


群落结构在森林生态系统中具有重要作用, 一直是森林生态学的研究热点。森林群落的结构特征不仅表现在水平方向, 也表现在垂直方向上(孙儒泳等, 2007)。森林群落垂直结构的主要表现形式是林冠分层, 这是群落的基本特征之一(赵志模和郭依泉, 1990)。常用树冠层次(即林层)来描述和分析森林垂直结构(庄崇洋等, 2014)。关于林层的研究早在19世纪初就开始了, 但其发展历程中一直存在诸多争议, 一方面是因为林层分层的定义不明且划分方法多样, 另一方面则是因为缺乏对林冠层的有效接近手段, 难以直观准确地获取其特征(Neto, 1989)。关于分层的定义, 不同学者给出了不同标准, 一般从叶片生物量、个体和物种3个方面(Grubb et al, 1963)来定义。在分层方法上包括定性和定量两大类。定性方法主要是通过研究者的主观意识根据森林群落外貌特征来进行划分, 常用的如Watt (1924)提出的剖面图法等。定量方法则避免了研究者的主观性, 主要根据林内物种的描述性特征量如树高、胸径、枝下高、龄组、冠幅、叶片面积等参数的关系或变化来进行划分, 常用的如林冠分层指数(stratification index, SI)法(Ashton & Hall, 1992), TSTRAT分层算法(Latham et al, 1998)和遥感方法(Gillespie et al, 2004; 赵静等, 2013)等。正是由于垂直结构划分的困难性, 使得关于森林各林层林木分布格局和变化特征的研究仍不多见。
值得注意的是, 虽然定量方法更加客观, 但由于冠层接近技术的缺乏, 通常只能通过获取下层枝叶的样本资料来进行推断(李德志和臧润国, 2004); 或通过地面大致估计来获取物种描述性特征参数,导致对森林垂直结构的划分结果也较为粗略。直到20世纪70年代后期, Perry (1978)提出单绳攀爬技术(single rope technique)之后, 人们才初步找到了接近森林冠层的方法, 此后还有热气球、冠层步道、塔吊等技术手段, 极大地提高了观测数据的准确性, 同时还更大程度地保证了接近林冠时研究者的安全(Lowman & Wittman, 1996; Lowman & Rinker, 2004; 杨龙等, 2016; 沈浩等, 2017)。有研究表明, 林冠塔吊是研究森林群落冠层的有效手段, 具有高精度、非破坏性、可重复等特征, 已成为当前林冠研究的重要手段之一(吴毅等, 2016)。
对群落构建机制的理解与阐释一直是生态学的核心, 探究群落间物种多样性的空间变化格局是理解群落构建机制的关键所在。描述局域群落的物种多样性主要有α多样性和β多样性两个层次。α多样性是指群落或特定立地的物种丰富度; β多样性体现的是群落物种组成的差异, 即“与环境梯度或环境格局相关的群落组成变化的幅度或群落分化的程度” (Rosenzweig, 1995; 陈圣宾等, 2010; Legendre & De Cáceres, 2013)。β多样性常用来描述物种组成在时空尺度的变化。相异性指数(如Jaccard相异性指数和Sørensen相异性指数)是常用的β多样性度量方法之一(陈圣宾等, 2010)。
Harrison等(1992)、Williams (1996)和Lennon等(2001)提出群落间物种组成差异(β多样性)由两种不同的过程造成, 其一为物种周转或替换(species turnover, or replacement), 其二为物种丧失或增加(species loss or gain)。β多样性分解就是要区分这两种过程对总体β多样性的作用, 并探讨这两种过程如何共同影响不同时空尺度的物种分布格局(Baselga, 2010; Podani & Schmera, 2011; 斯幸峰等, 2017)。常用的β多样性分解方法有Podani和Schmera方法(POD法, Podani & Schmera, 2011)和Baselga方法(BAS法, Baselga, 2013)两种。POD法将两个群落间的差异分解为物种替换组分和丰富度差异组分两部分, 其中, 物种替换组分表示不同群落间由物种替代引起的差异部分; 而物种丧失会导致群落间物种丰富度产生差异, 即丰富度差异组分表示由物种丧失引起的部分(Lennon et al, 2001)。BAS法则将其分解为物种周转组分和嵌套组分两部分, 其中, 嵌套是丰富度差异的一种特殊情况, 当物种丰富度差异沿着某一梯度呈现出有序排列, 拥有较少物种的群落是拥有较多物种的群落的严格子集, 此时会呈现嵌套格局。β多样性分解方法被系统地提出之后, 该研究领域的成果迅速增加, 但主要集中在北美和欧洲, 我国对此的研究还较少(斯幸峰等, 2017)。另外, 研究者们对于β多样性的研究也不再仅仅停留在描述不同地点间的物种组成差异, 更试图揭示这些差异的形成机制。不同组分格局的形成机制有所差异, 多数研究表明, 导致物种周转的潜在机制包括环境过滤、竞争和地理隔离等作用(Angeler, 2013; Legendre, 2014)。而形成物种丰富度差异(或嵌套格局)可能的驱动机制包括生态位的多样性(diversity of niches)、选择性灭绝(selective extinction)、选择性迁入(selective colonization)和生境嵌套(habitat nestedness) (Ulrich et al, 2009)等生态过程。
因此, 探究β多样性及其分解格局将有助于揭示群落的构建机理。由于研究条件的限制, 已有研究多在水平结构上进行, 忽略了群落垂直结构的影响。由于林冠塔吊设备可以便利地获取准确的林层描述参数, 本文利用林冠塔吊测量和地面测量结合的方法对鼎湖山南亚热带常绿阔叶林林冠塔吊监测样地(简称: 塔吊样地)内所有DBH ≥ 1 cm的植物进行调查, 以获取准确树高, 再根据个体树高数据对塔吊样地进行分层, 以获得其垂直结构各层次的基本特征。并将每个林层的全部植物分别视为一个群落, 计算其α多样性指数; 将两个林层间物种组成的差异沿森林垂直层次上的变化幅度视为这两个群落间的β多样性。采用POD法进行β多样性的计算和分解, 分析β多样性的格局, 同时分析物种组成差异来源的两个组分(周转和丰富度差异)对塔吊样地林层间β多样性的作用。以期结合每个林层的物种组成特征和林内环境特征, 从垂直结构层面初步探究影响本样地群落构建的过程。
1 材料与方法
1.1 研究地概况
鼎湖山塔吊样地于2014年在广东省肇庆市鼎湖山国家级自然保护区(23°09'21"-23°11'30" N, 112°30'39"-112°33'41" E)内建立, 是中国森林冠层生物多样性监测专项网的一部分。塔吊塔身高度60 m、机械臂臂长60 m, 以塔吊为中心建立的样地东西长120 m, 南北长120 m, 面积为1.44 ha。所在区域属南亚热带湿润型季风气候, 冬夏季气候交替明显, 年均温和年降水量分别为20.9℃和1,927 mm, 年均蒸发量1,115 mm, 年均相对湿度80.3% (黄忠良等, 1998; 叶万辉等, 2008)。样地海拔高度48-100 m, 坡度4°-36°。土壤以发育于砂页岩母质的赤红壤为主, 地形复杂。植被类型为典型的南亚热带常绿阔叶林, 样地内共有木本植物(DBH ≥ 1 cm) 4,140株, 分属50科84属121种。
1.2 样地调查数据
按照美国Smithsonian热带研究所热带森林科学研究中心ForestGEO (Forest Global Earth Observatory, 原CTFS)森林动态样地建设技术规范(Condit, 1998), 利用全站仪将样地划分为36个20 m × 20 m的小样方, 调查样地内所有DBH ≥ 1 cm的木本植物个体, 记录种名、编号、胸径、坐标等。2014年10月完成第1次调查, 2018年11月进行了第2次调查。本次分析采用第2次的调查数据。
在第2次调查中胸径数据全部由胸径尺实测获得, 利用林冠塔吊和测高杆准确测量样地内每树的实际高度。具体方法为: 对于样地中从地面难以直接尺测的高树, 测量人员利用塔吊吊篮在森林林冠层之上用卷尺向下垂直测量, 记录该树树冠顶部到根部的高度, 即该树树高。对于剩下的高度较低的树木则利用测高杆地面手持直接测量。
1.3 群落内小气候特征
选取分布于群落内不同方位和不同树高的荷木(Schima superba)、马尾松(Pinus massoniana)、厚壳桂(Cryptocarya chinensis)和锥栗(Castanopsis chinensis) 4个物种, 每种各3棵共12个个体, 在每棵树上的5个不同位置(冠表层及之上两处、树冠最下方处、枝下高处和小于5 m树高下处)分别放置一套微气象环境监测系统(监测设备放置的具体高度见附录1), 包括: 空气温湿度传感器、雨量传感器、光合有效辐射传感器、无线数据采集器、数据采集扩展单元、太阳能供电系统等, 整个系统实现了对群落垂直方向上不同高度林层林内环境的实时立体监测。本次分析主要用到的微气象指标包括: 太阳辐射(光照, μE)、温度(℃)和相对空气湿度(RH%)。
将气象监测系统采集到的环境数据做如下处理: 选取样地8月监测数据为处理对象, 由于系统采集数据为每小时的实时数据, 因此对一天24 h的光照、温度和空气相对湿度进行日平均, 获得每日平均环境数据, 再获得月平均。
2 数据分析
2.1 群落垂直结构的划分
将第2次调查获得的样地内全部个体(DBH ≥ 1 cm)的准确树高进行K-means非监督聚类以获得垂直结构层次和各层树高范围。首先由于非监督分类需要根据经验或其他条件事先设定分类数, 所以要先确定树高的最佳分类数。用于确定最佳分类数的常用指标有calinski值、ssi (simple structure index)值,还可以根据不同分类情况下的组内平方和变化情况来判断。本文综合ssi指数和组内平方和变化情况这两个指标来确定最佳分层数。一般来说ssi值最大时对应的分类数为最佳分类组数(Dolnicar et al, 1999)。在不同分类组数的组内平方和变化折线图中, 随分类组数的增大, 当组内平方和的大小变化趋缓时, 对应的分类组数可为最佳分类组数。选取由这两个指标综合确定的最佳分类组数为最佳树高分层层次, 即塔吊样地垂直结构层数。最后将最佳分类组数代入K-means聚类法中, 输出得到每一层的树高范围, 根据树高大小将样地内各树(DBH ≥ 1 cm)划分到相应的层次中。
2.2 群落各林层的α多样性
在20 m × 20 m塔吊样地地面空间尺度上, 计算各林层的α多样性指数, 具体采用Shannon-Wiener多样性指数(H)和Pielou均匀度指数(J)(Pielou, 1975; 马克平和刘玉明, 1994)两个指数。为避免各层总多度差异较大的影响, 本文采用重要值为测度指标计算多样性指数(Whittaker, 1972)。其计算公式如下:
式中, Pi = Ni / N, 表示第i个物种的相对重要值, N为总体重要值, Ni为第i个种的重要值, S为物种数。另外, 采用Duncan法对各林层间α多样性指数大小进行多重比较, 判断差异是否显著。
2.3 群落垂直的β多样性及分解
因为从垂直结构来看, 鼎湖山各林层间物种组成随林层高度上升, 拥有较少物种的林层并非拥有较多物种林层的严格子集, 因此本文选择基于Sørensen相异性指数的POD法对鼎湖山森林各林层物种组成差异进行分解, 即分解为物种替换和丰富度差异组分两部分。具体方法为: 将塔吊样地中每一林层的全部植株视为一个群落, 并计算两两林层间的整体β多样性和两组分的大小(物种数据为每林层内DBH ≥ 1 cm的全部植物)。具体计算和分解公式如下:
Sørensen相异性指数:
物种替换组分:
丰富度差异组分(Richness difference):
考虑到本群落物种多度的不均匀性, 故本文中用于β多样性分解的数据为物种多度数据。A为两个林层共有物种的多度, B、C分别为两个林层各自物种的多度。
2.4 群落林层间的物种比较
本文利用Chazdon等(2010)提出的基于两个生境物种相对多度的多项式: CLAM (Classification Method)法分类模型, 将塔吊样地各林层的物种组成进行两两比较。两林层(林层A、林层B)的物种比较结果分为4个部分: 两林层共有物种(generalist)、林层A的特有种(A group specialist)、林层B的特有种(B group specialist)和相对多度太小无法分类的物种(too rare to classify)。其中因相对多度太小无法进行分类的物种是指在该模型中无统计学意义, 不能被划入前3种任何一类的物种的集合。
该模型主要影响参数有两个, 一个是决定阈值是否严格的K值, 可取2/3或1/2; 另一个是检验区分是否显著的P值, 可取值0.05、0.01、0.005或0.001。本文在进行分类时, 为将全部物种进行分类, 并达到统计学上的显著水平, K值取2/3, P值取0.005。此过程在CLAM程序(http://purl.oclc.org/clam)中完成。
本文数据计算和分析过程主要在R 3.4.3中完成, 其中聚类使用clust包, α多样性指数计算使用vegan包, β多样性及其分解使用betapart包(Baselga & Orme, 2012)。
3 结果
3.1 群落垂直层次及各林层物种组成特征
ssi指数最大时对应分类组数为最佳分类组数。由图1a的右图可知, 分类组数为5类时ssi指数最大; 同时, 根据不同分类组数时各分类组的组内平方和变化趋势图(图1b)可知, 当分类组数为4时组内平方和大小变化开始趋于平缓, 4类之后基本不变。因此, 综合考虑这两个指数, 确定最优树高聚类组数为5类, 即将塔吊样地群落垂直结构划分为5层: 即林层从下到上树高范围依次为[1.4, 4) m、[4, 6.8) m、[6.8, 11.3) m、[11.3, 17.4) m、[17.4, 27.1] m, 对应于传统垂直结构分类中的灌木层、亚冠层、林冠层, 林冠层进一步划分为林冠下层、林冠中层和林冠上层(表1)。
图1
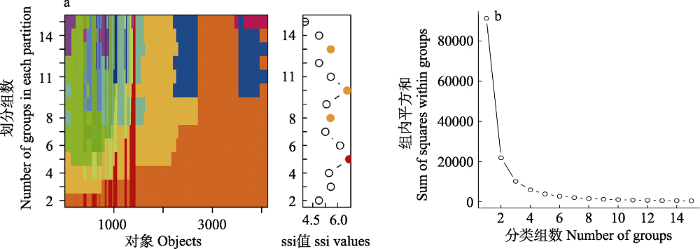
图1
最优分类组数判断指标。(a)不同分类情况下的各对象归属和各分类组数对应的ssi (simple structure index)值, 其中左边的图表示每个对象在不同分类情况下的归属, 一种颜色代表一个分类组; 右图展示了不同分类组数对应的ssi值大小。(b)不同分类组数时分成的各类的组内平方和。
Fig. 1
The judgment index partitioning optimal groups. (a) Attribution of the objects and the corresponding ssi (simple structure index) values for different number of groups. The panel on the left shows the groups that each object is categorized with different conditions and different groups were distinguished by different colors; the panel on the right shows the value of ssi for different number of groups. (b) Value of sum of squares within groups for different number of groups.
表1 鼎湖山塔吊样地垂直各层概况
Table 1
| 最低树高 Minimum height (m) | 最高树高 Maximum height (m) | 多度 Abundance | 物种丰富度 Species richness | 优势种例举(多度, 最大胸径) Dominant species (abundance, maximum DBH) | ||
|---|---|---|---|---|---|---|
| 灌木层 Shrub | 1.4 | 3.9 | 1,966 | 89 | 银柴 Aporosa dioica (207, 7.3) | 黄果厚壳桂 Cryptocarya concinna (251, 4.5) |
| 亚冠层 Sub-canopy | 4.0 | 6.7 | 1,173 | 75 | 鸭脚木 Schefflera octophylla (161, 16.0) | 银柴 Aporosa dioica (122, 10.1) |
| 林冠下层 Lower canopy | 6.8 | 11.2 | 503 | 56 | 荷木 Schima superba (54, 39.4) | 鸭脚木 Schefflera octophylla (51, 18.8) |
| 林冠中层 Middle canopy | 11.3 | 17.2 | 271 | 27 | 荷木 Schima superba (101, 41.9) | 锥栗 Castanopsis chinensis (31, 41.0) |
| 林冠上层 Upper canopy | 17.4 | 27.1 | 226 | 17 | 荷木 Schima superba (111, 66.0) | 马尾松 Pinus massoniana (34, 47.1) |
| 总体 Total | - | - | 4,140 | 121 | 荷木 Schima superba (301,49.0) | 马尾松 Pinus massoniana (66, 47.1) |
由表1可知, 随垂直高度上升, 塔吊样地群落各林层的物种丰富度和多度均明显减小。从各林层的优势种种类、数量及其径级特征可以发现, 群落不同林层的优势种种类差异大, 径级大小随群落林层向上明显增大, 但物种多度不断减小。综合可知, 塔吊样地各林层特征如下: 灌木层群落均为小径级个体(DBH < 10 cm), 多数为群落内乔木种的幼树, 还有一部分灌木树种, 该林层内植物个体数量和丰富度均最多, 基本包含了塔吊样地森林林层中的全部植物种类, 该层内的优势树种为银柴(Aporosa dioica)和黄果厚壳桂(Cryptocarya concinna)。亚冠层多为中径级个体(DBH < 20 cm), 物种数量占样地总物种丰富度的84%, 多度为灌木层的60%。林内物种多为小乔木或乔木, 优势种为鸭脚木(Schefflera octophylla)和银柴。林冠下层中等径级和大径级个体均有, 物种数量占样地总物种丰富度的46%, 多度仅为灌木层的25.6%, 基本为乔木, 优势种为荷木和鸭脚木。林冠中层和上层均多为大径级个体(DBH ≥ 20 cm), 中层优势种为荷木和锥栗, 上层物种数和多度最少, 优势种为荷木和马尾松, 且上层的优势种也是整个样地的优势物种。同时, 林冠中层和上层优势种个体数量分布十分不均匀, 如林冠中层的荷木多度(101株)明显大于锥栗(31株)和马尾松(34株)。
表2 鼎湖山塔吊样地灌木层与其他各林层及相邻林层间物种组成比较
Table 2
| 共有种总数量 Generalist (%) | 特有种总数量 Specialist (%) | 太少未能分类 Too rare to clarify (%) | 总计 Total | |
|---|---|---|---|---|
| 灌木层和亚冠层 Shrub vs sub-canopy | 24 (23.08) | 4 (3.98) | 76 (73.08) | 104 |
| 灌木层和林冠下层 Shrub vs lower canopy | 17 (16.50) | 7 (6.79) | 79 (76.70) | 103 |
| 灌木层和林冠中层 Shrub vs middle canopy | 8 (8.42) | 11 (11.58) | 76 (80.00) | 95 |
| 灌木层和林冠上层 Shrub vs upper canopy | 1 (1.06) | 14 (14.89) | 79 (84.08) | 94 |
| 亚冠层和林冠下层 Sub-canopy vs lower canopy | 19 (20.43) | 3 (3.23) | 71 (76.34) | 93 |
| 林冠下层和林冠中层 Lower canopy vs middle canopy | 10 (17.24) | 4 (6.89) | 44 (75.86) | 58 |
| 林冠中层和林冠上层 Middle canopy vs upper canopy | 5 (21.80) | 4 (3.13) | 23 (75.00) | 32 |
3.2 群落各林层α多样性特征
塔吊样地各林层α多样性指数大小如表3所示, Shannon-Wiener指数随林层向上而降低, 灌木层最高, 林冠上层最低, 且各林层之间差异显著(P < 0.05)。但Pielou均匀度指数表现却不同: 随林层向上, 均匀度指数呈单峰型变化, 林冠下层的均匀度指数最高, 其次是林冠中层和林冠上层, 灌木层最小, 且除林冠下层与灌木层之外, 其他层间差异均不显著。
表3 鼎湖山塔吊样地各林层的α多样性
Table 3
| 林层 Layers | Shannon-Wiener指数 Shannon-Wiener index | Peliou均匀度指数 Peliou evenness index |
|---|---|---|
| 灌木层 Shrub | 2.556 ± 0.335a | 0.910 ± 0.040a |
| 亚冠层 Sub-canopy | 2.300 ± 0.529b | 0.917 ± 0.033ab |
| 林冠下层 Lower canopy | 1.955 ± 0.334c | 0.940 ± 0.027ab |
| 林冠中层 Middle canopy | 1.440 ± 0.497d | 0.925 ± 0.232ab |
| 林冠上层 Upper canopy | 1.172 ± 0.519e | 0.896 ± 0.275b |
Different letters indicate the significant difference among layers in the same column.
同一列中, 不同字母表示不同林层间差异显著。
3.3 群落各林层β多样性及分解特征
图2
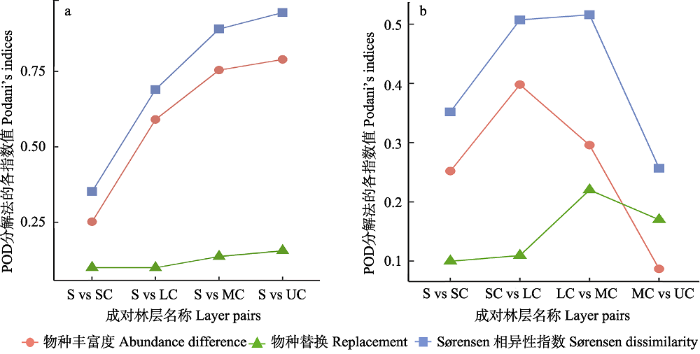
图2
鼎湖山塔吊样地各林层基于物种多度数据的β多样性大小(Sørensen相异性指数)与分解结果。(a)以灌木层为参照, 沿林层向上灌木层与其他各林层间β多样性大小和分解组分大小。(b)相邻两林层间的β多样性大小和分解组分大小。S: 灌木层; SC: 亚冠层; LC: 林冠下层; MC: 林冠中层; UC: 林冠上层。
Fig. 2
Results of β diversity (Sørensen dissimilarity) and their partitions based on species abundance data of each layers in the crane plot of Dinghushan. (a) Layers relative to the shrub layer. (b) Neighbouring layers. S, Shrub; SC, Sub-canopy; LC, Lower canopy; MC, Middle canopy; UC, Upper canopy.
与上述结果相比, 相邻两林层间的β多样性随林层变化则呈现不同趋势。随林层向上, β多样性大小先增大后减小, 呈单峰型(图2b)。即林冠下层与林冠中层间的β多样性最大, 林冠中层与林冠上层间最小, 林冠下层与亚冠层间的β多样性略低于最高值(0.507 vs 0.516), 即林冠下层与亚冠层和林冠上层间物种组成差异均较大, 以林冠下层为分界, 非林冠层之间及林冠层之间物种组成差异均较小。从分解的两个过程来看, 除林冠中层与林冠上层这两个林层间外, 其他相邻林层间物种丰富度差异组分均大于物种替换组分。但两个过程在各层间的变化趋势不同。物种丰富度差异组分大小在β多样性达到峰值的林层间开始降低, 而替换组分却明显增大并达到最大值。说明林冠下层与林冠中层间物种种类差异大, 群落间物种组成差异大。
4 讨论
4.1 群落垂直层次结构特征
根据样地内所有个体的树高进行聚类, 可将鼎湖山塔吊样地群落的垂直结构划分为5个林层, 分别命名为灌木层、亚冠层、林冠下层、林冠中层、林冠上层。这与叶万辉等(2008)依据群落外貌特征和物种不同生态型将鼎湖山南亚热带常绿阔叶林划分为灌木层、小乔木层和乔木层的结果相似。另外由塔吊样地所有个体的树高散点图(图3)可以看出, 塔吊样地垂直高度上的自然分层现象明显, 聚类划分的各层之间树高分布密集。
图3
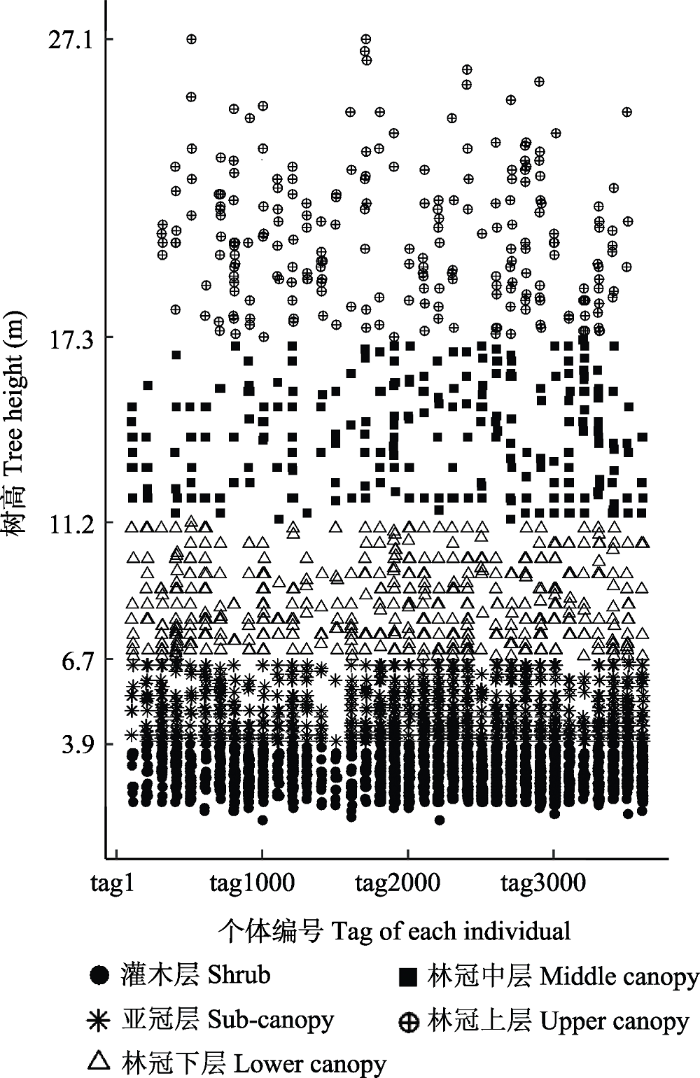
图3
鼎湖山塔吊样地所有个体树高分布散点图
Fig. 3
Tree height scatter points of each individual in the crane plot of Dinghushan
在一些研究中, 研究者常根据经验以5 m或10 m为间隔划分垂直各林层。如划分北方森林垂直结构时便以5 m下为一层, 其上以10 m为间隔 (Harms et al, 2001; Hao et al, 2007)。本文各林层间树高高度间隔先小后大, 层次更丰富(表1), 这与塔吊样地处于南亚热带区域, 森林结构较北方森林更为复杂有关(Smith, 1973)。同时塔吊样地各林层间隔与经验值5 m、10 m相差不大, 可见利用5 m或10 m树高间隔粗略划分森林垂直结构具有一定的合理性。
4.2 群落各林层α多样性特征
在塔吊样地的5个林层中, 各类α多样性指数均随林层向上呈递减趋势, 灌木层最大; 而均匀度呈单峰型, 在林冠下层时最大。基于物种重要值计算的多样性指数大小排除了个体数量的干扰, 说明该森林群落内灌木层α多样性最高的主要原因是其物种丰富度最大, 且物种多度分布并不均匀(表1)。对比各层间的物种特征, 灌木层丰富度最高的原因是: 首先, 除少数仅在林冠层出现的物种(如马尾松、枫香(Liquidambar formosana)等)和少数仅在亚冠层出现的稀少种(如岭南杜鹃(Rhododendron mariae)、铁冬青(Ilex rotunda)等)外, 群落内的其他物种均有小径级个体存在于灌木层中; 其次, 群落内还有一些仅存在于该层的灌木树种, 如九节(Psychotria rubra)、岭南山竹子(Garcinia oblongifolia)等(附录2)。可以说灌木层物种基本上是该群落的物种总集。通过均匀度指数的比较发现, 亚冠层的均匀度最高, 这可能是因为一方面如荷木、锥栗等林冠层优势种在该层中的个体多度开始增多, 另一方面非林冠层(灌木层和亚冠层)的优势种如鸭脚木、黄果厚壳桂等多度减少, 使得该层物种间多度差异变小。
4.3 群落各林层β多样性及其分解特征
随林层向上, 塔吊样地群落的灌木层与各林层间的总体β多样性呈递增趋势, 且主要由物种丰富度差异引起。而物种丰富度差异组分表示在特定环境梯度下物种的丧失或增加(Legendre, 2014; Li et al, 2016)。说明该样地森林从灌木层到林冠上层, 各林层物种不断丧失。这与随林层向上各林层物种丰富度不断下降一致(表1, 2)。随林层向上, 对比各层群落与灌木层的共有种和特有种种类及数量可知, 这种减少主要表现在共有种的减少, 如黄果厚壳桂、银柴和鸭脚木等非林冠层优势种在林冠层少量存在或消失, 仅荷木、锥栗等少数物种到达林冠上层并成为优势种。造成这一现象的原因可能是物种本身特性差异和环境资源变化的共同影响(Si et al, 2016)。
值得注意的是, 相邻林层间β多样性大小及其组分随林层向上的变化格局均呈单峰型, 但峰值点位置不同。林冠下层与亚冠层和林冠中层间β多样性均较大, 且物种周转组分明显增大。结合林冠下层与这两个林层的物种比较结果, 发现与亚冠层相比, 林冠下层的特有种为林冠中层的优势种荷木; 而与林冠中层相比, 林冠下层特有种为黄果厚壳桂等非林冠层优势种。Rahbek (2005)研究发现, β多样性随海拔上升也呈单峰型变化。不少研究者认为出现单峰型的原因可能是因为高海拔物种与低海拔物种在单峰型峰处交汇而发生了物种周转(替换) (Ricklefs, 1987; Brown, 2001; McCain & Beck, 2016)。鼎湖山塔吊样地森林群落垂直结构上的变化与之相似, 极有可能是林冠层与非林冠层在林冠下层交汇, 且发生了明显的物种替换, 同时也暗示林冠下层在该森林垂直结构中所处位置的特殊性。
另外, 林冠中层与林冠上层间β多样性最低, 这与丰富度差异组分非常低有关, 同时物种替换组分相对更大, β多样性主要由物种替换引起。在林冠上层中, 枫香、朴树(Celtis sinensis)等物种仅在该层中出现, 它们是塔吊样地群落中演替早期遗留下来的先锋种, 或早期人工培植的乡土物种。这些物种先到达群落并先占据林冠层, 在群落演替后期, 它们排斥后续到达的物种(Fukami, 2015), 因此认为优先效应可能是导致林冠中层与林冠上层间发生物种替换的原因。
4.4 鼎湖山森林群落垂直结构的构建机制
HilleRisLambers等(2012)认为群落构建是多个尺度生态学过程共同作用的结果, 局域尺度上的物种共存是环境过滤和物种相互作用共同驱动的结果。森林群落垂直方向上的成层现象导致了林内环境因子的高异质性。根据小气候监测数据可知, 从灌木层到林冠上层, 塔吊样地各林层平均温度和平均光照均呈增大趋势, 而平均相对空气湿度呈减小趋势, 日平均光照整体上也呈上升趋势。另外, 从相邻林层间的环境因子来看, 林冠中层的日平均光照较之林冠下层变化最大(附录3)。该群落β多样性及其分解组分的变化情况表明, 林冠下层与林冠中层间物种组成差异最大且物种替换组分最大。前人的研究认为, 导致物种替换的潜在机制包括环境过滤、竞争、地理隔离等(Angeler, 2013; Legendre, 2014)。塔吊样地各林层在垂直空间上并无地理隔离, 结合群落内各林层的环境因子变化, 最有可能是环境过滤限制了某些物种不能到达林冠中层, 而能通过环境筛选的物种在林冠中层出现, 且限制因子主要为光照。竞争作用对本群落构建的影响具体如何还有待进一步的研究。最后需要指出的是, 物种本身的生物学特性也会影响群落垂直结构的构建, 如物种的生活型, 灌木种和乔木种的树高本身就存在差异, 多数灌木物种树高较低, 仅在灌木层中出现。因此研究森林群落垂直结构的构建过程可以从物种本身属性、非生物因素(环境过滤)和生物因素3个方面考虑。
综上所述, 鼎湖山塔吊样地垂直层次可划分为灌木层、亚冠层、林冠下层、林冠中层和林冠上层5层。随林层向上各林层物种多度不断减小, 且丰富度指数不断减小, 林层间β多样性增大主要由物种丰富度差异所致, 而相邻林层间的β多样性呈单峰型。林冠下层可能是垂直结构上的特殊位置, 有较强的环境筛选作用, 限制因子主要为光照。
本研究仅仅是对塔吊样地垂直结构及其群落构建机制的初步探究, 希望通过本研究能进一步从更多的视角(如谱系、功能性状等)量化各林层内物种共存的影响过程, 深入阐释有关群落构建机制的问题。
附录 Supplementary Material
附录1 鼎湖山塔吊样地微气候监测仪器放置高度
Appendix 1 Height of all micro-climate monitoring equipments placement sites in the crane plot of Dinghushan
附录2 鼎湖山塔吊样地灌木层与其他各林层及相邻林层间物种组成比较
Appendix 2 Comparison of species component between shrub and other layers as well as among neighbouring layers in pair in the crane plot of Dinghushan
附录3 鼎湖山塔吊样地各林层微环境
Appendix 3 Microenvironment factors of each layer in the crane plot of Dinghushan
参考文献
Revealing a conservation challenge through partitioned long-term beta diversity: Increasing turnover and decreasing nestedness of boreal lake metacommunities
DOI:10.1111/ddi.2013.19.issue-7 URL [本文引用: 2]
Comparisons of structure among mixed dipterocarp forests of north-western Borneo
DOI:10.2307/2260691 URL [本文引用: 1]
Partitioning the turnover and nestedness components of beta diversity
DOI:10.1111/j.1466-8238.2009.00490.x URL [本文引用: 1]
Betapart: An R package for the study of beta diversity
DOI:10.1111/j.2041-210X.2012.00224.x URL [本文引用: 1]
Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients
DOI:10.1111/2041-210X.12029 URL [本文引用: 1]
Mammals on mountainsides: Elevational patterns of diversity
DOI:10.1046/j.1466-822x.2001.00228.x URL [本文引用: 1]
A novel statistical method for classifying habitat generalists and specialists
DOI:10.1890/10-1345.1 URL [本文引用: 1]
A review of beta diversity studies
Beta多样性研究进展
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
A tale of three cities: Perceptual charting for analyzing destination images
Historical contingency in community assembly: Integrating niches, species pools, and priority effects. Annual Review of Ecology,
DOI:10.1146/annurev-ecolsys-110411-160340 URL [本文引用: 1]
Prospects for quantifying structure, floristic composition and species richness of tropical forests
A comparison of montane and lowland rain forest in ecuador. I. The forest structure, physiognomy, and floristics
DOI:10.2307/2257748 URL [本文引用: 1]
Vertical structure and spatial associations of dominant tree species in an old- growth temperate forest
DOI:10.1016/j.foreco.2007.06.026 URL [本文引用: 1]
Habitat associations of trees and shrubs in a 50-ha neotropical forest plot
DOI:10.1111/jec.2001.89.issue-6 URL [本文引用: 1]
Beta diversity on geographic gradients in Britain
DOI:10.2307/5518 URL [本文引用: 1]
Rethinking community assembly through the lens of coexistence theory
Plant species diversity dynamics in Dinghu Mountain forests
鼎湖山植物物种多样性动态
A method for quantifying vertical forest structure
DOI:10.1016/S0378-1127(97)00254-5 URL [本文引用: 1]
Interpreting the replacement and richness difference components of beta diversity
DOI:10.1111/geb.12207 URL [本文引用: 3]
Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning
DOI:10.1111/ele.2013.16.issue-8 URL [本文引用: 1]
The geographical structure of British bird distributions: Diversity, spatial turnover and scale
DOI:10.1046/j.0021-8790.2001.00563.x URL [本文引用: 2]
The research advances on the structure and function of forest canopy, as well as their temporal and spatial changes
森林冠层结构与功能及其时空变化研究进展
Convergence and divergence in a long-term old-field succession: The importance of spatial scale and species abundance
DOI:10.1111/ele.12647 URL [本文引用: 1]
Forest canopies: Methods, hypotheses, and future directions
DOI:10.1146/annurev.ecolsys.27.1.55 URL [本文引用: 1]
Measurement of biotic community diversity. I. α diversity (Part 2)
生物群落多样性的测度方法. I. α多样性的测度方法(下)
Species turnover in vertebrate communities along elevational gradients is idiosyncratic and unrelated to species richness
DOI:10.1111/geb.12410 URL [本文引用: 1]
A method of access into the crowns of emergent and canopy trees
DOI:10.2307/2388019 URL [本文引用: 1]
A new conceptual and methodological framework for exploring and explaining pattern in presence-absence data
DOI:10.1111/j.1600-0706.2011.19451.x URL [本文引用: 2]
The role of spatial scale and the perception of large-scale species-richness patterns
Community diversity: Relative roles of local and regional processes
DOI:10.1126/science.235.4785.167 URL [本文引用: 1]
On Chinese forest canopy biodiversity monitoring
中国森林冠层生物多样性监测
Selective extinction drives taxonomic and functional alpha and beta diversities in island bird assemblages
DOI:10.1111/1365-2656.12478 URL [本文引用: 1]
Beta-diversity partitioning: Methods, applications and perspectives
Beta多样性分解: 方法、应用与展望
Stratification of temperate and tropical forests
DOI:10.1086/282866 URL [本文引用: 1]
A consumer’s guide to nestedness analysis
DOI:10.1111/oik.2009.118.issue-1 URL [本文引用: 1]
On the ecology of British beechwoods with special reference to their regeneration. Part II. The development and structure of beech communities on the Sussex Downs
DOI:10.2307/2255242 URL [本文引用: 1]
Evolution and measurement of species diversity
DOI:10.2307/1218190 URL [本文引用: 1]
Mapping variations in the strength and breadth of biogeographic transition zones using species turnover
DOI:10.1098/rspb.1996.0087 URL [本文引用: 1]
Advances in ecological studies of epiphytes using canopy cranes
基于林冠塔吊的附生植物生态学研究进展
Identifying canopy species of subtropical forest by lightweight unmanned aerial vehicle remote sensing
基于微型无人机遥感的亚热带林冠物种识别
Community structure of a 20 hm 2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China
鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究
Review of forest vertical structure parameter inversion based on remote sensing technology
森林垂直结构参数遥感反演综述
Patterns and determinants of beta diversity in the understory vegetation layers of montane boreal conifer forest in Luya Mountain
芦芽山寒温性针叶林冠层下植被beta多样性格局及其成因

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}