanjiang River
DOI:10.17520/biods.2015245 URL [本文引用: 1]
地理距离及环境差异对云南元江干热河谷植物群落beta多样性的影响
DOI:10.17520/biods.2015245
[本文引用: 1]
beta多样性反映了群落间物种组成的差异, 是生物多样性研究的热点之一。本研究通过对云南元江干热河谷41个植物群落样方进行调查, 用Jaccard相异系数表征物种beta多样性, 用样方之间的最近谱系距离(mean nearest taxon distance, MNTD)及平均谱系距离(mean pairwise distance, MPD)表征谱系beta多样性, 采用基于距离矩阵的多元回归和方差分解方法, 探讨了该区域干热河谷典型植物群落的物种beta多样性和谱系beta多样性与样方间环境差异(主要是气候)及地理距离之间的关系。结果表明: (1)群落间的地理距离和年平均温度差异对干热河谷植物群落的物种beta多样性和谱系beta多样性有显著影响; (2)地理距离对物种beta多样性和MNTD的影响最大; 地理距离和年平均温度差异对MPD的影响均较大; (3)样方间年平均温度与年平均降水量的差异和地理距离能够解释群落间beta多样性及谱系beta多样性11-13%的变异。以上结果表明, 生态位分化和扩散限制对该地区植物群落的beta多样性均有显著影响, 其中扩散限制的影响可能更大。此外, 人类活动等其他因素也很可能对元江干热河谷的群落组成具有非常重要的影响。
Seasonal dynamics of soil phosphorus for main forest types in Liangshui Nature Reserve and its availability
凉水自然保护区主要森林类型土壤磷季节动态及有效性
Effects of forest gap on tree species regeneration and diversity of mixed broadleaved Korean pine forest in Xiaoxing’an Mountains
林隙对小兴安岭阔叶红松林树种更新及物种多样性的影响
Use the growth cone-like wood method to determine the number of stands at maturity
用生长锥样木法确定林分数量成熟龄
A new conceptual and methodological framework for exploring and explaining pattern in presence-absence data
DOI:10.1111/j.1600-0706.2011.19451.x URL [本文引用: 1]
Beta-diversity partitioning: Methods, applications and perspectives
DOI:10.17520/biods.2017024 URL [本文引用: 1]
Beta多样性分解: 方法、应用与展望
DOI:10.17520/biods.2017024
[本文引用: 1]
Beta多样性是指不同群落间物种组成的差异, 由物种周转(或物种替换)和嵌套(或丰富度差异)这两种过程决定。Beta多样性分解是将这两种过程对总体beta多样性的作用进行拆分, 然后分别探讨这两种过程对群落间物种组成差异的影响。2010年之后, 人们提出了beta多样性分解的方法, 其中占据主导地位的是由Andrés Baselga于2010年提出的BAS法(总体beta多样性分解为物种周转和嵌套组分)和由János Podani和Dénes Schmera于2011年以及José C. Carvalho等于2012年提出的POD法(总体beta多样性分解为物种替换和丰富度差异组分)。这两种分解方法引起了持续的争论, 促进了该领域的快速发展。作者归纳分析了2010年后有关beta多样性分解的文献后发现, 使用BAS法的论文无论在发表量和引用次数上都多于POD法(75% vs. 20%)。Beta多样性分解的研究主要集中在欧洲(45%), 研究类群则以动物(64%)为主。本文在回顾beta多样性分解方法的提出及其发展过程的基础上, 从时空维度(纬度梯度、海拔梯度、生境片断化过程以及季节和年际动态)、多样性的不同方面(物种、功能和谱系多样性)和不同生物类群之间的比较等研究角度出发, 进一步阐述了beta多样性分解方法在探讨生物多样性分布格局以及形成机制中的应用。对于beta多样性分解的研究, 我们认为需要深入探讨的问题有: (1) beta多样性分解方法的比较分析和整合; (2)结合物种多度信息探讨beta多样性及其组分的分布格局; (3)对大尺度下beta多样性分解的结果进行普适性验证。
How should beta-diversity inform biodiversity conservation
DOI:S0169-5347(15)00289-X
PMID:26701706
[本文引用: 2]
To design robust protected area networks, accurately measure species losses, or understand the processes that maintain species diversity, conservation science must consider the organization of biodiversity in space. Central is beta-diversity--the component of regional diversity that accumulates from compositional differences between local species assemblages. We review how beta-diversity is impacted by human activities, including farming, selective logging, urbanization, species invasions, overhunting, and climate change. Beta-diversity increases, decreases, or remains unchanged by these impacts, depending on the balance of processes that cause species composition to become more different (biotic heterogenization) or more similar (biotic homogenization) between sites. While maintaining high beta-diversity is not always a desirable conservation outcome, understanding beta-diversity is essential for protecting regional diversity and can directly assist conservation planning.Copyright © 2015 Elsevier Ltd. All rights reserved.
A meta-analysis of nestedness and turnover components of beta diversity across organisms and ecosystems
DOI:10.1111/geb.2018.27.issue-1 URL [本文引用: 1]
Beta diversity of plant communities in Baishanzu Nature Reserve
DOI:10.5846/stxb URL [本文引用: 1]
百山祖自然保护区植物群落beta多样性
Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains
DOI:10.17520/biods.2004014 URL [本文引用: 1]
秦岭太白山木本植物物种多样性的梯度格局及环境解释
DOI:10.17520/biods.2004014
[本文引用: 1]
物种多样性沿环境梯度的分布格局是生物多样性研究的重要议题,而海拔梯度包含了各种环境因子的综合影响,因此对于探讨物种多样性沿环境梯度的分布格局具有重要意义。秦岭山脉地处我国暖温带向亚热带的过渡带,其植被垂直带和物种多样性分布格局对于研究我国南北部植被分异特征具有重要意义。基于对秦岭山脉太白山南北坡海拔1200–3750m之间的垂直样带调查的83个样方,本文利用植被数量分析方法(DCA和TWINSPAN)和Shannon-Wiener多样性指数、Pielou指数以及Jaccard相异性系数对太白山木本植物物种多样性在南北坡沿海拔梯度分布格局进行了初步研究。结果表明:太白山的木本植物群落具有明显的环境梯度格局,海拔是决定太白山植物群落分布的主要因素,而坡向起到次要作用。植物群落类型与坡向的关系不大,当考虑群落的环境梯度格局时,DCA第一轴主要与年均温密切相关,而第二轴则取决于年平均相对湿度。乔木层和灌木层的物种具有相似的海拔梯度格局,植物群落中木本植物物种丰富度和多样性随着海拔的升高单调下降;群落均匀度随着海拔变化的规律不明显;灌木层的物种多样性比乔木层更为丰富,而南坡具有比北坡更多的物种数和更高的多样性。相邻海拔之间群落的相异性在南北坡具有不同的分布格局,在北坡2800 m以下,群落相异性沿海拔梯度变化不大,而在2800 m以上的高海拔地区,群落相异性随海拔的升高而降低; 在南坡,随着海拔的升高,群落相异性不断减少。太白山南坡群落比北坡分布更连续。
Projected impacts of climate change on spatio-temporal patterns of freshwater fish beta diversity: A deconstructing approach
DOI:10.1111/j.1466-8238.2012.00773.x URL [本文引用: 1]
Dispersal, environment, and floristic variation of western Amazonian forests
The distribution of plant species, the species compositions of different sites, and the factors that affect them in tropical rain forests are not well understood. The main hypotheses are that species composition is either (i) uniform over large areas, (ii) random but spatially autocorrelated because of dispersal limitation, or (iii) patchy and environmentally determined. Here we test these hypotheses, using a large data set from western Amazonia. The uniformity hypothesis gains no support, but the other hypotheses do. Environmental determinism explains a larger proportion of the variation in floristic differences between sites than does dispersal limitation; together, these processes explain 70 to 75% of the variation. Consequently, it is important that management planning for conservation and resource use take into account both habitat heterogeneity and biogeographic differences.
Understory β diversity differences and influential factors between Pinus tabuliformis plantation and natural Quercus wutaishanica forest on the Loess Plateau
黄土高原油松和辽东栎林下植物β多样性差异及影响因素
Differences in plant composition and forest structure among of 3 forest types in Liangshui National Nature Reserve
凉水自然保护区3种森林类型的植物组成和林分结构特征
Relating macrofungal diversity and forest characteristics in boreal forests in China: Conservation effects, inter-forest-type variations, and association decoupling
Ecological drivers of spatial community dissimilarity, species replacement and species nestedness across temperate forests
DOI:10.1111/geb.2018.27.issue-5 URL [本文引用: 1]
Evolution and measurement of species diversity
DOI:10.2307/1218190 URL [本文引用: 1]
Functional diversity of typical broad-leaved Korean pine forest communities in Changbai Mountains and its relationship with topographical factors
长白山阔叶红松林典型森林群落功能多样性及其与地形因子的关系
Driving forces underlying the beta diversity of tree species in subtropical mid-mountain moist evergreen broad-leaved forests in Ailao Mountains
DOI:10.17520/biods.2019356 URL [本文引用: 2]
哀牢山亚热带中山湿性常绿阔叶林树种beta多样性格局形成的驱动力
DOI:10.17520/biods.2019356
[本文引用: 2]
Beta多样性通常指群落在时间和空间上物种组成的差异, 包括物种周转组分和物种丰富度差异组分。驱动beta多样性格局形成的生态过程决定了群落的时空动态, 然而关于beta多样性及其两个组分格局形成的驱动力还存在较多争议。以往研究表明, beta多样性的格局存在取样尺度的依赖性, 驱动其形成的生态过程在不同取样尺度下的相对重要性也随之改变。本研究以哀牢山亚热带中山湿性常绿阔叶林20 ha动态监测样地为研究对象, 在不同取样尺度上, 将样方间的Bray-Curtis指数分解为物种周转组分和物种丰富度差异组分, 通过典范冗余分析和方差分解的方法揭示环境过滤和扩散限制对于beta多样性及其两个组分格局形成的相对重要性及其尺度依赖性。结果表明: (1) beta多样性、物种周转组分和物种丰富度差异组分均随取样尺度的增大而减小。在不同取样尺度下, 物种周转组分对于beta多样性的贡献始终占主导地位。(2)随着取样尺度的增大, 环境过滤驱动beta多样性格局形成的相对重要性逐渐增加, 而扩散限制的相对重要性逐渐降低。本研究进一步证实了取样尺度在beta多样性格局形成及其驱动力定量评价中的重要性, 今后的研究需要进一步解析上述尺度效应的形成机制。
Differences in community characteristics, species diversity, and their coupling associations among three forest types in the Huzhong area, Daxinganling Mountains
大兴安岭呼中地区3种林分的群落特征、物种多样性差异及其耦合关系
Characteristics of plant diversity and community structure in Shuanghe Nature Reserve in Daxing’anling area of northeastern China
大兴安岭双河保护区植物多样性和群落结构特征分析
Spatial distribution of soil total nitrogen in Liangshui National Nature Reserve based on local model
Based on LiDAR data of Liangshui National Nature Reserve, digital elevation model (DEM) was constructed and both primary terrain attributes (slope, aspect, profile curvature, etc.) and secondary terrain attributes (wetness index, sediment transport index, relative stream power index, etc.) were extracted. According to the theory of soil formation, geographically weighted regression (GWR) was applied to predict soil total nitrogen (TN) of the area, and the predicted results were compared with those of three traditional interpolation methods including inverse distance weighting (IDW), ordinary Kriging (OK) and universal Kriging (UK). Results showed that the prediction accuracy of GWR (77.4%) was higher than that of other three interpolation methods and the accuracy of IDW (69.4%) was higher than that of OK (63.5%) and UK (60.6%). The average of TN predicted by GWR reached 4.82 g. kg-1 in the study area and TN tended to be higher in the region with higher elevation, bigger wetness index and stronger relative stream power index than in other areas. Further, TN also varied partly with various aspects and slopes. Thus, local model using terrain attributes as independent variables was effective in predicting soil attribute distribution.
基于局域模型的凉水国家自然保护区土壤全氮空间分布
Plant community composition and tree species diversity on eastern and western Nanling Mountains, China
DOI:10.17520/biods.2004007 URL [本文引用: 1]
南岭东西段植物群落物种组成及其树种多样性垂直格局的比较
DOI:10.17520/biods.2004007
[本文引用: 1]
根据沿海拔梯度设置的样地的调查资料,对南岭东段的莽山和西段的猫儿山植物群落乔木层物种多样性的垂直分布格局进行了比较研究。莽山和猫儿山的植被类型差异不大,但相同植被类型的物种组成有比较明显的差异。随海拔的上升,两山体乔木的最大树高均呈现显著下降趋势。猫儿山乔木的最大胸径、胸高断面积之和以及立木密度在海拔梯度上呈单峰曲线变化趋势。莽山乔木的最大胸径、胸高断面积之和与海拔呈明显的负相关性,而立木密度的变化规律不明显。这种单峰曲线格局或负相关关系与中尺度上的海拔高度以及小尺度上的土壤特性、小地形以及人为干扰强度等因素有关。在海拔1000 m以下的常绿阔叶林群落和1700 m以上的山顶矮林群落,莽山乔木的物种丰富度均明显高于猫儿山。两山体的乔木种数与海拔之间都显示出负相关性,但莽山由于高海拔群落受人为干扰的影响,这种相关性不显著。乔木层α多样性沿海拔梯度没有表现出明显的变化规律,并且在两山体之间也看不出明显的差别。本文最后还对两山脉低海拔区段的常绿阔叶林群落的物种组成和多样性进行了对比分析,结果表明人为干扰导致猫儿山常绿阔叶林的物种丰富度(S)和α多样性(H')明显低于莽山的原生性常绿阔叶林群落,而且落叶阔叶树种和毛竹(Phyllostachys pubescens)开始入侵到群落中。
Characteristics of canopy disturbance for a typical broadleaf-Korean pine mixed forest in Xiaoxing’an Mountains, Liangshui, northeastern China
小兴安岭凉水典型阔叶红松林林冠干扰特征分析

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}