荞麦是世界上广为种植的谷物类粮食作物, 因其具有药用价值, 现在人们开发了荞麦茶、荞麦酒等农副产品。人们俗称的荞麦是指栽培的两个种, 即甜荞和苦荞(图1 )。已有的证据表明甜荞(Fagopyrum esculentum )和苦荞(F. tataricum )是独立的两个种, 均起源于我国西南地区长江流域的上游。荞麦是唯一不属于禾本科的、世界主要的谷物类粮食作物; 其他谷物类是通过风媒或自花传粉, 而荞麦则是虫媒或自花传粉。另外, 有别于其他作物, 甜荞是二型花柱植物, 居群中有两种基因型不同的个体(Wu et al, 2018 )。中国被认为是最早栽培荞麦的国家, 而且荞麦属的大多数野生近缘物种分布在中国。因此荞麦为我国研究栽培植物的起源、散布和野生资源的利用、植物性系统的演化以及长江流域文明的起源提供了一个好的研究系统。本文试图对前人的研究工作进行小结, 提出值得深入研究的问题, 以期为我国的粮食安全、资源保护以及生态文明建设提供参考依据。
图1 栽培甜荞和苦荞的花和果实。(A)、(B)、(C)分别代表二型花柱甜荞的花序、长花柱花和短花柱花; (D)同型花柱的苦荞正在结实。
Fig. 1 Flowers and fruits in the two cultivated buckwheat species. (A), (B) and (C) indicate flowering individuals of sweet buckwheat (Fagopyrum esculentum ), a distylous crop, illustrating long-styled, and short-styled flower, respectively; (D) Fruiting individuals of bitter buckwheat (F. tartaricum ), a homostylous crop.
1 荞麦的栽培历史、种植现状及经济药用价值
对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》。前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 )。长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 )。另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 )。在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 )。自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 )。这些出土的实物、文字记载表明我国种植荞麦的历史悠久。
荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本。公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 )。在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 )。也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 )。据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 )。在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 )。据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 )。生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二。我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 )。由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用。荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物。
图2 世界范围内荞麦种植现状, 显示欧亚大陆普遍栽培, 数据来源于FAO (2014) 。
Fig. 2 Buckwheat planting status in the world, showing the prevalence of cultivation in Eurasia, data from FAO (2014)
近年来, 对荞麦理化性质、药用功效及营养价值的研究较多。苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 )。制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品。荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用。临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用。尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 )。Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因。荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 )。研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 )。荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 )。另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物。荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 )。此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 )。
2 荞麦的分类地位、系统关系及可能的起源地
2.1 分类地位的归并
荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum )。林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench)。Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 )。Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议。Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组。荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属。自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller。
《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道。Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum 。Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium 。Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense 。夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium )。刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum )。Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense )。Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai )。Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense )。Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense )。Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense )。目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位。
2.2 荞麦属种间系统关系
国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系。Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系。Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系。Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析。Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组。胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系。Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系。以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致。目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础。
2.3 栽培荞麦的起源及祖先
关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 )。Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方。根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部。后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富。近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 )。亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 )。甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 )。任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心。目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 )。
关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说。早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点。另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点。一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析。Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交。野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 )。野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 )。甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 )。形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 )。同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 )。野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 )。另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus )。Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区。甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 )。推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞。虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径。
3 荞麦属植物的繁殖生物学特征
荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 )。生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 )。荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天。花期一般为6-9月, 果期8-10月(Li & Hong, 2003 )。栽培荞麦在我国南方可以春秋两季播种。瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落。属的模式种为甜荞(Fagopyrum esculentum Moench)。
荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 )。二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置。甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实。与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 )。苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 )。另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小。据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 )。研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 )。利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础。
甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 )。较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 )。荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 )。
自交不亲和的甜荞依赖传粉者传递花粉才结实, 调查表明访花传粉昆虫是泛化的, 主要包括膜翅目(蜜蜂、熊蜂、独居蜂)、双翅目(食蚜蝇科、丽蝇科等), 另外还有鳞翅目、半翅目、脉翅目等其他昆虫(Jacquemart et al, 2007 ; Wu et al, 2017 )。在许多国家, 蜜蜂被认为是栽培荞麦最普遍且最主要的访花者(Jacquemart et al, 2007 )。但不同访花昆虫的传粉效率仍需进一步研究。
4 展望
荞麦是一种很好的药食同源植物。由于荞麦果实成熟后容易脱落的落粒特性, 结实率及产量低下, 对其开发和利用的空间还很大。甜荞两型花之间授粉才能结实, 而苦荞可以自交, 但即使是人工补充授粉, 甜荞的结实率都不到50%, 而苦荞结实率可达80% (作者未发表数据)。由于荞麦自身的生物学特性以及其他大宗作物的发展导致荞麦播种面积呈减少趋势, Jacquemart等(2012) 质疑了荞麦是否仍然是有前途的作物。为保障国家粮食安全, 开发新的主粮作物被我国政府提上日程。荞麦作为一种起源于中国的粮食作物, 与长江流域文明的发展密不可分。但是我们目前对其生物学特性、起源地, 如何提高荞麦产量和种植技术方面还有待深入研究, 培育出自交亲和的甜荞品种有利于提高甜荞的产量, 从而扩大其种植范围。该属具有二型和同型花柱物种, 为性系统演化的研究提供了好的模式系统。我们正在同国内基因组学的同行合作, 将在近期完成甜荞的全基因组测序, 为回答这一演化问题和挖掘优良的农艺性状奠定基础。
目前的研究主要集中在栽培的甜荞和苦荞, 对于我国西南丰富的野生荞麦资源研究甚少, 对于这些野生种质资源应该加以保护和利用。据研究野生金荞麦的根茎具有抑菌、抗炎和抗肿瘤等作用(赵佐成等, 2007 ), 我们2012-2015年在四川、云南进行野外调查时, 注意到大量野生资源遭到破坏, 有些种群已经消失。对二型花柱植物, 种群中两种表型的比例偏离1:1将影响有性繁殖, 加剧了地方种群的灭绝。对于这些珍贵的资源植物的家底需要调查并实行有效的保护措施, 避免陷入濒临灭绝的境地。对于荞麦属的分类地位及栽培荞麦的祖先种仍存在诸多争议, 在开展野外种质资源收集的同时, 需要厘清该属的系统发生与演化关系。
致谢: 感谢赵佐成教授给予荞麦项目的指导和大力帮助, 感谢陈家宽教授从南昌大学专项中支付本文的出版费。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Björkman T 1995 ) The effectiveness of heterostyly in preventing illegitimate pollination in dish-shaped flowers
. Sex Plant Reproduction , 8 , 143 -146 .
[本文引用: 1]
[2]
Boivin N Fuller DQ Crowther A 2012 ) Old World globalization and the Columbian exchange: Comparison and contrast
. World Archaeology , 44 , 452 -469 .
[本文引用: 1]
[3]
Campbell CG 1976 ) Buckwheat: Fagopyrum (Polygonaceae). In: Evolution of Crop Plants (ed. Simmonds NW) , pp. 235 -237 . Longman , London & New York .
[本文引用: 1]
[4]
Cawoy V Ledent JF Kinet JM Jacquemart AL 2009 ) Floral biology of common buckwheat (Fagopyrum esculentum Moench)
. The European Journal of Plant Science and Biotechnology , 3 , 1 -9 .
[本文引用: 2]
[5]
Chen QF 1999 ) A study of resources of Fagopyrum (Polygonaceae) native to China
. Botanical Journal of the Linnean Society , 130 , 53 -64 .
[本文引用: 2]
[6]
Chen QF 2001 ) Discussion on the origin of cultivated buckwheat in genus Fagopyrum (Polygonaceae). In: Proceedings of the 8th International Symposium on Buckwheat ,pp. 206 -213 , August,Chunchon , Republic of Korea .
[本文引用: 1]
[7]
Chen QF 2012 ) Plant Sciences on Genus Fagopyrum . Science Press , Beijing . (in Chinese)
[本文引用: 8]
[陈庆富 (2012 ) 荞麦属植物科学 . 科学出版社 , 北京 .]
[本文引用: 8]
[8]
Chen XR 1960 ) Excavation of Han Tombs in Mozuizi, Wuwei, Gansu Province
. Archaeology , (9 ), 15 -28 . (in Chinese)
[本文引用: 1]
[陈贤儒 (1960 ) 甘肃武威磨咀子汉墓发掘
. 考古 , (9 ), 15 -28 .]
[本文引用: 1]
[9]
Cho KS Yun BK Yoon YH Hong SY Mekapogu M Kim KH Yang TJ 2015 ) Complete chloroplast genome sequence of tartary buckwheat (Fagopyrum tataricum ) and comparative analysis with common buckwheat (F. esculentum )
. PLoS ONE , 10 , e0125332 .
[本文引用: 1]
[10]
Food and Agriculture Organization of the United Nations (FAO)
(2014 ) Production of Buckwheat by Countries . .
URL
[本文引用: 3]
[11]
Gondola I Papp PP 2010 ) Origin, geographical distribution and phylogenic relationships of common buckwheat (Fagopyrum esculentum Moench)
. Buckwheat , 2 , 17 -32 .
[本文引用: 1]
[12]
Gross MH 1913 ) Remarques sur les Polygonées de l’Asie Orientale
. Bulletin de Géographie Botanique , 23 , 7 -32 .
[本文引用: 2]
[13]
Hedberg O 1946 ) Pollen morphology in the genus Polygonum L. (s. lat.) and its taxonomical significance
. Svensk Botanisk Tidskrift , 40 , 371 -404 .
[本文引用: 2]
[14]
Hou LL Zhou ML Zhang Q Qi LP Yang XB Tang Y Zhu XM Shao JR 2015 ) Fagopyrum luojishanense , a new species of Polygonaceae from Sichuan, China
. Novon , 24 , 22 -26 .
[本文引用: 1]
[15]
Hu YN Zhang ZW Wu B Gao J Li YQ 2016 ) Genetic relationships of buckwheat species based on the sequence analysis of ITS and ndh F-rpl 32
. Biodiversity Science , 24 , 296 -303 . (in Chinese with English abstract)
[本文引用: 1]
[胡亚妮 , 张宗文 , 吴斌 , 高佳 , 李艳琴 (2016 ) 基于ITS和ndh F-rpl 32序列的荞麦种间亲缘关系分析
. 生物多样性 , 24 , 296 -303 .]
[本文引用: 1]
[16]
Jacquemart AL Cawoy V Kinet JM Ledent JF Quinet M 2012 ) Is buckwheat (Fagopyrum esculentum Moench) still a valuable crop today?
The European Journal of Plant Science and Biotechnology , 6 , 1 -10 .
[本文引用: 1]
[17]
Jacquemart AL Gillet C Cawoy V 2007 ) Floral visitors and the importance of honey bee on buckwheat (Fagopyrum esculentum Moench) in central Belgium
. The Journal of Horticultural Science and Biotechnology , 82 , 104 -108 .
[本文引用: 2]
[18]
Kishima Y Ogura K Mizukami K Mikami T Adachi T 1995 ) Chloroplast DNA analysis in buckwheat species: Phylogenetic relationships, origin of the reproductive systems and extended inverted repeats
. Plant Science , 108 , 173 -179 .
[本文引用: 1]
[19]
Kreft I 2001 ) Buckwheat research, past, present and future perspectives 20 years of internationaly coordinated research. In: Proceedings of the 8th International Symposium on Buckwheat, pp . 361 -366 , August, Chunchon , Republic of Korea .
[本文引用: 1]
[20]
Levent H Bilgiçli N 2011 ) Enrichment of gluten-free cakes with lupin (Lupinus albus L.) or buckwheat (Fagopyrum esculentum M.) flours
. International Journal of Food Sciences and Nutrition , 62 , 725 -728 .
[本文引用: 1]
[21]
Li AR Hong SP 2003 ) Fagopyrum
.2003 ) Fagopyrum .
URL
[本文引用: 4]
[22]
Li X Dodson J Zhou X Zhang H Masutomoto R 2007 ) Early cultivated wheat and broadening of agriculture in Neolithic China
. Holocene , 17 , 555 -560 .
[本文引用: 1]
[23]
Li YF 1979 ) Western Han Tombs in Maquan, Xianyang, Shaanxi Province
. Archaeology , (2 ), 125 -135 . (in Chinese)
[本文引用: 1]
[李毓芳 (1979 ) 陕西咸阳马泉西汉墓
. 考古 , (2 ), 125 -135 .]
[本文引用: 1]
[24]
Li YY Willis KJ Zhou LP Cui HT 2006 ) The impact of ancient civilization on the northeastern Chinese landscape: Palaeoecological evidence from the Western Liaohe River Basin, Inner Mongolia
. Holocene , 16 , 1109 -1121 .
[本文引用: 1]
[25]
Liu JL Tang Y Xia MZ Shao JR Cai GZ Luo Q Sun JX 2008 a) Fagopyrum crispatifolium J. L. Liu, a new species of Polygonaceae from Sichuan, China
. Journal of Systematics and Evolution , 46 , 929 -932 . (in Chinese with English abstract)
[本文引用: 1]
[刘建林 , 唐宇 , 夏明忠 , 邵继荣 , 蔡光泽 , 罗强 , 孙俊秀 (2008 a) 中国四川蓼科荞麦属一新种——皱叶野荞麦
. 植物分类学报 , 46 , 929 -932 .]
[本文引用: 1]
[26]
Liu JL Tang Y Xia MZ Shao JR Cai GZ Luo Q Sun JX 2008 b) Fagopyrum densovillosum J. L. Liu, a new species of Polygonaceae from Sichuan, China
. Bulletin of Botanical Research , 28 , 530 -533 . (in Chinese with English abstract)
[本文引用: 1]
[刘建林 , 唐宇 , 夏明忠 , 邵继荣 , 蔡光泽 , 罗强 , 孙俊秀 . (2008 b). 中国荞麦属(蓼科)一新种——密毛野荞麦
. 植物研究 , 28 , 530 -533 .]
[本文引用: 1]
[27]
Logacheva MD Kasianov AS Vinogradov DV Samigullin TH Gelfand MS Makeev VJ Penin AA 2011 ) De novo sequencing and characterization of floral transcriptome in two species of buckwheat (Fagopyrum )
. BMC Genomics , 12 , 30 .
[本文引用: 1]
[28]
Logacheva MD Samigullin TH Dhingra A Penin AA 2008 ) Comparative chloroplast genomics and phylogenetics of Fagopyrum esculentum ssp. ancestrale —a wild ancestor of cultivated buckwheat
. BMC Plant Biology , 8 , 59 .
[本文引用: 1]
[29]
Ma KH Kim NS Lee GA Lee SY Lee JK Yi JY Park YJ Kim TS Gwag JG Kwon SJ 2009 ) Development of SSR markers for studies of diversity in the genus Fagopyrum
. Theoretical and Applied Genetics , 119 , 1247 -1254 .
[本文引用: 1]
[30]
Matano T Ujihara A 1979 ) Agroecological classification and geographical distribution of the common buckwheat, Fagopyrum esculentum M. in the East Asia
. Japan Agricultural Research Quarterly , 13 , 157 -162 .
[本文引用: 1]
[31]
Murai M Ohnishi O 1996 ) Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench X. Diffusion routes revealed by RAPD markers
. Genes & Genetic Systems , 71 , 211 -218 .
[本文引用: 1]
[32]
Nakao S 1957 ) Transmittance of cultivated plants through the Sino-Himalayan route. In: Peoples of Nepal Himalaya (ed. Kihara H) , pp. 397 -420 . Fauna and Flora Research Society , Kyoto .
[本文引用: 1]
[33]
Nishimoto Y Ohnishi O Hasegawa M 2003 ) Topological incongruence between nuclear and chloroplast DNA trees suggesting hybridization in the urophyllum group of the genus Fagopyrum (Polygonaceae)
. Genes & Genetic Systems , 78 , 139 -153 .
[本文引用: 1]
[34]
Ohnishi O 1988 ) Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. VII. Allozyme variability in Japan, Korea, and China
. The Japanese Journal of Genetics , 63 , 507 -522 .
[本文引用: 2]
[35]
Ohnishi O 1993 ) Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. VIII. Local differentiation of land races in Europe and the silk road
. The Japanese Journal of Genetics , 68 , 303 -316 .
[本文引用: 1]
[36]
Ohnishi O 1995 ) Discovery of new Fagopyrum species and its implication for the studies of evolution of Fagopyrum and of the origin of cultivated buckwheat. In: Proceedings of the 6th International Symposium on Buckwheat ,pp. 175 -190 , August,Shinshu , Japan .
[本文引用: 6]
[37]
Ohnishi O 1998 ) Search for the wild ancestor of buckwheat. I. Description of new Fagopyrum (Polygonaceae) species and their distribution in China and the Himalayan hills
. Fagopyrum , 15 , 18 -28 .
[本文引用: 1]
[38]
Ohnishi O 2004 ) On the origin of cultivated buckwheat. In: Proceedings of the 9th International Symposium on Buckwheat ,pp. 18 -22 , August,Prague , Czech .
[本文引用: 3]
[39]
Ohnishi O Matsuoka Y 1996 ) Search for the wild progenitor of buckwheat. II. Taxonomy of Fagopyrum (Polygonaceae) species based on morphology, isozymes and cpDNA variability
. Genes & Genetic Systems , 71 , 383 -390 .
[本文引用: 1]
[40]
Ohsako T Ohnishi O 1998 ) New Fagopyrum species revealed by morphological and molecular analyses
. Genes & Genetic Systems , 73 , 85 -94 .
[本文引用: 1]
[41]
Ohsako T Ohnishi O 2000 ) Intra- and interspecific phylogeny of wild Fagopyrum (Polygonaceae) species based on nucleotide sequences of noncoding regions in chloroplast DNA
. American Journal of Botany , 87 , 573 -582 .
[本文引用: 1]
[42]
Ohsako T Yamane K Ohnishi O 2002 ) Two new Fagopyrum (Polygonaceae) species, F. gracilipedoides and F. jinshaense from Yunnan, China
. Genes & Genetic Systems , 77 , 399 -408 .
[本文引用: 1]
[43]
Ren CZ Zhao G 2015 ) Chinese Buckwheat Science . China Agriculture Press , Beijing . (in Chinese)
[本文引用: 5]
[任长忠 , 赵钢 (2015 ) 中国荞麦学 . 中国农业出版社 , 北京 .]
[本文引用: 5]
[44]
Shao JR Zhou ML Zhu XM Wang DZ Bai DQ 2011 ) Fagopyrum wenchuanense and Fagopyrum qiangcai , two new species of Polygonaceae from Sichuan, China
. Novon 21 , 256 -261 .
[本文引用: 1]
[45]
Sharma TR Jana S 2002 ) Species relationships in Fagopyrum revealed by PCR-based DNA fingerprinting
. Theoretical and Applied Genetics , 105 , 306 -312 .
[本文引用: 1]
[46]
Slawinska J Obendorf RL 2001 ) Buckwheat seed set in planta and during in vitro inflorescence culture: Evaluation of temperature and water deficit stress
. Seed Science Research , 11 , 223 -233 .
[本文引用: 1]
[47]
Steward AN 1930 ) The Polygoneae of Eastern Asia
. Contributions from the Gray Herbarium of Harvard University , 5 , 1 -129 .
[本文引用: 4]
[48]
Tang Y Ding MQ Tang YX Wu YM Shao JR Zhou ML 2016 ) Germplasm resources of buckwheat in China. In: Molecular Breeding and Nutritional Aspects of Buckwheat (eds Zhou ML, Kreft I, Woo SH, Chrungoo NK, Wieslander G) , pp. 13 -20 . Academic Press , London .
[本文引用: 2]
[49]
Tang Y Zhou ML Bai , DQ , Shao JR Zhu XM Wang DZ Tang YX 2010 ) Fagopyrum pugense (Polygonaceae), a new species from Sichuan, China
. Novon , 20 , 239 -242 .
[本文引用: 1]
[50]
Tsuji K Ohnishi O 2000 ) Origin of cultivated tartary buckwheat (Fagopyrum tataricum Gaertn.) revealed by RAPD analyses
. Genetic Resources and Crop Evolution , 47 , 431 -438 .
[本文引用: 1]
[51]
Tsuji K Ohnishi O 2001 ) Phylogenetic relationships among wild and cultivated tartary buckwheat (Fagopyrum tataricum Gaert.) populations revealed by AFLP analyses
. Genes & Genetic Systems , 76 , 47 -52 .
[本文引用: 1]
[52]
Wang CL Li ZQ Ding MQ Tang Y Zhu XM Liu JL Shao JR Zhou ML 2017 ) Fagopyrum longzhoushanense , a new species of Polygonaceae from Sichuan, China
. Phytotaxa , 291 , 73 -80 .
[本文引用: 1]
[53]
Wei YM 1995 ) Buckwheat production in China. In: Proceeding of the 6th International Symposium on Buckwheat ,pp. 24 -29 , August,Shinshu , Japan .
[本文引用: 2]
[54]
Weisskopf A Fuller DQ 2014 ) Buckwheat: Origins and development. In: Encyclopedia of Global Archaeology (ed. Smith C) , pp. 1025 -1028 . Springer-Verlag , New York .
[本文引用: 1]
[55]
Wu LY Chang FF Liu SJ SW Armbruster Huang SQ 2018 ) Heterostyly promotes compatible pollination in buckwheats: Comparisons of intraflower, intraplant, and interplant pollen flow in distylous and homostylous Fagopyrum
. American Journal of Botany , 105 , 108 -116 .
[本文引用: 1]
[56]
Wu LY Wang B Schoen DJ Huang SQ 2017 ) Transitions from distyly to homostyly are associated with floral evolution in the buckwheat genus (Fagopyrum )
. American Journal of Botany , 104 , 1232 -1240 .
[本文引用: 2]
[57]
Xia MZ Wang AH Cai GZ Yang P Liu JL 2007 ) A new variety of Fagopyrum Polygonaceae discovered in Sichuan, China—Fagopyrum polychromofolium A. H. Wang, J. L. Liu et P. Yang
. Journal of Xichang College , 21 , 11 -13 . (in Chinese with English abstract)
[夏明忠 , 王安虎 , 蔡光泽 , 杨坪 , 刘建林 (2007 ) 中国四川荞麦属(蓼科)一新种——花叶野荞麦
. 西昌学院学报 , 21 , 11 -13 .]
[58]
Yamane K Ohnishi O 2001 ) Phylogenetic relationships among natural populations of perennial buckwheat, Fagopyrum cymosum Meisn., revealed by allozyme variation
. Genetic Resources and Crop Evolution , 48 , 69 -77 .
[本文引用: 1]
[59]
Yang J Lu C Shen Q Yan Y Xu C Song C 2016 ) The complete chloroplast genome sequence of Fagopyrum cymosum
. Mitochondrial DNA Part A , 27 , 2410 -2411 .
[本文引用: 1]
[60]
Yang MJ Guo ZX Yang Y Chen YQ 2008 ) A brief history of buckwheat cultivation in China
. Inner Mongolia Agricultural Science and Technology , (5 ), 85 -86 . (in Chinese)
[本文引用: 2]
[杨明君 , 郭忠贤 , 杨媛 , 陈有清 (2008 ) 我国荞麦种植简史
. 内蒙古农业科技 , (5 ), 85 -86 .]
[本文引用: 2]
[61]
Yasui Y Hirakawa H Ueno M Matsui K Katsube-Tanaka T Yang SJ Aii J Sato S Mori M 2016 ) Assembly of the draft genome of buckwheat and its applications in identifying agronomically useful genes
. DNA Research , 23 , 215 -224 .
[本文引用: 1]
[62]
Yasui Y Mori M Aii J Abe T Matsumoto D Sato S Hayashi Y Ohnishi O Ota T 2012 ) S-LOCUS EARLY FLOWERING 3 is exclusively present in the genomes of short-styled buckwheat plants that exhibit heteromorphic self-incompatibility
. PLoS ONE , 7 , e31264 .
[本文引用: 1]
[63]
Yasui Y Ohnishi O 1998 a) Interspecific relationships in Fagopyrum (Polygonaceae) revealed by the nucleotide sequences of the rbcL and accD genes and their intergenic region
. American Journal of Botany , 85 , 1134 -1142 .
[本文引用: 1]
[64]
Yasui Y Ohnishi O 1998 b) Phylogenetic relationships among Fagopyrum species revealed by the nucleotide sequences of the ITS region of the nuclear rRNA gene
. Genes & Genetic Systems , 73 , 201 -210 .
[本文引用: 1]
[65]
Yi S Saiton T Zhao Q Wang P 2003 ) Vegetation and climate changes in the Changjiang (Yangtze River) Delta, China, during the past 13,000 years inferred from pollen records
. Quaternary Science Reviews , 22 , 1501 -1519 .
[本文引用: 1]
[66]
Yukio D 1960 ) Cytological studies in Polygonum and related genera
. Botanical Magazine (Tokyo) , 73 , 337 -340 .
[本文引用: 1]
[67]
Zhang LJ Li XX Ma B Gao Q Du HL Han YH Li Y Cao YH Qi M Zhu YX Lu HW Ma MC Liu LL Zhou JP Nan CH Qin YJ Wang J Cui L Liu HM Liang CZ Qiao ZJ 2017 ) The Tartary buckwheat genome provides insights into rutin biosynthesis and abiotic stress tolerance
. Molecular Plant , 10 , 1224 -1237 .
[本文引用: 1]
[68]
Zhao ZC Li BG Zhou MD 2007 ) Resources of Tartary Buckwheat and Related Wild Species in China . Sichuan Science and Technology Press , Chengdu . (in Chinese with English abstract)
[本文引用: 13]
[赵佐成 , 李伯刚 , 周明德 (2007 ) 中国苦荞及其近缘野生种资源 . 四川科技出版社 , 成都 .]
[本文引用: 13]
[69]
Zhou ML Wang CL Wang DZ Zheng YD Li FL Zhu XM Tang YX Shao JR Tang Y Wu YM 2014 ) Phylogenetic relationship of four new species related to southwestern Sichuan Fagopyrum based on morphological and molecular characterization
. Biochemical Systematics and Ecology , 57 , 403 -409 .
[本文引用: 1]
[70]
Zhou ML Zhang Q Zheng YD Tang Y Li FL Zhu XM Shao JR 2015 ) Fagopyrum hailuogouense (Polygonaceae), one new species from Sichuan, China
. Novon , 24 , 222 -224 .
[本文引用: 4]
[71]
Zhou X Wen L Li Z Zhou Y Chen Y Lu Y 2015 ) Advance on the benefits of bioactive peptides from buckwheat
. Phytochemistry Reviews , 14 , 381 -388 .
The effectiveness of heterostyly in preventing illegitimate pollination in dish-shaped flowers
1
1995
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
Old World globalization and the Columbian exchange: Comparison and contrast
1
2012
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
1
1976
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Floral biology of common buckwheat (Fagopyrum esculentum Moench)
2
2009
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
A study of resources of Fagopyrum (Polygonaceae) native to China
2
1999
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
1
2001
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
8
2012
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... 荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
8
2012
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... 荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
甘肃武威磨咀子汉墓发掘
1
1960
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
甘肃武威磨咀子汉墓发掘
1
1960
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
Complete chloroplast genome sequence of tartary buckwheat (Fagopyrum tataricum ) and comparative analysis with common buckwheat (F. esculentum )
1
2015
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Food and Agriculture Organization of the United Nations (FAO)
3
2014
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
... 世界范围内荞麦种植现状, 显示欧亚大陆普遍栽培, 数据来源于FAO (2014) . ...
... Buckwheat planting status in the world, showing the prevalence of cultivation in Eurasia, data from FAO (2014) ...
Origin, geographical distribution and phylogenic relationships of common buckwheat (Fagopyrum esculentum Moench)
1
2010
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
Remarques sur les Polygonées de l’Asie Orientale
2
1913
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Pollen morphology in the genus Polygonum L. (s. lat.) and its taxonomical significance
2
1946
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Fagopyrum luojishanense , a new species of Polygonaceae from Sichuan, China
1
2015
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
基于ITS和ndh F-rpl 32序列的荞麦种间亲缘关系分析
1
2016
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
基于ITS和ndh F-rpl 32序列的荞麦种间亲缘关系分析
1
2016
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Is buckwheat (Fagopyrum esculentum Moench) still a valuable crop today?
1
2012
... 荞麦是一种很好的药食同源植物.由于荞麦果实成熟后容易脱落的落粒特性, 结实率及产量低下, 对其开发和利用的空间还很大.甜荞两型花之间授粉才能结实, 而苦荞可以自交, 但即使是人工补充授粉, 甜荞的结实率都不到50%, 而苦荞结实率可达80% (作者未发表数据).由于荞麦自身的生物学特性以及其他大宗作物的发展导致荞麦播种面积呈减少趋势, Jacquemart等(2012) 质疑了荞麦是否仍然是有前途的作物.为保障国家粮食安全, 开发新的主粮作物被我国政府提上日程.荞麦作为一种起源于中国的粮食作物, 与长江流域文明的发展密不可分.但是我们目前对其生物学特性、起源地, 如何提高荞麦产量和种植技术方面还有待深入研究, 培育出自交亲和的甜荞品种有利于提高甜荞的产量, 从而扩大其种植范围.该属具有二型和同型花柱物种, 为性系统演化的研究提供了好的模式系统.我们正在同国内基因组学的同行合作, 将在近期完成甜荞的全基因组测序, 为回答这一演化问题和挖掘优良的农艺性状奠定基础. ...
Floral visitors and the importance of honey bee on buckwheat (Fagopyrum esculentum Moench) in central Belgium
2
2007
... 自交不亲和的甜荞依赖传粉者传递花粉才结实, 调查表明访花传粉昆虫是泛化的, 主要包括膜翅目(蜜蜂、熊蜂、独居蜂)、双翅目(食蚜蝇科、丽蝇科等), 另外还有鳞翅目、半翅目、脉翅目等其他昆虫(Jacquemart et al, 2007 ; Wu et al, 2017 ).在许多国家, 蜜蜂被认为是栽培荞麦最普遍且最主要的访花者(Jacquemart et al, 2007 ).但不同访花昆虫的传粉效率仍需进一步研究. ...
... ).在许多国家, 蜜蜂被认为是栽培荞麦最普遍且最主要的访花者(Jacquemart et al, 2007 ).但不同访花昆虫的传粉效率仍需进一步研究. ...
Chloroplast DNA analysis in buckwheat species: Phylogenetic relationships, origin of the reproductive systems and extended inverted repeats
1
1995
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
1
2001
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
Enrichment of gluten-free cakes with lupin (Lupinus albus L.) or buckwheat (Fagopyrum esculentum M.) flours
1
2011
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
Fagopyrum
4
2003
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
... 荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
Early cultivated wheat and broadening of agriculture in Neolithic China
1
2007
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
陕西咸阳马泉西汉墓
1
1979
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
陕西咸阳马泉西汉墓
1
1979
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
The impact of ancient civilization on the northeastern Chinese landscape: Palaeoecological evidence from the Western Liaohe River Basin, Inner Mongolia
1
2006
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
a) 中国四川蓼科荞麦属一新种——皱叶野荞麦
1
2008
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
a) 中国四川蓼科荞麦属一新种——皱叶野荞麦
1
2008
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
b). 中国荞麦属(蓼科)一新种——密毛野荞麦
1
2008
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
b). 中国荞麦属(蓼科)一新种——密毛野荞麦
1
2008
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
De novo sequencing and characterization of floral transcriptome in two species of buckwheat (Fagopyrum )
1
2011
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
Comparative chloroplast genomics and phylogenetics of Fagopyrum esculentum ssp. ancestrale —a wild ancestor of cultivated buckwheat
1
2008
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Development of SSR markers for studies of diversity in the genus Fagopyrum
1
2009
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Agroecological classification and geographical distribution of the common buckwheat, Fagopyrum esculentum M. in the East Asia
1
1979
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench X. Diffusion routes revealed by RAPD markers
1
1996
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
1
1957
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Topological incongruence between nuclear and chloroplast DNA trees suggesting hybridization in the urophyllum group of the genus Fagopyrum (Polygonaceae)
1
2003
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. VII. Allozyme variability in Japan, Korea, and China
2
1988
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. VIII. Local differentiation of land races in Europe and the silk road
1
1993
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
6
1995
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Search for the wild ancestor of buckwheat. I. Description of new Fagopyrum (Polygonaceae) species and their distribution in China and the Himalayan hills
1
1998
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
3
2004
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Search for the wild progenitor of buckwheat. II. Taxonomy of Fagopyrum (Polygonaceae) species based on morphology, isozymes and cpDNA variability
1
1996
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
New Fagopyrum species revealed by morphological and molecular analyses
1
1998
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
Intra- and interspecific phylogeny of wild Fagopyrum (Polygonaceae) species based on nucleotide sequences of noncoding regions in chloroplast DNA
1
2000
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Two new Fagopyrum (Polygonaceae) species, F. gracilipedoides and F. jinshaense from Yunnan, China
1
2002
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
5
2015
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
5
2015
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Fagopyrum wenchuanense and Fagopyrum qiangcai , two new species of Polygonaceae from Sichuan, China
1
2011
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
Species relationships in Fagopyrum revealed by PCR-based DNA fingerprinting
1
2002
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Buckwheat seed set in planta and during in vitro inflorescence culture: Evaluation of temperature and water deficit stress
1
2001
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
The Polygoneae of Eastern Asia
4
1930
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
2
2016
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... ; Tang et al, 2016 ). ...
Fagopyrum pugense (Polygonaceae), a new species from Sichuan, China
1
2010
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
Origin of cultivated tartary buckwheat (Fagopyrum tataricum Gaertn.) revealed by RAPD analyses
1
2000
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Phylogenetic relationships among wild and cultivated tartary buckwheat (Fagopyrum tataricum Gaert.) populations revealed by AFLP analyses
1
2001
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
Fagopyrum longzhoushanense , a new species of Polygonaceae from Sichuan, China
1
2017
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
2
1995
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
... ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
1
2014
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Heterostyly promotes compatible pollination in buckwheats: Comparisons of intraflower, intraplant, and interplant pollen flow in distylous and homostylous Fagopyrum
1
2018
... 荞麦是世界上广为种植的谷物类粮食作物, 因其具有药用价值, 现在人们开发了荞麦茶、荞麦酒等农副产品.人们俗称的荞麦是指栽培的两个种, 即甜荞和苦荞(图1 ).已有的证据表明甜荞(Fagopyrum esculentum )和苦荞(F. tataricum )是独立的两个种, 均起源于我国西南地区长江流域的上游.荞麦是唯一不属于禾本科的、世界主要的谷物类粮食作物; 其他谷物类是通过风媒或自花传粉, 而荞麦则是虫媒或自花传粉.另外, 有别于其他作物, 甜荞是二型花柱植物, 居群中有两种基因型不同的个体(Wu et al, 2018 ).中国被认为是最早栽培荞麦的国家, 而且荞麦属的大多数野生近缘物种分布在中国.因此荞麦为我国研究栽培植物的起源、散布和野生资源的利用、植物性系统的演化以及长江流域文明的起源提供了一个好的研究系统.本文试图对前人的研究工作进行小结, 提出值得深入研究的问题, 以期为我国的粮食安全、资源保护以及生态文明建设提供参考依据. ...
Transitions from distyly to homostyly are associated with floral evolution in the buckwheat genus (Fagopyrum )
2
2017
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
... 自交不亲和的甜荞依赖传粉者传递花粉才结实, 调查表明访花传粉昆虫是泛化的, 主要包括膜翅目(蜜蜂、熊蜂、独居蜂)、双翅目(食蚜蝇科、丽蝇科等), 另外还有鳞翅目、半翅目、脉翅目等其他昆虫(Jacquemart et al, 2007 ; Wu et al, 2017 ).在许多国家, 蜜蜂被认为是栽培荞麦最普遍且最主要的访花者(Jacquemart et al, 2007 ).但不同访花昆虫的传粉效率仍需进一步研究. ...
中国四川荞麦属(蓼科)一新种——花叶野荞麦
2007
中国四川荞麦属(蓼科)一新种——花叶野荞麦
2007
Phylogenetic relationships among natural populations of perennial buckwheat, Fagopyrum cymosum Meisn., revealed by allozyme variation
1
2001
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
The complete chloroplast genome sequence of Fagopyrum cymosum
1
2016
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
我国荞麦种植简史
2
2008
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
我国荞麦种植简史
2
2008
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
Assembly of the draft genome of buckwheat and its applications in identifying agronomically useful genes
1
2016
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
S-LOCUS EARLY FLOWERING 3 is exclusively present in the genomes of short-styled buckwheat plants that exhibit heteromorphic self-incompatibility
1
2012
... 荞麦属植物的性系统的调查显示该属有同型花柱(homostyly)和二型花柱(distyly)物种(表1 , 图1 ), 我们最近的分析表明该属至少发生过3次从二型花柱向同型花柱的演化(Wu et al, 2017 ).二型花柱是指同一物种的不同个体上花药高度与柱头位置交互对应, 表现出交互的雌雄异位现象; 同型花柱是指花药柱头高度没有空间上的分离, 处在同一位置.甜荞为二型花柱的物种, 同型花授粉和自交均不亲和、长花柱花与短花柱花之间的异交授粉才能结实.与典型的异型花柱植物类似, 甜荞长柱花的花粉产量比短柱型高, 而花粉大小比短柱型小(Björkman, 1995 ).苦荞为花柱同型、自交亲和(Nishimoto et al, 2003 ).另外, 甜荞花相对较大、粉色、花蜜量较大; 而苦荞花小、偏绿色、花蜜量小.据甜荞和苦荞的转录组测序的结果表明, 这些差别可能与基因的差异性表达有关, 甜荞中与二糖代谢相关的潜在差异表达基因相对丰富, 而葡萄糖、果糖和蔗糖是甜荞花蜜的主要成分(Logacheva et al, 2011 ).研究发现S-ELF3 基因只在甜荞短柱型植株表达, 可能是控制甜荞短柱表型的候选基因, 此基因也可能与自交不亲和性有关(Yasui et al, 2012 ).利用构建的甜荞基因组草图作为参考序列, 通过高通量测序标记技术, Yasui等(2016) 成功确定了控制甜荞自交不亲和的新的候选基因, 基因组数据库和基因组草图序列可为开发具有优良农艺性状的荞麦品种提供基础. ...
Interspecific relationships in Fagopyrum (Polygonaceae) revealed by the nucleotide sequences of the rbcL and accD genes and their intergenic region
1
1998
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Phylogenetic relationships among Fagopyrum species revealed by the nucleotide sequences of the ITS region of the nuclear rRNA gene
1
1998
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Vegetation and climate changes in the Changjiang (Yangtze River) Delta, China, during the past 13,000 years inferred from pollen records
1
2003
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
Cytological studies in Polygonum and related genera
1
1960
... 荞麦是蓼科荞麦属(Fagopyrum )植物, 曾隶属于蓼属(Polygonum ).林奈在《植物种志》中把荞麦放于广义的蓼属(Polygonum sensu lato.)中; Miller于1754年建立荞麦属(Fagopyrum Miller), 随后Moench也独立建立了荞麦属(Fagopyrum Moench).Meissner将荞麦各种归为蓼属中的一个组(Fagopyrum sect. Meissn.), 随后他又认为荞麦应该作为一个独立的属(Fagopyrum Meissn.) (陈庆富, 2012 ).Gross (1913) 曾试图对荞麦属进行系统的修订, 但对于荞麦属的分类地位仍有争议.Steward (1930) 根据植物的形态特征, 如花序类型、托叶鞘形状、瘦果形状等, 认为荞麦应该作为蓼属中的一个组.荞麦属具有不同的花粉外壁纹饰(Hedberg, 1946 ), 其染色体基数为8, 而蓼属的基数为10、11、12 (Yukio, 1960 ), 这些差别支持荞麦独立成属.自国际命名法出台后, 根据优先的原则荞麦属名统一为Fagopyrum Miller. ...
The Tartary buckwheat genome provides insights into rutin biosynthesis and abiotic stress tolerance
1
2017
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
13
2007
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... 荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
... ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
... 目前的研究主要集中在栽培的甜荞和苦荞, 对于我国西南丰富的野生荞麦资源研究甚少, 对于这些野生种质资源应该加以保护和利用.据研究野生金荞麦的根茎具有抑菌、抗炎和抗肿瘤等作用(赵佐成等, 2007 ), 我们2012-2015年在四川、云南进行野外调查时, 注意到大量野生资源遭到破坏, 有些种群已经消失.对二型花柱植物, 种群中两种表型的比例偏离1:1将影响有性繁殖, 加剧了地方种群的灭绝.对于这些珍贵的资源植物的家底需要调查并实行有效的保护措施, 避免陷入濒临灭绝的境地.对于荞麦属的分类地位及栽培荞麦的祖先种仍存在诸多争议, 在开展野外种质资源收集的同时, 需要厘清该属的系统发生与演化关系. ...
13
2007
... 对于荞麦的认识及利用, 我国自古就有诸多记载, 如北魏贾思勰所著的《齐民要术》和明代李时珍撰写的《本草纲目》.前者对荞麦的栽种技术作了专门介绍, 后者对荞麦的药用价值进行了描述(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).长江中下游的孢粉证据表明早在4,500年前长江流域的南部丘陵地区就有荞麦的种植, 这与长江地区的农耕文化时间上相吻合(良渚文化, 距今5,900-4,500年) (Yi et al, 2003 ).另外, 在中国的辽河流域、甘肃西山坪发现的孢粉学证据也表明我国的荞麦文明可能在4,500年前就开始了(Li et al, 2006 , 2007 ; Boivin et al, 2012 ).在陕西省和甘肃省汉代墓葬中出土了距今已有约2,000年的荞麦种子实物(陈贤儒, 1960 ; 李毓芳, 1979 ).自西周至春秋时期、南北朝、唐、宋、元、明等各朝不仅记载了荞麦的种植, 还记载了其种植技术, 而真正开始大范围普及种植的时间是在唐朝, 其后对荞麦的药用价值也有了记载, 在明代荞麦种植技术更趋于完善(杨明君等, 2008 ).这些出土的实物、文字记载表明我国种植荞麦的历史悠久. ...
... 荞麦在东亚及其邻近区域广为种植, 经朝鲜半岛传入日本.公元800年, 荞麦在日本是一种很重要的粮食作物(Wei, 1995 ).在13-14世纪, 经西伯利亚和俄罗斯南部传入欧洲(Gondola & Papp, 2010 ).也有基于等位酶分析的研究认为, 欧洲荞麦是从中国北方沿着“丝绸之路”进行扩散的(Ohnishi, 1993 ).据报道, 德国是欧洲最早种植荞麦的国家, 随后在17世纪, 荞麦被引入比利时、法国、意大利和英国 (Wei, 1995 ), 并由移民带入北美、阿根廷、巴西和南非(Kreft, 2001 ).在17-19世纪, 荞麦在西方国家是非常流行的食物(Cawoy et al, 2009 ).据联合国粮农组织(FAO, 2014 )的统计, 全球荞麦的收获面积有2,011,289 ha, 总产量1,924,082 t, 而且主要分布于北温带(图2 ).生产国主要有俄罗斯、中国、乌克兰、法国、波兰、美国、巴西、哈萨克斯坦、立陶宛、日本等(图2 ); 其中在中国收获面积约708,000 ha, 年产量约564,900 t, 均居世界第二.我国四川省凉山彝族自治州具有种植苦荞的传统, 常年种植面积约有46,666 ha, 年产量约有10,000 t, 苦荞是该自治州的主要粮食作物之一, 播种面积约占所有粮食作物种植面积的40-50% (赵佐成等, 2007 ).由于水稻、小麦等农作物在云贵川等高海拔地区种植困难, 荞麦可能在孕育当地文明中起着极其重要的作用.荞麦在我国的栽培时间可能比藏族种植青稞(Hordeum vulgare Linn. var. nudum Hook. f.)更早, 可能是早期山区人们的重要粮食作物. ...
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
... 关于栽培甜荞与苦荞的祖先问题, 目前主要有两种假说.早期研究者认为金荞麦是甜荞和苦荞的祖先(Gross, 1913 ; Steward, 1930 ; Hedberg, 1946 ; Campbell, 1976 ), 在形态学上金荞麦与甜荞更为接近; 但后来分子水平(cpDNA和同工酶)的分析表明金荞麦与苦荞的亲缘关系更近(Kishima et al, 1995 ; Yamane & Ohnishi, 2001 ), 不支持该观点.另一观点认为甜荞和苦荞是独立起源的, 但对于二者的祖先种存在两种不同的观点.一是认为甜荞和苦荞的祖先分别为野生甜荞和野生苦荞, 主要基于甜荞和苦荞野生种的发现及其地理分布特性, 以及亲缘关系分析.Ohnishi (1995) 在中国云南发现野生甜荞(F. esculentum ssp. ancestralis )和野生苦荞(F. tataricum ssp. potanini ), 且与栽培荞麦可以杂交.野生甜荞的分布局限于云南西部和四川, 海拔1,000-1,500 m, 云南金沙江流域和四川雅砻江流域的贫瘠土壤及岩石生境(Ohnishi, 1995 , 2004 ), 其中长江流域上游的三江流域(金沙江、澜沧江和怒江)是野生甜荞的分布中心(赵佐成等, 2007 ; 陈庆富, 2012 ).野生苦荞分布则较为广泛, 青藏高原较寒冷地区都有分布(Ohnishi, 1995 , 2004 ).甜荞与苦荞的野生祖先的分布区域的不同, 也表明两种栽培荞麦是独立起源的(Ohnishi, 1995 ; 陈庆富, 2012 ).形态特征、同工酶变异和cpDNA的RFLP (restriction fragment length polymorphism)分析表明苦荞与野生苦荞亲缘关系很近、甜荞与野生甜荞亲缘关系很近(Ohnishi, 1995 , 1998; Ohnishi & Matsuoka, 1996 ).同工酶和AFLP (amplified fragment length polymorphism)分析表明, 我国西藏东部可能是栽培甜荞起源地之一(Ohnishi, 2004 ).野生苦荞与栽培苦荞的RAPD和AFLP分析表明, 栽培苦荞可能起源于中国西藏东部或云南西北部(Tsuji & Ohnishi, 2000 , 2001 ).另一观点认为甜荞的祖先种是大野荞(F. megaspartanium ), 而苦荞的祖先种是毛野荞(F. pilus ).Chen (1999)认为甜荞和苦荞分别起源于中国西南部较温暖地区和青藏高原东部海拔较高的冷凉地区.甜荞和苦荞的形态学、细胞学、生态适应性、繁殖特点等基本特征分别类似于大野荞和毛野荞(Chen, 1999 , 2001 ; 陈庆富, 2012 ).推测毛野荞生长于不利于虫媒传粉的冷凉气候中, 诱发了基因重组和突变, 最后进化成苦荞.虽然目前支持甜荞和苦荞独立起源的证据较多, 但其祖先种的确定主要是日本学者在20世纪90年代开展的; 利用新一代基因组学的方法探寻其可能祖先类群, 成为澄清这一问题的有效途径. ...
... 荞麦属植物是一年生或多年生草本, 少为半灌木(硬枝野荞麦、大野荞和毛野荞) (Li & Hong, 2003 ; 陈庆富, 2012 ).生境类型广泛, 包括山坡草地、山谷湿地、路边、农田和荒地等(Li & Hong, 2003 ; 赵佐成等, 2007 ).荞麦属的花序总状或伞房状, 辐射对称花两性, 白色、粉色或浅绿色; 花被片5, 雄蕊有8枚, 排成2轮, 外轮5, 内轮3; 花柱3, 胚珠1, 花基部有蜜腺, 单花寿命为1天.花期一般为6-9月, 果期8-10月(Li & Hong, 2003 ).栽培荞麦在我国南方可以春秋两季播种.瘦果具3棱, 成熟后由绿色变成灰褐色且易掉落.属的模式种为甜荞(Fagopyrum esculentum Moench). ...
... 甜荞和苦荞在生态适应性上也有差异, 甜荞适应比较温暖的气候(最适生长温度: 18-23℃), 若温度低于15℃, 开花就受到抑制, 温暖气候有利于虫媒传粉; 而苦荞能适应比较寒冷的气候(最适生长温度: 12-23℃), 对高温比较敏感, 高温对植株生长发育和产量不利(赵佐成等, 2007 ; Cawoy et al, 2009 ; 陈庆富, 2012 ).较高温度(25℃以上)会导致荞麦的花萎蔫、种子败育、胚囊畸形以及果实干瘪等(Slawinska & Obendorf, 2001 ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
... ).荞麦自古以来就有种植, 主要是由于它生长周期短(60-80天)、适应性强, 能有效地利用各种资源(赵佐成等, 2007 ; 杨明君等, 2008 ). ...
... 目前的研究主要集中在栽培的甜荞和苦荞, 对于我国西南丰富的野生荞麦资源研究甚少, 对于这些野生种质资源应该加以保护和利用.据研究野生金荞麦的根茎具有抑菌、抗炎和抗肿瘤等作用(赵佐成等, 2007 ), 我们2012-2015年在四川、云南进行野外调查时, 注意到大量野生资源遭到破坏, 有些种群已经消失.对二型花柱植物, 种群中两种表型的比例偏离1:1将影响有性繁殖, 加剧了地方种群的灭绝.对于这些珍贵的资源植物的家底需要调查并实行有效的保护措施, 避免陷入濒临灭绝的境地.对于荞麦属的分类地位及栽培荞麦的祖先种仍存在诸多争议, 在开展野外种质资源收集的同时, 需要厘清该属的系统发生与演化关系. ...
Phylogenetic relationship of four new species related to southwestern Sichuan Fagopyrum based on morphological and molecular characterization
1
2014
... 国内外对荞麦属内系统发生关系进行了一系列研究, Yasui和Ohnishi (1998a , b )分别利用rbc L, acc D及其之间的基因间隔区, 以及ITS和rRNA揭示了12个种的系统关系.Ohsako和Ohnishi (2000) 通过测叶绿体基因非编码区的核苷酸序列(trn K和trn C (GCA)-rpo B), 研究了荞麦小粒组10个种的种内和种间的系统演化关系.Sharma和Jana (2002) 利用随机扩增多态性DNA (random amplified polymorphic DNA, RAPD)分析了荞麦属14个种和2个亚种的系统关系.Nishimoto等(2003)利用2个核基因的核苷酸序列(FLO /LFY 和AG )和3个cpDNA (rbc L-acc D, trn K和trn C-rpo B)的片段对荞麦属小粒组系统进化关系进行了分析.Zhou等(2014) 分别利用ITS和mat K构建了10种荞麦的系统树, 认为汶川野荞麦与金荞麦(F. dibotrys )亲缘关系很近, 应属于大粒组.胡亚妮等(2016) 利用ITS和ndh F-rpl 32序列构建了10种荞麦的系统树, 并分析了种间关系.Ma等(2009) 概述了研究荞麦属遗传多样性的SSR分子标记的开发, 通过SSR标记可获得序列和多样性信息, 有利于了解遗传结构和种间关系.以上系统发生学的结果均表明, 荞麦可以分为两组: 大粒组(cymosum)和小粒组(urophyllum), 但是对各种间关系目前争论较大, 不同研究方法与分子标记得出的结果不全一致.目前, 甜荞、苦荞和金荞麦的叶绿体全基因组序列都已测出(Logacheva et al, 2008 ; Cho et al, 2015 ; Yang et al, 2016 ), 这将为确定荞麦的种间关系奠定基础. ...
Fagopyrum hailuogouense (Polygonaceae), one new species from Sichuan, China
4
2015
... 近年来, 对荞麦理化性质、药用功效及营养价值的研究较多.苦荞产量高于甜荞, 种植面积大, 是目前研究较多的物种(赵佐成等, 2007 ).制成的苦荞茶、苦荞酒、苦荞醋含有抗氧化、调节血糖、防治心血管疾病等多种生物活性物质, 是现代一类新兴保健产品.荞麦制成的食品(荞麦馒头、荞麦面条等)还具有促进消化、增强免疫力、预防癌症等作用.临床医学观察表明, 苦荞含丰富的黄酮类物质, 对糖尿病、高血压、高血脂、冠心病等疾病的治疗都有一定的辅助作用.尤其其中的芦丁(含量0.8-1.5%, 其他谷物中几乎没有)可以防治因毛细血管脆弱引起的各种出血病, 并用作高血压的辅助治疗剂(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).Zhang等(2017) 的研究获得了苦荞高质量(489.3 Mb)的基因组序列, 并解析了芦丁的生物合成, 鉴定了芦丁生物合成途径中编码参与代谢的酶的基因及调控这些基因表达的转录因子, 同时注释了苦荞中存在大量可能与植物耐铝、抗旱和耐寒相关的新基因.荞麦中含有丰富的蛋白质, 含量高达10-14.5%, 其中含有赖氨酸、精氨酸等氨基酸, 比例适当, 很容易被人体吸收利用(赵佐成等, 2007 ; 任长忠和赵钢, 2015 ).研究表明, 从荞麦种子中可以分离出多种自然产生的肽, 如荞麦抗菌肽、胰蛋白酶抑制剂、抗肿瘤蛋白、降压肽和抗氧化肽(Zhou et al, 2015 ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... ).荞麦胰蛋白酶抑制剂除了对胰蛋白酶具有抑制作用外, 还对真菌、革兰氏阳性和革兰氏阴性菌以及多种肿瘤细胞具有抗性(Zhou et al, 2015 ).另外荞麦含有丰富的B族维生素及维生素C、E等参与生物体内糖、蛋白质和脂肪代谢的重要微量有机物质, 还含有铁、锌、铜、锰、铬、硒等人体必需的微量元素, 是一种营养丰富的粮食作物.荞麦中脂肪的含量2.1-2.5%, 含有9种脂肪酸, 大部分为不饱和油酸和亚油酸(赵佐成等, 2007 ; 陈庆富, 2012 ).此外, 荞麦不同于其他禾本科的谷物, 无麸质(gluten-free), 适合对麸质过敏的人食用(Levent & Bilgiçli, 2011 ). ...
... 《中国植物志》记载荞麦属有15种, 广布于亚洲和欧洲; 其中中国有10种1变种, 有2种为栽培种(甜荞和苦荞, 图1 ) (Li & Hong, 2003 ); 至今仍不断有新种被报道.Ohnishi (1998) 报道了4个新种, F. pleioramosum , F. callianthum , F. capillatum 和F. homotropicum .Ohsako和Ohnishi (1998) 报道了分布于中国云南和四川的2个新种F. macrocarpum 和F. rubrifolium .Ohsako等(2002)又报道了2个新种F. gracilipedoides 和F. jinshaense .夏明忠等(2007)在中国四川阿坝州发现了花叶野荞麦(F. polychromofolium ).刘建林等(2008a , b )报道了中国四川凉山州2种荞麦新种皱叶野荞麦(F. crispatifolium )和密毛野荞麦(F. densivillosum ).Tang等(2010) 在四川普格县螺髻山发现1种新的荞麦普格野荞麦(F. pugense ).Shao等(2011) 在中国四川发现了汶川野荞麦(F. wenchuanense )和羌彩野荞麦(F. qiangcai ).Hou等(2015) 在中国四川凉山州发现了新种螺髻山野荞麦(F. luojishanense ).Zhou等(2015) 在中国四川甘孜藏族自治州发现1种荞麦新种海螺沟野荞麦(F. hailuogouense ).Wang等(2017) 在中国四川普格县发现新种龙肘山野荞麦(F. longzhoushanense ).目前在中国报道的荞麦有30个种(表1 ), 其中部分种的性系统、生活史和分布也已记录, 但该属的分类修订亟待开展, 特别是结合DNA序列、基因组学的性状以确认这些新命名的合法地位. ...
... 关于栽培荞麦的起源地, 早期1883年, De Candolle认为荞麦起源于西伯利亚或中国北方(黑龙江流域) (Matano & Ujihara, 1979 ).Steward (1930) 总结了蓼族(Trib. Polygoneae)植物的分类和分布, 通过调查标本, 认为荞麦原产于中国南方.根据Steward (1930) 对中国西南野生荞麦分类与分布的总结, Nakao (1957) 也认为栽培荞麦可能起源于中国南部.后来一系列地理分布的研究(Chen, 1999 ; 赵佐成等, 2007 ; 陈庆富, 2012 ; 任长忠和赵钢, 2015 )表明, 栽培种(甜荞和苦荞)以及2个野生种金荞麦和细柄野荞麦(F. gracilipes )分布范围较广, 而其他大多数野生荞麦主要分布在中国云南、四川、西藏、贵州等西南地区(表1 ), 特别是金沙江流域野生荞麦资源尤为丰富.近年来, 多个野生新种的发现地均在云南、四川地区(Ohnishi, 1998 ; Ohsako et al, 2002 ; Zhou et al, 2015 ; Tang et al, 2016 ).亚洲甜荞的等位酶分析结果表明, 中国南部居群内的变异最大(Ohnishi, 1988 ).甜荞的RAPD分析也表明中国南部的多态性比例最高, 因此研究者提出甜荞是从中国南部扩散至亚洲其他国家(Murai & Ohnishi, 1996 ).任长忠和赵钢(2015) 根据瓦维洛夫的作物起源中心学说及前人的研究, 认为云南西北部可能是栽培苦荞的初生起源中心, 西藏东部和四川中部与南部可能是其次生起源中心; 而野生荞麦主要分布于云南西部和中部, 且云南西部也是苦荞的主产区, 这两个野生荞麦主要分布区可能是中国荞麦的另一个次生起源中心.目前证据支持中国西南部是荞麦属的分布及多样性中心, 也是其起源中心(Ohnishi, 1995 ; 赵佐成等, 2007 ; Weisskopf & Fuller, 2014 ; Tang et al, 2016 ). ...
Advance on the benefits of bioactive peptides from buckwheat
2015




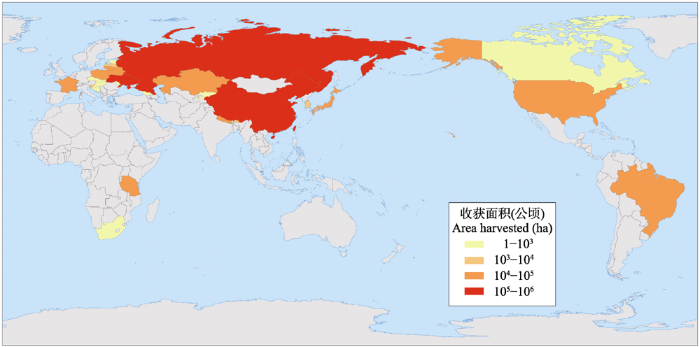
{kind=link}
{kind=link}
{kind=link}
{kind=link}