



作为当今世界环境和生态学的研究焦点之一, 生物入侵严重威胁被入侵地的生态系统功能、资源可利用性和经济可持续发展(Minteer et al, 2020)。大量研究发现, 除了入侵植物自身的生物学特性(Blossey & Notzold, 1995)之外, 本土植物群落的可入侵性(invasibility, 即本土群落抵抗外来物种入侵的能力)也是影响外来植物入侵成功与否的关键因素(Alpert et al, 2000)。学界先后提出群落多样性阻抗假说、资源竞争假说等尝试解释本土植物群落的可入侵性(Levine & D’Antonio, 1999; Shea & Chesson, 2002)。
在关于群落可入侵性的经典理论中, Elton (1958)在The Ecology of Invasions by Animals and Plants一书中最早提出生物抵抗假说(biotic resistance hypothesis), 认为多样性丰富的群落不易被外来物种入侵, 该观点得到国内外大量研究的支持(Pimm, 1984; Zhang et al, 2020)。另外, 达尔文归化假说(Darwin’s naturalization hypothesis, Darwin, 1859)强调近缘种往往具有相近的环境需求和强烈的种间竞争, 土著种会抑制其近缘外来种的归化和入侵, 因而本土植物群落对外来物种的抵抗力可能受群落组分种(即群落内物种组成)与入侵种的谱系距离(群落系统发育支长)影响。根据以上理论可以推测, 具有较高的物种多样性和与入侵种较近谱系距离的本土群落对入侵有更强的抵抗力。但这一论点仍缺乏针对不同生态系统和入侵物种的野外数据的广泛验证。
紫茎泽兰(Ageratina adenophora)为菊科紫茎泽兰属的多年生半灌木, 原产墨西哥、哥斯达黎加一带(O’Sullivan, 1979), 作为外来入侵植物最早于1944年在澳大利亚昆士兰岛被报道(Auld, 1970)。20世纪50、60年代曾作为观赏植物引种到欧洲、亚洲等, 因较强的抗逆性和广泛的生态适应性, 已经广布于世界多个热带、亚热带地区(Poudel et al, 2019)。20世纪40年代前后, 紫茎泽兰由中缅边境传入我国云南, 其后在广西、四川等地迅速扩散, 位居我国2003年公布的第一批外来入侵物种名单之首(强胜, 1998)。在其入侵区, 紫茎泽兰多以单优种群出现, 连片密集分布, 严重破坏当地生态环境(Poudel et al, 2019)。控制紫茎泽兰的入侵是我国西南地区生态环境保护的重要任务之一。认识不同群落结构对紫茎泽兰入侵的抵抗力, 探索基于群落学的入侵植物防控策略, 对区域生态环境保护具有重要意义。
目前国内外学者已对紫茎泽兰开展了种群分布与扩散特征(Horvitz et al, 2014; Zhu et al, 2007)、分子生物学特征(桂富荣, 2006)及人工防除技术(Wan et al, 2010)等研究, 但关于本土植物群落可入侵性的研究资料仍较为匮乏。云南松(Pinus yunnanensis)林为我国西南地区主要森林类型之一, 该区域是我国天然林资源保护和长江防护林工程的重点地区, 对区域水源涵养和生态环境维持具有重要意义。然而近年来云南松林下遭遇紫茎泽兰的入侵, 给当地自然景观、生态系统功能和社会经济带来严重危害。本研究以攀西地区云南松群落为研究对象, 从群落生态学入手研究紫茎泽兰入侵强度与环境因子、群落物种多样性和谱系多样性的关系, 探索紫茎泽兰在攀西地区的生态入侵机制。主要回答以下问题: (1)物种多样性高的群落中紫茎泽兰的入侵强度是否更低? (2)组分种与紫茎泽兰亲缘关系更近的群落中紫茎泽兰入侵强度是否更低? 以期为我国西南地区紫茎泽兰的有效防控、森林资源的科学管理与生物多样性保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区为横断山脉东缘的攀西地区(攀枝花市和凉山彝族自治州的简称, 100°15′-103°35′ E, 26°03′- 29°27′ N), 位于青藏高原、四川盆地和云贵高原的过渡带, 地形复杂, 地势西北高、东南低。该区为典型的亚热带季风气候区, 年平均气温约14-20℃, 年平均降水量700-1,400 mm (张利平等, 2020)。
20世纪50年代以来, 因大规模开垦、砍伐等人类活动, 攀西地区亚热带常绿阔叶林遭到严重破坏, 取而代之的是90年代人工种植或者飞播造林形成的云南松林, 其群落结构较为单一(汪建云等, 2015)。20世纪80年代初, 紫茎泽兰从云南省自然传播到攀枝花盐边县, 随后在凉山州盐源、德昌县等地快速扩散(侯太平, 2000), 现已在攀西地区广泛入侵林地、草地和农田等, 成为危害农林业生产最严重的入侵物种之一(周俗和谢永良, 1999)。紫茎泽兰多分布在海拔1,000-2,000 m、年均温12.5-19.3℃、水源丰富的河谷、林缘等环境中(周俗和谢永良, 1999), 适应能力极强, 在林隙、草地、沟边和荒坡等都能生长。一旦入侵后它会迅速排挤本土植物, 易形成单优群落, 给造林工程和生态系统功能造成极大威胁(刘伦辉等, 1985)。
1.2 样地设置
以攀西地区为主要研究区域, 向北延伸至雅安市石棉县, 南北跨度约320 km, 海拔1,015-2,278 m。根据遥感影像布设样方点(图1), 以充分代表该区域地形环境特征和云南松林分状况。2019年12月, 根据布设的样方点, 利用GPS结合实地踏查情况共调查40个样地, 每个样地面积10 m × 10 m, 按对角线取样法设置2个5 m × 5 m的灌木样方, 并在每个灌木样方内随机选取1个1 m × 1 m草本样方。
图1
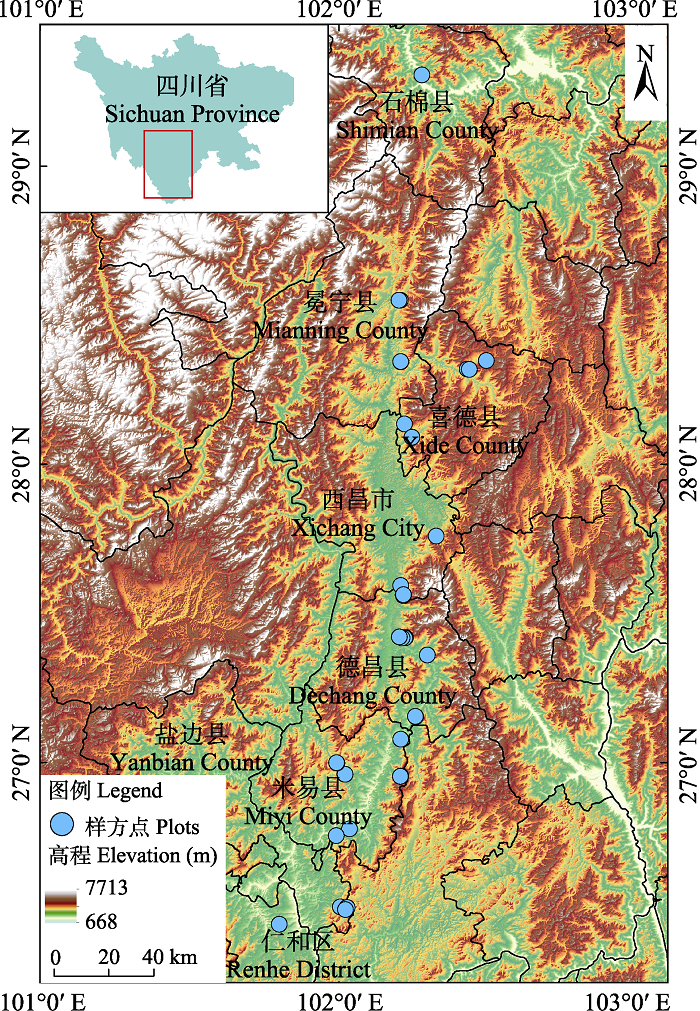
图1
四川省攀西地区样方位置分布图
Fig. 1
Distribution map of the sample plots in the Panxi region, Sichuan Province
1.3 群落调查
对乔木层进行每木检尺, 记录物种、胸径、株数和郁闭度等; 对灌木层和草本层植物鉴定到种, 记录每个物种的多度、高度、频度和盖度; 记录每个样方的地理坐标和海拔; 用罗盘仪测量坡向、坡度; 用土壤水分测定仪(Takeme-10, 哲勤科技有限公司, 大连)随机选取样方内5个点测量土壤5 cm处的温度和湿度(土壤温、湿度于2020年11月复查1次, 取两次调查平均值)。
因调查区域多处样地有不同程度火烧痕迹, 故参照刘经伦等(2019)并结合实际将火烧强度按火烧高度和面积分为4级: (1)未被火烧, 无痕迹; (2)轻度火烧, 火烧高度 < 1 m, 面积 < 0.3 m2; (3)中度火烧, 火烧高度为1-2 m, 面积为0.3-0.6 m2; (4)重度火烧, 火烧高度 > 2 m, 面积 > 0.6 m2。
参考张修玉等(2010)的方法, 野外调查群落以紫茎泽兰盖度代表群落入侵强度, 并划分为未入侵(完全以本土植物为优势)、轻度入侵(紫茎泽兰零星分布, 其盖度 < 20%)、中度入侵(紫茎泽兰与本土植物共存, 其盖度20%-60%)、重度入侵(紫茎泽兰为优势种群, 其盖度 > 60%) 4个等级。
1.4 重要值
采用重要值(importance value, IV)表示某一物种在群落中的重要性(方精云等, 2009):
由于研究区乔木层是以云南松为主的单优群落, 结构简单, 云南松重要值多接近于1, 因此未计算乔木层的物种多样性和谱系多样性, 仅将冠层郁闭度作为环境因子之一纳入统计分析。
1.5 物种多样性
式中, S为样方的物种数, Pi为第i个物种的重要值。
1.6 谱系多样性
1.6.1 群落谱系树的构建
参照《中国植物志》在线网站 (
1.6.2 谱系结构指数
为了检验群落谱系结构对外来植物入侵是否具有抵抗力, 本文采用Metz等(2010)的方法, 使用净种间亲缘关系指数(net relatedness index, NRI)代表物种间亲缘关系。假定样地中调查的所有物种组成局域物种库, 先计算出小样方内目标物种与其他物种的平均观测谱系距离, 保持物种数量与物种个体数不变, 将小样方内物种从物种库中随机抽取9,999次, 从而获得该小样方中物种在随机零模型下的期望谱系距离, 然后利用随机分布结果计算出小样方内期望谱系距离的平均值和标准差。NRI计算公式如下:
需要注意的是, 本研究除了采用Webb等(2002)的群落净种间亲缘关系指数(代表群落中任意两个物种对的平均谱系距离), 还参考Metz等(2010)采用目标物种净种间亲缘关系指数(NRIob), 该指数代表的是小样方内目标物种与群落中与其亲缘关系最近的一个物种的谱系距离, 而不是任意物种对最小谱系距离的平均值。它的值越大, 表示与目标物种间的亲缘关系越近, 基于植物性状保守性的假设, 亲缘关系越近的种对相似环境的适应能力越接近, 生态位也越相近(Cadotte et al, 2018)。本研究以紫茎泽兰作为目标物种计算群落的目标物种净种间亲缘关系指数。
净种间亲缘关系指数的计算利用R 4.0.2软件picante包(Kembel et al, 2010)完成。
1.7 统计分析
本研究分析的环境因子有10个, 包括海拔、纬度、坡向、坡度、火烧强度、土壤湿度、土壤温度、冠层郁闭度、灌木层和草本层(非紫茎泽兰)盖度; 生物因子包括灌木层和草本层的Shannon-Wiener指数、群落净种间亲缘关系指数和目标物种净种间亲缘关系指数。
为减少环境因子数量对模型解释度的稀释, 首先采用主成分分析(principal component analysis, PCA)方法提取表征样地特征的主要环境因子。以紫茎泽兰入侵盖度为因变量, 主成分分析所得主要环境因子轴和群落生物因子作为自变量, 进行广义线性模型(generalized linear models, GLMs, Dobson, 1990)分析。由于盖度数据为百分比, 无法满足正态分布假设, 因而参考Warton和Hui (2011)的方法, 在广义线性模型分析时采用逻辑斯蒂回归方法进行计算。
利用R 4.0.2软件lme4 (Bates et al, 2015)、vegan (Oksanen et al, 2019)、car (Fox & Weisberg, 2019)和ggplot2包(Wickham, 2016)等进行模型分析和图形绘制。
2 结果
2.1 群落基本特征
40个样地共调査到植物114种, 隶属于43科97属。其中, 乔木13科18属21种, 以云南松、栓皮栎(Quercus variabilis)、无患子(Sapindus saponaria)、槲栎(Quercus aliena)、珍珠花(Lyonia ovalifolia)占优势; 灌木49种, 隶属27科41属, 主要有矮高山栎(Quercus monimotricha)、斑鸠菊(Vernonia esculenta)、扁刺蔷薇(Rosa sweginzowii)、扁核木(Prinsepia utilis)、插田泡(Rubus coreanus)等; 草本44种, 隶属19科42属, 主要有紫茎泽兰、黄茅(Heteropogon contortus)、云南兔儿风(Ainsliaea yunnanensis)、白莲蒿(Artemisia stechmanniana)、野青茅(Deyeuxia pyramidalis)等。
2.2 环境因子与紫茎泽兰入侵强度
对10个代表云南松群落的环境变量进行主成分分析, 最终得到2个主成分(表1)。其中, 第一主成分反映云南松群落土壤温度和土壤湿度因子, 解释了57.85%的环境因子变异; 第二主成分反映群落林分郁闭度和草本层盖度因子, 解释了23.46%的环境因子变异。将第一主成分和第二主成分轴(共包含10个环境因子81.31%的变异)代入上述回归模型中进行分析。
表1 云南松群落环境变量在2个主成分上的载荷值
Table 1
| 环境变量 Environmental variables | PC1 | PC2 |
|---|---|---|
| 海拔 Altitude | 0.29 | -0.43 |
| 纬度 Latitude | -0.42 | -0.13 |
| 坡向 Aspect | 0.25 | -0.21 |
| 坡度 Slope | -0.26 | 0.34 |
| 土壤温度 Soil temperature | 0.51 | - |
| 土壤湿度 Soil moisture | 0.53 | - |
| 火烧强度 Fire intensity | 0.17 | 0.36 |
| 冠层郁闭度 Canopy closure | - | -0.46 |
| 灌木层盖度 Shrub coverage | - | -0.25 |
| 草本层盖度 Herb coverage | 0.17 | 0.47 |
| 变异解释量 Proportion of variance (%) | 57.85 | 23.46 |
广义线性回归结果表明: 第一主成分和第二主成分轴表征的海拔、坡向和火烧强度等环境因子与紫茎泽兰入侵强度无显著作用关系(表2, P > 0.05), 表明在研究范围(26.46°-29.31° N、海拔1,015-2,278 m)内, 海拔、坡向和火烧强度等环境因子对紫茎泽兰入侵强度没有显著影响。
表2 攀西地区云南松林环境因子和生物因子对紫茎泽兰入侵强度的影响
Table 2
| 因子 Factor | 数值范围 Numerical ranges | χ2 | P |
|---|---|---|---|
| 第一主成分 First principal component | -3.35 to 3.16 | 0.23 | 0.63 |
| 第二主成分 Second principal component | -3.02 to 3.43 | 0.72 | 0.39 |
| 灌木层Shannon-Wiener指数 Shannon-Wiener index of shrub layer | 0.14-2.26 | 4.06 | 0.04* |
| 灌木层净种间亲缘关系指数 Net relatedness index of shrub layer | -1.27 to 3.11 | 0.46 | 0.49 |
| 灌木层目标物种净种间亲缘关系指数 Net relatedness index of shrub layer to the target species | -1.29 to 2.39 | 0.05 | 0.81 |
| 草本层Shannon-Wiener指数 Shannon-Wiener index of herb layer | 0.93-2.37 | 1.99 | 0.15 |
| 草本层群落净种间亲缘关系指数 Net relatedness index of herb layer | -1.32 to 1.31 | 3.21 | 0.07 |
| 草本层目标物种净种间亲缘关系指数 Net relatedness index of herb layer to the target species | -1.74 to 1.44 | 4.28 | 0.03* |
* P < 0.05.
2.3 物种多样性指数与紫茎泽兰入侵强度
广义线性模型分析结果显示, 草本层Shannon-Wiener指数与紫茎泽兰入侵强度没有显著关系(表2, χ2 = 1.99, P > 0.05), 但灌木层Shannon-Wiener指数与紫茎泽兰入侵强度呈显著负相关关系, 即灌木层Shannon-Wiener指数越高, 紫茎泽兰入侵强度越低(表2, χ2 = 4.06, P < 0.05), 紫茎泽兰入侵强度随灌木层Shannon-Wiener指数的升高而降低(图2a)。调查结果显示: S2、S7等样地灌木层Shannon-Wiener指数较高(分别是2.26和2.19), 物种丰富度 > 15, 群落内主要灌木物种有矮高山栎、野拔子(Elsholtzia rugulosa)、金丝梅(Hypericum patulum)、密花合耳菊(Synotis cappa)、珍珠花等, 尽管群落周边路缘有紫茎泽兰分布, 但群落内没有发现紫茎泽兰; S32、S33等样地Shannon-Wiener指数较低(分别是0.40和0.14), 灌木层仅有2个种: 野拔子和圆锥山蚂蝗(Desmodium elegans), 群落内紫茎泽兰盖度均达90%以上。
图2
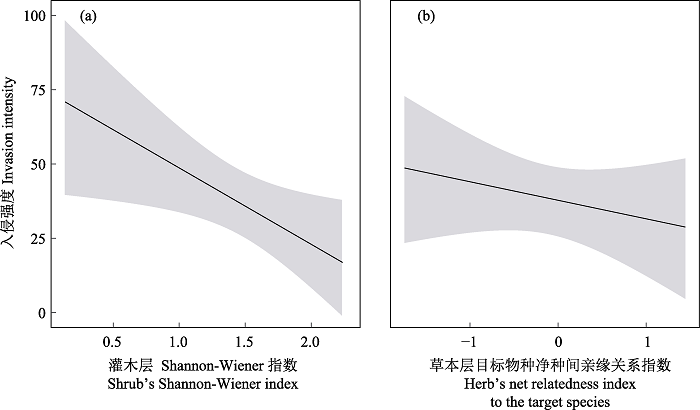
图2
灌木层Shannon-Wiener指数(a)、草本层目标物种净种间亲缘关系指数(b)对紫茎泽兰入侵影响的预测曲线
Fig. 2
Prediction curves of the effects of Shannon-Wiener index of shrub layer (a) and the net relatedness index of herb layer to the target species (b) on the invasion of Ageratina adenophora
2.4 NRI指数与紫茎泽兰入侵强度
模型显示, 灌木层的群落净种间亲缘关系指数、目标物种净种间亲缘关系指数和草本层的群落净种间亲缘关系指数与紫茎泽兰的入侵强度没有显著关系(表2, P > 0.05), 但草本层目标物种净种间亲缘关系指数与入侵强度呈显著负相关关系(表2, χ2 = 4.28, P < 0.05), 即同层物种与紫茎泽兰亲缘关系(重要值为权重)越近, 紫茎泽兰入侵强度越低(图2b)。该结果与实际调查基本吻合: 未入侵群落中主要物种有千里光(Senecio scandens)、密花合耳菊、云南兔儿风、蛛毛香青(Anaphalis busua)、鬼针草(Bidens pilosa)、白莲蒿等, 同为菊科植物; 而重度入侵群落中与紫茎泽兰亲缘关系相对较近的物种较少。
3 讨论
大量研究表明, 物种多样性对外来植物入侵有一定的指示作用(吴昊等, 2020; 张文馨等, 2020), 而群落谱系多样性从亲缘关系角度解释了本土与外来物种之间的关系(Elton, 1958; Webb et al, 2002), 可能对本土群落的可入侵性具有重要影响。本研究基于大量野外调查, 分析了攀西地区云南松群落环境因子、物种多样性和谱系多样性对紫茎泽兰入侵的影响。
本研究结果表明, 灌木层物种多样性越高, 紫茎泽兰入侵强度越低(图2a, 表2, P < 0.05), 表明较高的灌木层物种多样性有助于抑制外来物种的入侵。Symstad (2000)提出多样性高的群落内各个物种能够充分利用现有的资源, 留给外来物种的空余生态位大大减少, 降低了入侵发生的概率。这符合空余生态位假说(empty niche hypothesis), 即因种间竞争激烈, 群落内可利用资源较少, 导致外来物种难以进入(Hierro et al, 2005)。张震等(2010)通过对入侵植物飞机草(Chromolaena odorata)本土(墨西哥)和入侵地(中国)的群落调查发现, 随着群落中本土物种数量的上升, 飞机草的株数呈下降趋势, 群落可入侵性降低; 吴昊等(2020)以人工构建群落的方式, 探讨了物种多样性对喜旱莲子草(Alternanthera philoxeroides)入侵的影响, 发现高水平的物种多样性能够阻止其入侵。但也有学者认为物种多样性越高, 群落结构越脆弱, 反而有利于外来物种入侵(Stachowicz et al, 1999), 对于这种差异, Crawley等(1999)提出随着研究尺度变大, 物种多样性对入侵植物的负效应被复杂的外部因素模糊, 群落某些微环境有利于外来物种入侵。本文基于区域尺度野外调查发现, 群落内空间上高于紫茎泽兰的灌木层物种多样性越丰富, 紫茎泽兰的入侵强度越低, 即复杂的灌木层对光照资源的利用越充分, 留给相对喜光的紫茎泽兰的光照资源就有限, 从而限制其入侵, 该结论支持生物抵抗假说(Elton, 1958)。
群落净种间亲缘关系指数对紫茎泽兰入侵强度无影响(表2, P > 0.05), 但草本层目标物种净种间亲缘关系指数显著影响紫茎泽兰的入侵强度(图2b, 表2, P < 0.05), 即群落垂直结构上的同层组分种与紫茎泽兰亲缘关系越近, 对外来植物入侵的抵抗力也越强。达尔文归化假说(Darwin, 1859)指出亲缘关系相近的物种对光照、水分和土壤养分等资源需求相似, 种间竞争作用更强, 在含有同属本土种的地方, 被引入的外来植物建立种群的可能性减少, 即亲缘关系相近物种竞争排斥作用更大。近年来, 以物种亲缘关系为基础的群落谱系结构逐渐被应用于植物入侵机制的研究(Li et al, 2015; El-Barougy et al, 2020), Strauss等(2006)和Miller等(2016)先后分析了草原、森林群落的谱系结构, 发现群落谱系结构越聚集, 抵抗外来物种入侵的能力越强, 证明了群落总体谱系多样性对植物入侵的影响。然而Liendo等(2021)发现群落谱系结构对外来植物入侵没有显著影响。El-Barougy等(2020)认为入侵植物与本土群落植物的相似性可能对本土群落的入侵抵抗力有更好的预测作用, 用平均谱系距离(mean phylogenetic distance, MPD)计算入侵物种与本土物种的亲缘关系远近, 解释了生态位相似性对物种入侵的抑制/促进作用。但是平均谱系距离也存在一些缺陷, 即与物种丰富度(species richness)具有较强的关联性(Swenson, 2014), 因此本研究在El-Barougy等(2020)的基础上, 采用基于平均谱系距离的目标物种净种间亲缘关系指数, 剔除了物种丰富度的影响。通过研究发现, 虽然传统的群落整体谱系多样性指数对紫茎泽兰入侵无影响, 但同层物种与紫茎泽兰亲缘关系越近, 紫茎泽兰入侵强度越弱, 表明群落中相同生活型近缘种之间的相似性限制对外来植物入侵具有抵抗力。
另外, 云南松群落灌木层、草本层群落净种间亲缘关系指数在0上下波动(表2), 未表现出显著的谱系聚集或发散规律, 且与紫茎泽兰的入侵强度并无显著关系, 说明群落整体的谱系结构所反映的群落整体谱系关系对特定物种间的竞争关系表征能力有限, 因而群落净种间亲缘关系指数与紫茎泽兰的入侵强度并无显著关系, 而本文采用的目标物种净种间亲缘关系指数可以有效弥补其缺陷。此外, 近年来已有越来越多的研究基于群落功能多样性(functional diversity)来解释外来植物的入侵机制(Pokorny et al, 2005), 并得到学界的认可。本研究中相同生活型近缘种的抑制作用可能与功能性状和资源获取方式有紧密联系, 在未来的研究中有必要借助功能多样性方法从不同角度继续深入开展紫茎泽兰的群落入侵机制研究。
影响群落中生物入侵的因子包括生物和非生物因素, 生物因素主要包括种间互利和竞争等; 非生物因素包括气温、降水、土壤及光照等(Halpern & Underwood, 2006)。Alpert等(2000)发现海拔、水分、火灾强度等环境因子对植物入侵有显著影响, 万方浩等(2002)和汪建云等(2015)对云南松林下紫茎泽兰的入侵研究发现, 紫茎泽兰株高、株丛数等随坡度、土壤水分的增加而增加。但本研究并未发现海拔、坡向和火烧强度等环境因子对研究区域内紫茎泽兰入侵强度明显的作用规律, 可能与以下原因有关: 首先, 紫茎泽兰本身有较强的表型可塑性和适应性, 加之入侵该地区已有数十年, 使紫茎泽兰具有较宽的生态幅和更好的环境耐受性, 可以占据更广阔的地理范围和多样化的生境。其次, 尺度对群落学研究结果具有重要影响, Lu和Ma (2005)对我国西南地区外来物种入侵的研究发现, 小尺度范围内(25 m2)物种多样性影响群落可入侵性, 而在稍大尺度范围(400 m2)则环境因子影响群落的可入侵性。因此, 本研究中未发现海拔等环境因子对紫茎泽兰入侵的限制作用可能与调查的环境梯度范围和空间尺度有关。
云南松林天然更新容易, 是迹地更新和荒山造林主要树种。攀西地区现有云南松是在亚热带常绿阔叶林破坏后天然或人工更新起来的次生林, 对川西地区水土保持具有重要意义。但境内云南松林近80%为纯林, 林分年轻, 群落结构单一, 生物多样性低, 生物种群间相互作用弱, 对外来物种的抵抗能力较差(蔡年辉等, 2007)。而紫茎泽兰因其极强的繁殖能力和表型可塑性(Poudel et al, 2019), 对我国西南森林群落结构和生态系统功能造成了严重威胁。本研究结果表明, 本土植物群落结构包括物种多样性和谱系多样性等影响紫茎泽兰的入侵: 灌木层物种多样性高、草本层(同层)物种与紫茎泽兰亲缘关系更近的群落具有更强的入侵抵抗力。本研究结果对我国西南地区人工林管理具有重要的启示意义: 在紫茎泽兰入侵风险区的人工林的经营管理中可调整林分树种组成以提高群落的入侵抵抗力, 可通过人工播种等手段提高林下灌木层的物种多样性和草本层本土同科近缘种的比例, 加强群落内部竞争和资源占用, 可能有助于提高本土植物群落抵御紫茎泽兰入侵的能力。
另外, 紫茎泽兰入侵强度可能与入侵时间有关(Thuiller et al, 2006)。紫茎泽兰在我国自南向北蔓延(强胜, 1998), 国家农业部2002年组织调查了包括紫茎泽兰在内的有害植物并描述了紫茎泽兰分布扩散的大致情况(卢志军, 2005), 但本研究的尺度主要涉及县、乡镇及以下单元, 难以获得相对应的入侵时间记录, 在未来的研究中可以考虑通过分枝结构的年龄分析(于兴军, 2005)、分子标记(桂富荣, 2006)等手段将入侵时间加入模型。此外, 考虑到紫茎泽兰极强的入侵能力, 在未来研究中, 还应该开展室内及野外控制实验研究, 以加强生物入侵理论的实验验证并明确其背后的生态学过程与机制。
致谢
成都理工大学张晓娟、王延茹、唐自豪、张萌、车路璐和谢成晟参加了野外调查工作, 赵管乐和卢韩好帮助绘制了样方位置分布图; 中国科学院东北地理与农业生态研究所刘艳杰老师、本文责任编委和两位外审专家等对论文提出了宝贵意见, 在此一并致谢。
参考文献
Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants
DOI:10.1078/1433-8319-00004 URL [本文引用: 2]
Eupatorium weed species in Australia
Fitting linear mixed-effects models using lme4
Evolution of increased competitive ability in invasive nonindigenous plants: A hypothesis
DOI:10.2307/2261425 URL [本文引用: 1]
Preadaptation and naturalization of nonnative species: Darwin’s two fundamental insights into species invasion
DOI:10.1146/annurev-arplant-042817-040339
PMID:29489400
[本文引用: 1]

Predicting which nonnative species become invasive is critical for their successful management, and Charles Darwin provided predictions based on species' relatedness. However, Darwin provided two opposing predictions about the relatedness of introduced nonnatives to indigenous species. First, environmental fit is the dominant factor determining invader success; thus, we should expect that invasive species are closely related to local native residents. Alternatively, if competition is important, we should expect successful invaders are distantly related to the native residents. These opposing expectations are referred to as Darwin's naturalization conundrum. The results of studies that examine nonnative species relatedness to natives are largely inconsistent. This inconsistency arises from the fact that studies occur at different spatial and temporal scales, and at different stages of invasion, and so implicitly examine different mechanisms. Further, while species have evolved ecological differences, the mode and tempo of evolution can affect species' differences, complicating the predictions from simple hypotheses. We outline unanswered questions and provide guidelines for collecting the data required to test competing hypotheses.
A comparison study on the community structure between artificial and natural forests of Pinus yunnanensis
云南松人工林与天然林群落结构的比较研究
Invasion-resistance in experimental grassland communities: Species richness or species identity?
DOI:10.1046/j.1461-0248.1999.00056.x URL [本文引用: 1]
Richness, phylogenetic diversity, and abundance all have positive effects on invader performance in an arid ecosystem
Methods and protocols for plant community inventory
DOI:10.3724/SP.J.1003.2009.09253 URL [本文引用: 1]
植物群落清查的主要内容、方法和技术规范
Genetic Diversity and Population Structure Ageratina adenophora Spreng
紫茎泽兰的遗传多样性及其种群结构分析
Approaches for testing herbivore effects on plant population dynamics
DOI:10.1111/j.1365-2664.2006.01220.x URL [本文引用: 1]
A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range
DOI:10.1111/jec.2005.93.issue-1 URL [本文引用: 1]
A simple modeling approach to elucidate the main transport processes and predict invasive spread: River-mediated invasion of Ageratina adenophora in China
DOI:10.1002/2014WR015537 URL [本文引用: 1]
Study on the Active Substances and Preparations of Poisonous Grassland Plant Eupatorium adenophorum
草地有毒植物紫茎泽兰灭蚜活性物质及制剂研究
Picante: R tools for integrating phylogenies and ecology
DOI:10.1093/bioinformatics/btq166 URL [本文引用: 1]
Elton revisited: A review of evidence linking diversity and invasibility
DOI:10.2307/3546992 URL [本文引用: 1]
The effects of phylogenetic relatedness on invasion success and impact: Deconstructing Darwin’s naturalisation conundrum
DOI:10.1111/ele.2015.18.issue-12 URL [本文引用: 1]
Effects of disturbance and alien plants on the phylogenetic structure of riverine communities
Restoration of Pinus yunnanensis forest after fire and flora analysis of substitute tree species in Daxi Mountain
大西山云南松林火灾后的恢复情况及替代树种区系分析
Studies on the distribution, harmfulness and control of Eupatorium adenophorum Spreng
紫茎泽兰在我国的分布、危害与防除途径的探讨
Plant Community Resistance to the Invasion of Croftonweed (Eupatorium adenophorum) in Southwest China
中国西南地区植物群落的可入侵性与紫茎泽兰的入侵
Scale dependent relationships between native plant diversity and the invasion of croftonweed (Eupatorium adenophorum) in southwest China
DOI:10.1614/WS-04-188R2.1 URL [本文引用: 1]
Widespread density-dependent seedling mortality promotes species coexistence in a highly diverse Amazonian rain forest
DOI:10.1890/08-2323.1 URL [本文引用: 2]
Is invasion success of Australian trees mediated by their native biogeography, phylogenetic history, or both?
Is biological control for earleaf acacia (Acacia auriculiformis) feasible in the United States?
DOI:10.1080/09583157.2020.1833305 URL [本文引用: 1]
Crofton weed (Eupatorium adenophorum) toxicity in horses
DOI:10.1111/j.1751-0813.1979.tb09538.x URL [本文引用: 1]
The complexity and stability of ecosystems
DOI:10.1038/307321a0 URL [本文引用: 1]
Plant functional group diversity as a mechanism for invasion resistance
DOI:10.1111/rec.2005.13.issue-3 URL [本文引用: 1]
Biology and management of the invasive weed Ageratina adenophora (Asteraceae): Current state of knowledge and future research needs
DOI:10.1111/wre.12351
[本文引用: 3]
Biological invasion is increasing worldwide and the management of invasive species is becoming an important priority for vegetation managers. Success of invasive species management depends on a thorough understanding of the biology of the organism in question and the effectiveness of current management efforts, in order to identify the best practices for management improvement. In this review, we synthesised current biological knowledge of a noxious invasive weed Ageratina adenophora to identify knowledge gaps and assessed management efforts to identify best practices. Finally, we proposed some priority areas for future research to fill knowledge gaps and improve management. Our analysis showed that A. adenophora has already invaded 40 countries, mainly in Asia, Oceania, Africa and Europe. Phenotypic plasticity, allelopathic interference and invasion-mediated changes in the soil microbial community are the proposed mechanisms that facilitate rapid spread of this weed. However, allelopathy as a mechanism of invasion success of this weed has not been supported by ecologically meaningful experiments. Though mechanical, chemical and biological control measures have been used, their success remains limited and the weed continues to spread in new regions. Among seven biological control agents examined to date, gall fly (Procecidochares utilis) and leaf spot fungus (Passalora ageratinae) have been effective in limited areas to suppress growth of this weed. Some perennial native grasses (e.g. Setaria sphacellata and Lolium perenne) have shown potential to competitively suppress A. adenophora. In conclusion, understanding the invasion mechanisms, exploring further to identify effective biological control agents, combined with approaches of ecological restoration, could help in the management of this weed.
The history and status of the study on crofton weed (Eupatorium adenophorum Spreng.) A worst worldwide weed
世界性恶性杂草——紫茎泽兰研究的历史及现状
Community ecology theory as a framework for biological invasions
DOI:10.1016/S0169-5347(02)02495-3 URL [本文引用: 1]
Species diversity and invasion resistance in a marine ecosystem
Theory predicts that systems that are more diverse should be more resistant to exotic species, but experimental tests are needed to verify this. In experimental communities of sessile marine invertebrates, increased species richness significantly decreased invasion success, apparently because species-rich communities more completely and efficiently used available space, the limiting resource in this system. Declining biodiversity thus facilitates invasion in this system, potentially accelerating the loss of biodiversity and the homogenization of the world's biota.
Exotic taxa less related to native species are more invasive
A test of the effects of functional group richness and composition on grassland invasibility
DOI:10.1890/0012-9658(2000)081[0099:ATOTEO]2.0.CO;2 URL [本文引用: 1]
Interactions between environment, species traits, and human uses describe patterns of plant invasions
DOI:10.1890/0012-9658(2006)87[1755:IBESTA]2.0.CO;2 URL [本文引用: 1]
Alien invasive species in China: Their damages and management strategies
中国外来入侵生物的危害与管理对策
Invasive mechanism and control strategy of Ageratina adenophora (Sprengel)
DOI:10.1007/s11427-010-4080-7 URL [本文引用: 1]
Study on AIS-Ageratina adenophora population of Pinus yunnanensis pure forest from Longyang region, Baoshan, Yunnan
保山市隆阳区云南松纯林入侵物种紫茎泽兰种群的研究
The arcsine is asinine: The analysis of proportions in ecology
The arcsine square root transformation has long been standard procedure when analyzing proportional data in ecology, with applications in data sets containing binomial and non-binomial response variables. Here, we argue that the arcsine transform should not be used in either circumstance. For binomial data, logistic regression has greater interpretability and higher power than analyses of transformed data. However, it is important to check the data for additional unexplained variation, i.e., overdispersion, and to account for it via the inclusion of random effects in the model if found. For non-binomial data, the arcsine transform is undesirable on the grounds of interpretability, and because it can produce nonsensical predictions. The logit transformation is proposed as an alternative approach to address these issues. Examples are presented in both cases to illustrate these advantages, comparing various methods of analyzing proportions including untransformed, arcsine- and logit-transformed linear models and logistic regression (with or without random effects). Simulations demonstrate that logistic regression usually provides a gain in power over other methods.
Phylogenies and community ecology
DOI:10.1146/annurev.ecolsys.33.010802.150448 URL [本文引用: 2]
Interactive effects of climate warming and species diversity on the invasiveness of the alien weed Alternanthera philoxeroides
气候变暖和物种多样性交互效应对空心莲子草入侵的影响
Studies on Biological Invasion by Eupatorium adenpohorum
紫茎泽兰入侵生态学研究
Climatic ecological suitability and potential distribution for Boletus edulis in mountainous areas of Western Sichuan Plateau, China
川西高原山地美味牛肝菌气候生态适宜性及潜在分布
Impacts of black locust forest on understory plant species diversity and phylogenetic diversity in Shandong Province
山东刺槐林对林下植物物种多样性及谱系多样性的影响
Biodiversity of invaded area of Eupatorium adenophorum
紫茎泽兰(Eupatorium adenophorum)入侵地的生物多样性
Effect on species diversity of plant communities caused by invasion of Alternanthera philoxeroides in different habitats
喜旱莲子草对不同生境植物群落多样性的影响
Evidence for Elton’s diversity-invasibility hypothesis from belowground
The investigation report on the poisonous and injurious plant—Eupatorium adenophorum Spreng. in Sichuan Province
四川省毒害植物——紫茎泽兰调查报告
Predicting the spatial distribution of an invasive plant species (Eupatorium adenophorum) in China
DOI:10.1007/s10980-007-9096-4 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}