



从扩散的形式看, 有近距离扩散和远距离扩散两种。就节肢动物而言, 近距离扩散包括爬行(crawling)、下降(rappelling) (Hogg & Daane, 2018)等; 远距离扩散包括迁飞(migration)、飞航(ballooning) (Morley & Robert, 2018)等。关于昆虫的迁飞已有大量的研究(如Johnson, 1960; 江幸福等, 2016; 周燕等, 2020)。相似于昆虫的迁飞行为, 某些不具翅的节肢动物也能够借助自身产生的细丝来实现远距离扩散, 即飞航。多种节肢动物存在飞航行为, 如蜘蛛、叶螨、鳞翅目幼虫等(Bell et al, 2005; Stevens et al, 2019)。
众所周知, 蜘蛛是陆地生态系统中最丰富的捕食性天敌, 承担着生态平衡调控者的角色。蜘蛛的数量和分布直接与生态系统的功能相关, 而飞航在决定蜘蛛群落规模和物种组成方面具有重要意义(Suter, 1999; Gillespie et al, 2012), 尤其是一些受到干扰的环境(如农田系统)中, 飞航显得更加关键。以农田生态系统为例, 飞航蜘蛛是农田中主要的捕食者之一(Bishop & Riechert, 1990)。Hogg和Daane (2018)也指出蜘蛛飞航能力能够使得蜘蛛顺利到达农田系统, 且群落的再建立还依赖于其对栖息地的适应性和种群竞争能力。蜘蛛飞航的具体过程如何? 飞航机制又有哪些? 本文回顾了蜘蛛飞航的研究历程, 梳理了蜘蛛飞航的过程和影响因子, 比较了目前流行的几种关于飞航机制的解释, 并基于我国研究现状提出了蜘蛛飞航未来的研究方向。
1 蜘蛛飞航研究历程
蜘蛛飞航的研究最早可追溯到19世纪早期, Blackwall在1827年首次精确地记录了蜘蛛的飞航行为(Blackwall, 1827), 同时代的Darwin也在其专著中有过类似的描述(Darwin, 1845)。飞航作为一种非常有效的扩散方式, 尽管这些年来受到学者们的青睐(Richter, 1970; Greenstone et al, 1985, 1987; Szymkowiak et al, 2007; Cho et al, 2018), 但仍有许多问题尚未解决。在ISI Web of Science基本检索中以“spider balloon” “spider aerial” “spider aeronautic”作为检索词, 检索到1950-2019年间的文献有590篇, 去掉不相关的、重复的或关于螨类(spider mite)的结果, 共得到相关文献332篇。统计发现, 自1970年之后发文量逐年增加, 尤其是2000年之后; 近20年发文量占82.2% (图1)。
图1
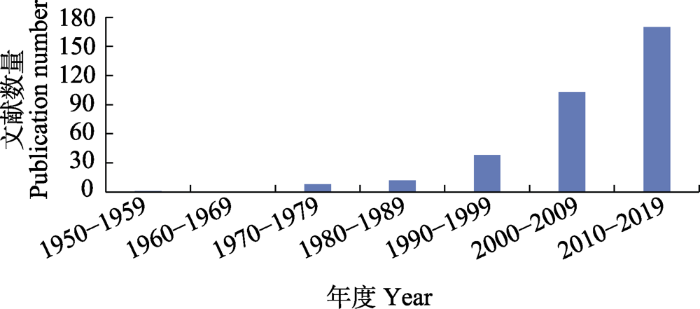
纵观已发表的文献资料, 蜘蛛飞航的研究历程大致可分为三个阶段: 第一阶段(1820s-1940s), 主要是对飞航现象的观察以及对飞航机制的猜测与假设, 以Blackwall (1827)、Darwin (1845)和Glick (1939)等为代表; 第二阶段(1950s-1990s), 更多关注飞航蜘蛛多样性及影响飞航的相关因子(如栖息地条件和气象因子), 以Vugts和van Wingerden (1976)、Greenstone等(1987)和Weyman (1993)等为代表; 第三阶段(2000s-), 深入分析飞航影响因子, 尝试构建飞航模型并提供验证飞航机制的试验证据, 以Reynolds等(2006, 2007)、Morley和Robert (2018)等为代表, 研究的广度和深度都明显增加。
表1 目前蜘蛛飞航研究中涉及的主要内容
Table 1
| 主要内容 Main contents | 关于内容的描述 Description on contents | 参考文献 References |
|---|---|---|
| 飞航的意义 Significance of ballooning | 蜘蛛扩散 Dispersal of spiders | Bristowe, 1939; Glick, 1939; Bell et al, 2005 |
| 飞航蜘蛛的多样性 Diversity of ballooning-spiders | 飞航蜘蛛组成及差异性 Composition and difference of spiders by ballooning | Greenstone, 1982; Greenstone et al, 1987; Schneider et al, 2001; Pearce et al, 2005; Blandenier, 2009; Lee et al, 2015 |
| 飞航过程和影响因素 Ballooning course and influencing factors | 栖息地因子、气象因子、大气电场 Habitat factors, meteorological factors, and atmospheric electric field | Richter, 1970; van Wingerden & Vugts, 1974; Vugts & van Wingerden, 1976; Greenstone, 1990; Bishop, 1990; Weyman, 1993; Bonte et al, 2003a; Morley & Robert, 2018 |
| 飞航机制解析 Analysis about spider ballooning | 模型、机制 Model and mechanism | Humphrey, 1987; Suter, 1991, 1999; Reynolds et al, 2006, 2007; Gorham, 2013; Cho et al, 2018; Morley & Robert, 2018; Stevens et al, 2019 |
2 飞航蜘蛛的多样性
2.1 飞航蜘蛛种类构成
许多科属的蜘蛛都有飞航行为, 目前已记录具有飞航行为的蜘蛛有27个科(表2)。有些研究明确记
录了具体飞航种类, 如狼蛛科的Pardosa ramulosa和P. tuoba (Greenstone, 1982)和园蛛科的斑络新妇(Nephila pilipes) (Lee et al, 2015), 但大多是见于多样性分析中。不同区域的飞航蜘蛛种类组成和丰富度存在差异。从全年的情况来看, 皿蛛科蜘蛛所占比例最高。Dean和Sterling (1985)在美国德克萨斯州利用Johnson-Taylor吸虫塔采集飞航蜘蛛, 鉴定出18个科的蜘蛛, 采集个体数量居于前3位的分别是皿蛛科、园蛛科和猫蛛科。Blandenier和Fürst (1998)在瑞士农田景观中利用吸虫塔做过类似的研究, 共采集到13个科的蜘蛛, 前3位分别是皿蛛科、逍遥蛛科和园蛛科。一般来说, 皿蛛是飞航的优势类群, 个体数量常居首位, 许多学者的研究结果均支持这一结论(Blandenier, 2009; Simonneau et al, 2016)。然而, 从具体月份的飞航蜘蛛组成来看, 结果又有所不同。Greenstone等(1987)分别于6-10月在美国密苏里州和11月在澳大利亚新南威尔士州采集飞航蜘蛛, 发现在这两个地点皿蛛都是优势科, 所占比例分别为42%和52%。而Bishop (1990)在美国田纳西州橡树岭9-10月采集的结果显示蟹蛛是优势科, 占比为65%, 皿蛛科仅占9%。
表2 目前已知的具有飞航行为的蜘蛛类群
Table 2
| 科名 Family | 种名 Species | 研究区域 Study area | 时间段 Time period | 参考文献 References | |
|---|---|---|---|---|---|
| 漏斗蛛科 Agelenidae | - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | |
| 近管蛛科 Anyphaenidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 园蛛科 Araneidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| Argiope bruennichi | 德国哈勒(室内实验) Halle, Germany (laboratory experiments) | - | Walter et al, 2005 | ||
| A. trifasciataandA. aurantia | 美国田纳西州 Tennessee, USA | 4-5月 Apr. to May | Tolbert, 1977 | ||
| Nephila pilipes | 新加坡乌敏岛(室内实验) Pulau Ubin, Singapore (laboratory experiments) | - | Lee et al, 2015 | ||
| 地蛛科 Atypidae | Sphodrosspp. | 美国北卡罗来纳州 North Carolina, USA | 3月 Mar. | Coyle, 1983 | |
| 管巢蛛科 Clubionidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 圆颚蛛科 Corinnidae | - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | |
| 螲蟷蛛科 Ctenizidae | Ummidiaspp. | 美国北卡罗来纳州 North Carolina, USA | 4月 Apr. | Coyle, 1985 | |
| 卷叶蛛科 Dictynidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 石蛛科 Dysderidae | - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | |
| 隆头蛛科 Eresidae | Stegodyphus mimosarum | 南非纳塔尔省 Natal, South Africa | 11月 Nov. | Wickler & Seibt, 1986 | |
| S. dumicola | 纳米比亚温得和克 Windhoek, Namibia | 1月 Jan. | Schneider et al, 2001 | ||
| 平腹蛛科 Gnaphosidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 栅蛛科 Hahniidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| 科名 Family | 种名 Species | 研究区域 Study location | 时间段 Time period | 参考文献 References | |
| 皿蛛科 Linyphiidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| Erigone atra | (室内实验) (laboratory experiments) | - | Bonte et al, 2008 | ||
| 光盔蛛科 Liocranidae | - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier & Fürst, 1998 | |
| 狼蛛科 Lycosidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| Pardosa amentata,P. lugubris,P. monticola,P. nigriceps,P. palustris,P. prativaga,P. pullata, andP. purbeckensis | 荷兰(室内实验) The Netherlands (laboratory experiments) | - | Richter, 1970 | ||
| P. ramulosaandP. tuoba | 美国旧金山 (室内实验) San Francisco, USA (laboratory experiments) | - | Greenstone, 1982 | ||
| Allocosa senex, A. marindia andSchizocosa malitiosa | 乌拉圭卡内洛内斯Canelones, Uruguay | 3-4月 Mar. to Apr. | Postiglioni et al, 2017 | ||
| 拟态蛛科 Mimetidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| 米图蛛科 Miturgidae | - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | |
| 蜜蛛科 Mysmenidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| 类球蛛科 Nesticidae | - | 中国岫岩Xiuyan, China | 5-9月 May to Sept. | 郑国等, 2011 | |
| 猫蛛科 Oxyopidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| 逍遥蛛科 Philodromidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 盗蛛科 Pisauridae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| 跳蛛科 Salticidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 肖蛸蛛科 Tetragnathidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 球蛛科 Theridiidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| 蟹蛛科 Thomisidae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
| - | 美国田纳西州 Tennessee, USA | 9-10月 Sept. to Oct. | Bishop, 1990 | ||
| - | 瑞士沃州 Canton de Vaud, Switzerland | 全年 The whole year | Blandenier, 2009 | ||
| Xysticus spp. | 德国 (室内实验) Germany (laboratory experiments) | - | Cho et al, 2018 | ||
| 妩蛛科 Uloboridae | - | 美国德克萨斯州 Texas, USA | 全年 The whole year | Dean & Sterling, 1985 | |
-表示文章中未记录蜘蛛具体种类。
- refers to no certain species of spider in references.
在统计飞航蜘蛛丰富度时, 需考虑季节的差异。Duffey (1956)提到了蜘蛛飞航具有季节性, 不同季节或气候条件下其丰富度不同。飞航行为多发生在5-10月, 冬季获取的飞航蜘蛛个体数量最少(Salmon & Horner, 1977; Dean & Sterling, 1985)。Simonneau等(2016)比较了连续3年(2000-2002年)的飞航蜘蛛采集数据, 发现无论是成体还是幼体其春末夏初的采集量都是最大的。Toft (1995)基于连续7年的采集数据发现, 温暖季节的蜘蛛倾向于远距离飞航, 而在早春和晚秋蜘蛛更多地进行近距离运动。除了物种组成之外, 不同飞航群体性别比例也存在差异, 有些雄性居多(Thomas & Jepson, 1999)、有些雌性居多(Duffey, 1956)或者雌雄相近(Meijer, 1977); 在年龄结构方面, 往往幼体居多, 所占比例甚至高达90%以上(Horner, 1975; Bishop, 1990; Postiglioni et al, 2017)。
2.2 飞航蜘蛛的个体大小
蜘蛛个体形态是限制飞航的一个重要因子(Humphrey, 1987), 尤其是个体大小。如上所述, 皿蛛是飞航蜘蛛的主体类群, 而其重要特征之一就是个体小; 飞航群体中幼体居多也从侧面体现了飞航对于蜘蛛重量和大小的要求。有研究表明, 蜘蛛飞航频率的差异可能与其重量有关(Weyman, 1993)。有的学者对飞航蜘蛛的重量和体型进行了试验性研究, 发现飞航蜘蛛的重量一般介于0.2-1.0 mg之间(Greenstone et al, 1987); 郑国等(2011)调查发现辽宁东部地区飞航蜘蛛体型小于3 mm的占91.44%。原蛛下目的蜘蛛同样具有飞航行为, 且个体都较新蛛下目蜘蛛略大(Coyle et al, 1985)。Coyle等(1985)发现Ummidiaspp.(螲蟷蛛科)和Sphodros. atlanticus(地蛛科)的重量分别是3.45 ± 0.13 mg (N = 9)、1.25 ± 0.03 mg (N = 15)。随着研究的深入, 一些学者认为较大个体同样能够进行飞航, 如隆头蛛科(Wickler & Seibt, 1986; Henschel et al, 1995; Schneider et al, 2001)、园蛛科(Lee et al, 2015)、蟹蛛科(Cho et al, 2018); 而且Wickler和Seibt (1986)提到Stegodyphus. mimosarum(隆头蛛科)重量最大能够达到150 mg, 体长接近12 mm。
尽管大个体蜘蛛也具有飞航行为, 但从飞航比例来看, 仍以小蜘蛛为主。Weyman等(1995)基于实验发现, 在农田生态系统中皿蛛科微蛛属(Erigone)的种类具备很强的飞航能力, 且飞航能够在各个季节、各种条件下进行。
3 蜘蛛飞航过程及影响因子
3.1 起飞‒空中航行‒降落
起飞(take-off)是蜘蛛飞航的首要步骤, 也是核心环节。目前学者们对蜘蛛飞航过程的研究重点也是蜘蛛的起飞, 因为这一环节更容易观察。Weyman (1993)提到蜘蛛不能直接从地上飞到空中, 需要到达高处(树枝或草丛)并进行预飞航。对于新蛛下目来说, 预飞航有两种形式: 一种是“脚尖式” (tip-toe), 即蜘蛛先爬到高处, 选好位置, 步足伸展, 尾部抬起并释放蛛丝(图2a); 另一种是“漂筏式” (dragline), 在蜘蛛爬到高处后, 通过蛛丝下降到一定高度, 然后再次释放蛛丝而实现飞行(图2b) (Bell et al, 2005; Sheldon et al, 2017; Cho et al, 2018)。还有一些原蛛下目的类群采用另一种漂筏式进行预飞航, 区别在于起飞时不释放蛛丝(Weyman, 1993)。目前鲜有文献比较两种预飞航方式在蜘蛛科属中所占比例及对后续飞航的影响。比较而言, “脚尖式”是一种更加主动(Richter, 1970)、更加进化的行为(Bell et al, 2005), 且同种蜘蛛可能只会采取其中的一种方式进行预飞航。
图2
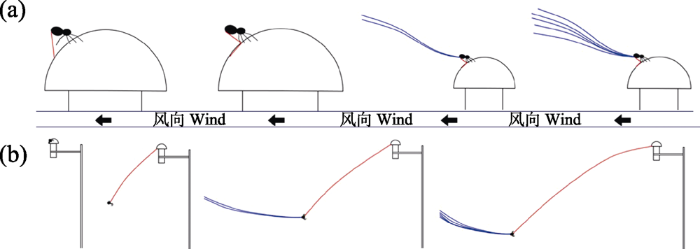
图2
飞航蜘蛛预飞航的两种方式(修改自Cho et al, 2018) (a) 脚尖式; (b) 漂筏式。红蓝色线都表示蛛丝, 红色蛛丝主要起到固定蜘蛛的作用, 蓝色蛛丝则参与飞航过程。
Fig. 2
Two ways of pre-ballooning of spiders (modified from Cho et al, 2018). a, Tiptoe; b, Dragline. Red and blue threads represent spider silk. The red ones are mainly related to the fixed function of spider and the blue ones take part in the course of ballooning.
目前, 对于空中航行(flight)和降落(setting)两个环节还鲜有报道。Thomas等(2003)指出, 如果天气条件适宜, 蜘蛛启动飞航, 且能够在6小时内飞航30 km; 如果考虑连续飞航, 水平方向上能够达到3,200 km (Gressitt, 1965); 在垂直方向上能够到达5 km (Foelix, 2011), 甚至在大气平流层也发现过飞航蜘蛛。不同蜘蛛的飞航能力不同, 整体来看结网型蜘蛛的飞航频率大于游猎型蜘蛛(Larrivée & Buddle, 2011)。飞航者能利用不同长度的蛛丝实现对飞航距离和降落点的控制(Humphrey, 1987)。然而, Suter (1999)认为“飞航者是否能够实现对飞航过程的有效控制”仍然是不清楚的, 因为大气环境条件是不可预测的。Reynolds等(2006)更进一步指出, 在混乱气流中飞航者对扩散距离的控制力较弱。
到底是什么原因促使了飞航的终止, 目前也缺乏证据(Weyman, 1993; Pearce et al, 2005)。有报道称蜘蛛对飞航的落点无法进行选择(Bonte, 2013), 但也有学者持不同观点, 如Tolbert (1977)发现当降落点环境状况不理想时, 蜘蛛会再次飞航, 直至到达合适位点; 某些情况下, 蜘蛛甚至可以连续飞航超过6次。Greenstone (1982)基于简易的飞航平台, 研究了两种狼蛛(Pardosa ramulosa和P. tuoba)的飞航行为, 发现P. ramulosa(栖息地适宜性低)的飞航频率高于P. tuoba(栖息地适宜性高), 肯定了栖息地适宜性与飞航频率间的相关性。总的来说, 在某些刺激下, 蜘蛛开启飞航过程; 而蜘蛛飞航的终止同样需要在内、外源因子的作用下发生。
3.2 影响蜘蛛飞航的主要因子
3.2.1 内源性因子
3.2.2 气象因子
如前所述, 20世纪中期, 学者们开始致力于研究引起蜘蛛飞航的关键因子, 尤其集中于外界刺激方面。气象因子(温度、湿度、风速等)、栖息地条件、大气电场等因子都会对蜘蛛飞航的各环节产生巨大影响。其中, 气象因子的研究最早也最深入。Duffey (1956)在英国牛津不同景观中研究蜘蛛飞航时, 就发现温度会影响飞航群体构成和飞航频率。Weyman (1993)综述了影响蜘蛛飞航的主要因子, 温度升降、湿度变化、气流运动、光强和压力等都曾被看作是蜘蛛飞航启动的刺激因素。当然, 目前普遍接受的观点是, 由太阳热量不均引起的空气对流能够启动飞航过程, 这是一种常见的飞航机制(见4.1.1)。当气象条件适宜蜘蛛起飞时, 飞航是一种非常有效的扩散方式(Reynolds et al, 2007)。
(1)温度。Duffey (1956)认为当温度上升且风小时, 蜘蛛飞航频率较高; Richter (1970)也发现当风速较小时(1.4 m/s), 蜘蛛飞航频率与温度间存在显著的正相关关系。在不同的温度下, 蜘蛛飞航频率是截然不同的, 如Bonte等(2003b)发现在20℃时黑微蛛(Erigone atra) (皿蛛科)飞航频率显著低于15℃时。温度对飞航的影响不仅体现在成体蜘蛛方面, 而且与幼蛛的关系也非常密切, 如Bonte等(2008)以黑微蛛为研究对象, 发现幼蛛发育温度能够强烈影响成体蜘蛛的扩散行为, 即当幼蛛在15℃时近距离扩散频率最低, 而在30℃时飞航频率最低。另外, 温差是否影响飞航也一度是学者们关注的问题, Duffey (1956)提出早晚温度差异无法解释飞航行为, 而Yeargen (1975)则认为蜘蛛飞航与最高温和最低温间温差有关。
在微气候的调节作用下, 蜘蛛会启动飞航过程, 而且主要是对飞航起飞环节产生影响。Weyman (1993)一方面肯定了各气象因子在蜘蛛飞航方面的作用, 同时也建议进行更广泛的研究来寻找影响飞航的其他因子。
3.2.3 栖息地条件
栖息地条件对蜘蛛飞航的影响表现在两个方面: 第一, 在不同的栖息地中飞航蜘蛛组成结构不同。Pearce等(2005)对比了澳大利亚昆士兰州大豆田和非作物区的飞航蜘蛛组成, 发现每天大豆田中飞航蜘蛛密度(14.8头/m2)高于非作物区(7头/m2)。Hogg和Daane (2018)比较了美国加利福尼亚州葡萄园和周围橡树林地中飞航蜘蛛构成, 结果显示葡萄园中的蜘蛛个体数量比橡树林中多25%。Blandenier等(2013)利用11年的吸虫塔数据, 调查了25种飞航蜘蛛的物候特征, 并探究了物候特征与栖息地因子间的关系, 发现生活于较封闭和稳定环境中的蜘蛛扩散呈现单峰模型。第二, 不同栖息地条件下蜘蛛飞航频率不同。当栖息地不适宜时, 蜘蛛倾向于启动飞航, 如Tolbert (1977)提到的连续飞航。Richter (1970)提出: 栖息地连续、面积大时, 蜘蛛倾向于短距离运动; 而在不连续或稀少的栖息地中, 蜘蛛会倾向于飞航以寻求新的栖息地。Greenstone (1982)和Postiglioni等(2017)的研究支持这一观点。Postiglioni等(2017)统计了3种狼蛛(Allocosa senex、A. marindia和S. malitiosa)的预飞航行为, 认为相比于泛化种, 栖息地特有种或处于破碎化栖息地中的种类具有更高的飞航频率。但也有学者认为栖息地空间异质性会降低飞航频率, 因为扩散到不适宜栖息地的风险增加了(Weyman et al, 2002)。Bonte等(2003a)也发现在破碎化栖息地中, 飞航方向性控制较差会直接导致远距离扩散成本增加, 因而窄域分布的物种具有较低的飞航频率。
3.2.4 大气电场
即使在晴朗的天气下, 大气中也经常存在着带电粒子。引起大气中分子电离的主要原因是贯穿整个大气的宇宙射线、高层大气中的太阳紫外辐射、低层大气中由地壳内的天然放射性物质发出的射线及人工放射性等(张三慧, 2009)。晴天时, 区域中的大气电场都指向下方, 地表附近的平坦地面上电场强度在100-200 V/m之间。大气电场强度随高度的增加而减小, 从而形成了大气电势梯度(atmospheric potential gradient, APG)。Wilson于1903年在Nature发表Atmospheric electricity的文章证实APG是广泛存在的。已有学者证实APG在昆虫传粉行为、昆虫间通信方面的作用(Clarke et al, 2013), 但关于APG对蜘蛛飞航影响的研究还较少。Gorham (2013)指出当蛛丝带电时, APG所产生的静电力能够实现飞航过程。Morley和Robert (2018)首次开展了蜘蛛电感受实验, 进一步证实了APG在蜘蛛飞航中的作用, 确认电场作用力足以驱动蜘蛛的飞航。
4 蜘蛛飞航机制解析
4.1 两种理论的博弈
在飞航机制方面, 目前普遍存在两种观点, 即空气动力理论(aerodynamics theory) (Blackwall, 1827; Greenstone, 1990; Cho et al, 2018)和大气静电理论(atmospheric electrostatic theory) (Murray, 1830; Gorham, 2013; Morley & Robert, 2018)。前者认为空气对流是蜘蛛起飞的主要驱动力; 而后者则认为大气电场的作用才是导致蜘蛛飞航的关键。实际上, 在著名的贝格尔号的第二次航行中(1831-1836年), Darwin曾观察到两类蜘蛛存在飞航行为, 小的个体约2-3 mm, 另外一种稍大个体约7 mm, 并在其论著中有如下描述(Darwin, 1845): 小蜘蛛从尾部发出一条丝, 然后迅速水平飞出; 大个体蜘蛛站在杆上, 同时发出4-5条蛛丝, 然后快速消失。Darwin曾推测, 空气对流对小蜘蛛的飞航起关键作用; 而大个体蜘蛛的飞航似乎与静电排斥有关。
4.1.1 空气动力理论
空气动力理论的基本观点是: 蜘蛛在一条(或数条)蛛丝的牵引下, 能够在上升气流的作用下向上运动(Blackwall, 1827)。从20世纪中期开始, 这一理论逐渐受到重视, 学者们发表了大量文章解释蜘蛛飞航的启动行为, 并分析飞航影响因子, 提出了飞航启动模型等(Duffey, 1956; van Wingerden & Vugts, 1974; Vugts & van Wingerden, 1976; Humphrey, 1987)。Greenstone (1990)则进一步基于实验证实了风在飞航启动中的作用, 发现热对流与蜘蛛飞航存在显著负相关关系, 但风速和飞航存在正相关关系。Cho等(2018)观察了蟹蛛(Xysticus spp., 10-25 mg)的飞航行为, 发现在近地气层中, 上升动力不是由热对流来提供, 而是由切变风产生的混乱气流来完成。
4.1.2 大气静电理论
随着飞航机制的深入研究, 有学者对空气动力理论产生了质疑, 并提出了如下问题: (1)飞航蜘蛛的数百条蛛丝如何形成扇形的蛛丝结构, 而且蛛丝间保持分离? (2)飞航蜘蛛如何能够快速地释放蛛丝, 是否有外力参与? (3)在极低的风速条件下, 蜘蛛如何获得非常高的起飞速度? (4)大个体蜘蛛的飞航中, 到底需要多少(或多长)的蛛丝来参与?
大气静电理论首先是Murray于1830年提出的, 基本观点是广泛存在的大气电势梯度(APG)能提供足够的静电力使得蜘蛛飞航。关于大个体蜘蛛的飞航, 对于穹蛛(Stegodyphusspp.)的研究最多, 如Wickler和Seibt (1986)曾观察到(S. mimosarum) (85-150 mg)具有飞航行为, 这个重量明显超出了基于空气对流的界限。Schneider等(2001)以另一种穹蛛(S. dumicola)为例, 得到了类似的结果, 推测可能是在静电的作用下启动了飞航。蜘蛛蛛丝从纺器喷出时能够迅速带电, 从而导致整个蛛丝间存在静电排斥, 在静电排斥的作用下, 数百条蛛丝形成了一个三角形的结构, 并给蜘蛛提供一种向上的分力。相比于空气动力理论, 这一理论一直未受到关注, 更谈不上量化层次。Morley和Robert (2018)以微蛛属(Erigone)蜘蛛为例, 统计了在两个APG梯度(1.25 kV/m和6.26 kV/m)下两种预飞航行为(tiptoe和dragline)的差异, 并发现电场的有无可以引起飞航蜘蛛向上或向下运动; 且比较了蜘蛛听毛在感受风、电场时所产生飞航行为的差异性。
4.2 常见的数学模型
蜘蛛起飞需要一个向上的分力, 而这个分力可能来自于热对流(也有风的作用)或者静电场。有的学者基于模型来模拟飞航的起飞动作, 用来找出完成起飞所需的最小指标。常见的有如下几种模型:
式中, (VZ)min为用于启动飞航的垂直方向最小速度(m/s); m为蜘蛛质量(mg); d和l分别为蛛丝的直径(um)和长度(m); u为空气黏度。
图3
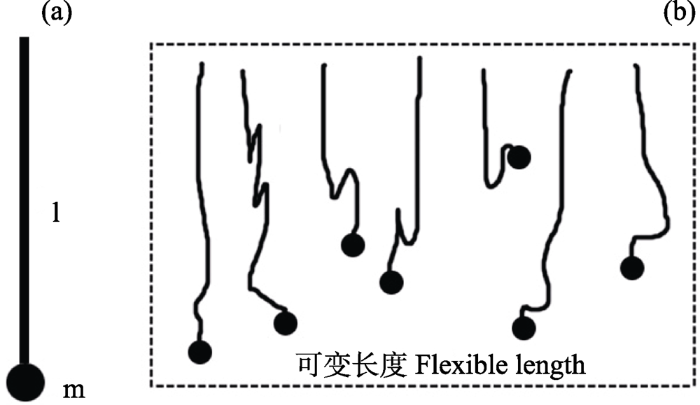
式中, Vsb为用于启动飞航的垂直方向速度(m/s); W为蜘蛛重量(μN; 1 μN ≈ 0.1 mg);L为蛛丝长度(m)。
式中, m0为蜘蛛质量(mg); mi为各个节点的质量(mi= ms/N, mg);ui为当地空气速度; τs为蛛丝的空气动力学响应时间; vi为蜘蛛从第1个节点i = 1到第N个节点i = N的速度集合; K为弹性常数; δi,0为克罗内克函数; pi,i+1为连接节点i、i+1的单位向量集合; si,i+1为在节点i、i+1上实际分离值和平衡分离值间的差异。
Gorham (2013)承认APG的存在, 并基于电场和电势, 给出了静电场模型和电量模型。
式中, E(h)为电场强度(V/m); E0为h = 0时电场强度, 即‒120 V/m; α为常数, 值为3 × 10‒4; h为高度(m); Qreq为用于启动飞航的电量; m为蜘蛛质量(mg); β为垂直方向初始加速度, 值为3-6 m/s2。
5 蜘蛛飞航研究的主要方法和技术
5.1 飞航蜘蛛获取方法
飞航蜘蛛的获取是研究其多样性、扩散的前提, 截至目前常用的方法包括粘贴法(sticky traps)、陷阱法(traps)、吸虫塔(suction traps) (图4)等。实际上, 这3种方法都是基于陷阱法原理, 只是捕获蜘蛛的具体过程略有区别。
图4
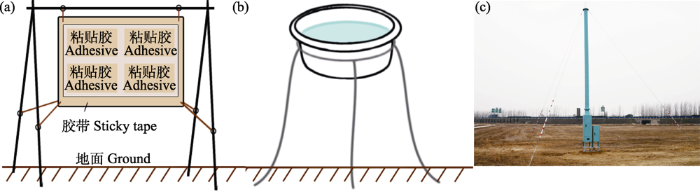
图4
飞航蜘蛛的3种采集方法。(a)粘贴法; (b)水域陷阱法; (c)吸虫塔(照片由李学军提供)。
Fig. 4
Three ways to capture ballooning spiders. (a) Sticky traps; (b) Water traps; (c) Suction traps (photo provided by Xuejun Li).
(1)粘贴法。这是较早使用的一种方法, 就是利用粘贴板捕获蜘蛛。在进行设置时, 一般会把粘贴板放置于离地1-2 m的高处, 并在粘贴板的外围固定胶带条, 以避免地面蜘蛛爬入而被收集(图4a)。
(2)陷阱法。利用各种形式的陷阱来实施捕获, 如飞机陷阱(airplane traps)、网陷阱(net traps)、旋转陷阱(rotary traps)、水域陷阱(water traps) (图4b)和瓶陷阱(bottle traps)等。Felt (1928)首次利用飞机陷阱法收集了空中的昆虫标本, Glick (1939)沿用此法在美国密西西比州Tallulah收集5年, 在20-15,000英尺的高度上共收集到3万余份昆虫和蜘蛛标本。网陷阱往往与粘贴法、飞机陷阱等结合使用, 如Reling和Taylor (1984)比较了4种网陷阱的优缺点, 并在152-610 m的高度进行了连续2年的收集试验, 确认了其有效性。Topping和Sunderland (1995)使用旋转陷阱(Topping et al, 1992)、水域陷阱和吸虫塔法(见下文)来采集飞航蜘蛛, 并基于4‒11月小麦田中的采集数据比较了3种方法的异同点。Pearce等(2005)分别用水域陷阱法和地面陷阱法在澳大利亚昆士兰州大豆田收集飞航蜘蛛和地面蜘蛛, 并比较了二者的群落组成差异。
5.2 飞航机制研究的技术手段
20世纪末期, 学者们开始利用多种手段和方法尝试模拟蜘蛛飞航过程, 探索飞航机制。首先, 在实验性工作的基础上, 结合物理学知识尝试并构建了诸多数学模型, 如Humphrey模型(1987)、Reynolds模型(2006)、Gorham模型(2013)等。模型具有简单化和直观化等特点, 而多学科交叉研究能够对飞航过程和飞航机制进行深入和细致地探索。另外, 有些学者在野外或实验室条件下, 人为搭建蜘蛛飞航平台, 对蜘蛛飞航动态进行精确监测。平台构建的优势是可以真实地观察到蜘蛛的飞航过程, 实现对各环节的严格把控。如Walter等(2005)对Follner和Klarenberg (1995)的观点提出质疑, 在实验条件下验证了横纹金蛛(Argiope bruennichi)二龄幼蛛的飞航行为, 并指出了飞航不是强制性的行为。Lee等(2015)首次在控制条件下验证了斑络新妇一龄幼蛛的飞航行为, 同时肯定了飞航在种群扩散中的意义和价值。Cho等(2018)通过人为平台模拟实验, 积累了大个体蜘蛛飞航的实验性数据。Morley和Robert (2018)则在人为构建蜘蛛飞航平台的基础上模拟了大气电场, 利用激光多普勒测振仪(laser Doppler vibrometry, LDV)对比研究了蜘蛛听毛在应对大气电场和风时的响应差异。总的来说, 目前飞航机制研究中的技术手段还相当有限, 相关研究也不是很多, 如何更广泛地开展这方面的研究并提出更优的方法, 是我们迫切需要解决的问题。
6 小结与展望
6.1 飞航是蜘蛛参与维持生态平衡的重要方式
蜘蛛是生态系统(尤其是农田)中重要的天敌, 在减少害虫威胁、增加农业产量、维持生态平衡等方面具有举足轻重的作用(Marc et al, 1999)。大多数蜘蛛是肉食性的(Foelix, 2011)。Nyffeler和Birkhofer (2017)报道蜘蛛每年吃掉的猎物总量约为4‒8亿吨。飞航蜘蛛是受害虫威胁的农田系统中的首批捕食者, 能够有效控制害虫的指数式增长(Sunderland et al, 1986)。Bishop和Riechert (1990)发现农田中约50%的蜘蛛来源于飞航; 据初步估算在4个月的生长季中, 每天每平方米农田中飞航蜘蛛数量至少达到0.18头。通过飞航扩散, 蜘蛛在不同空间上重新分布; 而且由于生活史、行为或应对外界刺激方式的差异, 会导致蜘蛛产生不同的空间扩散格局(Woolley et al, 2016)。飞航作为一种有效的扩散方式, 当农田中蜘蛛数量上升并达到足够数量时, 无疑会对猎物和农田产量产生巨大影响(Suter, 1999; Bell et al, 2005)。
6.2 未来研究方向和重点领域
针对目前研究现状, 笔者认为以后的工作重点应该立足于以下两个方面:
(1)深入蜘蛛多样性研究。一方面利用DNA条形码等技术, 查清飞航蜘蛛种类, 并归纳飞航蜘蛛总体特征。如前所述, 皿蛛作为飞航蜘蛛的优势类群, 与其个体大小直接相关, 是否也与皿蛛的结网特性(结网型蜘蛛种类最多)相关, 有待后续研究。另一方面, 利用多种方法分析飞航蜘蛛群体与环境因子间关系。利用不同采集法可以获得大量的、长期的飞航蜘蛛数据, 如何对这些数据进行深层次挖掘, 是未来研究的一个重要方面。如Blandenier等(2014)基于11年的调查数据, 采用非参数相关分析法探索了飞航蜘蛛数量高峰与气象因子间的关系。Simonneau等(2016)基于法国布列塔尼农田景观中2000-2002年吸虫塔采集数据, 利用广义线性模型分析了物候和气象变量对飞航蜘蛛数量的影响。
(2)鼓励多学科交叉, 探索新的方法和技术, 揭秘蜘蛛飞航全过程。无论是飞航模型的提出, 还是飞航机制的解析, 都离不开数学、物理等学科的共同参与。自然界任何一个生物过程都不是简简单单的, 其中包含着特定的规律性、复杂性, 而从复杂的现象中找到规律, 需要多学科知识的综合。飞航过程目前还有许多疑问无法解答, 迫切需要新方法和技术的涌现和应用。正如南非生物学家Sydney Brenner曾经说过的: 科学上的任何进展均离不开新的技术、新的发现和新的想法, 而且新的技术可能处于首位。
飞航究竟是属于主动行为还是被动行为呢? 这是飞航过程中一个值得讨论的问题。多数学者认为蜘蛛飞航属于被动行为(Bonte et al, 2003a; Szymkowiak et al, 2007; Blandenier et al, 2014), 因为在飞航的方向、速度、距离等方面都缺乏控制(Vugts & van Wingerden, 1976)。然而Bell等(2005)指出飞航者会对环境条件进行选择, 在最大可能成功扩散的情况下才启动飞航。Cho等(2018)也曾谈到蜘蛛飞航不是被动地、简单地依赖于风速条件的随机过程, 而是会主动抬起前足评价环境情况(持续时间5‒8秒), 并对起飞时间点进行主动选择的行为过程。另外也有报道称蜘蛛可以通过改变蛛丝的长度来对飞行时间和高度进行一定控制(Greenstone et al, 1985)。蜘蛛还可以对落点进行选择, 当落点不理想时, 甚至可以连续飞行超过6次(Tolbert, 1977)。从这些例证来看, 笔者倾向于认为飞航属于主动行为, 因为在飞航目的、飞航前准备、飞航过程及目的地选择中都有一定的主观性, 但未来需要开展更多的相关研究进一步证明。
参考文献
How to catch the wind: Spider hairs specialized for sensing the movement of air
Ballooning dispersal using silk: World fauna, phylogenies, genetics and models
Meteorological aspects of spider ballooning
Spider colonization of agroecosystems: Mode and source
Observations and experiments, made with a view to ascertain the means by which the spiders that produce gossamer effect their aerial excursions
Ballooning spiders caught by a suction trap in an agricultural landscape in Switzerland
Ballooning of spiders (Araneae) in Switzerland, general results from an eleven-year survey
Are phenological patterns of ballooning spiders linked to habitat characteristics?
Do spiders respond to global change? A study on the phenology of ballooning spiders in Switzerland
Low propensity for aerial dispersal in specialist spiders from fragmented landscapes
Environmental and genetic background of tiptoe-initiating behaviour in the dwarfspider Erigone atra
Thermal conditions during juvenile development affect adult dispersal in a spider
Cost-benefit balance of dispersal and the evolution of conditional dispersal strategies in spiders
In:
Observation of habits of ballooning spiders
蜘蛛飞航习性观察
An observational study of ballooning in large spiders, Nanoscale multifibers enable large spiders’ soaring flight
URL PMID:29902191 [本文引用: 11]
Detection and learning of floral electric fields by bumblebees
Aerial dispersal by mygalomorph spiderlings (Araneae, Mygalomorphae)
Ballooning behaviour of Ummidia spiderlings (Araneae, Ctenizidae)
Ballooning mygalomorphs: Estimates of the masses of Sphodros and Ummidia ballooners (Araneae: Atypidae, Ctenizidae)
Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage of H. M. S. Beagle Round the World, under the Command of Capt
Size and phenology of ballooning spiders at two locations in Eastern Texas
Aerial dispersal in a known spider population
Spider adaptation to artificial biotopes: The fauna of percolating filter beds in a sewage treatment works
Dispersal of insects by air currents
Aeronautic behaviour in the wasp-like spider, Argiope bruennichi (Scopoli) (Araneae, Argiopidae)
Long-distance dispersal: A framework for hypothesis testing
The distribution of insects, spiders, and mites in the air
Ballooning spiders, the case for electrostatic flight
https://arxiv.org/pdf/1309.4731v1.pdf. (accessed on 2020-07-10)
Ballooning frequency and habitat predictability in two wolf spider species (Lycosidae, Pardosa)
Spider ballooning: Development and evaluation of filed trapping methods (Araneae)
Ballooning spiders in Missouri, USA, and New South Wales, Australia, family and mass distributions
Meteorological determinants of spider ballooning: The roles of thermals vs. the vertical windspeed gradient in becoming airborne
Biogeography and ecology of land arthropods of Antarctica
In:
Dispersal mechanisms of Stegodyphus (Eresida): Do they balloon?
Aerial dispersal ability does not drive spider success in a crop landscape
Annual aerial dispersal of jumping spiders in Oklahoma (Araneae, Salticidae)
Fluid mechanic constraints on spider ballooning
Research advances and perspectives on migration-induced mechanisms promoting outbreaks of major Lepidopteran insect pests in China
中国主要鳞翅目农业害虫迁飞致灾机制研究与展望
A basis for a general system of insect migration and dispersal by flight
From individual dispersal to species ranges: Perspectives for a changing world
Ballooning propensity of canopy and understorey spiders in a mature temperate hardwood forest
Ballooning behavior in the golden orbweb spider Nephila pilipes (Araneae: Nephilidae)
Spiders (Araneae) useful for pest limitation and bioindication
The immigration of spiders (Araneida) into a new polder
Electric fields elicit ballooning in spiders
An estimated 400-800 million tons of prey are annually killed by the global spider community
Spider ballooning in soybean and non-crop areas of southeast Queensland
The dark side of ballooning: Nocturnal aerial dispersal in wolf spiders from the South American coastline
A new aphid-monitoring network system based on suction trapping and development of “green techniques” for aphid management
蚜虫新型预警网络的构建及其绿色防控技术研究
A collapsible tow net used for sampling arthropods by airplane
Ballooning dispersal in arthropod taxa with convergent behaviours: Dynamic properties of ballooning silk in turbulent flows
Ballooning dispersal in arthropod taxa: Conditions at take-off
DOI:10.1098/rsbl.2007.0109
URL
PMID:17389214
[本文引用: 3]

We have solved a long-standing and seemingly paradoxical set of questions that relate to the conditions which govern spider ballooning. We show that observations of spider ballooning excursions are best explained by meteorological conditions which maximize dispersal. Dispersal is predicted to be most effective in terms of distance when the stability of the atmosphere is non-ideally convective and is less effective during purely convective or neutrally stable conditions. Ballooners are most likely to travel a few hundred metres, but dispersal distances of several hundred kilometres are possible.
Aerial dispersal in relation to habitat in eight wolf spider species (Pardosa, Araneae, Lycosidae)
Aerial dispersion of spiders in North Central Texas
Dispersal of Stegodyphus dumicola (Araneae, Eresidae): They do balloon after all!
Revisiting the Physics of Spider Ballooning
Phenological and meteorological determinants of spider ballooning in an agricultural landscape
DOI:10.1016/j.crvi.2016.06.007
URL
PMID:27527898
[本文引用: 5]
Spiders are known to commonly use aerial dispersal, so-called ballooning, especially at juvenile stages. They produce a silk thread that allows them to rise up in the air to disperse, which serves as inbreeding avoidance or to find an optimal over-winter habitat. Studies of phenology, species and meteorological factors associated with aerial dispersal have been limited to laboratory settings, with few data obtained under natural settings and no studies to date executed in France. To understand aerial dispersal, we conducted daily sampling between 2000 and 2002 at a height of 12m. For adults, high proportions of
Ballooning spiders over the East China Sea
东海上空的飞航蜘蛛
Distribution of linyphiid spiders in relation to capture of prey in cereal fields
Ballooning in spiders: Results of wind tunnel experiments
An aerial lottery, the physics of ballooning in a chaotic atmosphere
Passive dispersal in arachnids
Modeling aerial dispersal of linyphiid spiders
Differential aerial dispersal of linyphiid spiders from a grass and a cereal field
Aerial activity of linyphiid spiders: Modelling dispersal distances from meteorology and behaviour
Validation of a simple method for monitoring aerial activity of spiders
Two functions of gossamer dispersal in spiders
In:
A large improved rotary trap for sampling aerial invertebrates
Methods for monitoring aerial dispersal by spiders
In:
Factors influencing aeronautic behaviour of spiders
Meteorological aspects of aeronautic behavior of spiders
The wasp spider Argiope bruennichi (Arachnida, Araneidae): Ballooning is not an obligate life history phase
A review on dispersal behavior of small mammals
小型哺乳动物的扩散行为研究进展
A review of the possible causative factors and significance of ballooning in spiders
The effect of food deprivation on aeronautic dispersal behaviour (ballooning) in Erigone spp. spiders
Do seasonal changes in numbers of aerially dispersing spiders reflect population density on the ground or variation in ballooning motivation?
DOI:10.1007/BF00329428
URL
PMID:28306964
[本文引用: 1]
Groups of linyphiid spiders (Erigone spp.) (Araneae, Linyphiidae), collected at intervals from arable land, were tested in laboratory bioassays to determine the proportion of individuals that exhibited ballooning behaviour on each field sampling occasion. There was no significant variation in the proportions of spiders in each test group ballooning in the laboratory over a year. Investigations of ground density and aerial dispersal, in a grass field and a winter-wheat field, confirmed that peaks in numbers of spiders observed ballooning in the field generally coincided with population peaks in the summer and autumn. Significant correlations between ground populations and aerial catches were found for total spiders, immature spiders, and Bathyphantes gracilis (Blackwall) in both fields, adult spiders in the grass field, Erigone spp. in the grass, and Meioneta rurestris (C.L. Koch) in the wheat. The other groups analysed, Lepthyphantes tenuis (Blackwall) in both fields, Erigone spp. in wheat, and M. rurestris in grass, showed similar but non-significant trends. No significant difference was found between overall ground-to-air ratios for males compared to females, but adult spiders were more likely to balloon than immatures.
A review of the evolution and mechanisms of ballooning by spiders inhabiting arable farmland
Aerial dispersal by ballooning in adult Stegodyphus mimosarum
Atmospheric electricity
A novel trap to capture ballooning spiders
Aerial dispersal activity of spiders sampled from farmland in southern England
Factors influencing the aerial dispersal of spiders (Arachnida: Araneida)
Flying spiders: Simulating and modeling the dynamics of ballooning
Community composition and its characteristics of ballooning spiders in east Liaoning Province of Northeast China
辽宁东部地区飞航蜘蛛的群落组成及特征
Frequency of migration of agricultural pests across the Bohai Sea in northern China and a control strategy for these species
农业害虫跨越渤海的迁飞规律与控制策略

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}