1 Institute of Zoology, Chinese Academy of Sciences, Beijing 1001012 University of Chinese Academy of Sciences, Beijing 1000493 Guangdong Key Laboratory of Animal Conservation and Resource Utilization, Guangdong Public Laboratory of Wild Animal Conservation and Utilization, Guangdong Institute of Applied Biological Resources, Guangzhou 5102604 School of Nature Conservation, Beijing Forestry University, Beijing 1000835 Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091
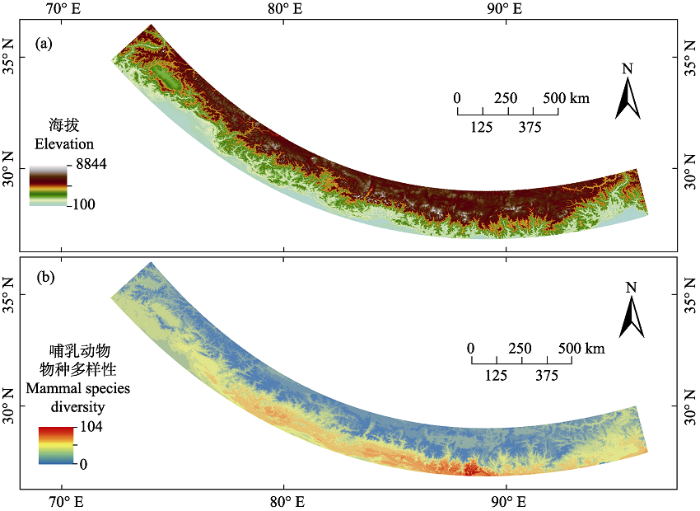
Studies of the distribution of biodiversity are hotspots in ecology and biogeography. Mountain ecosystems, which contain high habitat heterogeneity and biodiversity, play an important role in biodiversity conservation. The Himalayas are located in the southern rim of the Qinghai-Tibetan Plateau. The complicated geological topography, vast elevational gradients (100-8,844 m), significant vertical climatic zonation, and diverse biodiversity make it an ideal study area for biodiversity distribution research. By combining field data and data from museum records and literature, 313 mammal species were found for each 100 m elevational band along the gradient from 100 to 6,000 m above sea level (a.s.l.) in the Himalayas. Most of the elevational species richness patterns were hump-shaped. Species richness of the overall mammals peaked at an elevation of 900-1,400 m. Species composition along the elevational gradient in the Himalayas can be classified into five groups (100-1,500 m, 1,500-2,000 m, 2,000-3,000 m, 3,000-4,200 m and 4,200-6,000 m a.s.l.) using UPGMA clustering of elevation-based species assemblages, which was similar to the divisions of the vertical vegetation zone. The distribution of mammals is uneven, with low diversity found in the northern slope of the Himalayas and high diversity in the southern slope; the valleys on south slope of the Himalayas that contain higher biodiversity are biological corridors connecting the northern and southern fauna. To maintain the exchange of biodiversity, we should sustainably strengthen protection to the valley ecosystems.
Keywords:Himalayas
;
mammal species diversity
;
elevational patterns
;
distribution
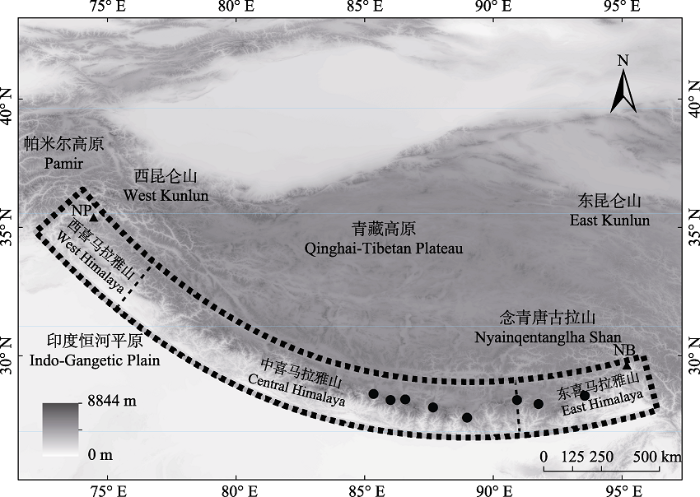
Fig. 1 Generalized topographic map of the Himalaya Mountains and Tibetan Plateau. The black dashed lines with shadow represent boundaries of the study area. Black dashed lines represent boundaries between the western, central, and eastern Himalaya Mountains. Black dots represent field survey areas, from left to right are: Gyirong County, Nyalam County, Tingri County, Dinggyê County, Yadong County, Lhozhag County, Cona County and Lhünzê County, respectively. NP, Nanga Parbat; NB, Namche Barwa.
整合野外调查数据和文献数据, 得出喜马拉雅山地区哺乳动物名录, 根据各物种的分布记录和海拔信息, 获取各物种分布区的海拔上、下限。另外, 基于野外考察观测值, 剔除和校正一些异常值。野外调查的哺乳动物物种编目参考《中国哺乳动物种和亚种分类名录与分布大全》(王应祥, 2003)和《中国哺乳动物多样性及地理分布》(蒋志刚等, 2015)。最后, 整体的喜马拉雅山地区哺乳动物物种名录根据The IUCN Red List of Threatened Species (IUCN, 2017)的分类体系重新合并整理, 确保各物种的准确(附录2)。
采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布。喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究。按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值。为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m)。
根据哺乳动物的运动方式与体型将其分为: 飞行哺乳动物(即蝙蝠类, Chiroptera)与非飞行哺乳动物(non-volant mammals), 非飞行小型哺乳动物(包括兔形目、食虫目、啮齿目和攀鼩目)和非飞行大中型哺乳动物(non-volant large and medium mammals)。
1.4 统计与分析
1.4.1 物种多样性垂直分布模式的检验
将海拔作为自变量, 各个海拔区间的物种多样性作为因变量, 构建多项式回归模型, 并根据校正赤池信息准则AICc (the corrected Akaike information criterion)的最小值, 从一阶和二阶的多项式(分别代表线性和单峰曲线)中选出最优模型来确定垂直分布格局的模式。
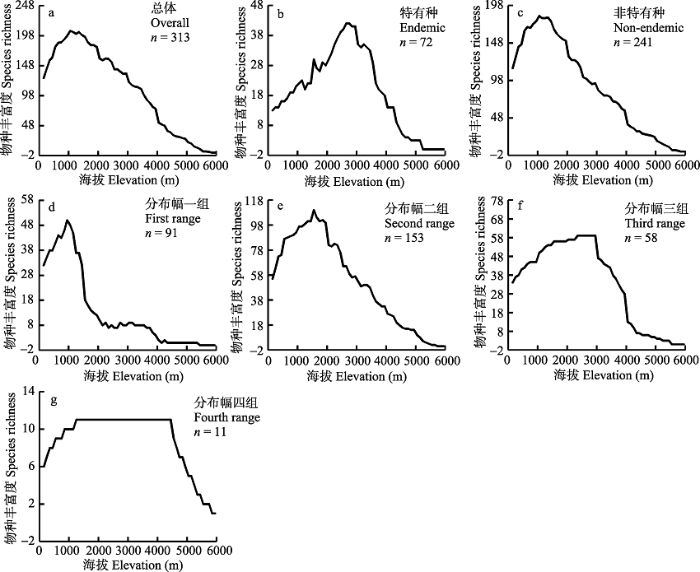
Fig. 2 Elevational patterns of mammal species richness for different distribution range subsets in the Himalaya Mountains. a, All mammals; b, Endemic mammals; c, Non-endemic mammals; d-g, Mammals in the first, second, third and fourth range size classes, respectively.
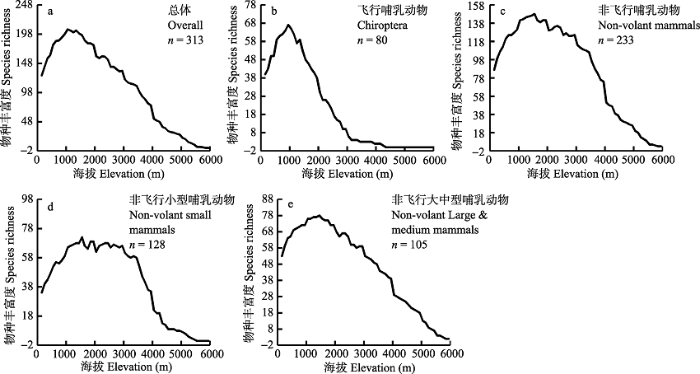
Fig. 3 Elevational patterns of mammal species richness for different biological habits and body size subsets in the Himalaya Mountains. a, All mammals; b, Chiroptera; c, Non-volant mammals; d, Non-volant small mammals; e, Non-volant large and medium mammals.
Table 1
表1
表1 喜马拉雅山地区哺乳动物不同子集的物种多样性沿海拔梯度的多项式回归
Table 1 Polynomial regressions for the mammal species richness along the elevational gradients in the Himalaya Mountains for each species subsets
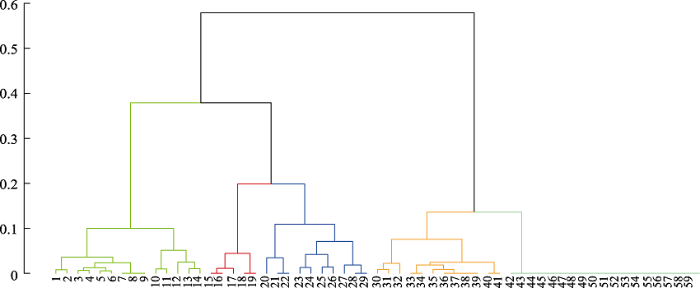
Fig. 4 Dendrograms resulting from UPGMA hierarchical clustering of 100 m elevational band assemblages based on βsim dissimilarity matrices for mammal species richness along the elevational gradient in the Himalayas. The agglomerative coefficient is 0.993. The numerals of x-axis represent the 59 100 m elevational bands. For example, “1” means 100-200 m a.s.l. The values of y-axis are dissimilarity (βsim). Cluster leaves of different colors represent different cluster assignments.
Elevational patterns of non-volant small mammal species richness in Gyirong Valley, Central Himalaya: Evaluating multiple spatial and environmental drivers
Avian species distribution along elevation at doon valley (foot hills of western Himalayas), Uttarakhand, and its association with vegetation structure
. Journal of Asia-Pacific Biodiversity, 8, 158-167.
The Comprehensive Scientific Expedition to the Qinghai-Xizang Plateau, Chinese Academy ofSciences(1986) The Mammals of Xizang. Science Press, Beijing. (in Chinese)
WangYX (2003) A Complete Checklist of Mammal Species and Subspecies in China: A Taxonomic and Geographic Reference. China Forestry Publishing House, Beijing. (in Chinese)
Past: Paleontological statistics software: Package for education and data analysis
1
2001
... 喜马拉雅山地区非特有种占哺乳动物种数的76.9%, 对总体物种多样性垂直分布格局贡献较大, 其峰值也和总体物种多样性的峰值更为接近.特有种与非特有种分布格局的差异可能是影响各自格局的潜在因素的差异所致.过去的研究表明, 特有种除了受到气候、植被等环境因子的影响, 同时还受到纯空间因子(如面积、中域效应等)的影响; 而非特有种则更易受到气候、植被和生境异质性等空间结构性的环境因子的影响(Hu et al, 2017).特有种分布区更狭窄, 扩散能力较弱, 所以受到限制扩散的环境因素影响; 非特有种活动扩散能力较强, 制约其扩散的往往是气候、生境类型等影响其生理需求的环境因子(Fu et al, 2006; Wu et al, 2013a, b; Rowe et al, 2015).哺乳动物垂直分布幅的分组中, 随着各哺乳动物子集的物种垂直分布幅加大, 物种多样性峰值从低海拔向中海拔地区移动.该现象符合中域效应的预测: 分布幅较广的物种在分布区中间重叠的几率较分布幅狭窄的物种要高; 分布幅较广的物种类群往往会在分布空间的中心形成物种多样性的峰值(Colwell & Hurtt, 1994). ...
Small mammal diversity along elevational gradients in the Philippines: An assessment of patterns and hypotheses
1
2001
... 本研究的多项式回归分析在PAST 2.17 (Hammer et al, 2001; http://folk.uio.no/ohammer/past/)中进行.聚类分析与相异性指数的计算在R 3.4.2 (R Development Core Team, 2017)软件中, 利用“vegan”程序包(Oksanen et al, 2007)和“NbClust”程序包(Charrad et al, 2014)完成. ...
Elevational patterns of non-volant small mammal species richness in Gyirong Valley, Central Himalaya: Evaluating multiple spatial and environmental drivers
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
... ; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
... 喜马拉雅山地区非特有种占哺乳动物种数的76.9%, 对总体物种多样性垂直分布格局贡献较大, 其峰值也和总体物种多样性的峰值更为接近.特有种与非特有种分布格局的差异可能是影响各自格局的潜在因素的差异所致.过去的研究表明, 特有种除了受到气候、植被等环境因子的影响, 同时还受到纯空间因子(如面积、中域效应等)的影响; 而非特有种则更易受到气候、植被和生境异质性等空间结构性的环境因子的影响(Hu et al, 2017).特有种分布区更狭窄, 扩散能力较弱, 所以受到限制扩散的环境因素影响; 非特有种活动扩散能力较强, 制约其扩散的往往是气候、生境类型等影响其生理需求的环境因子(Fu et al, 2006; Wu et al, 2013a, b; Rowe et al, 2015).哺乳动物垂直分布幅的分组中, 随着各哺乳动物子集的物种垂直分布幅加大, 物种多样性峰值从低海拔向中海拔地区移动.该现象符合中域效应的预测: 分布幅较广的物种在分布区中间重叠的几率较分布幅狭窄的物种要高; 分布幅较广的物种类群往往会在分布空间的中心形成物种多样性的峰值(Colwell & Hurtt, 1994). ...
... 同一类群不同动物子集间的物种多样性垂直分布格局模式及其成因有所差异; 相同区域、相同分布格局下, 不同类群或物种子集的物种多样性垂直分布格局的模式及其潜在生态机制也有差异(McCain, 2005, 2009, 2010; Wu et al, 2013a, b; Rowe et al, 2015; Hu et al, 2017).本研究结果与前人的研究结果一致, 即不同物种子集的物种多样性垂直分布格局的模式有所不同.为了更好地研究物种多样性垂直分布格局, 进一步了解全球范围山地生态系统的物种多样性垂直分布格局的模式, 还需要在不同地区、不同尺度开展跨类群的比较研究. ...
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
... ; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
... 喜马拉雅山地区非特有种占哺乳动物种数的76.9%, 对总体物种多样性垂直分布格局贡献较大, 其峰值也和总体物种多样性的峰值更为接近.特有种与非特有种分布格局的差异可能是影响各自格局的潜在因素的差异所致.过去的研究表明, 特有种除了受到气候、植被等环境因子的影响, 同时还受到纯空间因子(如面积、中域效应等)的影响; 而非特有种则更易受到气候、植被和生境异质性等空间结构性的环境因子的影响(Hu et al, 2017).特有种分布区更狭窄, 扩散能力较弱, 所以受到限制扩散的环境因素影响; 非特有种活动扩散能力较强, 制约其扩散的往往是气候、生境类型等影响其生理需求的环境因子(Fu et al, 2006; Wu et al, 2013a, b; Rowe et al, 2015).哺乳动物垂直分布幅的分组中, 随着各哺乳动物子集的物种垂直分布幅加大, 物种多样性峰值从低海拔向中海拔地区移动.该现象符合中域效应的预测: 分布幅较广的物种在分布区中间重叠的几率较分布幅狭窄的物种要高; 分布幅较广的物种类群往往会在分布空间的中心形成物种多样性的峰值(Colwell & Hurtt, 1994). ...
... 同一类群不同动物子集间的物种多样性垂直分布格局模式及其成因有所差异; 相同区域、相同分布格局下, 不同类群或物种子集的物种多样性垂直分布格局的模式及其潜在生态机制也有差异(McCain, 2005, 2009, 2010; Wu et al, 2013a, b; Rowe et al, 2015; Hu et al, 2017).本研究结果与前人的研究结果一致, 即不同物种子集的物种多样性垂直分布格局的模式有所不同.为了更好地研究物种多样性垂直分布格局, 进一步了解全球范围山地生态系统的物种多样性垂直分布格局的模式, 还需要在不同地区、不同尺度开展跨类群的比较研究. ...
... 整合野外调查数据和文献数据, 得出喜马拉雅山地区哺乳动物名录, 根据各物种的分布记录和海拔信息, 获取各物种分布区的海拔上、下限.另外, 基于野外考察观测值, 剔除和校正一些异常值.野外调查的哺乳动物物种编目参考《中国哺乳动物种和亚种分类名录与分布大全》(王应祥, 2003)和《中国哺乳动物多样性及地理分布》(蒋志刚等, 2015).最后, 整体的喜马拉雅山地区哺乳动物物种名录根据The IUCN Red List of Threatened Species (IUCN, 2017)的分类体系重新合并整理, 确保各物种的准确(附录2). ...
Geometric constraints explain much of the species richness pattern in African birds. Proceedings of the National Academy of Sciences,
Avian species distribution along elevation at doon valley (foot hills of western Himalayas), Uttarakhand, and its association with vegetation structure
Biodiversity in Ecosystems: Principles and Case Studies of Different Complexity Levels
1999
A criterion for determining the number of groups in a data set using sum-of-squares clustering
1
1988
... 整合野外调查数据和文献数据, 得出喜马拉雅山地区哺乳动物名录, 根据各物种的分布记录和海拔信息, 获取各物种分布区的海拔上、下限.另外, 基于野外考察观测值, 剔除和校正一些异常值.野外调查的哺乳动物物种编目参考《中国哺乳动物种和亚种分类名录与分布大全》(王应祥, 2003)和《中国哺乳动物多样性及地理分布》(蒋志刚等, 2015).最后, 整体的喜马拉雅山地区哺乳动物物种名录根据The IUCN Red List of Threatened Species (IUCN, 2017)的分类体系重新合并整理, 确保各物种的准确(附录2). ...
The geographical structure of British bird distributions: Diversity, spatial turnover and scale
1
2001
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
珠穆朗玛峰国家级自然保护区鸟类区系及其垂直分布特征
1
2013
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
珠穆朗玛峰国家级自然保护区鸟类区系及其垂直分布特征
1
2013
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
神农架自然保护区非飞行哺乳动物的物种丰富度: 沿海拔梯度的分布格局
2003
神农架自然保护区非飞行哺乳动物的物种丰富度: 沿海拔梯度的分布格局
2003
Species turnover in vertebrate communities along elevational gradients is idiosyncratic and unrelated to species richness
1
2016
... 利用非加权组平均法(UPGMA)对物种阵进行聚类分析.利用Simpson相异性指数(βsim, Simpson, 1943)来衡量各100 m海拔区间哺乳动物物种组成上的差异(Lennon et al, 2001; Mena & Vázquez- Domínguez, 2005; McCain & Beck, 2016).所有100 m海拔区间之间的βsim矩阵完整地反映出喜马拉雅山地区各个海拔带之间的物种组成差异. ...
The mid-domain effect applied to elevational gradients: Species richness of small mammals in Costa Rica
2004
Elevational gradients in diversity of small mammals
The Global 200: A representation approach to conserving the Earth’s most biologically valuable ecoregions
2
1998
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
... 同一类群不同动物子集间的物种多样性垂直分布格局模式及其成因有所差异; 相同区域、相同分布格局下, 不同类群或物种子集的物种多样性垂直分布格局的模式及其潜在生态机制也有差异(McCain, 2005, 2009, 2010; Wu et al, 2013a, b; Rowe et al, 2015; Hu et al, 2017).本研究结果与前人的研究结果一致, 即不同物种子集的物种多样性垂直分布格局的模式有所不同.为了更好地研究物种多样性垂直分布格局, 进一步了解全球范围山地生态系统的物种多样性垂直分布格局的模式, 还需要在不同地区、不同尺度开展跨类群的比较研究. ...
Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants
2005
珠穆朗玛峰国家级自然保护区两栖爬行动物多样性及区系
1
2013
... 利用非加权组平均法(UPGMA)对物种阵进行聚类分析.利用Simpson相异性指数(βsim, Simpson, 1943)来衡量各100 m海拔区间哺乳动物物种组成上的差异(Lennon et al, 2001; Mena & Vázquez- Domínguez, 2005; McCain & Beck, 2016).所有100 m海拔区间之间的βsim矩阵完整地反映出喜马拉雅山地区各个海拔带之间的物种组成差异. ...
珠穆朗玛峰国家级自然保护区两栖爬行动物多样性及区系
1
2013
... 利用非加权组平均法(UPGMA)对物种阵进行聚类分析.利用Simpson相异性指数(βsim, Simpson, 1943)来衡量各100 m海拔区间哺乳动物物种组成上的差异(Lennon et al, 2001; Mena & Vázquez- Domínguez, 2005; McCain & Beck, 2016).所有100 m海拔区间之间的βsim矩阵完整地反映出喜马拉雅山地区各个海拔带之间的物种组成差异. ...
Elevational pattern of bird species richness and its causes along a central Himalaya gradient, China
1
2016
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
Niche filling slows the diversification of Himalayan songbirds
1
2014
... 本研究的多项式回归分析在PAST 2.17 (Hammer et al, 2001; http://folk.uio.no/ohammer/past/)中进行.聚类分析与相异性指数的计算在R 3.4.2 (R Development Core Team, 2017)软件中, 利用“vegan”程序包(Oksanen et al, 2007)和“NbClust”程序包(Charrad et al, 2014)完成. ...
The elevational gradient of species richness: A uniform pattern?
1
1995
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
The relationship among area, elevation, and regional species richness in neotropical birds
1
1997
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
Elevational diversity gradients, biogeography and the structure of montane mammal communities in the intermountain region of North America
2001
Scale effects on the pattern and predictors of small mammal diversity along a local elevational gradient in the Great Basin
2
2015
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
1
1986
... 采用种分布区连续模型确定物种垂直分布范围(Rahbek, 1997; Kratochwil, 1999; McCain, 2009; Hu et al, 2017), 假定物种在它们的分布极限之间连续分布.喜马拉雅山地区海拔6,000 m以上为高山冰雪带, 没有哺乳动物记录, 故我们选择海拔100-6,000 m之间的地区开展研究.按照100 m海拔间隔, 在研究区域的海拔梯度内选取59个取样点(如150 m, 250 m, 350 m…5,850 m, 5,950 m), 然后根据物种分布区连续模型计算出各个海拔取样点的插值物种丰富度(interpolated species richness), 以该插值物种丰富度作为各100 m海拔区间的物种多样性的值.为了避免物种的“丢失”, 将仅有1个分布点的物种垂直海拔分布区进行适当扩展: 以该分布点为中点上下扩展100 m (同时将垂直海拔分布幅不足200 m的物种也扩展到200 m). ...
Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal
2
2002
... 喜马拉雅山脉坐落于青藏高原南缘, 先后被列入“全球200”和世界生物圈保护区(Olson & Dinerstein, 1998; Myers et al, 2000).目前喜马拉雅山地区生物多样性空间分布格局和相关机制的研究仍十分有限, 多数研究仅限于局部地区的单个生物类群, 且主要集中在植物类群(如: Vetaas & Grytnes, 2002; Bhattarai & Vetaas, 2003; Oommen & Shanker, 2005; Acharya et al, 2011a).脊椎动物类群中鸟类(如Acharya et al, 2011b; Price et al, 2014; Joshi & Bhatt, 2015; Pan et al, 2016)、两栖爬行类(如Chettri et al, 2010; 潘虎君等, 2013)和哺乳类(如胡一鸣等, 2014; Hu et al, 2017)虽有一些研究, 但相对较少.本研究的开展不仅有助于认识喜马拉雅山地区哺乳动物区系特征和物种多样性空间分布格局, 还为探讨大尺度范围物种多样性空间分布格局普遍模式和演化机制等宏生态学研究提供数据支撑. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}