1 Key Laboratory of Land Surface Pattern and Simulation, Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 1001012 School of Life Sciences, Sun Yat-sen University, Guangzhou 5102753 University of Chinese Academy of Sciences, Beijing 1000494 Guangdong Institute of Eco-environmental Science & Technology, Guangzhou 510650;
The Tibetan Plateau (TP) harbors numerous seed plants, however, the floristic characteristics and diversity patterns of plants endemic to this region have been rarely studied. Based on several monographs and online databases, we compiled a list of seed plants that exclusively occur on the TP as well as their distribution at the county level. We further explored their characteristics, floristic composition and spatial distribution patterns. We identified 3,764 endemic seed plants belonging to 519 genera and 113 families, 76.3% of which are herbaceous plants. Among them, 15 families (e.g. Asteraceae, Ranunculaceae, Orobanchaceae) and 7 genera (e.g. Pedicularis, Rhododendron, Corydalis) contain over 100 endemic species. Floristic composition analysis indicates that 67.5% of these endemic plants are temperate species. Species diversity declined gradually from the southeast to the northwest with hotspots located within the East Himalaya-Hengduan Mountains. Vertically, most species occurred at intermediate elevations. Understanding floristic characteristics and diversity patterns of Tibetan endemic flora shed light on future studies on the evolutional history and conservation practices in this area.
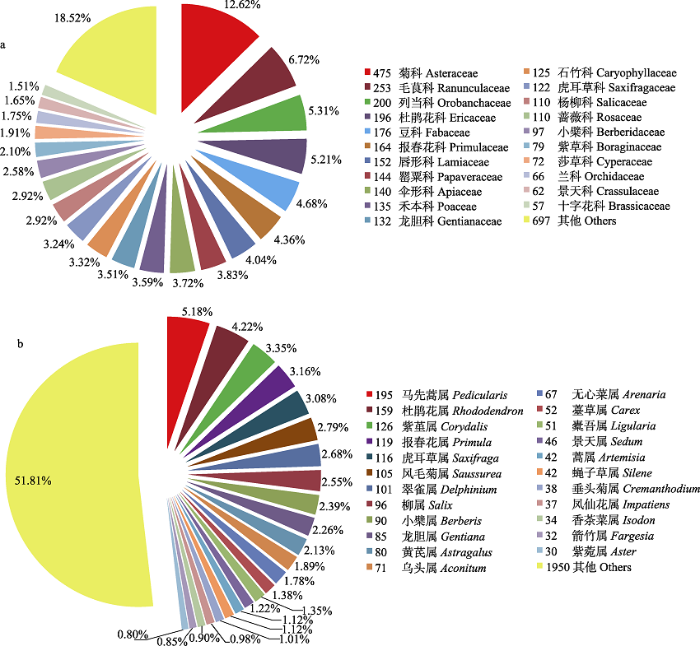
Fig. 1 Percentages of families containing over 50 endemic species (a) and genera containing over 30 endemic species (b) on the Tibetan Plateau. The numbers before family or genus represent number of species they contain.
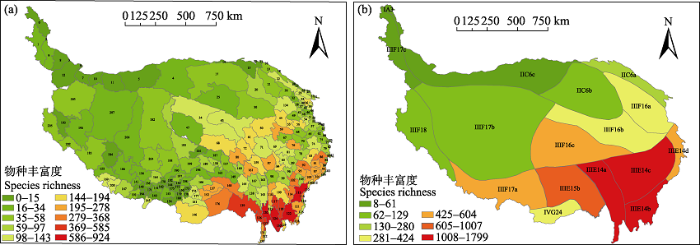
Fig. 2 Diversity patterns of endemic seed plants on the Tibetan Plateau at the county level (a) and floristic level (b). The codes correspond to the name of county and floristic region on the Tibetan Plateau in Appendix 1.
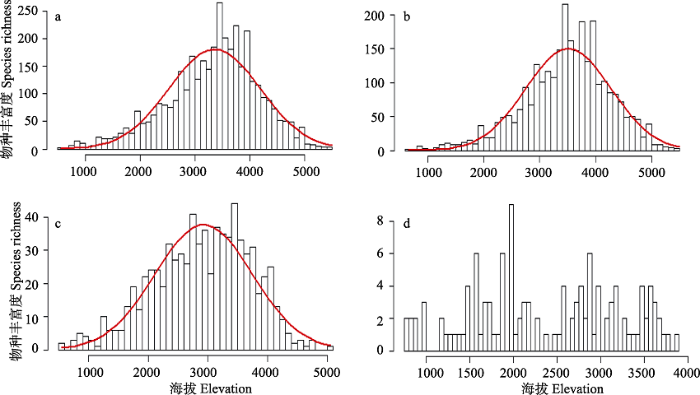
Fig. 3 Species richness of different growth forms of endemic seed plants along elevational gradients. (a), All species; (b), Herb; (c), Shrub; (d), Tree.
Geographic pat¬terns of endemic seed plant genera diversity in China
. Bio¬diversity Science, 19, 414-423. (in Chinese with English abstract)
[陈圣宾, 欧阳志云, 方瑜, 李振基 (2011)
中国种子植物特有属的地理分布格局
. 生物多样性, 19, 414-423.]
[3]
CunYZ, WangXQ (2010)
Plant recolonization in the Hima¬laya from the southeastern Qinghai-Tibetan Plateau: Geo¬graphical isolation contributed to high population differen¬tiation
. Molecular Phylogenetics and Evolution, 56, 972-982.
Dis¬persal routes between biodiversity hotspots in Asia: The case of the mountain genus Tripterospermum (Gentianinae, Gentianaceae) and its close relatives
Plant molecular phy¬logeography in China and adjacent regions: Tracing the ge¬netic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora
. Mo¬lecular Phylogenetics and Evolution, 59, 225-244.
R: A language and environment for statis¬tical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
. (accessed on 2016-11-10)
[20]
RenG, ContiE, SalaminN (2015)
Phylogeny and biogeography of Primula sect. Armerina: Implications for plant evolu¬tion under climate change and the uplift of the Qinghai- Ti¬bet Plateau
ShiYF, LiJJ, LiBY (1998) Uplift and Environmental Changes of Qinghai-Tibetan Plateau in the Late Cenozoic. Guang¬dong Science and Technology Press, Guangzhou. (in Chinese)
The Comprehensive Scientific Expedition to the Qinghai Xi¬zang Plateau, Chinese Academy ofSciences(1993) Vascular Plants of the Hengduan Mountains, Vol. 1. Science Press, Beijing. (in Chinese)
WuZY, ZhouZK, SunH, LiDZ, PengH (2006) The Areal- types of Seed Plants and Their Origin and Differentiation. Yunnan Science and Technology Press, Kunming. (in Chi¬nese)
Patterns of species diversity and phylogenetic structure of vascular plants on the Qinghai- Tibetan Plateau
. Ecology and Evolution, 3, 4584-4595.
[42]
YangFS, LiYF, DingX, WangXQ (2008)
Extensive population expansion of Pedicularis longiflora (Orobanchaceae) on the Qinghai-Tibetan Plateau and its correlation with the Quaternary climate change
Geographical sampling bias in a large distributional database and its effects on species richness-environment models
. Journal of Biogeography, 40, 1415-1426.
[44]
YuHB, ZhangYL, LiuLS, QiW, LiSC, HuZJ (2015)
Com¬bining the least cost path method with population genetic data and species distribution models to identify landscape connectivity during the Late Quaternary in Himalayan hemlock
Towards a comprehensive phylogeny of the large temperate genus Pedicularis (Orobanchaceae), with an emphasis on species from the Himalaya-Hengduan Mountains
. BMC Plant Biology, 15, 176.
[47]
ZhangDC, YeJX, SunH (2016)
Quantitative approaches to identify floristic units and centres of species endemism in the Qinghai-Tibetan Plateau, south-western China
. Journal of Biogeography, 43, 2465-2476.
[48]
ZhangJQ, MengSY, AllenGA, WenJ, RaoGY (2014)
Rapid radiation and dispersal out of the Qinghai-Tibetan Plateau of an alpine plant lineage Rhodiola (Crassulaceae)
. Molecular Phylogenetics and Evolution, 77, 147-158.
Plant recolonization in the Hima¬laya from the southeastern Qinghai-Tibetan Plateau: Geo¬graphical isolation contributed to high population differen¬tiation
1
2010
... 特有性是生物地理学研究的重要内容之一(Kier et al, 2009).对于局限分布于一定区域内的物种来说, 它们的分布格局和多样性不仅与当前地形、气候、土地利用等因素有关(Kreft & Jetz, 2007; 陈圣宾等, 2011), 更与过去的地质变迁和气候波动密切相关(Wiens & Donoghue, 2004; Sandel et al, 2011; Feng et al, 2016).因此, 了解一个地区特有种的多样性格局, 对于认识该地区的植物区系特征、起源、形成与演化具有十分重要的意义.此外, 特有性已成为生物多样性保护研究关注的焦点, 为预测生物多样性热点区域和确定优先保护区提供了重要依据(Huang et al, 2016). ...
Out-of-Tibet: The spatio-temporal evolution of Gentiana (Gentianaceae)
Dis¬persal routes between biodiversity hotspots in Asia: The case of the mountain genus Tripterospermum (Gentianinae, Gentianaceae) and its close relatives
2
2016
... 青藏高原自5,000万年前开始隆升, 尤其在中新世至上新世时期(新近纪, 23-2.6 Ma), 青藏高原经历了快速隆升阶段(施雅风等, 1998).高原隆升不仅改变了大气环流, 促进了亚洲季风气候的产生, 而且逐渐形成了高海拔特殊生境以及复杂的地形.来自分子钟的证据表明: 高原上的很多物种都是这个时期分化形成的, 如报春花属(Ren et al, 2015)、虎耳草属(Gao et al, 2015)、龙胆属(Gentiana) (Favre et al, 2016)和红景天属(Rhodiola) (Zhang et al, 2014)等.这些属的分化不仅与高原快速隆升加速了本地物种的适应性辐射有关, 也与复杂地形促进了物种的隔离分化有关, 可见青藏高原一些大属的形成与高原隆升密切相关.尤其是青藏高原东南部的东喜马拉雅-横断山脉地区(温带成分为主), 一系列高大山脉形成的地理障碍促进了物种的隔离分化和形成, 解释了为什么该地区特有种多样性异常丰富以及温带成分占主导的原因; 此外, 一些耐冷物种从青藏高原向东亚、北美、中亚、欧洲、甚至北极的高山或寒冷地区进行长距离迁移(如Matuszak et al, 2016).因此, 该地区常被称为北温带植物的辐射地以及全球生物多样性的热点地区(吴征镒, 1998; Myers et al, 2000).亚洲季风气候的形成, 造成了该区域暖湿(高原东南部)到冷干(高原西北部)的气候格局, 这种气候格局也深刻影响了青藏高原的物种多样性格局(Mao et al, 2013; Yan et al, 2013). ...
Plant molecular phy¬logeography in China and adjacent regions: Tracing the ge¬netic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora
2
2011
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
... 青藏高原自5,000万年前开始隆升, 尤其在中新世至上新世时期(新近纪, 23-2.6 Ma), 青藏高原经历了快速隆升阶段(施雅风等, 1998).高原隆升不仅改变了大气环流, 促进了亚洲季风气候的产生, 而且逐渐形成了高海拔特殊生境以及复杂的地形.来自分子钟的证据表明: 高原上的很多物种都是这个时期分化形成的, 如报春花属(Ren et al, 2015)、虎耳草属(Gao et al, 2015)、龙胆属(Gentiana) (Favre et al, 2016)和红景天属(Rhodiola) (Zhang et al, 2014)等.这些属的分化不仅与高原快速隆升加速了本地物种的适应性辐射有关, 也与复杂地形促进了物种的隔离分化有关, 可见青藏高原一些大属的形成与高原隆升密切相关.尤其是青藏高原东南部的东喜马拉雅-横断山脉地区(温带成分为主), 一系列高大山脉形成的地理障碍促进了物种的隔离分化和形成, 解释了为什么该地区特有种多样性异常丰富以及温带成分占主导的原因; 此外, 一些耐冷物种从青藏高原向东亚、北美、中亚、欧洲、甚至北极的高山或寒冷地区进行长距离迁移(如Matuszak et al, 2016).因此, 该地区常被称为北温带植物的辐射地以及全球生物多样性的热点地区(吴征镒, 1998; Myers et al, 2000).亚洲季风气候的形成, 造成了该区域暖湿(高原东南部)到冷干(高原西北部)的气候格局, 这种气候格局也深刻影响了青藏高原的物种多样性格局(Mao et al, 2013; Yan et al, 2013). ...
R: A language and environment for statis¬tical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
2016
Phylogeny and biogeography of Primula sect. Armerina: Implications for plant evolu¬tion under climate change and the uplift of the Qinghai- Ti¬bet Plateau
2
2015
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
... 青藏高原自5,000万年前开始隆升, 尤其在中新世至上新世时期(新近纪, 23-2.6 Ma), 青藏高原经历了快速隆升阶段(施雅风等, 1998).高原隆升不仅改变了大气环流, 促进了亚洲季风气候的产生, 而且逐渐形成了高海拔特殊生境以及复杂的地形.来自分子钟的证据表明: 高原上的很多物种都是这个时期分化形成的, 如报春花属(Ren et al, 2015)、虎耳草属(Gao et al, 2015)、龙胆属(Gentiana) (Favre et al, 2016)和红景天属(Rhodiola) (Zhang et al, 2014)等.这些属的分化不仅与高原快速隆升加速了本地物种的适应性辐射有关, 也与复杂地形促进了物种的隔离分化有关, 可见青藏高原一些大属的形成与高原隆升密切相关.尤其是青藏高原东南部的东喜马拉雅-横断山脉地区(温带成分为主), 一系列高大山脉形成的地理障碍促进了物种的隔离分化和形成, 解释了为什么该地区特有种多样性异常丰富以及温带成分占主导的原因; 此外, 一些耐冷物种从青藏高原向东亚、北美、中亚、欧洲、甚至北极的高山或寒冷地区进行长距离迁移(如Matuszak et al, 2016).因此, 该地区常被称为北温带植物的辐射地以及全球生物多样性的热点地区(吴征镒, 1998; Myers et al, 2000).亚洲季风气候的形成, 造成了该区域暖湿(高原东南部)到冷干(高原西北部)的气候格局, 这种气候格局也深刻影响了青藏高原的物种多样性格局(Mao et al, 2013; Yan et al, 2013). ...
The influence of Late Qua¬ternary climate-change velocity on species endemism
1
2011
... 本研究揭示的青藏高原特有种子植物的多样性分布格局, 为探讨高原植物区系的形成演化提供了重要启示.目前, 对生物多样性格局的形成原因更多地从历史和进化的角度来探讨, 而地质活动和过去的气候变化常被认为是改变生物多样性格局的两大历史因素(Wiens & Donoghue, 2004; Sandel et al, 2011).在青藏高原地区, 来自历史生物地理研究的一系列证据表明: 新近纪的高原快速隆升以及第四纪的周期性气候波动促进了植物的辐射分化和多样化(Qiu et al, 2011; Wen et al, 2014), 我们推测这两大因素可能影响了青藏高原的植物分布格局和多样性. ...
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
黄河源头地区植物的区系特征
1
1995
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
长江源区植物区系特征
1
2000
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
长江源区植物区系特征
1
2000
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
2008
2008
澜沧江源区种子植物区系研究
1
2009
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
澜沧江源区种子植物区系研究
1
2009
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
1
1998
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
1
1998
... 从物种垂直分布格局来看, 青藏高原多数植物分布在中海拔地带(3,000-4,000 m), 对高黎贡山和喜马拉雅山脉植物垂直分布格局的研究也有相似的结论(Vetaas & Grytnes, 2002; Wang et al, 2007), 这很可能是由于该海拔带的植物具有更多的时间来积累和保存物种, 即Montane Museum假说, 该假说不仅在龙胆属的多样化研究中得到验证(Favre et al, 2016), 而且普遍存在于华北植物区系的演变和东亚植物的进化历史中(Thorne, 1999; 王荷生, 1999).另外一种可能就是中高海拔范围内的物种具有更高的多样化率, 对19个科(属)的多样性研究发现, 分布在横断山区物种的多样化率更高(Xing & Ree, 2017). ...
1
2010
... 本研究揭示的青藏高原特有种子植物的多样性分布格局, 为探讨高原植物区系的形成演化提供了重要启示.目前, 对生物多样性格局的形成原因更多地从历史和进化的角度来探讨, 而地质活动和过去的气候变化常被认为是改变生物多样性格局的两大历史因素(Wiens & Donoghue, 2004; Sandel et al, 2011).在青藏高原地区, 来自历史生物地理研究的一系列证据表明: 新近纪的高原快速隆升以及第四纪的周期性气候波动促进了植物的辐射分化和多样化(Qiu et al, 2011; Wen et al, 2014), 我们推测这两大因素可能影响了青藏高原的植物分布格局和多样性. ...
1
2010
... 本研究揭示的青藏高原特有种子植物的多样性分布格局, 为探讨高原植物区系的形成演化提供了重要启示.目前, 对生物多样性格局的形成原因更多地从历史和进化的角度来探讨, 而地质活动和过去的气候变化常被认为是改变生物多样性格局的两大历史因素(Wiens & Donoghue, 2004; Sandel et al, 2011).在青藏高原地区, 来自历史生物地理研究的一系列证据表明: 新近纪的高原快速隆升以及第四纪的周期性气候波动促进了植物的辐射分化和多样化(Qiu et al, 2011; Wen et al, 2014), 我们推测这两大因素可能影响了青藏高原的植物分布格局和多样性. ...
2
2006
... 特有性是生物地理学研究的重要内容之一(Kier et al, 2009).对于局限分布于一定区域内的物种来说, 它们的分布格局和多样性不仅与当前地形、气候、土地利用等因素有关(Kreft & Jetz, 2007; 陈圣宾等, 2011), 更与过去的地质变迁和气候波动密切相关(Wiens & Donoghue, 2004; Sandel et al, 2011; Feng et al, 2016).因此, 了解一个地区特有种的多样性格局, 对于认识该地区的植物区系特征、起源、形成与演化具有十分重要的意义.此外, 特有性已成为生物多样性保护研究关注的焦点, 为预测生物多样性热点区域和确定优先保护区提供了重要依据(Huang et al, 2016). ...
... 本研究揭示的青藏高原特有种子植物的多样性分布格局, 为探讨高原植物区系的形成演化提供了重要启示.目前, 对生物多样性格局的形成原因更多地从历史和进化的角度来探讨, 而地质活动和过去的气候变化常被认为是改变生物多样性格局的两大历史因素(Wiens & Donoghue, 2004; Sandel et al, 2011).在青藏高原地区, 来自历史生物地理研究的一系列证据表明: 新近纪的高原快速隆升以及第四纪的周期性气候波动促进了植物的辐射分化和多样化(Qiu et al, 2011; Wen et al, 2014), 我们推测这两大因素可能影响了青藏高原的植物分布格局和多样性. ...
2
2006
... 特有性是生物地理学研究的重要内容之一(Kier et al, 2009).对于局限分布于一定区域内的物种来说, 它们的分布格局和多样性不仅与当前地形、气候、土地利用等因素有关(Kreft & Jetz, 2007; 陈圣宾等, 2011), 更与过去的地质变迁和气候波动密切相关(Wiens & Donoghue, 2004; Sandel et al, 2011; Feng et al, 2016).因此, 了解一个地区特有种的多样性格局, 对于认识该地区的植物区系特征、起源、形成与演化具有十分重要的意义.此外, 特有性已成为生物多样性保护研究关注的焦点, 为预测生物多样性热点区域和确定优先保护区提供了重要依据(Huang et al, 2016). ...
... 本研究揭示的青藏高原特有种子植物的多样性分布格局, 为探讨高原植物区系的形成演化提供了重要启示.目前, 对生物多样性格局的形成原因更多地从历史和进化的角度来探讨, 而地质活动和过去的气候变化常被认为是改变生物多样性格局的两大历史因素(Wiens & Donoghue, 2004; Sandel et al, 2011).在青藏高原地区, 来自历史生物地理研究的一系列证据表明: 新近纪的高原快速隆升以及第四纪的周期性气候波动促进了植物的辐射分化和多样化(Qiu et al, 2011; Wen et al, 2014), 我们推测这两大因素可能影响了青藏高原的植物分布格局和多样性. ...
Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proceedings of the National Academy of Sciences,
2017
Plant diversity and floristic characters of the alpine subnival belt flora in the Hengduan Mountains, SW China
2
2014
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
... 目前, 对中国特有种子植物的特征已经有了较深入的了解(Huang et al, 2011; 黄继红等, 2014), 这为我们理解青藏高原特有种的特征提供了很好的比较材料.与中国特有种的生长型所占比例相比(草本占67.8%), 青藏高原特有种中草本植物的比例更高(76.3%), 这与高海拔环境下较为适合草本植物生存有关.一些大的属如马先蒿属、虎耳草属包含较多的中国特有种(黄继红等, 2014), 其中大部分为青藏高原特有种, 这可能与青藏高原地区是许多类群的分布和分化中心有关(Gao et al, 2015; Yu WB et al, 2015).从属的分布区类型来看, 中国特有种子植物属以热带成分居多(42.0%), 而青藏高原则以温带成分为主(67.5%), 对三江源区、横断山高山冰缘带和滇西北香格里拉植物区系的分析结果也得到温带成分为主的结论(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015). ...
Patterns of species diversity and phylogenetic structure of vascular plants on the Qinghai- Tibetan Plateau
2013
Extensive population expansion of Pedicularis longiflora (Orobanchaceae) on the Qinghai-Tibetan Plateau and its correlation with the Quaternary climate change
2
2008
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
... 目前, 对中国特有种子植物的特征已经有了较深入的了解(Huang et al, 2011; 黄继红等, 2014), 这为我们理解青藏高原特有种的特征提供了很好的比较材料.与中国特有种的生长型所占比例相比(草本占67.8%), 青藏高原特有种中草本植物的比例更高(76.3%), 这与高海拔环境下较为适合草本植物生存有关.一些大的属如马先蒿属、虎耳草属包含较多的中国特有种(黄继红等, 2014), 其中大部分为青藏高原特有种, 这可能与青藏高原地区是许多类群的分布和分化中心有关(Gao et al, 2015; Yu WB et al, 2015).从属的分布区类型来看, 中国特有种子植物属以热带成分居多(42.0%), 而青藏高原则以温带成分为主(67.5%), 对三江源区、横断山高山冰缘带和滇西北香格里拉植物区系的分析结果也得到温带成分为主的结论(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015). ...
Geographical sampling bias in a large distributional database and its effects on species richness-environment models
2013
Com¬bining the least cost path method with population genetic data and species distribution models to identify landscape connectivity during the Late Quaternary in Himalayan hemlock
3
2015
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
Diverse range dynamics and routes of plants on the Tibetan Plateau during the Late Quaternary
2017
Towards a comprehensive phylogeny of the large temperate genus Pedicularis (Orobanchaceae), with an emphasis on species from the Himalaya-Hengduan Mountains
2015
Quantitative approaches to identify floristic units and centres of species endemism in the Qinghai-Tibetan Plateau, south-western China
2016
Rapid radiation and dispersal out of the Qinghai-Tibetan Plateau of an alpine plant lineage Rhodiola (Crassulaceae)
2
2014
... 青藏高原地区拥有丰富的植物物种(吴玉虎, 2008; Zhang et al, 2016), 尤其是高原东南部的东喜马拉雅和横断山脉地区, 不仅是全球生物多样性的热点地区(Myers et al, 2000), 也是我国特有植物最丰富的地区和高山植物区系的中心(吴征镒, 1998; Huang et al, 2016).一直以来, 对青藏高原一些热点地区(如三江源地区、横断山高山冰缘带、滇西北的香格里拉)的植物本底资料和区系特征不断有报道(吴玉虎, 1995, 2000, 2009; Xu et al, 2014; Zhu, 2015), 对青海西藏两省区维管植物的多样性格局也有研究(Mao et al, 2013; Yan et al, 2013).然而, 到目前为止, 对整个青藏高原地区的特有种子植物特征和多样性分布格局还鲜有报道.我们选取了与青藏高原环境密切相关的特有种子植物作为研究对象, 通过物种名录的确定和分布数据的收集, 初步分析了青藏高原特有种子植物的特征和多样性格局, 希望为青藏高原的生物地理学、谱系地理学和生物多样性保护等研究提供参考. ...
... 青藏高原自5,000万年前开始隆升, 尤其在中新世至上新世时期(新近纪, 23-2.6 Ma), 青藏高原经历了快速隆升阶段(施雅风等, 1998).高原隆升不仅改变了大气环流, 促进了亚洲季风气候的产生, 而且逐渐形成了高海拔特殊生境以及复杂的地形.来自分子钟的证据表明: 高原上的很多物种都是这个时期分化形成的, 如报春花属(Ren et al, 2015)、虎耳草属(Gao et al, 2015)、龙胆属(Gentiana) (Favre et al, 2016)和红景天属(Rhodiola) (Zhang et al, 2014)等.这些属的分化不仅与高原快速隆升加速了本地物种的适应性辐射有关, 也与复杂地形促进了物种的隔离分化有关, 可见青藏高原一些大属的形成与高原隆升密切相关.尤其是青藏高原东南部的东喜马拉雅-横断山脉地区(温带成分为主), 一系列高大山脉形成的地理障碍促进了物种的隔离分化和形成, 解释了为什么该地区特有种多样性异常丰富以及温带成分占主导的原因; 此外, 一些耐冷物种从青藏高原向东亚、北美、中亚、欧洲、甚至北极的高山或寒冷地区进行长距离迁移(如Matuszak et al, 2016).因此, 该地区常被称为北温带植物的辐射地以及全球生物多样性的热点地区(吴征镒, 1998; Myers et al, 2000).亚洲季风气候的形成, 造成了该区域暖湿(高原东南部)到冷干(高原西北部)的气候格局, 这种气候格局也深刻影响了青藏高原的物种多样性格局(Mao et al, 2013; Yan et al, 2013). ...
论青藏高原范围与面积
2002
论青藏高原范围与面积
2002
Biogeography of Shangri-la flora in southwestern China
 , 刘林山
, 刘林山

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}