



自然杂交是生物演化和物种形成的主要动力之一, 在植物界中普遍存在(Barton, 2001; Mallet, 2007; Abbott et al, 2013)。Mallet (2005)曾对相关研究进行了统计, 发现存在种间杂交或基因渐渗的植物物种数量占整体的25%以上。虽然并非所有的杂交事件最终都能产生新的物种, 但健康且可育的杂交个体一旦与亲本物种之间产生生态位分化, 就可能摆脱“少数细胞型劣势”, 不断演化形成独立的物种(Husband, 2000; Mallet, 2007)。随着大量杂交物种被鉴定, 代表谱系融合的杂交物种形成在物种多样性发生中的重要作用已得到普遍认同(Mallet, 2007; 王玉国, 2017; Taylor & Larson, 2019)。
根据杂交物种的倍性是否增加, 可以将其成种过程分为异源多倍化杂交物种形成(allopolyploid hybrid speciation)和同倍体杂交物种形成(homoploid hybrid speciation)。异源多倍化杂交物种是指两个物种杂交产生的染色体数量加倍的新物种, 它的形成相对常见, 这是因为倍性差异不仅保证了杂交个体具有正常的育性, 还可以使其与亲本物种之间快速产生较强的生殖隔离, 加快物种形成(Soltis et al, 2004; Mallet, 2007)。与之相比, 同倍体杂交物种的形成条件较为苛刻。同倍体杂交物种是指染色体倍性相同的物种之间通过杂交产生染色体倍性未发生变化的新物种, 由于倍性相同, 新产生的杂交后代不能立即阻断来自亲本的基因流, 在处于少数细胞型劣势的情况下很可能在与亲本的不断回交中消失(Buerkle et al, 2000)。由此可见, 杂交后代与其亲本之间是否能够产生生殖隔离, 是同倍体杂交物种形成的关键。越来越多的研究表明生态分化可能是导致同倍体杂交物种形成中产生生殖隔离的主要原因(Gross & Rieseberg, 2005; Mallet, 2007; Yakimowski & Rieseberg, 2014)。然而, 目前已经报道的同倍体杂交物种形成案例相对整个植物界而言非常少, 并且除向日葵属(Helianthus) (Rieseberg, 1991; Rieseberg et al, 2003)、高山松(Pinus densata) (Ma et al, 2006; Zhao et al, 2014)、居中虎榛子(Ostryopsis intermedia) (Wang et al, 2021)和峨眉锥栗(Castanea henryi var. omeiensis) (Sun et al, 2020)等少数研究以外, 大多数研究还处于描述性阶段, 缺乏对同倍体杂交物种形成机制的深入探讨。因此, 未来需要鉴定更多不同类型的杂交物种及种间杂交带, 这样才能更好地解释杂交对适应性产生及生殖隔离的影响, 更深入地探讨杂交物种形成不同阶段的遗传基础, 尤其是维持杂交物种表型变异相关的适应性位点(王玉国, 2017)。
红砂(Reaumuria soongarica)是柽柳科红砂属内无特殊分类地位的物种(Zhang et al, 2014a), 是起源于第三纪的超旱生灌木, 属于典型的地带性植物, 其分布区与亚洲中部干旱区基本重合(刘家琼等, 1982; Shi et al, 2013)。作为荒漠区重要的优势种和建群种, 红砂在维持温带荒漠生态系统的物种多样性和稳定性方面发挥着重要作用(刘家琼等, 1982; Ma et al, 2005)。红砂的遗传多样性较高, 其种内存在明显的遗传分化和杂交(Qian et al, 2008; Yang et al, 2015; Shi et al, 2020), 并且不同遗传支系(种内分化出来的具有不同遗传组分的谱系)的形态和生境已出现差异(Fan et al, 2020) (附录1)。目前, 红砂的西部支系(塔克拉玛干沙漠)与东部支系(巴丹吉林-腾格里沙漠)之间已完全分化, 该过程受到青藏高原东北部隆升以及东亚季风系统形成和演化的驱动(Li et al, 2012; Yin et al, 2015; Shi et al, 2020)。Shi等(2020)进一步指出红砂北疆支系(古尔班通古特沙漠)起源于东、西支系经两次杂交事件产生的同倍体杂交后代, 目前该支系与其亲本之间已出现明显的生态分化, 正处于物种形成的早期阶段。不同于那些已经和亲本产生有效生殖隔离的同倍体杂交物种, 如高山松(Zhao et al, 2014), 红砂北疆支系为研究杂交物种形成初期的生态适应性变化和生殖隔离的产生等关键过程提供了新的模型。此外, 相比于我国其他沙漠地区, 古尔班通古特沙漠植物区系形成时间较晚, 但物种多样性较高(刘媖心, 1995; 张立运和陈昌笃, 2002), 红砂北疆支系与之相同的特点(后衍支系却具有较高的群体遗传分化程度)对于研究该地区物种多样性的发生机制具有重要价值。但令人困惑的是, 红砂北疆支系内不同群体的多个叶绿体基因和核基因的系统发育关系相互矛盾(Yin et al, 2015; Shi et al, 2020)。比如, 沙湾群体中, 母系遗传的叶绿体基因单倍型与东部支系聚为一支, 双亲遗传的ITS单倍型与西部支系相近; 而火烧山群体的叶绿体基因单倍型则与西部支系聚为一支, ITS单倍型与东部支系相近。目前红砂属中仅兰州地区红砂群体的倍性已被调查, 该群体为二倍体(2n = 2x = 22), 与大多数已报道的柽柳科柽柳属(Tamarix)的物种的倍性相同, 同时在一些柽柳属物种中还存在多倍体植株(Wang et al, 2011; Samadi et al, 2013)。在环境恶劣的地区, 多倍化可能是植物适应极端环境的重要机制(Van de Peer et al, 2021)。考虑到Shi等(2020)分别对来自北疆阜康群体及两个亲本支系的共3个个体进行了核型调查, 但群体水平的倍性变化尚不清楚, 在无明显地理隔离但存在环境差异的狭窄区域内, 同为杂交后代的北疆群体之间存在显著的遗传障碍(Fan et al, 未发表数据), 因此我们推测倍性差异是导致北疆群体间产生生殖隔离的主要原因之一。
基因组大小(DNA C-值)是指一个物种的配子核中(染色体数目为n)未复制的DNA含量, 是重要的生物多样性参数, 在物种水平上较为固定, 而在物种间存在较大差异(Greilhuber et al, 2005; Smarda & Bures, 2010)。因此, 该值可用于判断倍性水平、辅助物种分类和鉴定杂交物种(Mahelka et al, 2005; Loureiro et al, 2010; 邹璇等, 2020; Urfusová et al, 2021), 如Zonneveld和Duncan (2010)利用DNA C-值数据发现了风信子科的一个新二倍体物种Eucomis grimshawii; Mahelka等(2005)利用DNA C-值区分了偃麦草属(Elytrigia)的E. repens和E. intermedia, 并发现了两个物种的同倍体杂交种。由于上述的成种特点, 同倍体杂交物种的DNA C-值一般介于两个亲本之间, 而异源多倍化杂交物种的DNA C-值接近两个亲本的DNA C-值之和(Loureiro et al, 2010)。此外, DNA C-值在植物种内和种间的变异对于研究生态适应性、系统发育和基因组进化等有重要意义(郭水良等, 2011; Guignard et al, 2016; Pellicer et al, 2018)。
1 材料与方法
1.1 样品采集
根据前期有关红砂群体遗传学的研究结果(Yin et al, 2015; Shi et al, 2020), 并充分考虑野外采样工作的可操作性, 选择新疆阿克苏(AKS)和策达雅(CDY)作为红砂西部支系的代表样地, 甘肃红古(HG)和宁夏四马营(SMY)作为东部支系的代表样地, 新疆阜康(FK)、沙湾(SW)、火烧山(HSS)和五彩城(WCC)作为北疆支系的代表样地(表1, 图1)。2018年秋在上述每个样地内随机采集20株红砂的种子和叶片, 并保证单株之间至少相距30 m。此外, 我们还在青海德令哈(DLH)采集了五柱红砂(R. kaschgarica)的样本用于红砂属内基因组大小比较。采集到的植物样本立即用硅胶干燥, 带回甘肃省寒区旱区逆境生理与生态重点实验室后放入种子低湿贮存柜中进行长期保存。经DNA条形码(叶绿体atpH-atpL和ndhA序列)鉴定技术分析(Yin et al, 2015), 所有样本的物种信息与野外采集时记录的信息一致。由于野生番茄(Solanum pimpinellifolium)的DNA C-值(739 Mb; The Tomato Genome Consortium, 2012)接近Wang等(2011)测量的兰州红砂群体的DNA C-值(778 Mb), 因此本研究选用野生番茄作为流式测定的内标物种, 其种子取自中国科学院植物研究所。
表1 红砂采样群体的基本信息
Table 1
| 简称 Code | 地理位置 Location | 经纬度 Locality | 海拔 Altitude (m) | 年均降水量 Mean annual precipitation (mm) | 遗传支系 Lineage |
|---|---|---|---|---|---|
| HG | 甘肃红古 Honggu, Gansu | 103.03° E, 36.26° N | 1,775 | 369 | 东部支系 Eastern |
| SMY | 宁夏四马营 Simaying, Ningxia | 106.01° E, 38.50° N | 1,175 | 193 | 东部支系 Eastern |
| AKS | 新疆阿克苏 Akesu, Xinjiang | 79.70° E, 41.57° N | 1,491 | 164 | 西部支系 Western |
| CDY | 新疆策达雅 Cedaya, Xinjiang | 84.86° E, 42.01° N | 1,078 | 80 | 西部支系 Western |
| SW | 新疆沙湾 Shawan, Xinjiang | 85.33° E, 44.32° N | 519 | 150 | 北疆支系 Northern |
| FK | 新疆阜康 Fukang, Xinjiang | 88.13° E, 44.32° N | 491 | 170 | 北疆支系 Northern |
| HSS | 新疆火烧山 Huoshaoshan, Xinjiang | 88.99° E, 44.86° N | 478 | 155 | 北疆支系 Northern |
| WCC | 新疆五彩城 Wucaicheng, Xinjiang | 88.99° E, 45.16° N | 773 | 176 | 北疆支系 Northern |
图1
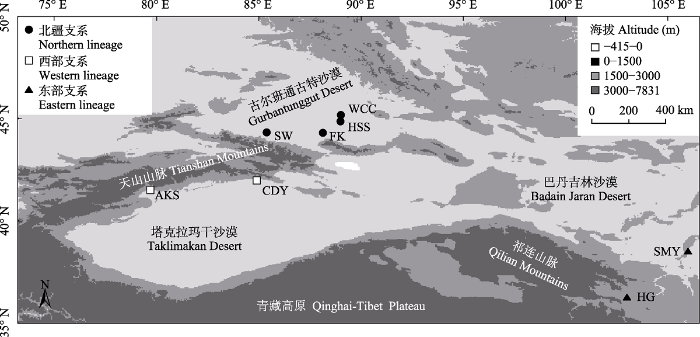
图1
本研究测量的8个红砂群体的地理分布图。群体的具体信息见
Fig. 1
Geographic locations of the eight Reaumuria soongarica populations measured in this study. The specific information of the populations is shown in
1.2 种子萌发
首先, 从每个群体内随机挑选2个单株的种子, 置于‒20℃下贮藏一周以削弱种子休眠(石勇等, 2016), 而后在常温下用蒸馏水浸泡18 h, 再用75%酒精浸泡1 min, 蒸馏水冲洗后再用3% NaClO消毒30 s, 经无菌水反复冲洗3次后置于铺有双层滤纸的培养皿中, 并加入适量无菌水使滤纸完全浸湿。之后将培养皿放入25℃的恒温培养箱中, 黑暗条件下进行萌发, 48 h后在16 h/8 h的光周期下继续培养备用。在此期间, 每两天补水一次, 以保持滤纸浸湿。
1.3 流式细胞仪检测
红砂细胞核悬液的制备利用了CyStain UV Precise P试剂盒。具体步骤如下: 收集来自同一母本的幼苗, 剪取其幼嫩的根, 混匀后称取0.2 g; 将材料放入加有250 μL细胞核解离液(nuclei extraction buffer)的培养皿中, 用锋利的刀片快速将根切碎, 混匀后孵育30-90 s; 混合液经孔径50 μm的滤膜过滤至流式专用管中。在测定细胞核DNA含量之前, 向上述细胞核悬液中加入500 μL染液(495.5 μL Staining Buffer + 3 μL Propidium Iodide (PI, 碘化丙啶) + 1.5 μL RNaseA), 并在避光条件下孵育15 min。五柱红砂和番茄细胞核悬液的制备也采用同样的方法。
本实验使用BD (美国)公司的LSRFortessa流式细胞仪对所有样品的细胞核DNA含量进行测定, 采用的激发波长为488 nm, 收集并分析样本中PI发射的荧光信号。首先, 对所有样品的细胞核悬液进行单独检测, 确定各自PI荧光强度的大致范围, 以优化仪器的电压、阈值等检测参数。之后, 将内标物种番茄的细胞核悬液分别与每个待测样品的细胞核悬液等比例混合, 混匀后上机检测, 每个样品重复检测3次, 并使每次检测中低速收集到的细胞个数大于10,000个, 变异系数小于5%。最终, 红砂样品的检测电压全部为445 V; 五柱红砂单独检测的电压为400 V, 混合样品的测定电压为430 V。
利用流式细胞术检测植物样本的DNA倍性而非DNA绝对含量时, 硅胶干燥的样品也可以得到理想的峰图(Suda & Trávníček, 2006; Wan et al, 2019)。由于红砂分布范围广, 对于大多数红砂群体而言, 难以在野外对样本进行保鲜处理并带回实验室保存。此外, 红砂种子的收集也存在一定难度。红砂不同支系的花期差异显著(Fan et al, 2020); 种子一旦成熟, 会在其表面密布长柔毛的协助下随风传播(张元明等, 1998)。因此, 要想获得更多红砂群体的倍性数据, 最简单且经济的办法就是利用野外采集并干燥处理的红砂叶片进行流式测定。为了检测红砂干燥叶片替代新鲜材料用于倍性测定的可行性, 本研究还制备了AKS红砂干燥叶片(采种期间收集)的细胞核悬液并上机检测, 检测电压为445 V。
1.4 数据分析
使用软件BD FACSDiva 7.0对流式细胞仪检测数据进行分析和制图。由于本实验采用已知基因组大小为739 Mb的野生番茄(2x)作为内标, 并且PI嵌入细胞核DNA双链中的量与DNA含量成正比, 因此待测样品的细胞核DNA含量(2C, 单位为pg)按照以下公式计算(Doležel & Bartoš, 2005):
其中, R为待测红砂属植物样本G0/G1期细胞所形成的DNA含量峰(在所有图中标注为P2峰)的荧光强度, L为内标番茄G0/G1期峰(标注为P1峰)的荧光强度, 1 pg DNA约等于978 Mb (Doležel et al, 2003)。
2 结果
2.1 红砂东、西支系的DNA C-值测定
图2
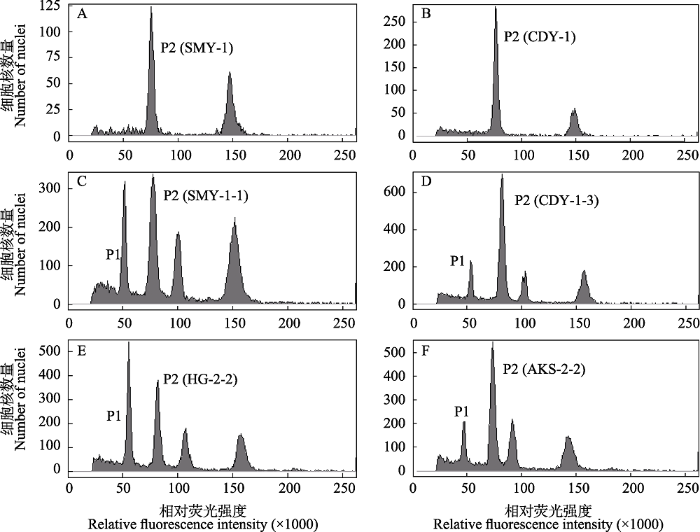
图2
红砂东西支系的流式测定结果。A, B: SMY和CDY群体的样品分别单独上机检测; C-F: 每个红砂群体与内标番茄的混合样品的检测结果, 包括SMY、CDY、HG和AKS群体。P1代表番茄的G0/G1峰, P2代表红砂的G0/G1峰。群体代号见
Fig. 2
Test results of different populations of the eastern and western lineages of Reaumuria soongarica using flow cytometry. A, B, The samples of SMY and CDY populations were measured without the internal standard; C-F, the mixed samples of Solanum pimpinellifolium and each Reaumuria soongarica population were tested, including SMY, CDY, HG, and AKS populations. P1 represents the nuclear DNA content of S. pimpinellifolium in G0/G1 phase, and P2 represents that of R. soongarica samples in G0/G1 phase. The population codes see
表2 红砂属植物样品的流式细胞仪测定结果(平均值 ± 标准差)
Table 2
| 支系 Lineage | 群体 Population | 红砂属样品G0/G1峰值 Peak values of Reaumuria (G0/G1) | 内标番茄G0/G1峰值 Peak values of Solanum pimpinellifolium (G0/G1) | DNA 1C-值 DNA 1C-values (pg) | 倍性 Ploidy | 一倍体基因组大小 Monoploid genome size (Mb) |
|---|---|---|---|---|---|---|
| 西部支系 Western lineage of R. soongarica | AKS | 74,192.667 ± 1,381.685 | 45,988.667 ± 1,264.577 | 1.219 ± 0.025 | 2n = 2x | 1,192.639 ± 24.701 |
| CDY | 76,255.167 ± 2,643.503 | 49,239.500 ± 1,725.806 | 1.170 ± 0.005 | 2n = 2x | 1,144.481 ± 4.816 | |
| 平均值 Average | - | - | 1.195 ± 0.031 | 2n = 2x | 1,168.560 ± 30.338 | |
| 东部支系 Eastern lineage of R. soongarica | HG | 75,210.667 ± 7,241.285 | 49,762.333 ± 5,100.729 | 1.143 ± 0.014 | 2n = 2x | 1,117.586 ± 13.338 |
| SMY | 74,570.500 ± 2,960.292 | 48,737.333 ± 1,845.872 | 1.156 ± 0.006 | 2n = 2x | 1,130.655 ± 5.787 | |
| 平均值 Average | - | - | 1.149 ± 0.012 | 2n = 2x | 1,124.120 ± 11.944 | |
| 北疆支系 Northern lineage of R. soongarica | SW | 77,156.167 ± 3,492.550 | 49,210.000 ± 1,972.505 | 1.185 ± 0.009 | 2n = 2x | 1,158.498 ± 9.009 |
| FK | 68,354.000 ± 4,150.933 | 44,074.167 ± 2,919.187 | 1.172 ± 0.013 | 2n = 2x | 1,146.490 ± 12.237 | |
| HSS | 141,698.000 ± 8,912.053 | 46,786.333 ± 2,805.389 | 2.288 ± 0.015 | 2n = 4x | 1,118.938 ± 7.437 | |
| WCC | 144,188.167 ± 5,140.113 | 47,052.500 ± 1,598.277 | 2.316 ± 0.023 | 2n = 4x | 1,132.291 ± 11.403 | |
| 五柱红砂 R. kaschgarica | DLH | 200,628.333 ± 1,704.432 | 38,574.000 ± 351.629 | 3.930 ± 0.006 | - | - |
| 红砂干燥叶片 Dehydrated leaves of R. soongarica | AKS | 72,426.000 ± 2,382.788 | 43,371.000 ± 1,417.229 | 1.262 ± 0.014 | 2n = 2x | 1,234.122 ± 14.060 |
一倍体基因组大小对应DNA Cx值; 除标注采用干燥叶片的AKS群体以外, 其余群体均采用幼嫩的根为材料。群体简称同
The monoploid genome size corresponds to the DNA Cx-value. Except for the AKS population labeled with dry leaves, the experimental material of all the other populations was young roots. The codes of the populations are the same as
2.2 红砂北疆支系的DNA C-值测定
图3
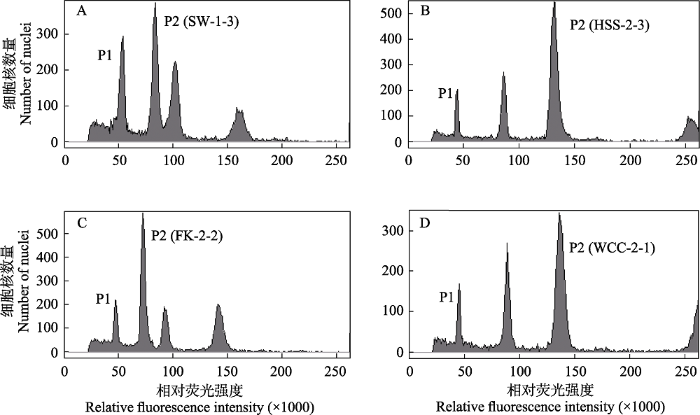
图3
番茄和不同北疆红砂群体混合样品的流式测定结果。(A) SW群体; (B) HSS群体; (C) FK群体; (D) WCC群体。P1代表番茄的G0/G1峰, P2代表红砂的G0/G1峰。
Fig. 3
Test results of the mixed sample of Solanum pimpinellifolium and different populations of the northern lineage of Reaumuria soongarica using flow cytometry. The populations of the northern lineage include SW (A), HSS (B), FK (C), and WCC (D). P1 represents the nuclear DNA content of S. pimpinellifolium in G0/G1 phase, and P2 represents that of R. soongarica samples in G0/G1 phase.
2.3 红砂干燥叶片进行流式测定的可行性
在AKS红砂干燥叶片单独进行流式测定的结果中, 虽然能检测到明显的G0/G1期峰, 但峰图的变异系数大于10% (图4A)。不同于AKS新鲜幼嫩组织的峰图, 当干燥叶片与内标番茄一同上机检测时, 红砂叶片样品的G0/G1期峰与番茄的G2/M期峰重叠在一起, 二者不易区分, 并且番茄的G0/G1期峰不明显(图4B), 说明干燥叶片样本的细胞核悬液中DNA碎片或者杂质较多从而影响了信号峰的形成。虽然干燥叶片峰图的稳定性和准确性还需进一步提升, 但是利用干燥叶片测定得到AKS红砂的DNA 1C-值(1.262 ± 0.014 pg)接近其新鲜样本的1C-值(1.219 ± 0.025 pg), 该结果可以准确判断样本的倍性。可见利用红砂干燥叶片检测不同群体的倍性合理可行, 但采样时最好收集嫩叶。
图4
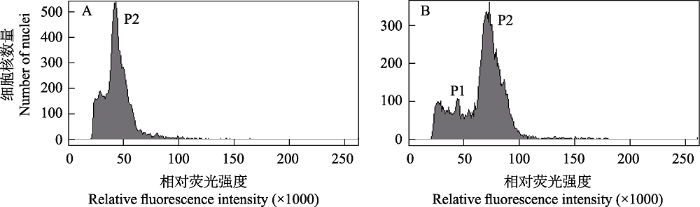
图4
红砂AKS群体干燥叶片样品的DNA含量直方图。(A)单独的红砂叶片样品; (B)红砂叶片(P2, G0/G1)和内标番茄(P1, G0/G1)的混合样品。
Fig. 4
Histograms of the nuclear DNA content of Reaumuria soongarica dry leaves (the AKS population). (A) R. soongarica sample alone; (B) the mixed sample of R. soongarica (P2, G0/G1) and Solanum pimpinellifolium (P1, G0/G1).
2.4 五柱红砂DNA C-值的测定结果
图5
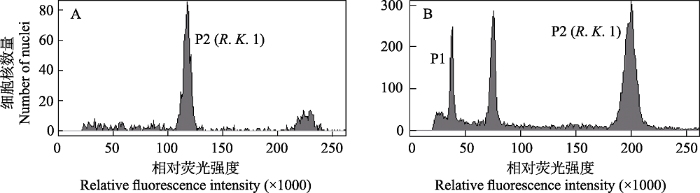
图5
五柱红砂样品的DNA含量直方图。(A)单独的五柱红砂样品(R. K. 1); (B)五柱红砂(P2, G0/G1)和内标番茄(P1, G0/G1)混合样品。
Fig. 5
Histograms of the nuclear DNA content of Reaumuria kaschgarica. (A) The pure sample of R. kaschgarica; (B) the mixed sample of R. kaschgarica (P2, G0/G1) and Solanum pimpinellifolium (P1, G0/G1).
3 讨论
3.1 红砂与柽柳科植物DNA C-值比较
然而, 本研究测定的红砂DNA C-值与Wang等(2011)报道的数据相差较大。Wang等(2011)以红砂嫩叶为材料, 选取水稻作为内标, 通过流式细胞术测得采集自兰州的二倍体红砂样本的DNA 1C-值为0.796 pg (1 pg = 978 Mb), 约为本研究结果的65%-70%。这两项研究相比, 对测定结果有重要影响的细胞核解离液、植物材料和内标物种(汪艳等, 2015)都不同。由于不同植物组织的结构和内含物不同, 当选用的解离液与植物材料不匹配时, 会使细胞核悬液中存在较多碎片和杂质, 从而影响流式测定结果的准确性(Doležel et al, 2007; 邹璇等, 2020)。本研究以幼嫩的根为材料, 所得流式峰图清晰且主峰的变异系数均小于5%, 该结果与Wang等(2011)给出的峰图效果一致, 表明两项研究中红砂幼嫩的叶片和根均有良好的裂解效果。此外, 无论以番茄还是水稻作为内标, 内标峰与红砂样品的DNA含量峰均不重叠, 表明内标未影响到红砂DNA含量峰的形成。造成这两项研究的测定结果存在较大差异的原因可能有两点: (1)在Wang等(2011)测定的流式峰图中, 红砂样品峰值对应的细胞核数量不足20个, 远小于其他研究中的数量(Doležel et al, 2007; 林峰等, 2018; 邹璇等, 2020), 细胞核较少可能会影响统计结果的准确性; (2) Wang等(2011)以水稻为内标, 基因组更小(389 Mb), 在估计远大于内参基因组的物种时, 结果的误差更大。以Wang等(2011)的估计来看, 如果红砂基因组大小接近水稻的两倍, 就应当与本研究选取的番茄基因组大小相近(739 Mb); 但是我们估计的多个群体的基因组都显著大于739 Mb, 因此我们对红砂基因组大小的估计更为客观。当然, 红砂DNA C-值的最终确定还需要选取与其更加相近的物种进一步验证。
DNA C-值在被子植物种间差异极大, 最高可达2,400倍, 分析这些差异对于理解植物的生态适应性、生长策略、群落组成和物种进化等具有重要意义(Guignard et al, 2016; Pellicer et al, 2018; Guignard et al, 2019; Qiu et al, 2019)。例如, Qiu等(2019)发现在向日葵属内, 一年生二倍体物种具有更小的基因组和更快的进化速率, 其基因组大小的演化可能与生长季节的长短有关。本文研究的红砂隶属于柽柳科, 该科植物起源于特提斯海沿岸地区, 目前间断分布于亚洲、欧洲和北非的草原和荒漠地区, 对于研究荒漠植物的演化及耐盐、耐旱机理具有重要价值(张道远等, 2003; Zhang et al, 2014a, b)。然而, 植物DNA C-值数据库(Pellicer & Leitch, 2020)中仅收录了3个柽柳科物种的信息, 分别为Tamarix canariensis (DNA 1C-值为1.53 pg) (Garcia et al, 2010)、T. tetrandra (1.55 pg) (Zonneveld et al, 2005)和T. africana (1.65 pg) (Loureiro et al, 2007), 这些1C-值约是二倍体红砂的1.3倍, 远小于五柱红砂的1C-值。相较于柽柳科已知的约100种植物, 该科物种DNA C-值的研究非常少, 并且其基因组学研究也严重滞后。因此, 测定更多柽柳科物种的DNA C-值十分必要, 这将有助于加快对该科植物系统发育关系、抗旱机理和基因组学等方面的研究。
3.2 北疆红砂的复杂起源
同倍体和异源多倍化杂交物种形成的过程和条件明显不同(Mallet, 2007; 王玉国, 2017), 因此鉴定杂交物种与其亲本的倍性是深入解析杂交物种形成机制的必要前提。Shi等(2020)利用9个随机核基因片段模拟了红砂的群体动态历史, 结果表明北疆支系起源于两次杂交事件: 在古尔班通古特沙漠区域, 东西支系第一次杂交产生了ghost群体(统计分析推断出的未获得真实数据的群体), ghost群体再与东部支系杂交产生北疆红砂, 目前北疆支系处于物种形成的早期阶段。通过核型分析, Shi等(2020)发现来自北疆FK群体及东西支系的共3个个体都是二倍体; 虽然STRUCTURE分析结果显示HSS群体的遗传组分不同于FK等其他3个北疆群体, 但在分析群体动态历史时, Shi等将北疆支系当做一个整体, 并基于核型数据推测北疆支系都是二倍体且没有在杂交过程中发生染色体加倍。本研究中, 流式测定结果显示北疆支系中的FK (1.172 ± 0.013 pg)和SW群体(1.185 ± 0.009 pg)的DNA 1C-值介于东部支系(1.149 ± 0.012 pg)和西部支系(1.195 ± 0.031 pg) 1C-值之间, 而HSS和WCC群体的DNA 1C-值约等于东西支系1C-值的2倍。结合多个叶绿体基因片段(cpDNA)和核基因的遗传信息(Yin et al, 2015; Shi et al, 2020), 我们推测FK和SW群体属于同倍体杂交种群, 其倍性与东西支系相同, 都为二倍体; 而HSS和WCC群体属于异源四倍体杂交种群。此外, HSS群体的母本为西部支系, 但是FK和SW群体的母本为东部支系(Yin et al, 2015), 说明HSS群体并不是同倍体杂交种群(FK和SW)通过染色体加倍形成的。
综上所述, 本研究支持北疆红砂起源于多次杂交事件, 但该区域内不同倍性群体不属于单系类群。Fan等(未发表数据)基于简化基因组测序数据对红砂的群体遗传结构和种群动态历史进行了全面分析, 发现北疆红砂进一步分成两组, 其中一组(命名为GuD-S)的代表群体是FK和SW群体, 另一组(GuD-N)的代表群体正是HSS和WCC群体; 最优进化模型显示, 东西支系第一次杂交产生了GuD-S组(也就是Shi等研究中未找到的ghost群体), GuD-S组与东部支系进一步杂交产生GuD-N组。Shi等(2020)调查的北疆群体较少且集中于北疆的东南缘, 在分析种群动态历史时未考虑北疆支系内部的分化, 这应该是该研究得到的物种进化模型中存在ghost群体的主要原因。然而, 在GuD-N组中, 只有HSS和WCC群体的cpDNA单倍型来自西部支系(Fan et al, 未发表数据), 因此GuD-N真实的演化过程可能比模型显示的更加复杂。HSS和WCC群体的多倍化起源可能与其所处的环境更加恶劣有关: 该地区为砾石戈壁区, 附近有大型露天煤矿, 并且几乎无地表水流(侯艳军, 2015)。倍性差异会使四倍体种群与二倍体种群之间产生较强的生殖隔离(Mallet, 2007), 这是导致北疆红砂群体在无明显地理隔离的情况下仍然保持各自遗传特性的原因之一。两个物种可以通过多次自然杂交事件形成一个或多个新的同倍体杂交物种, 这已在少数植物物种形成研究中得到证实, 如向日葵属(Rieseberg, 1991; Schwarzbach & Rieseberg, 2002)和高山松(Ma et al, 2006)等; 但是在相同或者相邻区域内, 两个亲本分别杂交形成同倍体杂交物种和异源多倍化杂交物种的案例较为罕见。因此, 北疆红砂的复杂起源为研究同倍体杂交物种形成机制提供了新的模型, 可以通过比较两类杂交物种形成方式之间的异同进而揭示物种形成的遗传基础, 如适应性性状的产生与维持、自然选择与生殖隔离的关系等。为了更准确地利用这一材料, 下一步拟全面调查北疆红砂群体的倍性。
3.3 红砂不同支系的分类地位
红砂的分类地位曾经存在极大争议, 有些学者认为红砂应单独成属(琵琶柴属, Hololachna) (Gaskin et al, 2004); 而有些学者将琵琶柴属处理为红砂属的亚属, 红砂是该亚属的唯一物种(郝晓莉等, 2014); 还有一些学者认为红砂在分类系统上并不特殊, 只是红砂属内的一个物种(Zhang et al, 2014a)。大多数有关红砂分类学的研究都是以植物标本的形态性状差异为分类依据, 而几乎没有统计群体水平的性状差异(尤其是数量性状), 因此参考的分类依据可能受到采集样品偏差的影响, 存在不确定性; 而Zhang等(2014a)根据18个红砂属物种(共21种)的ITS和3个cpDNA片段的遗传信息构建了所含红砂属物种最全的系统发育树, 其中红砂、五柱红砂和R. fruticosa聚为一支, 证明红砂不应归于单独的亚属。
通过多个cpDNA片段和低拷贝核基因序列变异分析, 红砂还应归于一个独立的分类群, 但是种内不同群体可显著分为东部支系、西部支系和北疆支系(Yin et al, 2015; Shi et al, 2020), 民丰群体并未单独分化出来, 所以不建议将民丰红砂作为变种或种处理(崔大方等, 1999, 2000)。红砂的东西支系分布于不同的地理区域, cpDNA和核基因片段均支持它们各为单系类群; 北疆群体不仅占据了一个新的地理分布区和生态位, 而且在遗传组分上与其亲本存在明显的差异。Fan等(2020)发现东西支系的花期几乎不重叠, 北疆群体的花期物候处于亲本的花期之间, 表明三者之间已经出现了一定程度的合子前生殖隔离; 北疆群体中与生产力相关的叶长和叶宽更是显著大于亲本支系, 尤其是WCC群体具有最长的叶片。结合本文红砂群体间的基因组大小和倍性变化, 显然按照3个支系处理红砂不合理, 不利于对不同地区红砂抗旱机制等方面的进一步研究。
目前, 业界越来越认可“整合物种概念”, 这个概念强调物种的界定和分类应该考虑物种分化的多个方面: 稳定的形态学差异、可靠的分子证据、明显的生态位分化和一定程度的生殖隔离等; 由于物种始终处于分化路上, 处于分化早期阶段的物种之间可能只存在上述一两种区分特征, 而处于分化后期的物种则能表现出更多的特征(刘建全, 2016; 莫日根高娃等, 2019)。基于上述讨论, 我们认为红砂东西支系和北疆不同倍性杂交种群的分化和杂交历史清晰, 分布区及生态位相互不重叠, 叶片表型和花期物候也存在显著分化, 不同支系均符合“整合物种概念”的多个要素。虽然目前对红砂不同支系形态学上的描述和比较还很欠缺, 人工杂交实验和后代育性检测还未完成, 北疆两个杂种的分布区未全面调查, 但已有的证据表明这些支系可能是处于分化早中期阶段的年轻物种, 而不是难以区分的种复合体。因此本文建议将红砂不同地理种群按照4个物种处理: (1)主要分布于河西走廊、柴达木盆地、蒙古高原、阿拉善高原和噶顺戈壁的红砂东部支系为一个种; (2)分布于塔里木盆地的西部支系为一个种; (3)分布于天山以北准噶尔盆地南缘的红砂种群为二倍体杂交种; (4)分布于准噶尔盆地东部的红砂种群为异源四倍体杂交种。
附录 Supplementary Material
附录1 红砂不同支系及其生境
Appendix 1 Different genetic lineages of Reaumuria soongarica and their habitats
参考文献
Hybridization and speciation
DOI:10.1111/j.1420-9101.2012.02599.x
PMID:23323997
[本文引用: 1]

Hybridization has many and varied impacts on the process of speciation. Hybridization may slow or reverse differentiation by allowing gene flow and recombination. It may accelerate speciation via adaptive introgression or cause near-instantaneous speciation by allopolyploidization. It may have multiple effects at different stages and in different spatial contexts within a single speciation event. We offer a perspective on the context and evolutionary significance of hybridization during speciation, highlighting issues of current interest and debate. In secondary contact zones, it is uncertain if barriers to gene flow will be strengthened or broken down due to recombination and gene flow. Theory and empirical evidence suggest the latter is more likely, except within and around strongly selected genomic regions. Hybridization may contribute to speciation through the formation of new hybrid taxa, whereas introgression of a few loci may promote adaptive divergence and so facilitate speciation. Gene regulatory networks, epigenetic effects and the evolution of selfish genetic material in the genome suggest that the Dobzhansky-Muller model of hybrid incompatibilities requires a broader interpretation. Finally, although the incidence of reinforcement remains uncertain, this and other interactions in areas of sympatry may have knock-on effects on speciation both within and outside regions of hybridization.© 2013 The Authors. Journal of Evolutionary Biology © 2013 European Society For Evolutionary Biology.
The role of hybridization in evolution
Hybridization may influence evolution in a variety of ways. If hybrids are less fit, the geographical range of ecologically divergent populations may be limited, and prezygotic reproductive isolation may be reinforced. If some hybrid genotypes are fitter than one or both parents, at least in some environments, then hybridization could make a positive contribution. Single alleles that are at an advantage in the alternative environment and genetic background will introgress readily, although such introgression may be hard to detect. 'Hybrid speciation', in which fit combinations of alleles are established, is more problematic; its likelihood depends on how divergent populations meet, and on the structure of epistasis. These issues are illustrated using Fisher's model of stabilizing selection on multiple traits, under which reproductive isolation evolves as a side-effect of adaptation in allopatry. This confirms a priori arguments that while recombinant hybrids are less fit on average, some gene combinations may be fitter than the parents, even in the parental environment. Fisher's model does predict heterosis in diploid F1s, asymmetric incompatibility in reciprocal backcrosses, and (when dominance is included) Haldane's Rule. However, heterosis arises only when traits are additive, whereas the latter two patterns require dominance. Moreover, because adaptation is via substitutions of small effect, Fisher's model does not generate the strong effects of single chromosome regions often observed in species crosses.
The likelihood of homoploid hybrid speciation
DOI:10.1046/j.1365-2540.2000.00680.x URL [本文引用: 1]
Substantial intraspecific genome size variation in golden-brown algae and its phenotypic consequences
DOI:10.1093/aob/mcaa133 URL [本文引用: 1]
Determination of flavonoid compounds of Reaumuria L. (Tamaricaceae) and their taxonomical significance
琵琶柴属植物黄酮类化合物的测定及分类学意义
A new species of Reaumuria L. from Xinjiang
新疆琵琶柴属一新种
Plant DNA flow cytometry and estimation of nuclear genome size
DOI:10.1093/aob/mci005 URL [本文引用: 1]
Nuclear DNA content and genome size of trout and human
Estimation of nuclear DNA content in plants using flow cytometry
DOI:10.1038/nprot.2007.310 URL [本文引用: 4]
Leaf size variations in a dominant desert shrub, Reaumuria soongarica, adapted to heterogeneous environments
DOI:10.1002/ece3.v10.18 URL [本文引用: 4]
First genome size estimations for some eudicot families and genera
DOI:10.3989/collectbot.2010.v29 URL [本文引用: 1]
A systematic overview of Frankeniaceae and Tamaricaceae from nuclear rDNA and plastid sequence data
The origin, evolution and proposed stabilization of the terms ‘genome size' and ‘C-value' to describe nuclear DNA contents
Perusing the literature on nuclear 'genome size' shows that the term is not stabilized, but applied with different meanings. It is used for the DNA content of the complete chromosome complement (with chromosome number n), for which others use 'C-value', but also for the DNA content of the monoploid chromosome set only (with chromosome number x). Reconsideration of the terminology is required.Our purpose is to discuss the currently unstable usage of the terms 'genome size' and 'C-value', and to propose a new unified terminology which can describe nuclear DNA contents with ease and without ambiguity.We argue that there is a need to maintain the term genome size in a broad sense as a covering term, because it is widely understood, short and phonetically pleasing. Proposals are made for a unified and consensual terminology. In this, 'genome size' should mean the DNA content based on chromosome number x and n, and should be used mainly in a general sense. The necessary distinction of the kinds of genome sizes is made by the adjectives 'monoploid' and the neology 'holoploid'. 'Holoploid genome size' is a shortcut for the DNA content of the whole chromosome complement characteristic for the individual (and by generalization for the population, species, etc.) irrespective of the degree of generative polyploidy, aneuploidies, etc. This term was lacking in the terminology and is for reasons of linguistic consistency indispensable. The abbreviated terms for monoploid and holoploid genome size are, respectively, Cx-value and C-value. Quantitative data on genome size should always indicate the C-level by a numerical prefix, such as 1C, 1Cx, 2C, etc. The proposed conventions cover general fundamental aspects relating to genome size in plants and animals, but do not treat in detail cytogenetic particularities (e.g. haploids, hybrids, etc.) which will need minor extensions of the present scheme in a future paper.
The ecological genetics of homoploid hybrid speciation
Our understanding of homoploid hybrid speciation has advanced substantially since this mechanism of species formation was codified 50 years ago. Early theory and research focused almost exclusively on the importance of chromosomal rearrangements, but it later became evident that natural selection, specifically ecological selection, might play a major role as well. In light of this recent shift, we present an evaluation of ecology's role in homoploid hybrid speciation, with an emphasis on the genetics underlying ecological components of the speciation process. We briefly review new theoretical developments related to the ecology of homoploid hybrid speciation; propose a set of explicit, testable questions that must be answered to verify the role of ecological selection in homoploid hybrid speciation; discuss published work with reference to these questions; and also report new data supporting the importance of ecological selection in the origin of the homoploid hybrid sunflower species Helianthus deserticola. Overall, theory and empirical evidence gathered to date suggest that ecological selection is a major factor promoting homoploid hybrid speciation, with the strongest evidence coming from genetic studies.
Genome size and ploidy influence angiosperm species' biomass under nitrogen and phosphorus limitation
DOI:10.1111/nph.13881
PMID:26875784
[本文引用: 2]
Angiosperm genome sizes (GS) range c. 2400-fold, and as nucleic acids are amongst the most phosphorus- (P) and nitrogen (N)-demanding cellular biomolecules, we test the hypothesis that a key influence on plant biomass and species composition is the interaction between N and P availability and plant GS. We analysed the impact of different nutrient regimes on above-ground biomass of angiosperm species with different GS, ploidy level and Grime's C-S-R (competitive, stress-tolerant, ruderal) plant strategies growing at the Park Grass Experiment (Rothamsted, UK), established in 1856. The biomass-weighted mean GS of species growing on plots with the addition of both N and P fertilizer were significantly higher than that of plants growing on control plots and plots with either N or P. The plants on these N + P plots are dominated by polyploids with large GS and a competitive plant strategy. The results are consistent with our hypothesis that large genomes are costly to build and maintain under N and P limitation. Hence GS and ploidy are significant traits affecting biomass growth under different nutrient regimes, influencing plant community composition and ecosystem dynamics. We propose that GS is a critical factor needed in models that bridge the knowledge gap between biodiversity and ecosystem functioning.© 2016 The Authors. New Phytologist © 2016 New Phytologist Trust.
Interspecific and intraspecific variations of plant DNA C-values and its biological significance
植物DNA C-值在种间和种内的变异及其生物学意义
Classification and distribution of the genus Reaumuria L. in Tamaricaceae
柽柳科红砂属(Reaumuria L.)的分类与分布
Genome size variation in gymnosperms under different growth conditions
DOI:10.1080/00087114.2015.1024546 URL [本文引用: 1]
Study on the Transport Processes and Ecological Effect of Heavy Metals in Dust-soil-plant in the East Junggar Region
PhD dissertation, Xinjiang University, Urumqi.
准东地区降尘-土壤-植物重金属迁移过程及生态效应研究
博士学位论文, 新疆大学, 乌鲁木齐.]
Constraints on polyploid evolution:A test of the minority cytotype exclusion principle
Response of a desert shrub to past geological and climatic change: A phylogeographic study of Reaumuria soongarica (Tamaricaceae) in western China
DOI:10.1111/j.1759-6831.2012.00201.x URL [本文引用: 1]
Estimation of genomic C value of 25 samples of Iris plants by flow cytometry
25份鸢尾属植物基因组DNA C值的流式测定
“The integrative species concept” and “species on the speciation way”
DOI:10.17520/biods.2016222 URL [本文引用: 1]
“整合物种概念”和“分化路上的物种”
The typical extreme xerophyte Reaumuria soongorica in the desert of China
我国荒漠典型超旱生植物-红砂
A study on origin and formation of the Chinese desert floras
试论我国沙漠地区植物区系的发生与形成
Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species
After the initial boom in the application of flow cytometry in plant sciences in the late 1980s and early 1990s, which was accompanied by development of many nuclear isolation buffers, only a few efforts were made to develop new buffer formulas. In this work, recent data on the performance of nuclear isolation buffers are utilized in order to develop new buffers, general purpose buffer (GPB) and woody plant buffer (WPB), for plant DNA flow cytometry.GPB and WPB were used to prepare samples for flow cytometric analysis of nuclear DNA content in a set of 37 plant species that included herbaceous and woody taxa with leaf tissues differing in structure and chemical composition. The following parameters of isolated nuclei were assessed: forward and side light scatter, propidium iodide fluorescence, coefficient of variation of DNA peaks, quantity of debris background, and the number of particles released from sample tissue. The nuclear genome size of 30 selected species was also estimated using the buffer that performed better for a given species.In unproblematic species, the use of both buffers resulted in high quality samples. The analysis of samples obtained with GPB usually resulted in histograms of DNA content with higher or similar resolution than those prepared with the WPB. In more recalcitrant tissues, such as those from woody plants, WPB performed better and GPB failed to provide acceptable results in some cases. Improved resolution of DNA content histograms in comparison with previously published buffers was achieved in most of the species analysed.WPB is a reliable buffer which is also suitable for the analysis of problematic tissues/species. Although GPB failed with some plant species, it provided high-quality DNA histograms in species from which nuclear suspensions are easy to prepare. The results indicate that even with a broad range of species, either GPB or WPB is suitable for preparation of high-quality suspensions of intact nuclei suitable for DNA flow cytometry.
The use of flow cytometry in the biosystematics, ecology and population biology of homoploid plants
Correlations between foliar stable carbon isotope composition and environmental factors in desert plant Reaumuria soongorica (Pall.) Maxim
DOI:10.1111/jipb.2005.47.issue-9 URL [本文引用: 1]
Genetic structure and evolutionary history of a diploid hybrid pine Pinus densata inferred from the nucleotide variation at seven gene loci
DOI:10.1093/molbev/msj100 URL [本文引用: 2]
Genome size discriminates between closely related taxa Elytrigia repens and E. intermedia (Poaceae: Triticeae) and their hybrid
DOI:10.1007/BF02804286 URL [本文引用: 2]
Hybridization as an invasion of the genome
DOI:10.1016/j.tree.2005.02.010 URL [本文引用: 1]
One or more species? GBS sequencing and morphological traits evidence reveal species diversification of Sphaeropteris brunoniana in China
DOI:10.17520/biods.2019146
[本文引用: 1]
Species are not static, but in the process of continuous differentiation and evolution. In this context, reproductive isolation is considered the key to most speciation events. However, species that may be only partially reproductively isolated may disturb our understanding of a species, as stated by ‘The integrative species concept’. In the Flora of China, Chinese Sphaeropteris contains two species, S. brunoniana and S. lepifera. S. brunoniana is thought to be the same species as a Hainan native, S. hainanensis, however, S. hainanensis seems to have differentiated morphologically. This study further explores the genetic relationship between S. brunoniana and its related species by collecting 21 samples from 9 populations for GBS reduced-representation genome sequencing for phylogenetic analysis. Using both genetic and morphological data, our results indicate that S. hainanensis differs genetically from S. brunoniana, but is also morphologically distinct based on leaf characteristics and spore ornamentation. Although the reproductive isolation of the two populations is weak, hybrids are formed in the coastal areas of Guangxi and leaves mimic intermediate morphology between parents. Therefore, our results suggest that S. hainanensis is in the process of speciation due to geographic isolation and suggests to restore its species status. S. brunoniana × hainanensis which the sample from Guangxi should be considered an independent natural hybrid taxon.
一个种还是多个种? 简化基因组及其形态学证据揭示中国白桫椤植物的物种多样性分化
DOI:10.17520/biods.2019146
[本文引用: 1]
物种是生物多样性的基本单元, 生殖隔离被认为是物种形成的关键; 然而物种并不是静止的而是处于不断的分化演变之中, 已经稳定成型但尚未到达分化后期的物种可能存在不完全的生殖隔离。对于物种的认识不能单从某一侧面或局部特征来界定, 而应通过“整合物种概念”来确定物种地位。Flora of China记载了中国产白桫椤属(Sphaeropteris) 2种, 即白桫椤(S. brunoniana)和笔筒树(S. lepifera), 并认为原产中国海南的海南白桫椤(S. hainanensis)和白桫椤为同一物种而将其并入白桫椤; 但海南白桫椤在形态上已出现了分化。为探讨白桫椤及其近缘物种的亲缘关系和物种多样性分化的情况, 本文采集到9个居群共21个样本, 通过GBS简化基因组测序技术获得单核苷酸变异位点(SNP), 进行系统发育树的构建和主成分及遗传结构的分析, 并结合叶片数量性状的统计分析和孢子形态的观察测量。结果表明, 海南白桫椤不仅与云南产白桫椤的基因型不同, 且在叶片特征和孢子纹饰上有明显差异; 但两个居群的生殖隔离较弱, 在广西沿海地区形成杂交产物, 其叶片特征为亲本的中间类型。因此, 我们认为海南白桫椤是由于地理隔离而形成的一个处在分化路上的物种, 建议恢复其物种地位; 广西产白桫椤为自然杂交群体, 应另处理为独立的自然杂交分类群——广西白桫椤(S. brunoniana × hainanensis)。
Genome size diversity and its impact on the evolution of land plants
DOI:10.3390/genes9020088 URL [本文引用: 2]
The Plant DNA C-values database (release 7.1): An updated online repository of plant genome size data for comparative studies
Ecological genetics of Reaumuria soongorica (Pall.) Maxim. population in the oasis-desert ecotone in Fukang, Xinjiang, and its implications for molecular evolution
DOI:10.1016/j.bse.2008.01.008 URL [本文引用: 1]
Phylogenetic trends and environmental correlates of nuclear genome size variation in Helianthus sunflowers
DOI:10.1111/nph.2019.221.issue-3 URL [本文引用: 2]
Homoploid reticulate evolution in Helianthus (Asteraceae): Evidence from ribosomal genes
DOI:10.1002/ajb2.1991.78.issue-9 URL [本文引用: 2]
Major ecological transitions in wild sunflowers facilitated by hybridization
Hybridization is frequent in many organismal groups, but its role in adaptation is poorly understood. In sunflowers, species found in the most extreme habitats are ancient hybrids, and new gene combinations generated by hybridization are speculated to have contributed to ecological divergence. This possibility was tested through phenotypic and genomic comparisons of ancient and synthetic hybrids. Most trait differences in ancient hybrids could be recreated by complementary gene action in synthetic hybrids and were favored by selection. The same combinations of parental chromosomal segments required to generate extreme phenotypes in synthetic hybrids also occurred in ancient hybrids. Thus, hybridization facilitated ecological divergence in sunflowers.
Meiotic behaviour, karyotype analyses and pollen viability in species of Tamarix (Tamaricaceae)
DOI:10.3372/wi.43.43121 URL [本文引用: 1]
Likely multiple origins of a diploid hybrid sunflower species
The recurrent origin of diploid hybrid species is theoretically improbable because of the enormous diversity of hybrid genotypes generated by recombination. Recent greenhouse experiments, however, indicate that the genomic composition of hybrid lineages is shaped in part by deterministic forces, and that recurrent diploid hybrid speciation may be more feasible than previously believed. Here we use patterns of variation from chloroplast DNA (cpDNA), nuclear microsatellite loci, cross-viability and chromosome structure to assess whether a well-characterized diploid hybrid sunflower species, Helianthus anomalus, was derived on multiple occasions from its parental species, H. annuus and H. petiolaris. Chloroplast DNA and crossability data were most consistent with a scenario in which H. anomalus arose three times: three different H. anomalus fertility groups were discovered, each with a unique cpDNA haplotype. In contrast, there was no clear signature of multiple, independent origins from the microsatellite loci. Given the age of H. anomalus (> 100 000 years bp), it may be that microsatellite evidence for recurrent speciation has been eroded by mutation and gene flow through pollen.
Effects of different isolation buffers on DNA resolution in nucleus suspension of root tip cell of Cunninghamia lanceolata (Lamb.) Hook
不同分离缓冲液对杉木根尖细胞核悬液DNA分辨率的影响
Seed germination characteristics and local adaptation of Reaumuria soongarica
红砂(Reaumuria soongarica)种子萌发特性及其局部适应性
Divergence and hybridization in the desert plant Reaumuria soongarica
DOI:10.1111/jse.v58.2 URL [本文引用: 11]
Transcriptomic analysis of a tertiary relict plant, extreme xerophyte Reaumuria soongorica to identify genes related to drought adaptation
DOI:10.1371/journal.pone.0063993 URL [本文引用: 1]
Understanding intraspecific variation in genome size in plants
Advances in the study of polyploidy since plant speciation
DOI:10.1046/j.1469-8137.2003.00948.x URL [本文引用: 1]
Reliable DNA ploidy determination in dehydrated tissues of vascular plants by DAPI flow cytometry-New prospects for plant research
DOI:10.1002/(ISSN)1552-4930 URL [本文引用: 1]
Genomic basis of homoploid hybrid speciation within chestnut trees
DOI:10.1038/s41467-020-17111-w URL [本文引用: 1]
Insights from genomes into the evolutionary importance and prevalence of hybridization in nature
The tomato genome sequence provides insights into fleshy fruit evolution
DOI:10.1038/nature11119 URL [本文引用: 1]
Evidence of widespread hybridization among couch grasses (Elymus, Poaceae)
DOI:10.1111/jse.v59.1 URL [本文引用: 1]
Polyploidy: An evolutionary and ecological force in stressful times
DOI:10.1093/plcell/koaa015 URL [本文引用: 1]
Ecological niche shift between diploid and tetraploid plants of Fragaria (Rosaceae) in China
DOI:10.1016/j.sajb.2018.10.027
[本文引用: 1]
Ecological niche shifts between diploid and tetraploid plants remains a topic with potential for plant ecology and evolution. Here, we identified populations of Fragaria (Rosaceae) from 169 sampling plots, classifying populations into two species groups (diploid and tetraploid) across China using flow cytometry. We then applied niche modeling to assess the ecological niche shift between diploid and tetraploid plants of Fragaria in China based on fieldwork and herbarium data. We explored the variables that best explained niche shifts between diploid and tetraploid plants. Compared to diploid plants, tetraploid plants are more likely to inhabit areas of higher altitude, lower temperature and lower precipitation, indicating that different ploidy levels are mainly associated with niche changes of altitude, temperature and precipitation. Diploid and tetraploid plants of Fragaria exhibit significant niche shifts (i.e. niche divergence between types). Ecological niche divergence between diploid and tetraploid plants is best explained by a shift in climate and altitude in biomes of conifer forests of southwestern China. Moreover, heterogeneity of soil further contributed to the observed niche shift between diploid and tetraploid plants. Our study provides novel insights into the role of niche shifts on genome mergers and duplications of Fragaria in China. (C) 2018 SAAB. Published by Elsevier B.V.
The chromosome number, karyotype and genome size of the desert plant diploid Reaumuria soongorica (Pall.) Maxim
DOI:10.1007/s00299-011-1020-3 URL [本文引用: 8]
Operation skills of flow cytometer for detecting nuclear DNA contents in higher plant cells
流式细胞仪检测高等植物细胞核DNA含量的方法
Natural hybridization and speciation
DOI:10.17520/biods.2017041 URL [本文引用: 3]
自然杂交与物种形成
DOI:10.17520/biods.2017041
[本文引用: 3]
在生物进化过程中, 自然杂交对物种形成有着重要的影响。多倍化和同倍体杂交物种形成是通过杂交产生新种的两种主要方式。普遍的观点认为多倍化是植物物种形成的主要动力, 而越来越多染色体倍性相同物种之间杂交现象的发现, 说明同倍体杂交也是物种形成的重要方式。目前按严格标准确定的同倍体物种形成案例相对较少, 还亟需对更多不同类型的种间杂交带进行形态与分子等多方面证据的调研, 并通过模型预测与实测数据分析, 来探讨杂交物种形成初期乃至整个过程的遗传基础; 更多杂交物种形成的新模式材料, 有待于通过分化谱系的历史基因流重建进行鉴定, 用于进一步阐释自然选择对于生殖隔离形成的影响、物种形成过程中的生态适应变化和多样性遗传创新的产生规律。本文简要综述了自然杂交的研究历史, 阐述其相关概念、研究方法和基因组时代杂交与物种形成研究的最近进展, 并针对拟解决的关键问题提出相应的分析见解, 以期为后续研究和涉及杂交的物种多样性保护提供借鉴。
Hybrid speciation via inheritance of alternate alleles of parental isolating genes
DOI:10.1016/j.molp.2020.11.008 URL [本文引用: 1]
The role of homoploid hybridization in evolution: A century of studies synthesizing genetics and ecology
DOI:10.3732/ajb.1400201
PMID:25156978
[本文引用: 1]
While homoploid hybridization was viewed as maladaptive by zoologists, the possibility that it might play a creative role in evolution was explored and debated by botanists during the evolutionary synthesis. Owing to his synthetic work on the ecological and genetic factors influencing the occurrence and effects of hybridization, G. Ledyard Stebbins' contributions to this debate were particularly influential. We revisit Stebbins' views on the frequency of hybridization, the evolution of hybrid sterility, and the evolutionary importance of transgressive segregation, introgression, and homoploid hybrid speciation in the context of contemporary evidence. Floristic surveys indicate that ∼10% of plant species hybridize, suggesting that natural hybridization is not as ubiquitous as Stebbins argued. There is stronger support for his contention that chromosomal sterility is of greater importance in plants than in animals and that selection drives the evolution of hybrid sterility. Stebbins' assertions concerning the frequent occurrence of transgressive segregation and introgressive hybridization have been confirmed by contemporary work, but few studies directly link these phenomena to adaptive evolution or speciation. Stebbins proposed a mechanism by which chromosomal rearrangements partially isolate hybrid lineages and parental species, which spurred the development of the recombinational model of homoploid speciation. While this model has been confirmed empirically, the establishment of reproductively independent hybrid lineages is typically associated with the development of both intrinsic and extrinsic reproductive barriers. We conclude by reflecting on outcomes of hybridization not considered by Stebbins and on possible future research that may extend our understanding of the evolutionary role of hybridization beyond Stebbins' legacy. © 2014 Botanical Society of America, Inc.
The effects of different growth regulators on the Reaumuria soongorica (Pall.) Maxim seedling emergence and growth
植物生长调节剂对红砂出苗及苗期生长的影响
Genetic diversity and drivers of genetic differentiation of Reaumuria soongorica of the Inner Mongolia Plateau in China
DOI:10.1007/s11258-015-0479-3 URL [本文引用: 1]
The role of East Asian monsoon system in shaping population divergence and dynamics of a constructive desert shrub Reaumuria soongarica
DOI:10.1038/srep15823 URL [本文引用: 7]
Optimization of strawberry ploidy identification method using flow cytometry
利用流式细胞仪检测草莓倍性方法的优化
The photogeographical studies of Tamarix (Tamaricaceae)
柽柳科柽柳属的植物地理研究
On the general characteristics of plant diversity of Gurbantunggut sandy desert
论古尔班通古特沙漠植物多样性的一般特点
Spatiotemporal evolution of Reaumuria (Tamaricaceae) in Central Asia: Insights from molecular biogeography
DOI:10.11646/phytotaxa.167.1 URL [本文引用: 4]
Himalayan origin and evolution of Myricaria (Tamaricaeae) in the Neogene
DOI:10.1371/journal.pone.0097582 URL
Seed morphology of Tamaricaceae in China arid areas and its systematic evolution
中国干旱区柽柳科植物种子形态特征及其系统学意义
Weak crossability barrier but strong juvenile selection supports ecological speciation of the hybrid pine Pinus densata on the Tibetan Plateau
DOI:10.1111/evo.12496
PMID:25065387
[本文引用: 2]
Determining how a new hybrid lineage can achieve reproductive isolation is a key to understanding the process and mechanisms of homoploid hybrid speciation. Here, we evaluated the degree and nature of reproductive isolation between the ecologically successful hybrid species Pinus densata and its parental species P. tabuliformis and P. yunnanensis. We performed interspecific crosses among the three species to assess their crossability. We then conducted reciprocal transplantation experiments to evaluate their fitness differentiation, and to examine how natural populations representing different directions of introgression differ in adaptation. The crossing experiments revealed weak genetic barriers among the species. The transplantation trials showed manifest evidence of local adaptation as the three species all performed best in their native habitats. Pinus densata populations from the western edge of its distribution have evolved a strong local adaptation to the specific habitat in that range; populations representing different directions of introgressants with the two parental species all showed fitness disadvantages in this P. densata habitat. These observations illustrate that premating isolation through selection against immigrants from other habitat types or postzygotic isolation through selection against backcrosses between the three species is strong. Thus, ecological selection in combination with endogenous components and geographic isolation has likely played a significant role in the speciation of P. densata. © 2014 The Author(s). Evolution © 2014 The Society for the Study of Evolution.
Genome sizes of Eucomis L'Hér. (Hyacinthaceae) and a description of the new species Eucomis grimshawii G. D. Duncan & Zonneveld
DOI:10.1007/s00606-009-0236-y URL [本文引用: 1]
First nuclear DNA amounts in more than 300 angiosperms
Genome size (DNA C-value) data are key biodiversity characters of fundamental significance used in a wide variety of biological fields. Since 1976, Bennett and colleagues have made scattered published and unpublished genome size data more widely accessible by assembling them into user-friendly compilations. Initially these were published as hard copy lists, but since 1997 they have also been made available electronically (see the Plant DNA C-values database http://www.kew.org/cval/homepage.html). Nevertheless, at the Second Plant Genome Size Meeting in 2003, Bennett noted that as many as 1000 DNA C-value estimates were still unpublished and hence unavailable. Scientists were strongly encouraged to communicate such unpublished data. The present work combines the databasing experience of the Kew-based authors with the unpublished C-values produced by Zonneveld to make a large body of valuable genome size data available to the scientific community.C-values for angiosperm species, selected primarily for their horticultural interest, were estimated by flow cytometry using the fluorochrome propidium iodide. The data were compiled into a table whose form is similar to previously published lists of DNA amounts by Bennett and colleagues.The present work contains C-values for 411 taxa including first values for 308 species not listed previously by Bennett and colleagues. Based on a recent estimate of the global published output of angiosperm DNA C-value data (i.e. 200 first C-value estimates per annum) the present work equals 1.5 years of average global published output; and constitutes over 12 % of the latest 5-year global target set by the Second Plant Genome Size Workshop (see http://www.kew.org/cval/workshopreport.html). Hopefully, the present example will encourage others to unveil further valuable data which otherwise may lie forever unpublished and unavailable for comparative analyses.
Establishment of flow cytometry system for DNA C-value determination of polyploid species in subgenus Yulania
玉兰亚属多倍体植物DNA C-值流式细胞术测定体系的建立

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}