


物种的时空分布及其驱动因素是生态学和保护生物学的重要研究内容(Connell, 1983; Case & Bolger, 1991)。环境变量和动物间的相互作用(如捕食关系、竞争关系等)是决定物种分布的重要因素(Ritchie et al, 2009)。捕食关系是自然生态系统中十分重要和复杂的生态关系, 影响着捕食者和被捕食者的空间分布。Creel和Winnie (2005)在美国西北部的研究发现, 当狼(Canis lupus)群出现时, 马鹿(Cervus elaphus)会通过减少在草地上的觅食活动并进入安全的林地来躲避狼的捕食。Rabelo等(2019)对美洲豹(Panthera onca)栖息地选择的研究结果表明猎物密度可以影响美洲豹的栖息地选择, 美洲豹更倾向于使用其树栖类猎物红吼猴(Alouatta seniculus)和褐喉树懒(Bradypus variegatus)数量较多的区域。当同一地区存在多种捕食者或多种猎物, 且捕食者对不同种类猎物的捕食偏好不同时, 不同种类猎物对捕食者空间分布的影响也不同。Steinmetz等(2013)对虎(Panthera tigris)的研究表明, 其对生境位点的使用主要受印度野牛(Bos gaurus)、水鹿(Cervus unicolor)、野猪(Sus scrofa)和印度麂(Muntiacus muntjak)等4种猎物丰富程度的影响, 由于虎对野猪这一猎物的捕食偏好, 野猪的数量与虎对位点的使用率存在显著正相关关系。也有研究认为捕食者与被捕食者的空间分布关系是捕食关系和环境因子共同作用的结果, Magle等(2014)对郊狼(Canis latrans)与白尾鹿(Odocoileus virginianus)空间分布的研究表明不同环境因子对郊狼和白尾鹿位点使用情况的影响不同, 郊狼的占域率和白尾鹿的占域率存在显著正相关关系。尽管在“生态恐惧”的理论框架下, 白尾鹿会避开有郊狼的区域, 但由于优质生存环境稀少的原因, 白尾鹿仍会选择有郊狼的区域生存。Murphy等(2019)在马达加斯加对外来捕食者与27个猎物物种的空间分布关系研究结果显示, 当地居民放养的狗(Canis familiaris)、小灵猫(Viverricula indica)、野猫(Felis silvestris)会影响当地猎物马岛鼠(Nesomys spp.)、大马岛猬(Setifer setosus)、马岛林秧鸡(Mentocrex kioloides)的空间分布, 且这种影响会随着环境的变化而变化。虽然已经有一些研究, 但有关作为捕食动物主要食物来源的雉类与捕食者间的空间分布关系及其驱动因素的研究仍然较少。
赤狐(Vulpes vulpes)属食肉目犬科, 广泛分布于欧亚大陆各种生境, 主要选择食物丰富度较高的植被和生境类型(李路云等, 2014)。赤狐是川西高原常见的捕食动物, 主要以啮齿动物、鸡形目鸟类和野兔等为食(马勇等, 2014)。鸡形目鸟类生活史大多数时间局限于地面, 飞行和扩散能力较弱, 是典型的地栖型鸟类。黄喉雉鹑(Tetraophasis szechenyii)、血雉(Ithaginis cruentus)和白马鸡(Crossoptilon crossoptilon)是川西高原具有代表性的鸡形目鸟类, 主要栖息于亚高寒地区的针阔混交林、针叶林、高山灌丛及高山草甸中, 具有相似的生活习性和资源需求。尽管3种雉类间存在一定的种间竞争, 但已有研究表明雉类的种间竞争并不影响其共存模式(Chen & Luiselli, 2009; Wang et al, 2021)。本研究利用红外相机数据对黄喉雉鹑、血雉和白马鸡与其捕食者——赤狐分别建立条件型双物种占域模型(conditional two-species occupancy model)研究其在相机位点尺度上的空间分布关系, 分析影响川西高原3种雉类空间分布的生物和非生物因素, 探究3种雉类与捕食者赤狐的空间分布关系及驱动因素, 为物种空间分布关系的研究提供新的案例。
1 研究方法
1.1 研究区域概况
研究区域位于川西高原中部四川省甘孜藏族自治州新龙县和白玉县境内, 地理坐标为98°36°- 100°54° E, 30°22°-31°40° N, 海拔2,587-5,870 m。属于高原季风气候, 昼夜温差大, 垂直气候变化显著。植被群落主要有河谷旱生灌丛、针阔叶混交林、亚高山针叶林、高山灌丛、高山草甸和流石滩(邓林和杨海真, 2010; 游章强等, 2014)。
1.2 红外相机布设
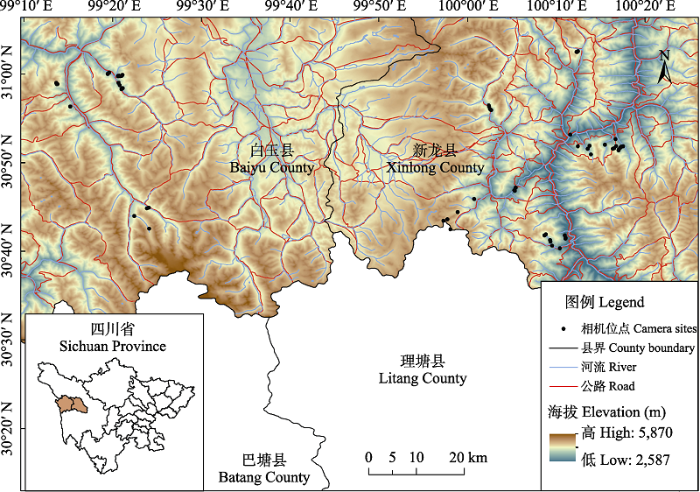
根据研究区域内的地形和植被因素选择野生动物丰富的区域, 将调查区域划分为2 km × 2 km的网格, 在考虑交通、地形条件下, 选取有森林分布的网格内布设1-3台红外相机(图1)。相机布设在动物痕迹较多的地点(如兽径、山脊等), 以最大限度提高物种的探测概率, 海拔范围在3,120-4,843 m。相机固定在距地面30-60 cm的树干上, 全天24 h工作, 参数设置为连续拍3张照片, 时间间隔为1 s。记录每台相机的GPS坐标(朱博伟等, 2019)。
图1
1.3 空间分布关系分析
红外相机和占域模型的结合是在考虑环境条件的情况下研究物种共存问题的一种有效手段(Burton et al, 2015; D’Amen et al, 2018)。本研究使用条件型双物种占域模型(Richmond et al, 2010)来探究3种雉类和其捕食者赤狐的空间分布关系。占域模型可以利用红外相机捕获到的物种出现或未出现情况, 在考虑不完全探测的情况下同时分析物种相互作用和环境因子对物种空间分布的影响(MacKenzie et al, 2004; Chen et al, 2019)。条件型双物种占域模型包含8个基本参数(表1)。根据条件型双物种占域模型的基本假设, 我们将赤狐作为优势物种(物种A), 3种雉类作为从属物种(物种B)。有研究表明赤狐的家域半径为1.5 km (Lesmeister et al,2015), 本研究的相机距离不大于其家域半径, 因此我们放宽了占域模型通常需要地理封闭的假设, 定义模型中的“占域”为“单个相机位点的使用” (Farris et al, 2016)。
表1 条件型双物种占域模型中的参数描述
Table 1
| 参数 Parameters | 描述 Description |
|---|---|
| psiA | 物种A对位点使用的概率 Probability of site use for species A |
| psiBA | 当物种A存在时, 物种B对位点使用的概率 Probability of site use for species B, given species A is present |
| psiBa | 当物种A不存在时, 物种B对位点使用的概率 Probability of site use for species B, given species A is absent |
| pA | 当物种B不存在时, 物种A的探测概率 Probability of detection for species A, given species B is absent |
| pB | 当物种A不存在时, 物种B的探测概率 Probability of detection for species B, given species A is absent |
| rA | 当两物种都存在时, 物种A的探测概率 Probability of detection for species A, given both species are present |
| rBA | 当两物种都存在且物种A被探测到时, 物种B的探测概率 Probability of detection for species B, given both species are present and species A is detected |
| rBa | 当两物种都存在且物种A未被探测到时, 物种B的探测概率 Probability of detection for species B, given both species are present and species A is not detected |
以10天作为1次探测周期对研究物种分别建立观测历史, 本次调查期间共进行了48次探测。将海拔、坡度、坡向、植被增强型指数(enhanced vegetation index, EVI)、除赤狐外其他食肉动物的相对多度指数(relative abundance index, RAI)、相机位点距最近河流距离和相机位点距最近道路距离7个变量作为占域协变量, 将每个探测周期中的具体探测天数作为探测协变量。海拔、坡度、坡向和植被增强型指数从地理空间数据云 (
为了简化候选模型的数量, 我们首先分别对每个物种建立单物种占域模型, 选取最优模型中的占域和探测协变量作为每个物种的最佳占域和探测协变量, 然后使用各物种的最佳占域和探测协变量对3个物种对建立条件型双物种占域模型(Richmond et al, 2010)。
我们分别对每个物种对建立了8个候选模型(附录1), 然后用赤池信息量准则(AIC)来对模型进行排序, 同时选择ΔAIC ≤ 2且模型权重最高的模型作为最优模型来提取各个参数并计算物种相互作用因子(species interaction factor, SIF)的估计值(Murphy et al, 2019)。物种相互作用因子表示两个物种共同出现的关系, 当SIF = 1时, 两物种的空间分布相互独立; 当SIF < 1时, 两物种在相机位点相比独立的假设下更不容易共同出现, 空间分布趋于分离; 当SIF > 1时, 两物种在相机位点相比独立的假设下更容易共同出现, 空间分布趋于重合(Richmond et al, 2010)。所有分析均在软件PRESENCE 2.12.43 (Hines, 2006)中完成。
2 结果
在2016年9月至2017年5月和2017年8月至2018年4月期间, 共设置了84个相机位点, 累计监测8,679个相机工作日, 共获得独立照片2,817张, 其中目标物种独立照片682张。拍到黄喉雉鹑的相机位点有31个(36.9%), 独立照片117张; 拍到血雉的相机位点有37个(44.1%), 独立照片175张; 拍到白马鸡的相机位点有49个(58.3%), 独立照片312张; 拍到赤狐的相机位点有25个(29.8%), 独立照片78张。在研究区域中还监测到其他9种食肉动物, 分别为金钱豹(Panthera pardus)、豹猫(Prionailurus bengalensis)、猞猁(Lynx lynx)、荒漠猫(Felis bieti)、兔狲(Otocolobus manul)、金猫(Catopuma temminckii)、狼、黄喉貂(Martes flavigula)、黄鼬(Mustela sibirica)。
2.1 赤狐与三种雉类的空间关系
4个物种的单物种占域模型结果表明, 赤狐的最优模型显示其对位点的使用概率不受占域协变量的影响。黄喉雉鹑的最优模型中占域协变量为植被增强型指数(EVI)和距最近河流距离; 白马鸡的最优模型中占域协变量为海拔和距最近道路距离; 血雉的最优模型中占域协变量为距最近河流距离(附录2)。
条件型双物种占域模型的最优模型结果显示, 赤狐对3种雉类位点使用概率的影响不同, 但都受到了环境变量的影响(表2)。在环境变量的介导下, 血雉在赤狐存在时对位点的使用概率显著高于赤狐不存在时(psiBA = 0.81 ± 0.30, psiBa = 0.29 ± 0.17, P < 0.01), 赤狐和血雉在研究区域内空间分布趋于重合(SIF = 1.31 ± 0.14) (表3)。赤狐和血雉随距河流距离的增加, 空间关系呈现先趋于重合后趋于分离的趋势(图2A)。黄喉雉鹑在赤狐存在时对位点使用概率也显著高于赤狐不存在时(psiBA = 0.70 ± 0.08, psiBa = 0.31 ± 0.38, P < 0.01)。赤狐和黄喉雉鹑在研究区域内空间分布趋于重合(SIF = 1.42 ± 0.41) (表3)。赤狐和黄喉雉鹑的空间关系随着距河流距离的增加呈现出由重合转为分离的趋势(图2B)。
表2 赤狐与3种雉类的条件型双物种占域模型选择结果
Table 2
| 模型 Models | 参数数量 Number of parameters | AIC | ΔAIC | AIC Wt | ||
|---|---|---|---|---|---|---|
| 赤狐 + 白马鸡 Vulpes vulpes + Crossoptilon crossoptilon | ||||||
| psiA, psiB (ELE + DTR); pA, pB, rB | 7 | 1,120.73 | 0 | 0.9731 | ||
| psiA, psiB; pA, pB, rB | 5 | 1,128.81 | 8.08 | 0.0171 | ||
| psiA, psiBA, psiBa; pA, pB, rB | 6 | 1,130.8 | 10.07 | 0.0063 | ||
| psiA, psiB (ELE + DTR); pA, pB | 6 | 1,132.21 | 11.48 | 0.0031 | ||
| psiA, psiBA (ELE + DTR), psiBa (ELE + DTR); pA, pB | 9 | 1,137.12 | 16.39 | 0.0003 | ||
| psiA, psiB; pA, pB | 4 | 1,141.3 | 20.57 | 0 | ||
| psiA, psiBA, psiBa; pA, pB | 5 | 1,142.01 | 21.28 | 0 | ||
| psiA, psiBA (ELE + DTR), psiBa (ELE + DTR); pA, pB, rB | 10 | 1,151.45 | 30.72 | 0 | ||
| 赤狐 + 黄喉雉鹑 Vulpes vulpes + Tetraophasis szechenyii | ||||||
| psiA, psiBA (DTW), psiBa (DTW); pA, pB, rB | 8 | 814.35 | 0 | 0.9847 | ||
| psiA, psiBA, psiBa; pA, pB, rB | 6 | 823.13 | 8.78 | 0.0122 | ||
| psiA, psiB (DTW); pA, pB, rB | 6 | 826.47 | 12.12 | 0.0023 | ||
| psiA, psiB; pA, pB, rB | 5 | 829.01 | 14.66 | 0.0006 | ||
| psiA, psiBA (DTW), psiBa (DTW); pA, pB | 7 | 833.91 | 19.56 | 0.0001 | ||
| psiA, psiB (DTW); pA, pB | 5 | 833.97 | 19.62 | 0.000 | ||
| psiA, psiB; pA, pB | 4 | 836.53 | 22.18 | 0 | ||
| psiA, psiBA, psiBa; pA, pB | 5 | 837.41 | 23.06 | 0 | ||
| 赤狐 + 血雉 Vulpes vulpes + Ithaginis cruentus | ||||||
| psiA, psiBA (DTW), psiBa (DTW); pA, pB, rB | 8 | 935.01 | 0 | 0.773 | ||
| psiA, psiB (DTW); pA, pB, rB | 6 | 937.74 | 2.73 | 0.1974 | ||
| psiA, psiBA, psiBa; pA, pB, rBa | 6 | 942.00 | 6.99 | 0.0235 | ||
| psiA, psiB; pA, pB, rBa | 5 | 944.68 | 9.67 | 0.0061 | ||
| psiA, psiB (DTW); pA, pB | 5 | 968.95 | 33.94 | 0 | ||
| psiA, psiBA (DTW), psiBa (DTW); pA, pB | 7 | 972.72 | 97.71 | 0 | ||
| psiA, psiB; pA, pB | 4 | 975.74 | 40.73 | 0 | ||
| psiA, psiBA, psiBa; pA, pB | 5 | 977.74 | 42.73 | 0 | ||
参数含义见
The meaning of parameters is shown in
表3 赤狐与3种雉类条件型双物种占域模型最优模型的参数估计值(平均值 ± 标准差)
Table 3
| 物种对 Species pairs | psiA ± SE | psiBA ± SE | psiBa ± SE | pA ± SE | pB ± SE | rB ± SE | SIF ± SE |
|---|---|---|---|---|---|---|---|
| 赤狐 + 白马鸡 Vulpes vulpes + Crossoptilon crossoptilon | 0.50 ± 0.10 | 0.68 ± 0.20 | 0.68 ± 0.20 | 0.10 ± 0.02 | 0.32 ± 0.03 | 0.14 ± 0.02 | 1.00 |
| 赤狐 + 黄喉雉鹑 Vulpes vulpes + Tetraophasis szechenyii | 0.56 ± 0.07 | 0.70 ± 0.08 | 0.31 ± 0.38 | 0.09 ± 0.01 | 0.42 ± 0.07 | 0.09 ± 0.02 | 1.42 ± 0.41 |
| 赤狐 + 血雉 Vulpes vulpes + Ithaginis cruentus | 0.62 ± 0.07 | 0.81 ± 0.30 | 0.29 ± 0.17 | 0.08 ± 0.01 | 0.59 ± 0.05 | 0.11 ± 0.01 | 1.31 ± 0.14 |
参数含义见
The meaning of parameters is shown in
图2
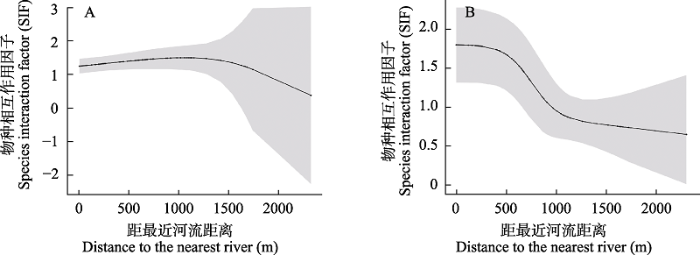
图2
基于最优模型预测的协变量对赤狐与血雉(A)和黄喉雉鹑(B)在位点水平上的物种相互作用因子(SIF)的影响。灰色部分为每个位点SIF值的95%置信区间。
Fig. 2
The effect of covariates predicted by the optimal model on the species interaction factor (SIF) of the red fox and blood pheasant (A), red fox and buff-throated partridge (B) at the single site level. Gray area indicates 95% confidence intervals for SIF value of each site.
赤狐的存在并不影响白马鸡对于位点的使用(psiBA = psiBa = 0.68 ± 0.20), 白马鸡与赤狐在研究区域内空间分布相互独立(SIF = 1)。白马鸡对位点的使用概率主要受环境因子影响, 随海拔升高而降低, 随距最近道路距离升高而升高(图3)。
图3
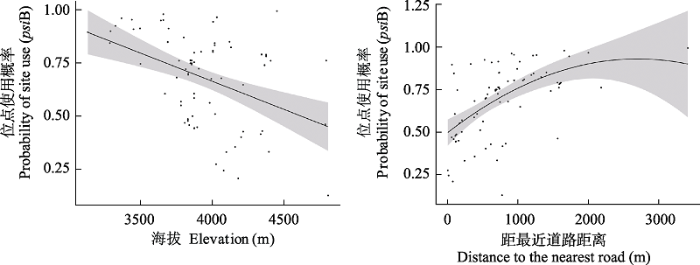
图3
基于最优模型预测的协变量在位点水平上对白马鸡位点使用概率的影响。黑色曲线为多项式回归拟合曲线, 灰色区域为95%置信区间。
Fig. 3
The effect of covariates predicted by the optimal model on the white eared-pheasant’s site use at the single site level. The black curve fitted with polynomial regression, gray area indicates 95% confidence intervals.
2.2 赤狐对3种雉类探测率的影响
单物种占域模型最优模型显示4个物种的探测率均不受探测协变量的影响(附录2), 所以在建立条件型双物种占域模型时不纳入探测协变量。条件型双物种占域模型结果显示当赤狐存在时3种雉类的探测概率低于赤狐不存在时(pB > rB) (表3)。即当赤狐存在时, 3种雉类可能会采取更加隐蔽的活动方式或减少觅食强度躲避来自赤狐的捕食风险。
3 讨论
单物种占域模型结果显示, 距最近河流距离是影响黄喉雉鹑和血雉对位点使用概率的重要因子。有研究表明黄喉雉鹑和血雉都偏好距水源距离近的生境(贾陈喜等, 1999; Wang et al, 2021)。已有对白马鸡的生境选择研究结果显示, 白马鸡主要偏好于距离水源近且灌木盖度高的生境(Jia et al, 2005; 贾非等, 2005), 而本研究结果显示白马鸡偏好于选择远离道路且海拔相对较低的生境, 对其他因子没有明显的偏好。差异来源可能是本研究的范围尺度比已有研究大, 因子更宏观。在这3种雉类中, 白马鸡的活动海拔相对偏低(Wang et al, 2021), 加之体型较大, 常集群, 活动范围较大, 对人类活动相对敏感(格玛嘉措等, 1999), 故其可能要相对远离道路。同时, 由于环境的差异性, 同一物种在不同区域本身就会表现出不同的环境因子偏好(杨春花等, 2006)。本研究结果还显示环境因子不影响赤狐对位点的使用, 这可能是由于赤狐可栖息的生境类型十分丰富, 其空间分布主要取决于食物的丰富度(马勇等, 2014), 受环境因子的影响相对较小。
根据条件型双物种占域模型的结果, 赤狐与3种同域分布的雉类物种对的分布关系都受到了环境变量的影响, 但赤狐只对黄喉雉鹑和血雉的位点使用概率产生影响(SIF ≠ 1)。赤狐与黄喉雉鹑和血雉的空间分布趋于重合(SIF > 1), 且当活动区域距河流距离越近, 赤狐与黄喉雉鹑和血雉的空间分布重合度越高, 也意味着发生捕食行为的概率更高。有研究表明捕食者偏好在食物丰富的环境中活动且大多数捕食者具有追踪猎物的能力, 这会导致捕食者和猎物密度呈正相关关系(Godin & Keenleyside, 1984; Luttbeg & Sih, 2004; Ngoprasert et al, 2012)。黄喉雉鹑和血雉都是赤狐捕食的猎物, 且赤狐也偏好距河流较近的生境(Cavallini & Lovari 1991), 所以其共同出现的概率高。赤狐不影响白马鸡对位点的使用, 二者的空间分布相互独立(SIF = 1)。白马鸡对位点的使用概率随海拔升高而降低, 随位点距道路距离的升高而升高。有研究表明在同一区域内, 捕食者与不同猎物的空间分布关系会受其对猎物偏好的影响, 从而与不同猎物表现出不同的空间分布关系(Steinmetz et al, 2013)。相比于黄喉雉鹑和血雉, 白马鸡体型更大, 可能不是赤狐最佳的捕食选择, 从而在相机位点表现出与黄喉雉鹑和血雉有更高的概率共同出现, 与白马鸡在空间分布上相互独立。
在占域模型中, 一个物种可能改变另一物种的探测概率, 进而影响它们在整个景观中的占域概率(Bailey et al, 2009)。捕食者会在一定程度上影响猎物的探测概率(Wang et al, 2015; Farris et al, 2016; Murphy et al, 2019)。本研究结果也显示当赤狐分别与3种雉类使用同一位点时, 3种雉类的探测概率因赤狐的存在而下降, 且3个物种对的探测概率仅受物种作用的影响, 不受环境因子影响。猎物可能会避开捕食者最可能出现的地点而选择更加隐蔽复杂的栖息地或是减少自身活动时间来减少与捕食者相遇的几率(Werner, 1992; Creel & Winnie, 2005)。尽管赤狐与黄喉雉鹑和血雉在空间分布上趋于重合, 与白马鸡在空间分布上相互独立, 但3种雉类都可能会在赤狐存在的区域中避开赤狐活动最活跃的地点或是调整自身活动时间和微生境来避开赤狐的捕食, 从而降低了3种雉类的探测率。
本研究结果显示捕食作用与环境因子促使赤狐与黄喉雉鹑和血雉在相机位点有更高的概率共同出现, 而赤狐与白马鸡的空间关系只受环境因子影响。同时捕食作用也降低了3种雉类的探测概率。
附录 Supplementary Material
附录1 赤狐与3种雉类的8种条件型双物种占域模型的候选模型
Appendix 1 Eight candidate conditional two-species occupancy model for red fox and three pheasant species
附录2 4个物种单物种占域模型选择结果
Appendix 2 Selection results of single species occupancy model for four target species
参考文献
Modeling co-occurrence of northern spotted and barred owls: Accounting for detection probability differences
DOI:10.1016/j.biocon.2009.07.028 URL [本文引用: 1]
Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes
DOI:10.1111/1365-2664.12432 URL [本文引用: 1]
The role of introduced species in shaping the distribution and abundance of island reptiles
DOI:10.1007/BF02214232 URL [本文引用: 1]
Environmental-factors influencing the use of habitat in the red fox, Vulpes vulpes
DOI:10.1111/jzo.1991.223.issue-2 URL [本文引用: 1]
Combined effects of habitat and interspecific interaction define co-occurrence patterns of sympatric Galliformes
Species richness and co-occurrence patterns of Galliformes in China at three large spatial scales: Does scale size matter? Revue D Ecologie-la Terre Et La Vie
On the prevalence and relative importance of interspecific competition: Evidence from field experiments
DOI:10.1086/284165 URL [本文引用: 1]
Responses of elk herd size to fine-scale spatial and temporal variation in the risk of predation by wolves
DOI:10.1016/j.anbehav.2004.07.022 URL [本文引用: 2]
Disentangling biotic interactions, environmental filters, and dispersal limitation as drivers of species co-occurrence
DOI:10.1111/ecog.2018.v41.i8 URL [本文引用: 1]
Estimation of natural ecological value for the plateau pasturing area—A case study in Xinlong County of Sichuan Province
高原牧区自然生态价值的测算——以四川省甘孜藏族自治州新龙县为例
Patterns of spatial co-occurrence among native and exotic carnivores in north-eastern Madagascar
DOI:10.1111/acv.12233 URL [本文引用: 2]
A preliminary observation on the ecological habits of white eared-pheasant
白马鸡生态习性的初步观察
Foraging on patchily distributed prey by a cichlid fish (Teleostei, Cichlidae): A test of the ideal free distribution theory
DOI:10.1016/S0003-3472(84)80330-9 URL [本文引用: 1]
PRESENCE—Software to estimate patch occupancy and related parameters
Social organization of blood pheasant (Ithaginis cruentus) in Wolong Nature Reserve
卧龙自然保护区血雉的社群组织
Winter habitat requirements of white eared-pheasant Crossoptilon crossoptilon and blood pheasant Ithaginis cruentus in south-west China
DOI:10.1017/S095927090500047X URL [本文引用: 1]
Habitat selection and spatial distribution of white eared-pheasant Crossoptilon crossoptilon during early breeding period
白马鸡繁殖早期栖息地选择和空间分布
Spatial and temporal structure of a mesocarnivore guild in midwestern North America
DOI:10.1002/wmon.v191.1 URL [本文引用: 1]
Research progress of habitat selection by red fox (Vulpes vulpes)
赤狐生境选择研究进展
Predator and prey habitat selection games: The effects of how prey balance foraging and predation risk
DOI:10.1560/L6QV-UA5T-RDR7-L7QG URL [本文引用: 1]
Food habits by red fox (Vulpes vulpes), a review
赤狐食性的研究进展
Investigating species co-occurrence patterns when species are detected imperfectly
DOI:10.1111/jae.2004.73.issue-3 URL [本文引用: 1]
Urban predator-prey association: Coyote and deer distributions in the Chicago metropolitan area
DOI:10.1007/s11252-014-0389-5 URL [本文引用: 1]
Using camera traps to investigate spatial co-occurrence between exotic predators and native prey species: A case study from northeastern Madagascar
DOI:10.1111/jzo.12645
[本文引用: 4]

The presence of exotic predators in ecosystems across the world is a leading driver of native species' declines. Exotic predators largely influence native species through predation and harassment, which may cause native species to avoid them spatially. We used a camera trap dataset from seven sites in Madagascar's largest protected area complex to investigate spatial co-occurrence patterns between three exotic predators (free-ranging domestic dogs Canis familiaris and cats Felis silvestris, and small Indian civet Viverricula indica) and native ground-dwelling forest birds (n = 5) and small mammals (n = 4). We created 216 two-species occupancy models for 27 exotic predator-native species pairings to examine how habitat, exotic predator presence or both, influenced each native species' occupancy and/or detection. We found that native bird and small mammal occupancy and/or detection were strongly related to an exotic predator's presence 52% of the time (i.e. 14 of 27 exotic predator-native species pairings). Six of the 14 species pairings (i.e. 43%) had non-independent co-occurrence patterns, four of which were habitat-mediated. The effect of exotic predator presence on native species' detection was largely negative, depressing native species' detection in seven out of 12 instances (i.e. 58% of the time). The small Indian civet and free-ranging cats each strongly influenced the occupancy and/or detection of six species for a combined impact on seven native species, while domestic dogs strongly influenced two species. By including habitat covariates in two-species occupancy models, we gained deeper insight into the effect exotic predator presence has on native species' distribution. We also note that the strong effect of exotic predator presence on our ability to detect native species can hinder the ability of researchers to provide accurate parameter estimates. We recommend future research into the synergistic effects that habitat degradation and exotic species presence has on native species.
Occurrence of three felids across a network of protected areas in Thailand: Prey, intraguild, and habitat associations
DOI:10.1111/btp.2012.44.issue-6 URL [本文引用: 1]
Prey abundance drives habitat occupancy by jaguars in Amazonian floodplain river islands
DOI:10.1016/j.actao.2019.04.004 URL [本文引用: 1]
Two-species occupancy models: A new parameterization applied to co-occurrence of secretive rails
DOI:10.1890/09-0470.1 URL [本文引用: 3]
Separating the influences of environment and species interactions on patterns of distribution and abundance: Competition between large herbivores
DOI:10.1111/j.1365-2656.2008.01520.x
PMID:19175695
[本文引用: 1]
1. Much recent research has focused on the use of species distribution models to explore the influence(s) of environment (predominantly climate) on species' distributions. A weakness of this approach is that it typically does not consider effects of biotic interactions, including competition, on species' distributions. 2. Here we identify and quantify the contribution of environmental factors relative to biotic factors (interspecific competition) to the distribution and abundance of three large, wide-ranging herbivores, the antilopine wallaroo (Macropus antilopinus), common wallaroo (Macropus robustus) and eastern grey kangaroo (Macropus giganteus), across an extensive zone of sympatry in tropical northern Australia. 3. To assess the importance of competition relative to habitat features, we constructed models of abundance for each species incorporating habitat only and habitat + the abundance of the other species, and compared their respective likelihoods using Akaike's information criterion. We further assessed the importance of variables predicting abundance across models for each species. 4. The best-supported models of antilopine wallaroo and eastern grey kangaroo abundance included both habitat and the abundance of the other species, providing evidence of interspecific competition. Contrastingly, models of common wallaroo abundance were largely influenced by climate and not the abundance of other species. The abundance of antilopine wallaroos was most influenced by water availability, eastern grey kangaroo abundance and the frequency of late season fires. The abundance of eastern grey kangaroos was most influenced by aspects of climate, antilopine wallaroo abundance and a measure of cattle abundance. 5. Our study demonstrates that where census and habitat data are available, it is possible to reveal species' interactions (and measure their relative strength and direction) between large, mobile and/or widely-distributed species for which competition is difficult to demonstrate experimentally. This allows discrimination of the influences of environmental factors and species interactions on species' distributions, and should therefore improve the predictive power of species distribution models.
Tigers, leopards, and dholes in a half-empty forest: Assessing species interactions in a guild of threatened carnivores
DOI:10.1016/j.biocon.2012.12.016 URL [本文引用: 2]
Niche partitioning among three montane ground-dwelling pheasant species along multiple ecological dimensions
DOI:10.1111/ibi.v163.1 URL [本文引用: 3]
Mesopredator spatial and temporal responses to large predators and human development in the Santa Cruz Mountains of California
DOI:10.1016/j.biocon.2015.05.007 URL [本文引用: 2]
Individual behavior and higher-order species interactions
DOI:10.1086/285395 URL [本文引用: 1]
Review of habitat selection in the giant panda (Ailuropoda melanoleuca)
大熊猫(Ailuropoda melanoleuca)生境选择研究进展
Summer habitat selection by white-lipped deer (Cervus albirostris) in Chaqingsongduo White-lipped Deer National Nature Reserve
察青松多白唇鹿国家级自然保护区白唇鹿对夏季生境的选择
Study on habitat use and activity rhythms of wild boar in eastern region of Northeast Tiger and Leopard National Park
东北虎豹国家公园东部的野猪生境利用和活动节律初步研究
Seasonal variation of daily activity patterns and diet of yellow-throated marten (Martes flavigula)
黄喉貂日活动节律及食性的季节变化

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}