


物种多样性分布格局及其影响因素是生物多样性研究中的热点问题。α和β多样性是物种多样性研究中的两个常用指标, 其中α多样性是对群落物种丰富度和均匀度的评估, β多样性则度量群落的物种组成沿环境梯度的周转速率(Whittaker, 1972; 马克平, 1994; 马克平等, 1995)。
森林群落的物种多样性分布格局受到多种因素影响(李亚藏等, 2006; Qian et al, 2014), 包括非生物环境因子、群落类型和空间距离等(唐志尧和方精云, 2004; Guillem et al, 2006; Gatti et al, 2015; Tello et al, 2015; 王月霞等, 2016)。很多研究发现非生物环境因子例如地形是影响森林群落α和β多样性的重要因子(Whittaker & Niering, 1965; Vazquez & Givnish, 1998; Wang et al, 2002), 但也有研究发现群落类型是影响森林α和β多样性的重要因子(王月霞等, 2016)。此外, 空间距离通过扩散限制等过程也可影响森林β多样性, 表现为随着空间距离增加, 物种组成相似程度减小, 群落间β多样性增大(Hubbell, 2001; 牛克昌等, 2009; Blundo et al, 2016)。
迄今, 国内外学者对于植物物种多样性格局已开展过大量的研究(Gallardo-Cruz et al, 2009; Qian & Shimono, 2012; 蒙洋等, 2017), 主要集中在两个尺度: (1)单个大型样地的尺度, 如Legendre等(2009)对古田山24 ha亚热带常绿阔叶林β多样性及影响因素的研究, Condit (2005)对热带地区13个大型样地森林物种α多样性的比较研究; (2)区域尺度, 如王志恒等(2004)对高黎贡山种子植物物种丰富度沿海拔梯度变化的研究, López-Martínez等(2012)对墨西哥尤卡坦半岛次生热带干林木本植物不同尺度β多样性影响因素的研究。但以往研究中对单个自然保护区尺度(~100 km2)上的多种类型森林群落α和β多样性及其差异驱动机制的研究还较少, 仅见卢训令等(2010)对河南省伏牛山自然保护区物种多样性的研究。
古田山国家级自然保护区(以下简称古田山保护区)地处中亚热带地区, 地形复杂, 森林群落类型较为丰富(吴征镒, 1980; 陈灵芝等, 1997; 宋永昌, 1999), 区内分布着常绿阔叶林、常绿落叶阔叶混交林、针阔叶混交林、针叶林和人工林等多种群落类型(于明坚等, 2019), 为我们提供了良好的研究对象。本文以古田山保护区内79个20 m × 20 m森林群落样地调查数据为基础, 拟探讨如下问题: (1)古田山森林的α和β物种多样性分布格局如何? (2) α和β多样性分布格局分别主要受到哪些因子的影响?
1 材料与方法
1.1 研究地点
古田山保护区(118°03°49.7-118°11°12.2 E; 29°10°19.4-29°17°41.4 N)属于白际山脉的一部分, 位于浙江省开化县西北部, 与江西省婺源县、德兴市毗邻。保护区总面积为8,107 ha, 主峰青尖海拔约为1,258 m。古田山属于亚热带季风气候区, 山体呈东北-西南走向, 受夏季风影响较大, 四季分明, 降水丰富, 年平均降水量约1,963.7 mm, 相对湿度92.4%。年平均气温约为15.3℃, 最高气温38.1℃, 最低气温-6.8℃, 生长期总积温为5,221.5℃, 无霜期约250 d。保护区的土壤类型主要有红壤、黄红壤、红黄壤及沼泽土, 前三者合计超过全区总面积的80%。
1.2 群落类型划分
森林群落类型划分主要参照《中国植被》(吴征镒等, 1980)的分类原则, 高级分类单位以生态外貌为主要依据, 中低级分类单位依据优势种和标志种相结合的原则。其中, 优势度以各木本植物种类的重要值表示。我们将所有种类先分为2类: 针叶植物和阔叶植物, 再将后者分为2类: 常绿阔叶植物和落叶阔叶植物。若某一类的重要值不超过另一类的2倍则划分为混交林, 超过则划分为纯林。根据上述分类原则, 将古田山保护区内的群落类型分为以下10种: 常绿阔叶林、常绿落叶阔叶混交林、黄山松(Pinus taiwanensis)林、黄山松针阔叶混交林、落叶阔叶林、马尾松(Pinus massoniana)林、马尾松针阔叶混交林、杉木(Cunninghamia lanceolata)林、杉木针阔叶混交林和油茶(Camellia oleifera)林(于明坚等, 2019)。
1.3 样地设置及调查
图1
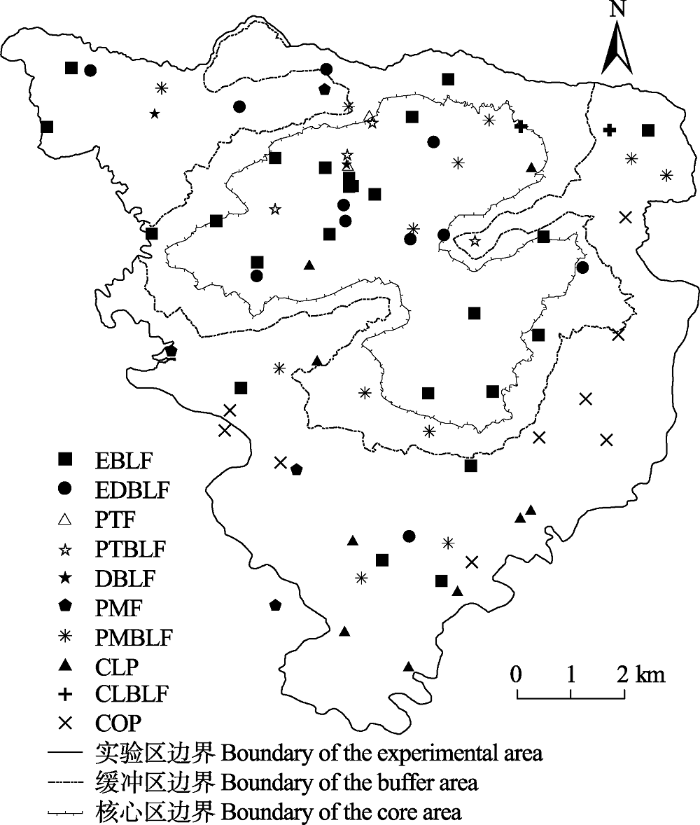
图1
79个森林群落样地分布图。EBLF、EDBLF、PTF、PTBLF、DBLF、PMF、PMBLF、CLP、CLBLF和COP的含义见
Fig. 1
The locations of 79 forest community plots. EBLF, EDBLF, PTF, PTBLF, DBLF, PMF, PMBLF, CLP, CLBLF and COP are the same as in
表1 各森林群落类型面积及样地个数、取样面积
Table 1
| 群落类型 Community type | 群落面积 Community area (ha) | 样地个数 Number of plots | 取样面积 Sample area (ha) |
|---|---|---|---|
| 常绿阔叶林 Evergreen broad-leaved forest (EBLF) | 4,778.9 | 24 | 0.96 |
| 常绿落叶阔叶混交林 Evergreen and deciduous broad-leaved mixed forest (EDBLF) | 444.1 | 11 | 0.44 |
| 黄山松林 Pinus taiwanensis forest (PTF) | 14.3 | 2 | 0.08 |
| 黄山松针阔叶混交林 Pinus taiwanensis and broad-leaved trees mixed forest (PTBLF) | 152.7 | 4 | 0.16 |
| 落叶阔叶林 Deciduous broad-leaved forest (DBLF) | 18.5 | 2 | 0.08 |
| 马尾松林 Pinus massoniana forest (PMF) | 217.8 | 4 | 0.16 |
| 马尾松针阔叶混交林 Pinus massoniana and broad-leaved trees mixed forest (PMBLF) | 329.6 | 12 | 0.48 |
| 杉木林 Cunninghamia lanceolata plantation (CLP) | 549.8 | 9 | 0.36 |
| 杉木针阔叶混交林 Cunninghamia lanceolata and broad-leaved trees mixed forest (CLBLF) | 30 | 2 | 0.08 |
| 油茶林 Camellia oleifera plantation (COP) | 1,516.1 | 9 | 0.36 |
记录每个样地的经纬度和各种地形因子(包括海拔、坡度和坡向等)。其中, 经纬度在样地几何中心点测定, 海拔为每个20 m × 20 m样地4个顶点海拔的平均值; 坡度为样地4个角中任意3个所构成的4个平面与水平面夹角的平均值; 坡向为这4个平面与正北方向夹角的平均值。空间距离为两个样地中心点水平面投影的直线距离。
1.4 多样性指数
α多样性用Shannon-Wiener指数(H')来衡量。采用Rarefaction方法比较不同森林群落类型间物种α多样性的差异, 方法如下: 针对一种群落类型, 先确定单个样地所含的平均植株个体数N; 然后从这种群落类型的所有个体中随机抽取N个植株, 重复100次, 得到100个物种丰富度值, 即为该群落类型的样地水平物种丰富度。
β多样性采用Horn-Morisita相异性指数(Horn-Morisita dissimilarity index, CMH)来衡量(Wolda, 1983)。
式中, pi为物种相对多度, ai和bi分别为A和B样地中第i种的个体数目, Na为样地A的物种数, Nb为样地B的物种数。
1.5 数据分析
为检验地形因子和群落类型等对α多样性的影响, 采用lme4软件包中的lmer函数(Bates et al, 2014)和stats软件包中的anova函数分别进行多元回归和方差分析。为比较森林群落β多样性分布格局的驱动因子, 进行了非度量多维标度排序(non-metric multidimensional scaling, NMDS)和偏Mantel检验等分析。NMDS用于分析不同群落间β多样性分布的大致情况, 通过vegan软件包中的metaMDS函数(Oksanen et al, 2016)完成。偏Mantel检验的多元回归手段用于检验空间距离(通过经纬度计算得出)、地形因子和群落类型对β多样性分布格局的影响, 通过phytools软件包中的multi.mantel函数(Revell, 2012)完成。组间比较采用t-检验, 多重比较的P值采用Holm法调整, 通过stats软件包中的pairwise.t.test函数完成。以上所有统计分析通过R3.2.4软件(R Core Team, 2016)完成。
2 结果
2.1 α多样性分布格局及其影响因素
图2
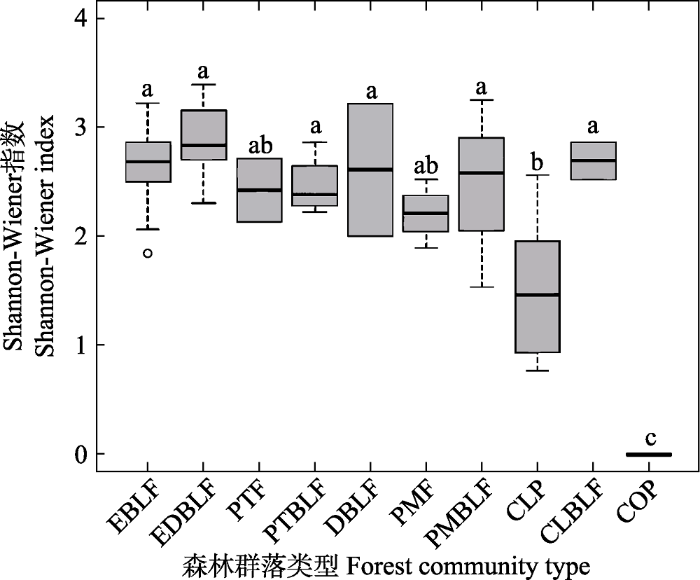
图2
古田山国家级自然保护区不同类型群落α多样性的差异。柱形上方的英文字母不同表示柱形间有显著差异(P < 0.05, Holm法修正), 否则无显著差异。EBLF、EDBLF、PTF、PTBLF、DBLF、PMF、PMBLF、CLP、CLBLF和COP的含义见
Fig. 2
The difference of α diversity in different community types in Gutianshan National Nature Reserve. The different letter on the top of the column shows that there is significant difference between the groups (P < 0.05, Holm method correction), otherwise there is no significant difference. EBLF, EDBLF, PTF, PTBLF, DBLF, PMF, PMBLF, CLP, CLBLF and COP are the same as in
图3
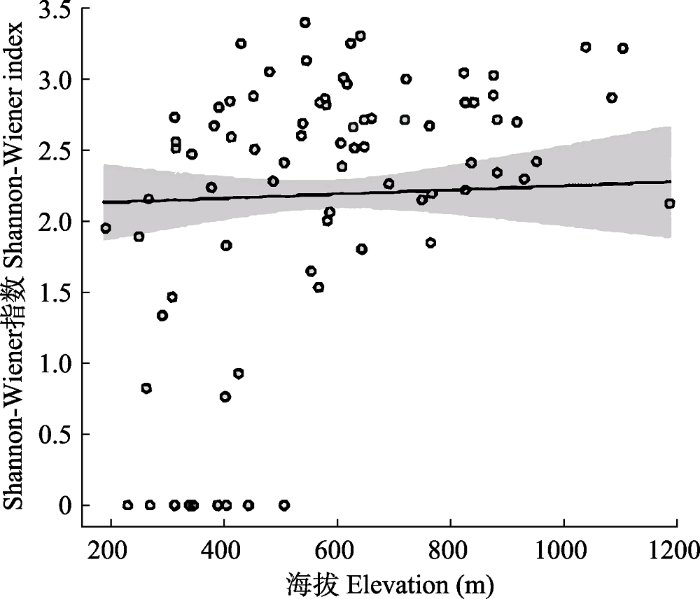
图3
古田山国家级自然保护区森林群落α多样性(Shannon- Wiener指数)随海拔梯度的变化。圆圈为实际观测值; 实线为拟合值; 阴影为95%置信区间。
Fig. 3
The forest community α diversity (Shannon-Wiener index) along an altitudinal gradient in Gutianshan National Nature Reserve. The circles are actual observations; the solid line is the fitted value; the shadow is 95% confidence intervals.
图4
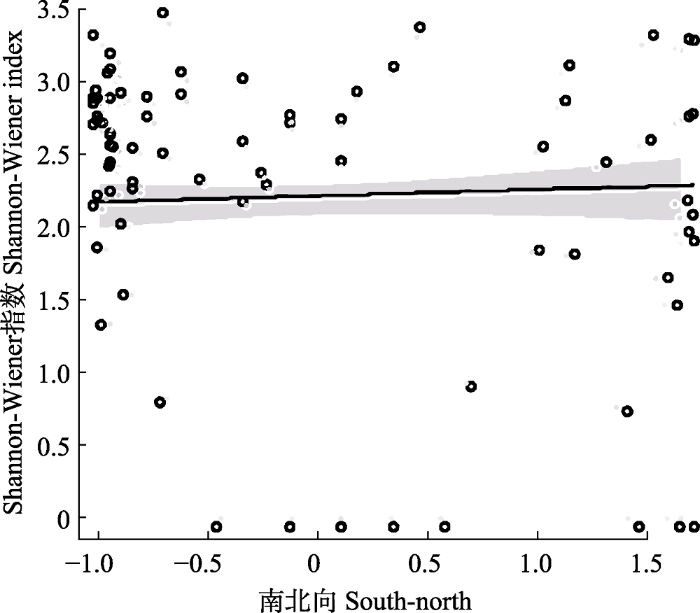
图4
古田山国家级自然保护区森林群落α多样性(Shannon- Wiener指数)随南北向的变化。圆圈为实际观测值; 实线为拟合值; 阴影为95%置信区间。
Fig. 4
The forest community α diversity (Shannon-Wiener index) along the north direction in Gutianshan National Nature Reserve. The circles are actual observations; the solid line is the fitted value; the shadow is 95% confidence intervals.
表2 地形和群落类型对古田山国家级自然保护区森林群落α多样性(Shannon-Wiener指数)影响的方差分析结果
Table 2
| 自由度 Degree of freedom | 总方差 Total variance | 均方差 Mean variance | F | P | |
|---|---|---|---|---|---|
| 海拔 Elevation | 1 | 15.716 | 15.716 | 91.393 | < 0.001 |
| 坡度 Slope | 1 | 0.550 | 0.550 | 3.200 | 0.078 |
| 东西向 East-west | 1 | 0.414 | 0.414 | 2.407 | 0.126 |
| 南北向 South-north | 1 | 0.828 | 0.828 | 4.817 | 0.032 |
| 群落类型 Community type | 9 | 42.872 | 4.764 | 27.702 | < 0.001 |
| 残差 Residuals | 65 | 11.177 | 0.172 |
2.2 β多样性分布格局及其影响因素
通过NMDS分析样地间β多样性的分布情况发现, 同一群落类型内部和不同群落类型间物种组成均存在差异(图5)。具体表现为: 同一群落类型的样地通常分布较为聚集, 如油茶林、杉木林、马尾松针阔叶混交林等; 不同森林类型的样地通常分布较为分散, 如油茶林、杉木林和马尾松针阔叶混交林等; 也有一些森林类型有交错分布, 如常绿阔叶林与黄山松针阔叶混交林、常绿落叶阔叶混交林、马尾松针阔叶混交林等。各森林群落类型的Horn-Morisita指数比较结果显示, 不同群落类型间的β多样性显著大于同一群落类型内部(图6)。偏Mantel检验的多元回归发现,空间距离、群落类型和地形共同解释了β多样性变异的26.21% (P < 0.001)。其中, 群落类型和海拔对β多样性有显著影响, 坡度、坡向和空间距离对β多样性无显著影响(表3)。具体表现为, 群落间的海拔差异增大, β多样性也随之增大(图7, 附录2)。空间距离单独对β多样性变异的解释度为1.44% (P < 0.001), 群落类型和地形共同的解释度达到26.16% (P < 0.001)。
图5
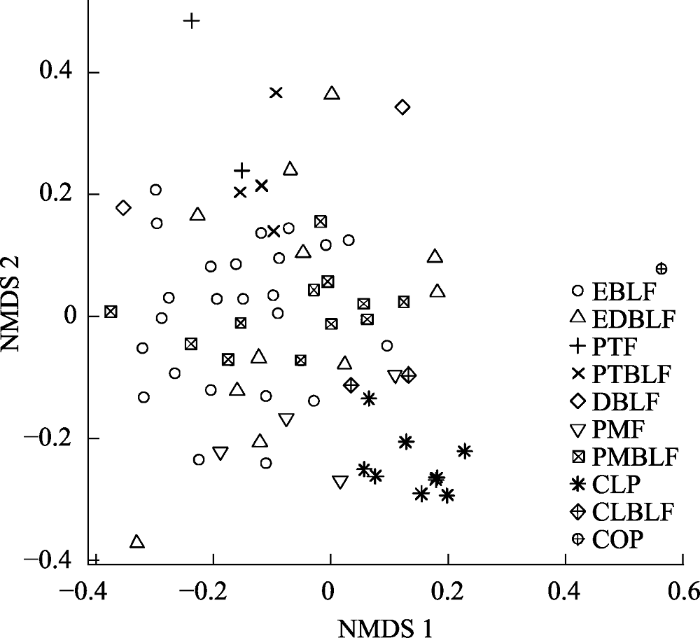
图5
古田山国家级自然保护区不同类型群落物种组成的非度量多维尺度分析排序结果。EBLF、EDBLF、PTF、PTBLF、DBLF、PMF、PMBLF、CLP、CLBLF和COP的含义见
Fig. 5
Non-metric Multidimensional scaling (NMDS) result of species composition of the different community types in Gutianshan National Nature Reserve. EBLF, EDBLF, PTF, PTBLF, DBLF, PMF, PMBLF, CLP, CLBLF and COP are the same as in
图6
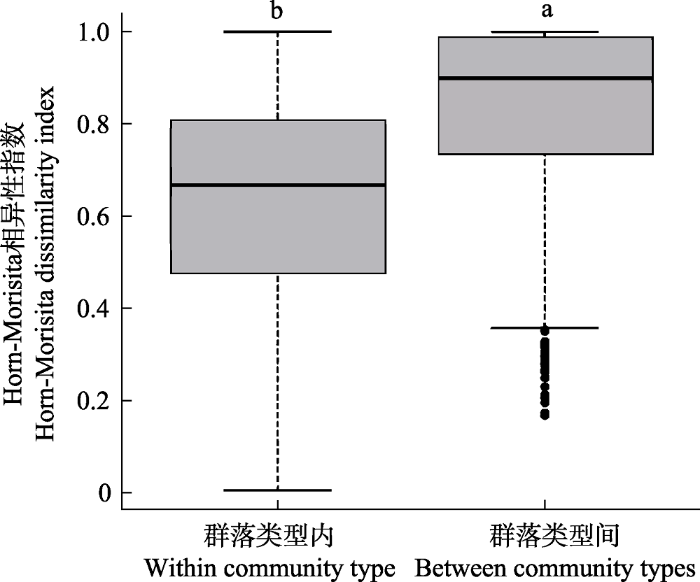
图6
同种群落类型内部和不同群落类型间β多样性(Horn-Morisita相异性指数)的比较。柱形图上方的英文字母不同表示柱形间有显著差异(P < 0.05, t-检验), 否则无显著差异。
Fig. 6
Comparison of β diversity (Horn-Morisita dissimilarity idex) within the same community type and between different community types. The different letter on the top of the column shows that there is significant difference between the groups (P < 0.05, t-test), otherwise there is no significant difference.
图7
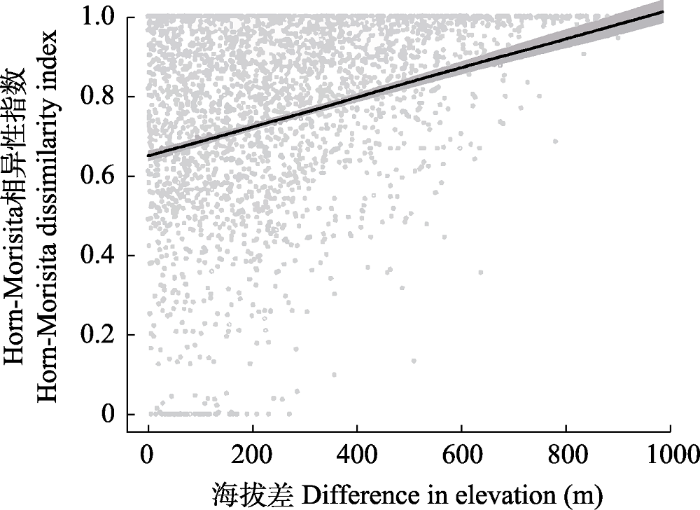
图7
古田山国家级自然保护区森林群落β多样性(Horn-Morisita相异性指数)随海拔梯度的变化。圆点为实际观测值; 实线为拟合值; 阴影为95%置信区间。
Fig. 7
The forest community β diversity (Horn-Morisita dissimilarity index) along an altitudinal gradient in Gutianshan National Nature Reserve. The circles are actual observations; the solid line is the fitted value; the shadow is 95% confidence intervals.
表3 空间距离、地形和群落类型对古田山国家级自然保护区森林群落β多样性(Horn-Morisita指数)的影响(R2 = 0.262)
Table 3
| 效应 Effect | t | P | |
|---|---|---|---|
| 空间距离 Spatial distance | 0.002 | 1.497 | 0.415 |
| 海拔 Elevation | 0.066 | 15.148 | 0.001 |
| 坡度 Slope | -0.006 | -1.635 | 0.391 |
| 南北向 North-South | 0.0052 | 1.370 | 0.347 |
| 东西向 East-West | 0.0035 | 0.836 | 0.635 |
| 群落类型 Community type | 0.2393 | 25.415 | 0.001 |
3 讨论
3.1 α多样性分布格局和影响因素
对古田山保护区10种森林群落类型Shannon- Wiener指数的比较发现, 不同森林群落类型间α多样性存在巨大差异。例如, 杉木林和油茶林这两种人工林的α多样性明显低于其他8种群落类型, 表明人类干扰可能降低了森林生物多样性。并且Rarefaction方法和方差分析均显示群落类型对α多样性有重要影响, 进一步说明群落类型在较大程度上决定了古田山保护区森林群落的α多样性格局。
海拔是一类特殊的环境因子, 反映了水分、温度和光照等因素的综合情况(Whittaker & Niering, 1965; Gallardo-Cruz et al, 2009; 郑江坤等, 2009)。一般认为, α多样性随海拔升高呈下降趋势; 也有研究表明, 随海拔升高, α多样性先增大后减小(Rahbek, 1977; 沈泽昊等, 2000); 或呈现出其他更复杂的变化规律(Peet, 1981; Baruch, 1984; Itow, 1991)。古田山保护区森林群落的α多样性随海拔升高而增大, 这一趋势与很多其他地区的研究结果并不一致。造成这种现象的原因可能是古田山海拔梯度较小(样地最高海拔不超过1,200 m), 高海拔样地光照等条件较为充足, 并且热量没有成为植物分布的限制因素, 受到人为干扰也较少, 因而高海拔样地反而具有较高的物种多样性。海拔对森林群落多样性的显著影响表明古田山群落物种在海拔梯度上的生态需求分化可能较明显, 有必要进一步研究驱动该格局的潜在环境因子。
坡向直接影响森林的光照, 进而作用于森林植物的光合作用和生境过滤, 影响到生物多样性分布格局(郑江坤等, 2009; 吴昊, 2017)。古田山保护区森林群落α多样性受到坡向的一定影响, 随样地坡向更加偏北, α多样性有升高趋势, 可能是由于南坡光照强、温度高、蒸发量大、湿度较小, 土壤风化强、有机质积累少等多种原因导致南坡物种丰富度较低。
3.2 β多样性分布格局和影响因素
本研究发现古田山保护区森林群落β多样性表现为, 同一类型群落样地分布通常较为集中, 其中以杉木林和油茶林分布最为集中。古田山保护区的杉木林和油茶林均属于人工林, 其中杉木林与马尾松针阔叶混交林、杉木针阔叶混交林等的分布有部分交错。杉木林在保护区内被严格保护, 正在向针阔叶混交林演替(马丹炜等, 2002; 钱海源等, 2018); 而油茶林因始终受到人为管理(如下层植物的清理), 物种组成单一, 与其他群落类型差异较大。
但是, 常绿阔叶林分布较为发散, 并且与黄山松针阔叶混交林、常绿落叶阔叶混交林、马尾松针阔叶混交林的分布有交错。该地区的常绿落叶阔叶混交林主要占据低海拔沟谷地带; 马尾松针阔叶混交林主要生长于较高海拔山脊; 黄山松针阔叶混交林除山脊外, 在更高海拔区域也有分布(于明坚等, 2001)。而常绿阔叶林广泛分布在海拔300-1,150 m范围内, 估计与常绿落叶阔叶混交林、马尾松针阔叶混交林和黄山松针阔叶混交林之间均有物种扩散和生境交错等过程, 故与这三种群落类型的物种组成均较为相似。
此外, 古田山保护区不同群落类型样地的分布通常较为分散。不同群落类型间的β多样性大于同一群落类型内部的β多样性, 说明群落类型在一定程度上代表了独特的物种组成。因此, 维持该地区群落类型的多样性也有利于维持该地区森林群落的物种多样性。该发现与王月霞等(2016)对于整个浙江省范围内多种群落类型β多样性的研究一致。
森林群落类型对古田山保护区物种β多样性的影响最大, 然后是地形因子。群落类型(效应值0.2393)的影响甚至高于海拔(效应值0.066)。群落类型与森林物种多样性格局的紧密关系, 可能与生境条件的过滤作用有关(Wilson & Gitay, 1995; 蒙洋等, 2017)。海拔差异对于β多样性的正效应反映了与海拔相关的环境因子例如水分、温度和光照等因素(Whittaker & Niering, 1965; 冶民生等, 2004)对于群落构建的重要作用。另一方面, 空间距离对β多样性格局没有显著作用, 可能是由于与其他相似研究的尺度相比(Myers et al, 2013; 王月霞等, 2016), 古田山保护区的空间尺度较小(样地距离范围绝大多数在10 km以内), 扩散限制等过程对于群落组成的影响较不明显。
综上所述, 古田山不同森林群落类型α、β多样性主要受到群落类型和地形(主要为海拔)的影响, 表明生境过滤过程对于保护区尺度上森林群落的构建起重要作用。该区域群落类型多样, 各群落类型有其独特的α、β多样性。对于森林保护来说, 自然森林群落类型的多样化有利于维持该区域森林群落物种多样性。
致谢: 感谢浙江大学王月霞、骆杨青等同学参与古田山样地调查及数据录入等工作, 感谢赖正淦、赖祯熙等在样地调查工作中付出的辛勤劳动, 感谢浙江大学刘金亮、仲磊和韦博良等同学在论文修改上提出宝贵意见。
附录 Supplementary Material
附录1 古田山国家级自然保护区不同类型群落稀疏化的α多样性差异
Appendix 1 The difference of rarefied α diversity between community types in Gutianshan National Nature Reserve
附录2 古田山国家级自然保护区不同群落类型内(A)和类型间(B) β多样性(Horn-Morisita相异性指数)随海拔梯度的变化
Appendix 2 The β diversity (Horn-Morisita dissimilarity index) along an altitudinal gradient within (A) and between (B) community types in Gutianshan National Nature Reserve
参考文献
Ordination and classification of vegetation along an altitudinal gradient in the Venezuelan Páramos
DOI:10.1007/BF00037333
URL
[本文引用: 1]

By means of ordination and classification techniques, the relationships between climate, soils, human activities and vegetation along an altitudinal gradient of the Venezuelan páramos are analyzed and interpreted. The altitudinal gradient chosen is characterized by decrease of temperature, precipitation, soil fertility, soil water-holding capacity, and plant cover as altitude increases. The ordination results suggest vegetation changes to be primarily related to environmental changes occurring with altitude, and secondly to disturbances caused mainly by grazing. Some results point toward a disjunction in the vegetational gradient occurring at ca. 3 500 m.a.s.l. and separating low and high páramo. This disjunction might have been caused by the glacial history of the páramos and the occurrence of frequent night-frosts.
lme4: Linear mixed-effects models using Eigen and S4
Relative contribution of niche and neutral processes on tree species turnover across scales in seasonal forests of NW Argentina
DOI:10.1007/s11258-016-0577-x
URL
[本文引用: 1]
Environmental heterogeneity and dispersal limitation influence tree species distribution, but their relative contributions change with the spatial scale of analysis. We analyzed tree species turnover...
Tropical tree α-diversity: Results from a worldwide network of large plots
DOI:10.1016/0022-4804(78)90159-2
URL
[本文引用: 1]
We assembled data on the diversity of tree species, genera, families, and orders in 13 large-scale forest inventory plots across the tropics. Each plot included at least 16 ha where every individual ≥1 cm in stem diameter was censused, providing a much more thorough measure of α-diversity than the typical 0.1- or 1-ha inventories. Amazonian and Southeast Asian plots had more than 1000 tree spec...
β-diversity and vegetation structure as influenced by slope aspect and altitude in a seasonally dry tropical landscape
DOI:10.1007/s10980-009-9332-1
URL
[本文引用: 2]
http://link.springer.com/article/10.1007%2Fs10980-009-9332-1
The impact of selective logging and clearcutting on forest structure, tree diversity and above-ground biomass of African tropical forests
DOI:10.1007/s11284-014-1217-3
URL
[本文引用: 1]
Tropical deforestation is well known to have serious negative consequences for biodiversity, terrestrial carbon sinks and the balance of atmospheric greenhouse gases. By contrast, selective logging of tropical forests is often regarded as having a lesser impact on the ecosystem particularly in long terms, even though there have been few critical evaluations of the practice, particularly in Africa. We compared field data from 511 plots in the tropical forest of Sierra Leone, Ghana, Cameroon and Gabon. These plots were subject to different forest management practices: no recent logging (primary forests), selective logging (up to 30 years old) and re-grown secondary forests post clear-cutting (at least 20 years ago). Our findings suggest that the vertical structure and plant richness of the selectively logged and secondary forests change in different amplitude from those of primary forests, but stem density and the prevalence of vine and weed species differ greatly. We show that the effects of selective logging are greater than those expected simply from the removal of commercial species, and can persist for decades. Selective logging, unless it is practiced at very low harvest intensities, can significantly reduce the biomass of a tropical forest for many decades, seriously diminishing aboveground carbon storage capacity, and create opportunities for weeds and vines to spread and slow down the ecological succession.
Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama
DOI:10.1111/j.1654-1103.2006.tb02426.x
URL
[本文引用: 1]
The objectives of this study were to clarify the extent to which environmental factors and geographical distance account for tropical floristic composition, and propose a methodology for delimiting the boundaries of floristic types based on species similarity.
Species turnover and diversity patterns along an elevation broad-leaved forest coenocline
DOI:10.2307/3236029
URL
[本文引用: 1]
Direct gradient analysis was applied to the evergreen broad-leaved forest coenocline in the Tatera Forest Reserve, Japan. 10 quadrats of 0.1-0.05 ha were laid out from 140 m to 560 m above sea level at intervals of 25-70 m. Gradient analysis revealed that distributions of many species terminated or started at ca. 400 m. Community similarity, calculated in Percentage Similarity (PS) and Community Coefficient (CC), changed abruptly below and above the 400 m contour, suggesting a change of vegetation structure at this altitude, which was also clear from population distributions. The spatial turnover rate of species along the altitudinal gradient was calculated in two ways: as the Average turnover rate along the whole range of the gradient, and as the Zone turnover rate at individual altitudes. The overall rates calculated for five categories of populations: DBH > 10 cm, DBH >3 cm, all woody species, herb-layer, and total vegetation were - 0.0011 to - 0.0021 for PS, and - 0.0009 to - 0.0019 for CC. The calculated rates (PS basis) indicate that a 95% change in species composition is reached at 1120 to 620 m altitude. Similarly, the rates -0.0009 to -0.0019 (CC) correspond to 1410 - 680 m. The altitudinal range expected here for a 95% change agrees with the actual elevation of forest zonation in northwestern Kyushu. The average rate of both PS and CC in the herb-layer population was 1.56 times higher than the rate in the woody species population, showing a more rapid change in herb-layer population than in the woody ones along the gradient. The Zone turnover rates were higher at the 370-440 m belt than those below and above the belt. This coincided with the interchanging pattern in population distributions and the abrupt change in similarity at about 400 m above sea level. This may be due to the change in environmental conditions such as physiography and air humidity. In the diversity measurements, the species density per 100 m2 showed a gradual increase in the DBH >3 cm population but a constant level in the DBH >10 cm population along the whole range of the forest coenocline studied, while index values of S(100) and Shannon's H' showed decreasing trends in the same gradient with a few exceptionally high and low values.
Partitioning beta diversity in a subtropical broad- leaved forest of China
DOI:10.1890/07-1880.1
URL
PMID:19341137
[本文引用: 1]
The classical environmental control model assumes that species distribution is determined by the spatial variation of underlying habitat conditions. This niche-based model has recently been challenged by the neutral theory of biodiversity which assumes that ecological drift is a key process regulating species coexistence. Understanding the mechanisms that maintain biodiversity in communities critically depends on our ability to decompose the variation of diversity into the contributions of different processes affecting it. Here we investigated the effects of pure habitat, pure spatial, and spatially structured habitat processes on the distributions of species richness and species composition in a recently established 24-ha stem-mapping plot in the subtropical evergreen broad-leaved forest of Gutianshan National Nature Reserve in East China. We used the new spatial analysis method of principal coordinates of neighbor matrices (PCNM) to disentangle the contributions of these processes. The results showed that (1) habitat and space jointly explained ~53% of the variation in richness and ~65% of the variation in species composition, depending on the scale (sampling unit size); (2) tree diversity (richness and composition) in the Gutianshan forest was dominantly controlled by spatially structured habitat (24%) and habitat-independent spatial component (29%); the spatially independent habitat contributed a negligible effect (6%); (3) distributions of richness and species composition were strongly affected by altitude and terrain convexity, while the effects of slope and aspect were weak; (4) the spatial distribution of diversity in the forest was dominated by broad-scaled spatial variation; (5) environmental control on the one hand and unexplained spatial variation on the other (unmeasured environmental variables and neutral processes) corresponded to spatial structures with different scales in the Gutianshan forest plot; and (6) five habitat types were recognized; a few species were statistically significant indicators of three of these habitats, whereas two habitats had no significant indicator species. The results suggest that the diversity of the forest is equally governed by environmental control (30%) and neutral processes (29%). In the fine-scale analysis (10 10 m cells), neutral processes dominated (43%) over environmental control (20%).
Partitioning the variation of woody plant β-diversity in a landscape of secondary tropical dry forests across spatial scales
DOI:10.1111/j.1654-1103.2012.01446.x
URL
[本文引用: 1]
QuestionWhat is the relative importance of forest successional age, environmental heterogeneity, landscape structure and spatial structure of sampling sites on β-diversity of tropical dry forests (TDF)? How do the magnitude of β-diversity and the relative influence of factors, processes and mechanisms driving β-diversity differ at different spatial grains? What are the effects of stand age on β-diversity?LocationYucatán Peninsula, México.MethodsFloristic composition was obtained from a hierarchical survey performed in 276 sites distributed across 23 sampling landscapes (SL) (12 sites per SL). Land-cover classes were derived from the classification of multi-spectral SPOT 5 satellite imagery. We calculated landscape metrics of patch type for each SL, and characterized local soil conditions for each sampling site. A principal coordinates of neighbour matrices (PCNM) analysis was performed to estimate spatial variables, and partial redundancy analysis was used to decompose variation into spatial, stand age and landscape structure components.ResultsThe magnitude of β-diversity varied with spatial scale (grain size), and was larger at the local than at the landscape grain. The magnitude of β-diversity also decreased slightly but significantly with successional age. There were significant differences in species composition among vegetation classes. Environmental factors (local soil conditions, as well as landscape structure) and spatial structure both contributed to woody plant β-diversity in our TDF landscape, but their relative importance was scale-dependent. At the local grain size, both the environment (mainly soil conditions) and the spatial structure strongly affected β-diversity, while at the landscape grain, environmental factors (variation in soil conditions, as well as landscape configuration) played a more prominent role.ConclusionsThe magnitude of β-diversity decreased with increasing spatial grain and successional age, while the relative importance of mechanisms influencing β-diversity was scale-dependent: both niche partitioning and dispersal limitation affect β-diversity at the local grain size, while niche partitioning prevails at the landscape grain.
Prelimarily discussion the influence factor and protection of forest biodiversity
DOI:10.3969/j.issn.1003-2630.2006.04.013
URL
[本文引用: 1]
森林是人类赖以生存的重要资源之一.是陆地生态系统中分布范围最广.生物总量最大的植被类型,是生物多样性最丰富的地方。本文通过对有关资料的查阅.简要论述了森林生物多样性的意义,并初步分析了影响森林生物多样性的因素,提出了相应的保护措施。
浅谈森林生物多样性的影响因素及其保护
DOI:10.3969/j.issn.1003-2630.2006.04.013
URL
[本文引用: 1]
森林是人类赖以生存的重要资源之一.是陆地生态系统中分布范围最广.生物总量最大的植被类型,是生物多样性最丰富的地方。本文通过对有关资料的查阅.简要论述了森林生物多样性的意义,并初步分析了影响森林生物多样性的因素,提出了相应的保护措施。
The pattern of plant species diversity of Funiu Mountain Nature Reserve
在伏牛山自然保护区典型地段设立样方,通过群落调查和多样性分析对物种多样性的垂直分布格局进行研究。发现:α多样性,乔木层在中海拔的落叶阔叶混交林带最高,灌木层随海拔升高稍呈上升趋势,草本层在低海拔次生林带和山顶矮林带物种多样性较高;β多样性总体上呈“~”型变化,在中海拔建群种发生更替的落叶阔叶混交林区最高,物种更替速率最快,其次是山顶附近。γ多样性大致沿海拔升高递减,以700m以下和过渡带落叶阔叶混交林区总物种丰富度最高。沿海拔梯度升高,水热组合发生变化,地带性植被总体上表现为由栓皮栎(Quercus variabilis)林向锐齿栎(Quercus aliena var. acuteserrata)林更替,在建群种发生更替的中海拔过渡带形成混交林带,短柄枹(Quercus glandulifera var. brevipetiolata)、槲栎(Quercus aliena)等夹杂其间在一些小生境中形成优势种群。总体上体现了过渡带具有高的物种多样性和更替速率,总物种丰富度隐含着沿海拔升高而降低的负相关关系。;We established experimental plots in representative areas in the Funiu Mountain nature reserve. We investigated plant communities in these plots to discern the pattern of species diversity distribution along the altitudinal gradient. We calculated α-diversity, β-diversity, and γ-diversity using the data thus collected. Alpha diversity of the tree layer peaks in the deciduous broadleaved mixed forest zone at mid-elevation, alpha diversity of the shrub layer increases gradually along the altitudinal gradient, while alpha diversity of the herb layer peaks in low altitude secondary forests and the mountaintop elfin forest. Beta diversity peaks twice, first at mid-elevations where the composition of edificatory species changes, then in areas near the mountaintop. Gamma diversity decreases along the altitudinal gradient. The highest species richness levels occur in areas below 700m and the mid-elevation deciduous broadleaved mixed forest. As elevation rises, Quercus variabilis forest gradually gives way to Quercus aliena var. acuteserrata forest, forming a mixed forest at mid-elevations; dispersed among the edificatory trees are Quercus glandulifera var. brevipetiolata and Quercus aliena, secondary edificatory species that are dominant in some niches. The transition zone is characterized by relatively high levels of species diversity and species turn-over rate. Overall species richness is negatively correlated with altitude.
伏牛山自然保护区物种多样性分布格局
在伏牛山自然保护区典型地段设立样方,通过群落调查和多样性分析对物种多样性的垂直分布格局进行研究。发现:α多样性,乔木层在中海拔的落叶阔叶混交林带最高,灌木层随海拔升高稍呈上升趋势,草本层在低海拔次生林带和山顶矮林带物种多样性较高;β多样性总体上呈“~”型变化,在中海拔建群种发生更替的落叶阔叶混交林区最高,物种更替速率最快,其次是山顶附近。γ多样性大致沿海拔升高递减,以700m以下和过渡带落叶阔叶混交林区总物种丰富度最高。沿海拔梯度升高,水热组合发生变化,地带性植被总体上表现为由栓皮栎(Quercus variabilis)林向锐齿栎(Quercus aliena var. acuteserrata)林更替,在建群种发生更替的中海拔过渡带形成混交林带,短柄枹(Quercus glandulifera var. brevipetiolata)、槲栎(Quercus aliena)等夹杂其间在一些小生境中形成优势种群。总体上体现了过渡带具有高的物种多样性和更替速率,总物种丰富度隐含着沿海拔升高而降低的负相关关系。;We established experimental plots in representative areas in the Funiu Mountain nature reserve. We investigated plant communities in these plots to discern the pattern of species diversity distribution along the altitudinal gradient. We calculated α-diversity, β-diversity, and γ-diversity using the data thus collected. Alpha diversity of the tree layer peaks in the deciduous broadleaved mixed forest zone at mid-elevation, alpha diversity of the shrub layer increases gradually along the altitudinal gradient, while alpha diversity of the herb layer peaks in low altitude secondary forests and the mountaintop elfin forest. Beta diversity peaks twice, first at mid-elevations where the composition of edificatory species changes, then in areas near the mountaintop. Gamma diversity decreases along the altitudinal gradient. The highest species richness levels occur in areas below 700m and the mid-elevation deciduous broadleaved mixed forest. As elevation rises, Quercus variabilis forest gradually gives way to Quercus aliena var. acuteserrata forest, forming a mixed forest at mid-elevations; dispersed among the edificatory trees are Quercus glandulifera var. brevipetiolata and Quercus aliena, secondary edificatory species that are dominant in some niches. The transition zone is characterized by relatively high levels of species diversity and species turn-over rate. Overall species richness is negatively correlated with altitude.
Studies on species diversity of forest vegetation on Qingcheng Mountain
DOI:10.3969/j.issn.0490-6756.2002.01.025
URL
[本文引用: 1]
以各个植被样地为对象 ,采用物种丰富度、Simpson指数、Shannon wiener指数及Pielou指数 ,研究了青城前山森林植被的物种多样性 .结果表明 :(1)在对青城山森林植被样地的物种多样性进行测度时 ,上述 4种指标表现出相同的变化趋势 .(2 )不同层次的物种多样性的大小依次是 :灌木层 ,乔木层和草本层 .(3)不同植被类型物种多样性的大小是常绿阔叶林大于暖性针叶林 .(4)人为因子的干扰对青城山森林植被的物种多样性有较大的影响 .
青城山森林植被物种多样性的研究
DOI:10.3969/j.issn.0490-6756.2002.01.025
URL
[本文引用: 1]
以各个植被样地为对象 ,采用物种丰富度、Simpson指数、Shannon wiener指数及Pielou指数 ,研究了青城前山森林植被的物种多样性 .结果表明 :(1)在对青城山森林植被样地的物种多样性进行测度时 ,上述 4种指标表现出相同的变化趋势 .(2 )不同层次的物种多样性的大小依次是 :灌木层 ,乔木层和草本层 .(3)不同植被类型物种多样性的大小是常绿阔叶林大于暖性针叶林 .(4)人为因子的干扰对青城山森林植被的物种多样性有较大的影响 .
Measurement of biotic community diversity. I. The measurement of α diversity
生物群落多样性的测度方法Ⅰα多样性的测度方法(上)马克平(中国科学院植物研究所,北京100044)1引言生物群落是在一定地理区域内,生活在同一环境下的不同种群的集合体,其内部存在着极为复杂的相互联系。由于功能整合原理,即系统的加合性或突生性(emer...
生物群落多样性的测度方法. I. α多样性的测度方法(上)
生物群落多样性的测度方法Ⅰα多样性的测度方法(上)马克平(中国科学院植物研究所,北京100044)1引言生物群落是在一定地理区域内,生活在同一环境下的不同种群的集合体,其内部存在着极为复杂的相互联系。由于功能整合原理,即系统的加合性或突生性(emer...
Measurement of biotic community diversity. II. The measurement of β diversity
生物群落多样性的测度方法. II. β多样性的测度方法
Effects of geographical distance and differences in climate and altitude on species dissimilarity of vascular plant communities in the Dulongjiang River Watershed Area
地理距离、海拔和气候差异对独龙江流域维管植物群落物种空间相异性的影响
Wetland management in New Zealand: Are current approaches and policies sustaining wetland ecosystems in agricultural landscapes?
DOI:10.1016/j.ecoleng.2012.12.097
URL
[本文引用: 1]
As a signatory to the Convention on Biological Diversity and to the Ramsar Convention on Wetlands, New Zealand has international responsibilities to protect and restore wetland ecosystems. The New Zealand Biodiversity Strategy also reflects New Zealand's commitment to help stem the loss of biodiversity worldwide, including wetlands.Wetland loss in New Zealand has been more significant than in most parts of the world, and ecosystems in fertile lowlands have been most severely impacted by agricultural development. Wetlands provide important ecosystem services filtering nutrients and controlling floodwaters but they are under continued pressure from agricultural land use, including drainage, grazing, nutrient runoff, and the impacts of pest animals and plants.Legislation in New Zealand identifies the protection of wetlands as a matter of national importance, and the protection of wetlands on private land has been identified as a national priority for action. While most of the larger nationally and internationally significant wetlands in New Zealand are in public ownership, the vast majority of smaller wetlands, which contribute to the full diversity of lowland ecosystems in New Zealand, are on private land in agricultural landscapes.Regional and district councils have responsibilities to implement legislation and develop policies and regulations to protect wetlands and prevent their damage and degradation. Most use a mix of regulatory mechanisms and voluntary incentives to encourage protection and restoration of wetlands. The strength of regulation for wetland protection varies across the country, with stronger more restrictive rules in more populated regions and where loss in extent has been more significant. While all regional plans have some form of rule restricting damaging activities in wetlands, less than half have strong regulations where drainage is non-compliant, and monitoring is sparse. The majority of plans (60%) restrict damaging activities only in wetlands that are in a schedule or meet criteria for ecological significance; rules in most plans do not protect smaller, often degraded wetlands.Although wetland loss and degradation still occurs in many regions, national and regional rates of loss are not reported. A response requires strong national policies on preventing further loss, the implementation of regulations in regional and district plans, and monitoring of the effectiveness of policies, rules, and non-statutory mechanisms. A combination of bottom lines for statutory regulation, voluntary incentives including support for fencing, and effective practical management is required. (C) 2013 Elsevier B.V. All rights reserved.
Community assembly: The relative importance of neutral theory and niche theory
DOI:10.3724/SP.J.1003.2009.09142
URL
[本文引用: 1]
物种共存和生物多样性维持一直是生态学研究的中心论题。基于物种生态位分化的群落构建理论已经发展了近一个世纪,但我们对群落构建和生物多样性维持的机理仍然不清楚。近年来,群落中性理论以其简约性和预测能力成为群落生态学研究的焦点,但由于其"物种在生态功能上等价"的假设与大量研究结果相悖,同时对自然群落结构的准确预测也只限于少数的生态系统,因而饱受质疑。如今,越来越多的生态学家认为群落构建的生态位理论与中性理论之争的最终归宿应该是二者的整合。在本文中,我们在简要回顾生态位理论和群落中性理论发展的基础上,分析二者之间的主要分歧和互补性,试图梳理二者整合的途径。我们认为,尽管中性理论的发展极大地丰富了群落构建理论,但二者的整合尚处于初级阶段;群落构建零模型假说、中性—生态位连续体假说、随机生态位假说等都不失为有价值的尝试,今后需要在其他类型的生态系统中进行实验验证,以更好地理解确定性过程和随机过程在决定群落构建和生物多样性维持中的作用。
群落构建的中性理论和生态位理论
DOI:10.3724/SP.J.1003.2009.09142
URL
[本文引用: 1]
物种共存和生物多样性维持一直是生态学研究的中心论题。基于物种生态位分化的群落构建理论已经发展了近一个世纪,但我们对群落构建和生物多样性维持的机理仍然不清楚。近年来,群落中性理论以其简约性和预测能力成为群落生态学研究的焦点,但由于其"物种在生态功能上等价"的假设与大量研究结果相悖,同时对自然群落结构的准确预测也只限于少数的生态系统,因而饱受质疑。如今,越来越多的生态学家认为群落构建的生态位理论与中性理论之争的最终归宿应该是二者的整合。在本文中,我们在简要回顾生态位理论和群落中性理论发展的基础上,分析二者之间的主要分歧和互补性,试图梳理二者整合的途径。我们认为,尽管中性理论的发展极大地丰富了群落构建理论,但二者的整合尚处于初级阶段;群落构建零模型假说、中性—生态位连续体假说、随机生态位假说等都不失为有价值的尝试,今后需要在其他类型的生态系统中进行实验验证,以更好地理解确定性过程和随机过程在决定群落构建和生物多样性维持中的作用。
vegan: Community Ecology Package
Forest vegetation of the Colorado Front Range: Patterns of species diversity
DOI:10.1007/BF00126830
URL
[本文引用: 1]
Plant species diversity patterns of the Rocky Mountain forests were found to be at variance with patterns reported from other regions. The most centrally located forests in terms of elevation, site...
Phylogenetic structure and phylogenetic diversity of angiosperm assemblages in forests along an elevational gradient in Changbaishan, China
DOI:10.1093/jpe/rtt072 URL [本文引用: 1]
Effects of geographic distance and climatic dissimilarity on species turnover in alpine meadow communities across a broad spatial extent on the Tibetan Plateau
DOI:10.1007/s11258-012-0095-4
URL
[本文引用: 1]
AbstractUnderstanding the underlying mechanisms that generate species turnover or beta diversity among biological communities is a central theme in ecology. Here, we distinguish the effects of geographic distance and climatic dissimilarity on species turnover of vascular plants in alpine meadow communities on the Tibetan Plateau in China. We calculated species turnover between each pair of 17 sites, using the Jaccard’s and Simpson’s indices. We selected six variables to quantify climate at each site, and subjected values of the climatic variables to a principal component analysis. We applied a variance partitioning approach to disentangle the effects of geographic distance and climatic dissimilarity on species turnover in alpine meadow communities. We also examined the effect of elevation variation on species turnover. Geographic distance and climate dissimilarity together explained 49.102% of the variation in compositional difference between alpine meadow communities; the amount of the variation explained purely by geographic distance and purely by climatic dissimilarity was 6.802% and 2.802%, respectively. When geographic distance, climate dissimilarity, and elevation difference were included in an analysis, they together explained 5502% of the variation in compositional difference between alpine meadow communities; the pure effect of each of the three sets of explanatory variables was 4.8, 4.3, and 3.502%, respectively. The fact that the vast majority of the variation explained by geographic distance and climatic dissimilarity cannot be independently attributed to either factor suggests that the two factors operate together in determining regional patterns of species composition in alpine meadows on the Tibetan Plateau.
Community structures and biodiversities of broad-leaved forest and two types of plantations in Gutianshan National Nature Reserve, Zhejiang Province
古田山自然保护区阔叶林与两种人工林的群落结构和生物多样性
The relationship among area, elevation, and regional species richness in neotropical birds
Phytools: An R package for phylogenetic comparative biology (and other things)
DOI:10.1111/j.2041-210X.2011.00169.x
URL
[本文引用: 1]
1. Here, I present a new, multifunctional phylogenetics package, phytools, for the R statistical computing environment.2. The focus of the package is on methods for phylogenetic comparative biology; however, it also includes tools for tree inference, phylogeny input/output, plotting, manipulation and several other tasks.3. I describe and tabulate the major methods implemented in phytools, and in addition provide some demonstration of its use in the form of two illustrative examples.4. Finally, I conclude by briefly describing an active web-log that I use to document present and future developments for phytools. I also note other web resources for phylogenetics in the R computational environment.
R: A language and environment for statistical computing
Perspective of the vegetation zonation of forest region in eastern China
DOI:10.3321/j.issn:1672-9072.1999.05.021
URL
[本文引用: 1]
简要回顾了中国东部森林植被带划分研究的历史及当前存在的争论。提出了中国东部植被带划分应以植被本身的特征,特别是地带性的生物群落集为主要依据,同时参照它们的区系组成和气候指标。根据上述原则将中国东部划分为6个植被带:北方针叶林带、凉温带针阔混交林带、温带落叶阔叶林带、暖温带常绿落叶阔叶混交林带、亚热带常绿阔叶林带和热带雨林、季雨林带,并对各植被带的特征作了简要的描述。阐述了对一些植被带名称、界线改动
中国东部森林植被带划分之我见
DOI:10.3321/j.issn:1672-9072.1999.05.021
URL
[本文引用: 1]
简要回顾了中国东部森林植被带划分研究的历史及当前存在的争论。提出了中国东部植被带划分应以植被本身的特征,特别是地带性的生物群落集为主要依据,同时参照它们的区系组成和气候指标。根据上述原则将中国东部划分为6个植被带:北方针叶林带、凉温带针阔混交林带、温带落叶阔叶林带、暖温带常绿落叶阔叶混交林带、亚热带常绿阔叶林带和热带雨林、季雨林带,并对各植被带的特征作了简要的描述。阐述了对一些植被带名称、界线改动
Spatial pattern analysis and topographical interpretation of species diversity in the forests of Dalaoling in the region of the Three Gorges
DOI:10.3321/j.issn:1672-9072.2000.06.014
URL
[本文引用: 1]
在对三峡大老岭地区森林的大量群落样方调查基础上 ,研究地形因子和取样尺度对群落不同层次α多样性水平及其空间格局的影响 ,结果表明 :①受地形、人为或自然干扰的影响 ,大老岭森林群落的α多样性存在海拔10 0 0m以下、海拔 110 0~ 170 0m和海拔 170 0m以上 3段不同的水平与变化特征 ;②对海拔 10 0 0m以上样方群落的 16种多样性指数的DCCA分析表明 ,群落乔、灌、草各层的多样性在多维地形因子梯度上表现出不同的格局 ;③本地地形因子对群落α多样性格局影响的大小顺序是 :坡位 海拔 坡向 坡面 坡度 坡形。④对海拔 10 5 0m和 16 70m处两个样方α多样性的多尺度对比分析表明 :高、低海拔的群落不同层次α多样性水平及其变化在各个尺度上差别明显。总之 ,地形对山地森林植物物种多样性格局显示了多方面和多尺度的影响 ;α多样性的海拔梯度格局在此未得到确证。
三峡大老岭森林物种多样性的空间格局分析及其地形解释
DOI:10.3321/j.issn:1672-9072.2000.06.014
URL
[本文引用: 1]
在对三峡大老岭地区森林的大量群落样方调查基础上 ,研究地形因子和取样尺度对群落不同层次α多样性水平及其空间格局的影响 ,结果表明 :①受地形、人为或自然干扰的影响 ,大老岭森林群落的α多样性存在海拔10 0 0m以下、海拔 110 0~ 170 0m和海拔 170 0m以上 3段不同的水平与变化特征 ;②对海拔 10 0 0m以上样方群落的 16种多样性指数的DCCA分析表明 ,群落乔、灌、草各层的多样性在多维地形因子梯度上表现出不同的格局 ;③本地地形因子对群落α多样性格局影响的大小顺序是 :坡位 海拔 坡向 坡面 坡度 坡形。④对海拔 10 5 0m和 16 70m处两个样方α多样性的多尺度对比分析表明 :高、低海拔的群落不同层次α多样性水平及其变化在各个尺度上差别明显。总之 ,地形对山地森林植物物种多样性格局显示了多方面和多尺度的影响 ;α多样性的海拔梯度格局在此未得到确证。
A review on the elevational patterns of plant species diversity
DOI:10.3321/j.issn:1005-0094.2004.01.004
URL
[本文引用: 1]
生物多样性沿环境梯度的变化趋势是生物多样性研究的一个重要议题,而海拔梯度包含了多种环境因子的梯度效应,因此研究生物多样性的海拔梯度格局对于揭示生物多样性的环境梯度变化规律具有重要意义。在不同的研究尺度,植物多样性沿海拔梯度具有不同的分布格局,而形成这种格局的因素有很大差异。本文从α多样性,β多样性和γ多样性三个尺度总结了植物物种多样性沿海拔梯度分布格局及其环境解释。α多样性沿海拔梯度的分布格局在不同生活型的物种之间差异很大,但对于木本植物来说,虽然也存在其他格局,但α多样性随海拔升高而降低是被广泛接受的一种格局。在一般情况下,β多样性随着海拔的升高而降低,并且对于不同生活型的物种,β多样性沿海拔梯度具有相似的分布格局。γ多样性沿海拔梯度具有两种分布格局: 偏峰分布格局和显著的负相关格局; 特有物种数往往随着海拔的升高而减少,而特有度则随着海拔的升高而增加。
植物物种多样性的垂直分布格局
DOI:10.3321/j.issn:1005-0094.2004.01.004
URL
[本文引用: 1]
生物多样性沿环境梯度的变化趋势是生物多样性研究的一个重要议题,而海拔梯度包含了多种环境因子的梯度效应,因此研究生物多样性的海拔梯度格局对于揭示生物多样性的环境梯度变化规律具有重要意义。在不同的研究尺度,植物多样性沿海拔梯度具有不同的分布格局,而形成这种格局的因素有很大差异。本文从α多样性,β多样性和γ多样性三个尺度总结了植物物种多样性沿海拔梯度分布格局及其环境解释。α多样性沿海拔梯度的分布格局在不同生活型的物种之间差异很大,但对于木本植物来说,虽然也存在其他格局,但α多样性随海拔升高而降低是被广泛接受的一种格局。在一般情况下,β多样性随着海拔的升高而降低,并且对于不同生活型的物种,β多样性沿海拔梯度具有相似的分布格局。γ多样性沿海拔梯度具有两种分布格局: 偏峰分布格局和显著的负相关格局; 特有物种数往往随着海拔的升高而减少,而特有度则随着海拔的升高而增加。
Elevational gradients in β-diversity reflect variation in the strength of local community assembly mechanicsms across spatial scales
DOI:10.1371/journal.pone.0121458
URL
[本文引用: 1]
Despite long-standing interest in elevational-diversity gradients, little is known about the processes that cause changes in the compositional variation of communities (β-diversity) across elevations. Recent studies have suggested that β-diversity gradients are driven by variation in species pools, rather than by variation in the strength of local community assembly mechanisms such as dispersal limitation, environmental filtering, or local biotic interactions. However, tests of this hypothesis have been limited to very small spatial scales that limit inferences about how the relative importance of assembly mechanisms may change across spatial scales. Here, we test the hypothesis that scale-dependent community assembly mechanisms shape biogeographic β-diversity gradients using one of the most well-characterized elevational gradients of tropical plant diversity. Using an extensive dataset on woody plant distributions along a 4,000-m elevational gradient in the Bolivian Andes, we compared observed patterns of β-diversity to null-model expectations. β-deviations (standardized differences from null values) were used to measure the relative effects of local community assembly mechanisms after removing sampling effects caused by variation in species pools. To test for scale-dependency, we compared elevational gradients at two contrasting spatial scales that differed in the size of local assemblages and regions by at least an order of magnitude. Elevational gradients in β-diversity persisted after accounting for regional variation in species pools. Moreover, the elevational gradient in β-deviations changed with spatial scale. At small scales, local assembly mechanisms were detectable, but variation in species pools accounted for most of the elevational gradient in β-diversity. At large spatial scales, in contrast, local assembly mechanisms were a dominant force driving changes in β-diversity. In contrast to the hypothesis that variation in species pools alone drives β-diversity gradients, we show that local community assembly mechanisms contribute strongly to systematic changes in β-diversity across elevations. We conclude that scale-dependent variation in community assembly mechanisms underlies these iconic gradients in global biodiversity.
Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlan
DOI:10.1046/j.1365-2745.1998.00325.x
URL
[本文引用: 1]
1 Data on the composition, structure and diversity of plant communities were gathered along a 1000-m altitudinal transect from tropical seasonal dry forest to cloud forest on calcareous Cerro Grande in Jalisco, México.
Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China
DOI:10.1023/A:1022236115186
URL
[本文引用: 1]
We studied the distribution pattern, species diversity and life-form spectra of plant communities along an altitudinal gradient in the mid-section of the northern slopes of Qilianshan Mountains by means of multivariate analyses. Two data sets (167 species 75 plots, 10 environmental variables 75 plots), originated from the fieldworks in 1998-1999, were subjected to TWINSPAN and DCCA, resulting in 8 major plant communities: 1) Asterothamnus centraliasiaticus-Halogeton arachnoideus desert grassland on azonal substrates from 1450 to 1600 m and 2) zonal Reaumuria soogorica desert grassland on gravels from 1470 to 1900 m; 3) Stipa przewalskii-Stipa purpurea montane grassland from 2200 to 2900 m; 4) Polygonum viviparum alpine grassland from 2900 to 3700 m; 5) Caragana stenophylla-Ajania fruticulosa dry-warm shrubland from 2350 to 2800 m; 6) Sabina przewalskii mid-wet warm forest from 2700 to 3300 m; 7) Picea crassifolia cold coniferous forest from 2450 to 3200 m; 8) Caragana jubata wet-cold alpine shrubland from 3100 to 3700 m. Species diversity and species richness of both grasslands and forests peaked at the intermediate portion of the elevational gradient. Evenness might be strongly influenced by either biotic or abiotic factors at a local scale, while seems quite independent of an elevational gradient at landscape scales. Beta-diversity decreased from 1500 to 3700 m, indicating that species turnover declined with increased elevation. Both richness of life-form and total species richness in a given altitudinal belt (gamma-diversity) peaked at intermediate elevations, while relative species richness of different life-form varied differently along the altitudinal gradient.
Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China
DOI:10.3321/j.issn:1005-0094.2004.01.010
URL
[本文引用: 1]
物种丰富度沿海拔梯度的分布格局成为生物多样性研究的热点.为探讨中尺度区域物种丰富度沿海拔梯度的分布,本文以高黎贡山为研究对象,利用该地区的地方植物志资料,结合通过GIS生成的区域数字高程模型(DEM)数据,分析了该区域全部种子植物和乔木、灌木、草本三种生活型种子植物物种丰富度的垂直分布格局以及物种密度沿海拔梯度的变化特征.结果表明:(1)全部种子植物和不同生活型植物物种丰富度随着海拔的升高呈现先增加后减小的趋势,最大值出现在海拔1500-2000m的范围;(2)物种密度与海拔也呈现单峰曲线关系;(3)物种丰富度和物种密度分布格局的形成主要受海拔所反映的水、热状况组合以及物种分布的边界影响.
高黎贡山种子植物物种丰富度沿海拔梯度的变化
DOI:10.3321/j.issn:1005-0094.2004.01.010
URL
[本文引用: 1]
物种丰富度沿海拔梯度的分布格局成为生物多样性研究的热点.为探讨中尺度区域物种丰富度沿海拔梯度的分布,本文以高黎贡山为研究对象,利用该地区的地方植物志资料,结合通过GIS生成的区域数字高程模型(DEM)数据,分析了该区域全部种子植物和乔木、灌木、草本三种生活型种子植物物种丰富度的垂直分布格局以及物种密度沿海拔梯度的变化特征.结果表明:(1)全部种子植物和不同生活型植物物种丰富度随着海拔的升高呈现先增加后减小的趋势,最大值出现在海拔1500-2000m的范围;(2)物种密度与海拔也呈现单峰曲线关系;(3)物种丰富度和物种密度分布格局的形成主要受海拔所反映的水、热状况组合以及物种分布的边界影响.
Taxonomic and phylogenetic α and β diversities of major subtropical forest community types in Zhejiang Province
DOI:10.17520/biods.2016114
URL
[本文引用: 4]
了解不同森林群落类型的物种和谱系水平的α和β多样性,有助于指导森林经营和生物多样性保护。本研究比较了浙江省内不同地点主要森林类型(包括常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林和针阔叶混交林)的物种α多样性和谱系α多样性,以及物种β多样性和谱系β多样性。研究表明,该地区主要森林类型的物种和谱系α多样性均存在较大差异,但控制了空间和地形因子的作用后,差异几乎全部消失;森林类型内部及相互间的物种和谱系β多样性均存在显著差异,同种森林类型内部的物种和谱系β多样性分别小于不同森林类型之间的物种和谱系β多样性,且在控制了空间和地形因子的作用后,以上差异仍然显著。本研究表明影响亚热带主要森林群落类型物种和谱系水平的α和β多样性的因素存在差异:α多样性可能主要受到空间和地形因子等的影响,而β多样性则可能受到森林类型的重要影响。
浙江省主要亚热带森林群落类型物种和谱系水平的α和β多样性比较
DOI:10.17520/biods.2016114
URL
[本文引用: 4]
了解不同森林群落类型的物种和谱系水平的α和β多样性,有助于指导森林经营和生物多样性保护。本研究比较了浙江省内不同地点主要森林类型(包括常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林和针阔叶混交林)的物种α多样性和谱系α多样性,以及物种β多样性和谱系β多样性。研究表明,该地区主要森林类型的物种和谱系α多样性均存在较大差异,但控制了空间和地形因子的作用后,差异几乎全部消失;森林类型内部及相互间的物种和谱系β多样性均存在显著差异,同种森林类型内部的物种和谱系β多样性分别小于不同森林类型之间的物种和谱系β多样性,且在控制了空间和地形因子的作用后,以上差异仍然显著。本研究表明影响亚热带主要森林群落类型物种和谱系水平的α和β多样性的因素存在差异:α多样性可能主要受到空间和地形因子等的影响,而β多样性则可能受到森林类型的重要影响。
Evolution and measurement of species diversity
DOI:10.2307/1218190 URL [本文引用: 1]
Vegetation of the Santa Catalina Mountains, Arizona: A gradient analysis of the south slope
DOI:10.2307/1934875
URL
[本文引用: 3]
Vegetation of the southwest slope of the Santa Catalina Mountains of southeastern Arizona was sampled and transects prepared for 1,000-ft (305 m) elevation belts on granite and gneiss soils from the summit forests (2,440-2,750 m) to the base of the mountains (900 m). Transects also represented subalpine forests above 2,750 m in the Pinaleno Mts. and vegetation of the valley plain or bajada below the mountains, and samples were taken from volcanic soils below 900 m in the Tucson Mts. Principal community-types from high elevations to low are: subalpine forest (Picca engelmanni in the Pinaleno Mts. and Abies lasiocarpa), montane fir forest (Abies concolor, Pseudotsuga menziesii), pine forests (Pinus ponderosa, P. strobiformis), pine-oak forests (P. ponderosa, Quercus hypoleucoides), pine-oak woodlands (P. ponderosa, P. chihuahuana, Q. hypoleucoides, Q. arizonica), pygmy conifer-oak scrub (Pinus cembroides, Juniperus deppeana, Q. arizonica, Q. emoryi, Arctostaphylos pringlei, A. pungens, monocot shrubs), open oak woodland (Q. emoryi, Q. oblongifolia, Vauquelinia californica, monocot shrubs, and grasses), desert-grassland (Agave schottii, Haplopappus laricifolius, and grasses), Sonora desert of mountain slopes (north-slope shrub phase, and south-slope spinose-suffrutescent phase), upper bajada desert (Cercidium microphyllum, Franseria deltoidea), and lower bajada desert (Larrea tridentata). Forests of canyons and arroyos are also described. Relations of communities to elevation and topograhic moisture gradients are represented in a mosaic chart. Physiognomic relations of communities are represented in charts of growth-form coverage in relation to elevation and topographic moisture gradients. Growth-form diversity increases from high-elevation forests strongly dominated by evergreen-needleleaf trees to desert of lower mountain slopes in which pinnate leguminous trees, spinose shrubs, suffrutescent semi-shrubs, and stem-succulents share dominance. Among Raunkiaer life-forms hemicryptophyte species are most numerous at middle and higher elevations, phanerophyte species at lower elevations. In open oak woodlands and desert grasslands phanerophytes, hemicryptophytes, and suffrutescent chamaephytes each make up about one-third of the perennial flora. Desert floras of mountain slopes are characterized by predominance of suffrutescent chamaephytes over both phanerophytes and hemicryptophytes, and large numbers of therophyte species. Analysis in terms of geographic areas of species shows decreasing numbers of Rocky Mountain, Western, and Northern species from high-elevation forests downward, increasing numbers of Southwestern and Latin American species at lower elevations. Madrean species of the Mexican Plateau and Southwestern species predomonate in pine-oak forests and woodlands and pygmy conifer-oak scrub. Sonoran, Chichuahuan, and Latin American species predominate in the desert of lower mountain slopes, and widely distributed Southwestern species in the Larrea desert. Flora of the Catalina Mountains is rich and community species diversities are high. Species diversities increase toward lower elevations desert-grasslands and deserts of lower mountain slopes are among the riches communities in the United States. Floristic diversity is higher in continental than maritime climates, as indicated by comparison of species diversities and community differentiation along topographic moisture gradients in the Catalinas and in California mountains.
Limitations to species coexistence: Evidence for competition from field observations, using a patch model
DOI:10.2307/3236236
URL
[本文引用: 1]
Abstract. Plant community structure in four dune slacks is examined for evidence of competition, expressed as local constraint on species biomass. Such constraint would result in a low variance in total quadrat biomass, compared to a null model. A method of analysis is introduced, using a patchbased null model, which is intended to discount most of the effects of environmental heterogeneity. The method is applied to data in which previous methods had failed to find such structure.
Diversity, diversity indices and tropical cockroaches
DOI:10.1007/BF00385226
URL
PMID:28310325
[本文引用: 1]
The diversity of samples of cockroaches (Blattaria) taken with light-traps in six localities in Panama is described. As a diversity index of the log series is found to be more satisfactory than either N2or N1of Hills's series or Hurlbert's Sm, even if the distribution of the relative abundances is significantly different from a log series. However, even the -index is of only limited usefulness. A simple statistic such as the number of species per unit effort can be at least as useful. The seasonal variation in the value of the diversity indices is negatively correlated with both the number of species and the number of individuals. Mountain sites have a lower alpha-diversity than lowland sites and relatively undisturbed sites have more individuals and more species than disturbed ones, although this difference is not reflected in the value of the diversity indices. To describe the beta-diversity the NESS similarity index, here called Cm, is far superior over any other indices available. The between site diversity is very high. Samples taken at the same site in successive years in most cases are significantly different from being random samples of the same fauna.
Species richness and environmental interpretation of pine and oak forest community in Qinling Mountains
DOI:10.16258/j.cnki.1674-5906.2017.06.004
URL
[本文引用: 1]
异质性环境是影响植物群落物种丰富度的重要因素。通过在秦岭中段(陕西省及甘肃省境内)设置45个样地,利用方差分析、典范对应分析等方法探讨研究区内锐齿槲栎(Quercusalienavar.acuteserrata)林、油松(Pinustabuliformis)-锐齿槲栎混交林和油松林3类群落的物种丰富度特征,旨在揭示环境因子(地形、气候等)对丰富度的影响,为深入认识秦岭森林群落演替规律及生物多样性保护提供理论依据。结果显示:(1)锐齿槲栎林记录植物215种,主要物种有锐齿槲栎、卫矛(Euonymusalatus)、木姜子(Litseapungens)和细叶苔草(Carexrigescens)等;松栎混交林记录植物142种,主要物种为锐齿槲栎、油松、悬钩子(Rubuscorchorifolius)和苔草(Carextristachya)等;油松林记录植物214种,主要物种为油松、胡枝子(Lespedezabicolor)、苦糖果(Lonicerastanishii)和牛尾蒿(Artemisiadubia)等;(2)锐齿槲栎群落不同层次的物种丰富度表现为灌木层>草本层>乔木层(F2,44=11.502,P乔木层(F2,44=10.019,P<0.001;F2,44=12.404,P<0.001),而灌、草层之间无显著差异;(3)决定锐齿槲栎群落、松栎混交林群落和油松群落CCA第1排序轴的主导性环境因子分别为海拔(r=0.696,P<0.01)、坡度(r=0.604,P<0.05)和纬度(r=0.580,P<0.05);(4)锐齿槲栎群落草本层丰富度随海拔上升而增加;随着坡度上升,松栎混交林乔木层丰富度增加而灌木层丰富度下降;油松群落乔木层丰富度随纬度上升而下降。该研究表明,秦岭中段典型森林群落物种丰富度对山地异质性生境的响应存在较大差异。
秦岭松栎林群落物种丰富度特征及其环境解释
DOI:10.16258/j.cnki.1674-5906.2017.06.004
URL
[本文引用: 1]
异质性环境是影响植物群落物种丰富度的重要因素。通过在秦岭中段(陕西省及甘肃省境内)设置45个样地,利用方差分析、典范对应分析等方法探讨研究区内锐齿槲栎(Quercusalienavar.acuteserrata)林、油松(Pinustabuliformis)-锐齿槲栎混交林和油松林3类群落的物种丰富度特征,旨在揭示环境因子(地形、气候等)对丰富度的影响,为深入认识秦岭森林群落演替规律及生物多样性保护提供理论依据。结果显示:(1)锐齿槲栎林记录植物215种,主要物种有锐齿槲栎、卫矛(Euonymusalatus)、木姜子(Litseapungens)和细叶苔草(Carexrigescens)等;松栎混交林记录植物142种,主要物种为锐齿槲栎、油松、悬钩子(Rubuscorchorifolius)和苔草(Carextristachya)等;油松林记录植物214种,主要物种为油松、胡枝子(Lespedezabicolor)、苦糖果(Lonicerastanishii)和牛尾蒿(Artemisiadubia)等;(2)锐齿槲栎群落不同层次的物种丰富度表现为灌木层>草本层>乔木层(F2,44=11.502,P乔木层(F2,44=10.019,P<0.001;F2,44=12.404,P<0.001),而灌、草层之间无显著差异;(3)决定锐齿槲栎群落、松栎混交林群落和油松群落CCA第1排序轴的主导性环境因子分别为海拔(r=0.696,P<0.01)、坡度(r=0.604,P<0.05)和纬度(r=0.580,P<0.05);(4)锐齿槲栎群落草本层丰富度随海拔上升而增加;随着坡度上升,松栎混交林乔木层丰富度增加而灌木层丰富度下降;油松群落乔木层丰富度随纬度上升而下降。该研究表明,秦岭中段典型森林群落物种丰富度对山地异质性生境的响应存在较大差异。
The α diversity of shrubs community in the arid valley of the Minjiang River
DOI:10.3321/j.issn:1000-0933.2004.06.005
URL
[本文引用: 1]
通过对岷江干旱河谷植被及其环境因子的系统取样调查,研究了该地 区植物群落的a多样性及其与环境因子的关系.土壤、植被、地形三者之间的典范相关分析结果表明,三者两两之间均存在着较高的相关性.土壤因子与地形因子之 间,海拔和坡向起较大的作用,主要影响土壤中的全氮、有机质和土壤含水量;地形因子和植被之间,海拔和坡向影响灌木层的多样性和盖度;植被和土壤因子之 间,土壤中的全氮量、有机质和土壤含水量影响灌木层的盖度和多样性.随着海拔的增加,草本和灌木群落的多样性都呈现出先增加后减小而后又增加的趋势,草本 层的α多样性明显高于灌木层的a多样性,草本层和灌木层均在1400~1600m和2000~2200m两个海拔段有较高的α多样性;华帚菊-小黄素馨灌 丛、金花小檗-忍冬灌丛、绣线菊灌丛有着较高的多样性,西南野丁香灌丛、莸灌丛、小马鞍羊蹄甲-白刺花灌丛的群落多样性较低;样带的多样性,灌木层:样带 3>样带1>样带2,草本层:样带1>样带3>样带2;不同坡向的多样性,灌木层:阴坡>半阴半阳坡>阳坡,草本层:半阴半阳坡>阴坡>阳坡;不同坡形上 的多样性,无论是灌木层还是草本层,多样性大小为凹坡>平坡>凸坡;灌木层和草本层在不同坡位上的多样性大小均为上坡位>下坡位>中坡位.
岷江干旱河谷灌丛α多样性分析
DOI:10.3321/j.issn:1000-0933.2004.06.005
URL
[本文引用: 1]
通过对岷江干旱河谷植被及其环境因子的系统取样调查,研究了该地 区植物群落的a多样性及其与环境因子的关系.土壤、植被、地形三者之间的典范相关分析结果表明,三者两两之间均存在着较高的相关性.土壤因子与地形因子之 间,海拔和坡向起较大的作用,主要影响土壤中的全氮、有机质和土壤含水量;地形因子和植被之间,海拔和坡向影响灌木层的多样性和盖度;植被和土壤因子之 间,土壤中的全氮量、有机质和土壤含水量影响灌木层的盖度和多样性.随着海拔的增加,草本和灌木群落的多样性都呈现出先增加后减小而后又增加的趋势,草本 层的α多样性明显高于灌木层的a多样性,草本层和灌木层均在1400~1600m和2000~2200m两个海拔段有较高的α多样性;华帚菊-小黄素馨灌 丛、金花小檗-忍冬灌丛、绣线菊灌丛有着较高的多样性,西南野丁香灌丛、莸灌丛、小马鞍羊蹄甲-白刺花灌丛的群落多样性较低;样带的多样性,灌木层:样带 3>样带1>样带2,草本层:样带1>样带3>样带2;不同坡向的多样性,灌木层:阴坡>半阴半阳坡>阳坡,草本层:半阴半阳坡>阴坡>阳坡;不同坡形上 的多样性,无论是灌木层还是草本层,多样性大小为凹坡>平坡>凸坡;灌木层和草本层在不同坡位上的多样性大小均为上坡位>下坡位>中坡位.
Forest vegetation types in Gutianshan Natural Reserve in Zhejiang. Journal of Zhejiang University (Agriculture &
DOI:10.3321/j.issn:1008-9209.2001.04.007
URL
[本文引用: 1]
根据群落调查和统计分析结果 ,并参考《中国植被》分类原则 ,将古田山自然保护区的森林植被分为四种基本类型 :常绿阔叶林、常绿落叶阔叶混交林、针叶阔叶混交林和针叶林 ,并对各个植被类型进行了进一步的分类和描述 .古田山森林植被从外貌、结构和种类组成上看 ,均具有我国典型常绿阔叶林的基本特征 ,在中部亚热带低海拔地区较为罕见 .植被的垂直分带现象明显 ,同时在同一海拔的不同地段植被类型有较大变化
浙江古田山自然保护区森林植被类型
DOI:10.3321/j.issn:1008-9209.2001.04.007
URL
[本文引用: 1]
根据群落调查和统计分析结果 ,并参考《中国植被》分类原则 ,将古田山自然保护区的森林植被分为四种基本类型 :常绿阔叶林、常绿落叶阔叶混交林、针叶阔叶混交林和针叶林 ,并对各个植被类型进行了进一步的分类和描述 .古田山森林植被从外貌、结构和种类组成上看 ,均具有我国典型常绿阔叶林的基本特征 ,在中部亚热带低海拔地区较为罕见 .植被的垂直分带现象明显 ,同时在同一海拔的不同地段植被类型有较大变化
[
Effects of landforms on α biodiversity in slope scale
DOI:10.3969/j.issn.1674-5906.2009.06.047
URL
[本文引用: 2]
为了解地貌在坡面尺度上对α生物多样性的影响,采用主观采样法在陕北吴起县合家沟流域不同地貌部位进行了样地调查。利用SPSS16.0统计软件先后对各地貌部位物种组成及各物种的重要值、地形因子要素间、地形因子和群落Ⅱ多样性之间分别做了聚类分析、相关分析和多元回归分析。结果表明:(1)地貌部位相似的群落聚类在一起,说明地形因子是影响物种组成、群落结构、生态系统等的重要因素。(2)海拔和坡位,坡向和坡度,地形指数和海拔、坡位、坡形之间的Pearson相关系数均大于0.8,双尾显著性检验概率小于0.05。(3)影响α生物多样性指标香农.维纳指数的地形因子按重要性从大到小依次是:坡位、坡向、海拔、坡形、坡度、地形指数,进一步分析得出在黄土高原丘陵沟壑区,沟沿线、光照、土壤水分和养分在影响Ⅱ生物多样性指标上依次递减。(4)通过多元线形回归检验,得出坡位、坡向、坡形、海拔这四个地形因子与群落α生物多样性关系密切,建立的回归模型显著性检验可信度大,与样本数据的拟合度高。各地形因子数据归一处理后的回归方程为:香农-维纳指数=2.417-0.581×坡形-1.333×坡位+1.449×海拔+0.631×坡向。地形地貌特征在黄土丘陵区表现明显,研究它对生物多样性的影响可为该区植被恢复提供参考,但由于调查样地尺度较小,在应用推广上尚待进一步研究。
坡面尺度上地貌对α生物多样性的影响
DOI:10.3969/j.issn.1674-5906.2009.06.047
URL
[本文引用: 2]
为了解地貌在坡面尺度上对α生物多样性的影响,采用主观采样法在陕北吴起县合家沟流域不同地貌部位进行了样地调查。利用SPSS16.0统计软件先后对各地貌部位物种组成及各物种的重要值、地形因子要素间、地形因子和群落Ⅱ多样性之间分别做了聚类分析、相关分析和多元回归分析。结果表明:(1)地貌部位相似的群落聚类在一起,说明地形因子是影响物种组成、群落结构、生态系统等的重要因素。(2)海拔和坡位,坡向和坡度,地形指数和海拔、坡位、坡形之间的Pearson相关系数均大于0.8,双尾显著性检验概率小于0.05。(3)影响α生物多样性指标香农.维纳指数的地形因子按重要性从大到小依次是:坡位、坡向、海拔、坡形、坡度、地形指数,进一步分析得出在黄土高原丘陵沟壑区,沟沿线、光照、土壤水分和养分在影响Ⅱ生物多样性指标上依次递减。(4)通过多元线形回归检验,得出坡位、坡向、坡形、海拔这四个地形因子与群落α生物多样性关系密切,建立的回归模型显著性检验可信度大,与样本数据的拟合度高。各地形因子数据归一处理后的回归方程为:香农-维纳指数=2.417-0.581×坡形-1.333×坡位+1.449×海拔+0.631×坡向。地形地貌特征在黄土丘陵区表现明显,研究它对生物多样性的影响可为该区植被恢复提供参考,但由于调查样地尺度较小,在应用推广上尚待进一步研究。
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}