戴云山脉为福建第二大山脉, 中段位于德化县境内, 以戴云山为主峰, 向四周倾斜, 呈层状逐级下降, 最高海拔1,856 m, 最低海拔650 m, 垂直海拔梯度大, 气候与植被垂直变化明显。戴云山区主峰周边建有福建戴云山国家级自然保护区和德化石牛山森林公园两个自然保护地, 拥有丰富的野生动物资源, 是该区重要的生物多样性基因库。自1985年戴云山国家级自然保护区建立以来, 保护区先后多次对区内野生动物资源进行调查, 2003年出版的《福建戴云山自然保护区综合科学考察报告》中记录该区共有哺乳动物8目21科58种, 鸟类17目45科194种(林鹏, 2003 ), 为戴云山山脉闽中地区野生动物监测奠定了基础。2016年保护区夏季鸟类样线调查共发现鸟类29科22属72种, 其中14种为保护区新记录种(林开淼等, 2017 )。然而, 受到调查手段等限制, 一些物种记录并不十分准确, 历史记录数据不能完全反映该区域当前动物资源的现状。尤其是随着人为活动对动物栖息环境的破坏以及气候变化, 一些20世纪80年代以前记录的物种如金钱豹(Panthera pardus )、华南虎(P. tigris huananensis ), 及2003年科考报告中记录的物种如云豹(Neofelis nebulosa )、穿山甲(Manis pentadactyla )等, 在本区域内可能已局域灭绝。
红外相机调查技术具有“准确性、长期性、非损伤性”等优点, 已成为野生动物本底资源调查和生物多样性监测的重要手段(李晟等, 2014 )。本研究于2017年2月至2018年3月, 对福建戴云山国家级自然保护区和德化石牛山森林公园内的野生哺乳动物和鸟类开展了为期1年的红外相机调查, 以了解该区野生动物资源现状, 掌握区内大中型哺乳动物与地面活动鸟类的种类、相对多度与分布, 为该区域大中型哺乳动物与地栖鸟类的研究和管理提供本底资料, 填补闽中地区这两个野生动物类群观测的空白。
1 研究方法
1.1 研究区域概况
在戴云山山脉中段戴云山区内, 选择戴云山国家级自然保护区(以下简称戴云山保护区)和德化石牛山森林公园两个区域开展野外红外相机监测。戴云山保护区总面积为134.72 km2 , 范围在118°05°22°°- 118°20°15°° E, 25°38°07°°-25°43°40°° N之间, 海拔跨度650-1,856 m; 石牛山森林公园面积86.82 km2 , 主峰位于水口镇, 海拔跨度300-1,782 m, 地理位置介于118°24°3.3°°-118°28°40°° E, 25°36°59°°-25°40°50°° N之间。戴云山区地处亚热带海洋性季风气候带, 是东南沿海典型的山地森林生态系统, 植被类型为亚热带常绿硬叶林、亚热带常绿阔叶林和大陆东南沿海分布最南端、面积最大、保存最完好的黄山松(Pinus taiwanensis )林, 是福建省重要的生物多样性基因库、水源涵养区、生态安全屏障和珍稀野生动植物的生物安全岛屿, 对于福建省中部生物多样性保护、生态安全保障、水源涵养、调节气候、净化空气、科研教学实习等方面具有重要意义(林鹏, 2003 )。
1.2 红外相机布设方案
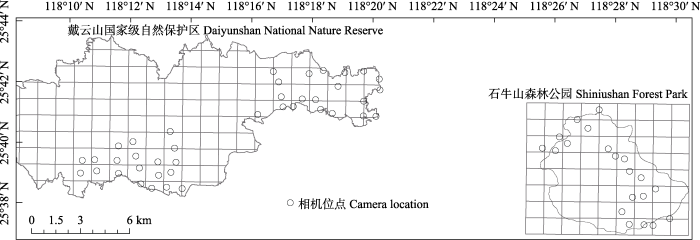
在戴云山保护区设置2个观测样地, 石牛山森林公园内设置1个样地。每个样地面积20 km2 (图1 ), 划分成20个1 km × 1 km的网格。在每个网格中心区域选择合适位置放置1台红外相机, 相邻网格的两台相机间距不少于500 m。共布设60台红外相机, 型号为猎科LTL 6511。戴云山保护区、石牛山森林公园布设的调查点位海拔跨度分别为765-1,630 m、649-1,355 m, 基本涵盖了戴云山脉中部不同生境, 包括常绿阔叶林、落叶阔叶林、竹林、针阔混交林、针叶林、草甸。
图1 福建戴云山国家级自然保护区与石牛山森林公园2017-2018年红外相机监测位点分布图
Fig. 1 Camera-trapping survey stations in Daiyunshan National Nature Reserve and Shiniushan Forest Park of Fujian Province during 2017-2018.
红外相机固定于粗细合适的树干上, 距离地面高度约60 cm。对每台相机进行定位和编号, 同时记录生境信息。相机镜头与地面平行, 避免阳光直射镜头。相机设置为24 h工作模式, 拍摄时间间隔为15 s, 灵敏度设为“中”。每台相机配置南孚5号碱性电池12节, SCAN 32 G储存卡1张。电池、卡每3个月更换1次, 下载回收数据; 如果发现不工作或存储卡已经写满的相机, 立即更换, 同时清理相机视场内的杂草、枯枝等。监测研究时间为2017年2月至2018年3月, 期间共收集照片4次。
1.3 数据整理与分析
整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定。哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 )。夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类。定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片。根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ):
RAI = Ai /N × 1000
其中, Ai 代表第i 类(i = 1, 2, …)动物出现的有效照片数, N 为总有效相机工作日。
2 结果
红外相机一年内累计监测19,924个相机工作日, 共获得有效照片25,286张, 其中鸟类4,216张、哺乳动物10,245张、家畜10,825张。独立有效照片3,499张, 占总有效照片数的13.83%, 其中野生鸟类独立有效照片1,040张, 占鸟类总有效照片数的24.67%; 野生哺乳动物2,054张, 占野生哺乳动物总有效照片数的20.04%。获得可准确鉴定的野生动物42种, 隶属9目21科38属, 其中鸟类5目11科21属24种, 哺乳动物4目10科17属18种。属于国家I级重点保护的野生动物2种, 即黄腹角雉(Tragopan caboti )、白颈长尾雉(Syrmaticus ellioti ); II级重点保护的野生动物5种, 即中华鬣羚(Capricornis milneedwardsii )、小灵猫(Viverricula indica )、黄喉貂(Martes flavigula )、白鹇(Lophura nythemera )、褐翅鸦鹃(Centropus sinens- is ) (附录1, 附录2)。
相对多度指数排名前5的鸟类依次为白鹇(RAI = 33.13)、紫啸鸫(Myophonus caeruleus , RAI = 7.28)、黑领噪鹛(Garrulax pectoralis , RAI = 4.97)、白眉山鹧鸪(Arborophila gingica , RAI = 2.71)、灰胸竹鸡(Bambusicola thoracica , RAI = 0.70)。国家I级重点保护野生动物黄腹角雉相对多度指数在鸟类中排名第6位(RAI = 0.55), 白颈长尾雉最低(RAI = 0.05) (附录1)。野生哺乳动物相对多度指数排名前5的物种分别为小麂(Muntiacus reevesi , RAI = 28.81)、野猪(Sus scrofa , RAI = 18.42)、赤麂(Muntiacus vaginalis , RAI = 14.71)、赤腹松鼠(Callosciurus erythraeus , RAI = 9.94)、猪獾(Arctonyx collaris , RAI = 4.12)。中华鬣羚(RAI = 0.35)、小灵猫(RAI = 0.30)相对多度指数较低(附录1, 附录2)。
3 讨论
通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 )。红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种。红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 )。与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种。开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足。
对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 )。鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低。松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到。该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到。结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀。另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证。在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 )。
本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致。同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究。但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少。毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 )。同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 )。戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础。赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 )。两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别。尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争。但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动。随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.)。
中华鬣羚是国家II级重点保护野生动物, 主要分布在树林茂密、地势险峻的地方, 随着多年来栖息地的破坏和人类的滥捕滥杀, 野生种已经较难发现。本次调查共有5个相机位点拍摄到中华鬣羚, 其中4个位点位于石牛山森林公园样区, 1个位点在戴云山国家级自然保护区样区, 表明该动物在德化戴云山区分布范围较为狭窄。另外石牛山森林公园样区还拍摄到黄腹角雉、白颈长尾雉两种国家I级保护动物, 由此可见戴云山区较多的珍稀保护动物并未包含在国家级自然保护区之内。因此, 石牛山森林公园中珍稀濒危物种的栖息地保护极为重要, 然而近年来该区所在政府为了发展县域旅游, 在其周边地区开展索道、玻璃栈道等旅游设施建设开发, 导致野生动物栖息地的破碎化, 而人为旅游活动极大可能会造成野生保护动物的消减或者灭绝。鉴于戴云山区区位的特殊性及对气候变化的敏感性, 未来应该积极推动戴云山国家公园试点和绿色发展, 探讨生态旅游和物种保护并存机制, 实现野生动物在环境变化和人为影响下的永续生存。
人为活动对野生哺乳动物的影响较大, 从拍摄到的照片分析来看, 戴云山区内周边社区存在家畜如牛、羊、狗等进入保护区内的现象, 较严重破坏野生动物栖息环境并争夺食物, 另外打猎、旅游、挖笋等人为活动较多, 需要加强进山人员的管理和监测。因此, 如何减少当地社区居民、家畜、家犬等活动对野生动物产生的直接或间接影响, 值得保护区管理人员深入思索。本研究所获得的数据和信息较为全面地反映了戴云山区哺乳动物和鸟类的本底信息, 为后续的保护管理和长期监测提供了数据支持和指导。目前, 戴云山区森林红外相机监测仍然在进行中, 更多的深入研究有待获取更多的后续监测数据。
附录 Supplementary Material
附录1 戴云山森林红外相机记录的野生哺乳动物和鸟类名录
Appendix 1 Wild mammals and birds recorded by camera trapping in Daiyunshan Forests
http://www.biodiversity-science.net/fileup/PDF/2018216-1.pdf
附录2 戴云山森林红外相机拍摄的野生哺乳动物和鸟类物种坐标记录
Appendix 2 Coordinate record of wild mammals and birds recorded by camera trapping in Daiyunshan Forests
http://www.biodiversity-science.net/fileup/PDF/2018216-2.xlsx
附录3 戴云山森林红外相机拍摄到的重要哺乳动物和鸟类照片
Appendix 3 Important mammals and birds recorded by camera trapping in Daiyunshan Forests
http://www.biodiversity-science.net/fileup/PDF/2018216-3.pdf
致谢: 本研究野外调查得到福建戴云山国家级自然保护区管理局陈文伟、蔡孝星、陈亚婷、林有星等同志的支持, 在此一并致谢!
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bai DF Chen Y Li JS Tao Q Wang LF Piao Y Shi K 2018 ) Mammal diversity in Shangyong Nature Reserve, Xishuangbanna, Yunnan Province
. Biodiversity Science , 26 , 75 -78 . (in Chinese with English abstract)
[本文引用: 1]
[白德凤 , 陈颖 , 李俊松 , 陶庆 , 王利繁 , 飘优 , 时坤 (2018 ) 西双版纳尚勇自然保护区哺乳动物物种多样性
. 生物多样性 , 26 , 75 -78 .]
[本文引用: 1]
[2]
Cai YS Gong YN Lu XL Xiao ZS Wang XC 2016 ) Camera trap survey of mammal diversity in Nanling Forests
. Ecological Science , 35 (2 ), 57 -61 . (in Chinese with English abstract)
[本文引用: 1]
[蔡玉生 , 龚粤宁 , 卢学理 , 肖治术 , 王新财 (2016 ) 南岭森林哺乳动物多样性的红外相机监测
. 生态科学 , 35 (2 ), 57 -61 .]
[本文引用: 1]
[3]
Chen W Hu JC 2012 ) Seasonal differences in microhabitat use by tufted deer (Elaphodus cephalophus ) in Tangjiahe Nature Reserve
. Acta Theriologica Sinica , 32 , 188 -192 .
[本文引用: 1]
[4]
Gao H Ma ZQ Teng LW Liu ZS Zhao C Sun YJ Guo ZH 2017 ) Tufted deer (Elaphodus cephalophus ) was discovered in the Liupan Mountain National Nature Reserve, Ningxia Hui Autonomous Region, China
. Chinese Journal of Zoology , 52 , 1066 -1068 . (in Chinese with English abstract)
[本文引用: 1]
[高惠 , 马忠其 , 滕丽微 , 刘振生 , 赵唱 , 孙玉姣 , 郭志宏 (2017 ) 宁夏回族自治区六盘山国家级自然保护区发现毛冠鹿
. 动物学杂志 , 52 , 1066 -1068 .]
[本文引用: 1]
[5]
Gordon CE Feit A Grüber J Letnic M 2015 ) Mesopredator suppression by an apex predator alleviates the risk of predation perceived by small prey
. Proceedings of the Royal Society B: Biological Sciences , 282 , 1 -8 .
[本文引用: 1]
[6]
Jiang ZG Liu SY Wu Y Jiang XL Zhou KY 2017 ) China’s mammal diversity (2nd edition)
. Biodiversity Science , 25 , 886 -895 . (in Chinese with English abstract)
[本文引用: 1]
[蒋志刚 , 刘少英 , 吴毅 , 蒋学龙 , 周开亚 (2017 ) 中国哺乳动物多样性(第2版)
. 生物多样性 , 25 , 886 -895 .]
[本文引用: 1]
[7]
Li J Liu F Ye LX Liu SL Peng H Li DQ 2018 ) Camera-trapping survey of the diversity of mammals and birds in Fengyang Mountain of Zhejiang Province, China
. Acta Theriologica Sinica , 38 , 95 -103 . (in Chinese with English abstract)
[本文引用: 1]
[李佳 , 刘芳 , 叶立新 , 刘胜龙 , 彭辉 , 李迪强 (2018 ) 利用红外相机调查浙江省凤阳山兽类和鸟类多样性
. 兽类学报 , 38 , 95 -103 .]
[本文引用: 1]
[8]
Li Q Wu JG Kou XJ Feng LM 2013 ) Applications of camera trap in wildlife population ecology
. Chinese Journal of Applied Ecology , 24 , 947 -955 . (in Chinese with English abstract)
[本文引用: 1]
[李勤 , 邬建国 , 寇晓军 , 冯利民 (2013 ) 相机陷阱在野生动物种群生态学中的应用
. 应用生态学报 , 24 , 947 -955 .]
[本文引用: 1]
[9]
Li S Wang DJ Xiao ZS Li XH Wang TM Feng LM Wang Y 2014 ) Camera-trapping in wildlife research and conservation in China: Review and outlook
. Biodiversity Science , 22 , 685 -695 . (in Chinese with English abstract)
[本文引用: 1]
[李晟 , 王大军 , 肖治术 , 李欣海 , 王天明 , 冯利民 , 王云 (2014 ) 红外相机技术在我国野生动物研究与保护中的应用与前景
. 生物多样性 , 22 , 685 -695 .]
[本文引用: 1]
[10]
Lin KM Huang YQ Chen WW Xu JG Hong LB Chen B 2017 ) Community structure and biodiversity of summer birds in Dehua County
. Inner Mongolia Forestry Investigation and Design , 40 (2 ), 51 -55 . (in Chinese with English abstract)
[本文引用: 2]
[林开淼 , 黄雅琼 , 陈文伟 , 徐建国 , 洪丽彬 , 陈斌 (2017 ) 德化县夏季鸟类群落结构及多样性调查
. 内蒙古林业调查设计 , 40 (2 ), 51 -55 .]
[本文引用: 2]
[11]
Lin P 2003 ) Comprehensive Scientific Investigation Report of Daiyunshan Nature Reserve in Fujian . Xiamen University Press , Xiamen . (in Chinese)
[本文引用: 6]
[林鹏 (2003 ) 福建戴云山自然保护区综合科学考察报告 . 厦门大学出版社 , 厦门 .]
[本文引用: 6]
[12]
Lu QB Yu JA Gao X Yang XY Zhou Q Zhang LB Zhang SY 2007 ) Winter habitat selection of Reeves’ muntjac and wild boars in the Qingliangfeng Mountains
. Acta Theriologica Sinica , 27 , 45 -52 . (in Chinese with English abstract)
[本文引用: 1]
[鲁庆彬 , 于江傲 , 高欣 , 杨仙玉 , 周圻 , 张良斌 , 章叔岩 (2007 ) 冬季清凉峰山区小麂和野猪的生境选择及差异
. 兽类学报 , 27 , 45 -52 .]
[本文引用: 1]
[13]
Shan XN Shi YF Zhang HJ Xu CH Li J Zheng AL 2004 ) mtDNA sequences variability of Munliacus reevesi , M. muntjak and M . crinifrons as well as the related phylogenetic relationship
. Chinese Journal of Zoology , 39 , 35 -39 . (in Chinese with English abstract)
[本文引用: 1]
[单祥年 , 施燕峰 , 张海军 , 徐春宏 , 李健 , 郑爱玲 (2004 ) 小麂、赤麂、黑麂mtDNA序列变异性及反映的进化关系
. 动物学杂志 , 39 , 35 -39 .]
[本文引用: 1]
[14]
Sheng HL Lu HJ 1982 ) Distribution, habits and resource status of the tufted deer (Elaphodus cepholophus )
. Acta Zoologica Sinica , 28 , 307 -311 . (in Chinese with English abstract)
[本文引用: 2]
[盛和林 , 陆厚基 (1982 ) 毛冠鹿的分布、资源和习性
. 动物学报 , 28 , 307 -311 .]
[本文引用: 2]
[15]
Shi XG Hu Q Li JQ Tang Z Yang J Li WJ Shen XL Li S 2017 ) Camera-trapping surveys of the mammal and bird diversity in Wolong National Nature Reserve, Sichuan Province
. Biodiversity Science , 25 , 1131 -1136 . (in Chinese with English abstract)
[本文引用: 1]
[施小刚 , 胡强 , 李佳琦 , 唐卓 , 杨建 , 李文静 , 申小莉 , 李晟 (2017 ) 利用红外相机调查四川卧龙国家级自然保护区鸟兽多样性
. 生物多样性 , 25 , 1131 -1136 .]
[本文引用: 1]
[16]
Wang QS 1990 ) The Mammal Fauna of Anhui . Anhui Science and Technology Publishing House , Hefei . (in Chinese)
[本文引用: 1]
[王岐山 (1990 ) 安徽兽类志 . 安徽科学技术出版社 , 合肥 .]
[本文引用: 1]
[17]
Yang F Hu SX Wang JJ Guo K Jiang GS 2018 ) Spatial interaction among free-ranging ungulates by season at the microhabitat scale
. Chinese Journal of Wildlife , 39 , 224 -230 . (in Chinese with English abstract)
[本文引用: 1]
[杨帆 , 胡素贤 , 王晶晶 , 郭楷 , 姜广顺 (2018 ) 微生境尺度下有蹄类动物空间季节利用互作关系研究
. 野生动物学报 , 39 , 224 -230 .]
[本文引用: 1]
[18]
Zheng GM 2017 ) A List of Classification and Distribution of Birds in China , 3rd edn. Science Press , Beijing . (in Chinese)
[本文引用: 1]
[郑光美 (2017 ) 中国鸟类分类与分布名录(第三版) . 科学出版社 , 北京 .]
[本文引用: 1]
[19]
Zhou DL 1997 ) A list of mammal species from Fujian, China
. Wuyi Science Journal , 13 , 256 -261 . (in Chinese with English abstract)
[本文引用: 1]
[周冬良 (1997 ) 福建哺乳动物种类名录
. 武夷科学 , 13 , 256 -261 .]
[本文引用: 1]
西双版纳尚勇自然保护区哺乳动物物种多样性
1
2018
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
西双版纳尚勇自然保护区哺乳动物物种多样性
1
2018
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
南岭森林哺乳动物多样性的红外相机监测
1
2016
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
南岭森林哺乳动物多样性的红外相机监测
1
2016
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
Seasonal differences in microhabitat use by tufted deer (Elaphodus cephalophus ) in Tangjiahe Nature Reserve
1
2012
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
宁夏回族自治区六盘山国家级自然保护区发现毛冠鹿
1
2017
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
宁夏回族自治区六盘山国家级自然保护区发现毛冠鹿
1
2017
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
Mesopredator suppression by an apex predator alleviates the risk of predation perceived by small prey
1
2015
... 对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 ).鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低.松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
中国哺乳动物多样性(第2版)
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
中国哺乳动物多样性(第2版)
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
利用红外相机调查浙江省凤阳山兽类和鸟类多样性
1
2018
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
利用红外相机调查浙江省凤阳山兽类和鸟类多样性
1
2018
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
相机陷阱在野生动物种群生态学中的应用
1
2013
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
相机陷阱在野生动物种群生态学中的应用
1
2013
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
红外相机技术在我国野生动物研究与保护中的应用与前景
1
2014
... 红外相机调查技术具有“准确性、长期性、非损伤性”等优点, 已成为野生动物本底资源调查和生物多样性监测的重要手段(李晟等, 2014 ).本研究于2017年2月至2018年3月, 对福建戴云山国家级自然保护区和德化石牛山森林公园内的野生哺乳动物和鸟类开展了为期1年的红外相机调查, 以了解该区野生动物资源现状, 掌握区内大中型哺乳动物与地面活动鸟类的种类、相对多度与分布, 为该区域大中型哺乳动物与地栖鸟类的研究和管理提供本底资料, 填补闽中地区这两个野生动物类群观测的空白. ...
红外相机技术在我国野生动物研究与保护中的应用与前景
1
2014
... 红外相机调查技术具有“准确性、长期性、非损伤性”等优点, 已成为野生动物本底资源调查和生物多样性监测的重要手段(李晟等, 2014 ).本研究于2017年2月至2018年3月, 对福建戴云山国家级自然保护区和德化石牛山森林公园内的野生哺乳动物和鸟类开展了为期1年的红外相机调查, 以了解该区野生动物资源现状, 掌握区内大中型哺乳动物与地面活动鸟类的种类、相对多度与分布, 为该区域大中型哺乳动物与地栖鸟类的研究和管理提供本底资料, 填补闽中地区这两个野生动物类群观测的空白. ...
德化县夏季鸟类群落结构及多样性调查
2
2017
... 戴云山脉为福建第二大山脉, 中段位于德化县境内, 以戴云山为主峰, 向四周倾斜, 呈层状逐级下降, 最高海拔1,856 m, 最低海拔650 m, 垂直海拔梯度大, 气候与植被垂直变化明显.戴云山区主峰周边建有福建戴云山国家级自然保护区和德化石牛山森林公园两个自然保护地, 拥有丰富的野生动物资源, 是该区重要的生物多样性基因库.自1985年戴云山国家级自然保护区建立以来, 保护区先后多次对区内野生动物资源进行调查, 2003年出版的《福建戴云山自然保护区综合科学考察报告》中记录该区共有哺乳动物8目21科58种, 鸟类17目45科194种(林鹏, 2003 ), 为戴云山山脉闽中地区野生动物监测奠定了基础.2016年保护区夏季鸟类样线调查共发现鸟类29科22属72种, 其中14种为保护区新记录种(林开淼等, 2017 ).然而, 受到调查手段等限制, 一些物种记录并不十分准确, 历史记录数据不能完全反映该区域当前动物资源的现状.尤其是随着人为活动对动物栖息环境的破坏以及气候变化, 一些20世纪80年代以前记录的物种如金钱豹(Panthera pardus )、华南虎(P. tigris huananensis ), 及2003年科考报告中记录的物种如云豹(Neofelis nebulosa )、穿山甲(Manis pentadactyla )等, 在本区域内可能已局域灭绝. ...
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
德化县夏季鸟类群落结构及多样性调查
2
2017
... 戴云山脉为福建第二大山脉, 中段位于德化县境内, 以戴云山为主峰, 向四周倾斜, 呈层状逐级下降, 最高海拔1,856 m, 最低海拔650 m, 垂直海拔梯度大, 气候与植被垂直变化明显.戴云山区主峰周边建有福建戴云山国家级自然保护区和德化石牛山森林公园两个自然保护地, 拥有丰富的野生动物资源, 是该区重要的生物多样性基因库.自1985年戴云山国家级自然保护区建立以来, 保护区先后多次对区内野生动物资源进行调查, 2003年出版的《福建戴云山自然保护区综合科学考察报告》中记录该区共有哺乳动物8目21科58种, 鸟类17目45科194种(林鹏, 2003 ), 为戴云山山脉闽中地区野生动物监测奠定了基础.2016年保护区夏季鸟类样线调查共发现鸟类29科22属72种, 其中14种为保护区新记录种(林开淼等, 2017 ).然而, 受到调查手段等限制, 一些物种记录并不十分准确, 历史记录数据不能完全反映该区域当前动物资源的现状.尤其是随着人为活动对动物栖息环境的破坏以及气候变化, 一些20世纪80年代以前记录的物种如金钱豹(Panthera pardus )、华南虎(P. tigris huananensis ), 及2003年科考报告中记录的物种如云豹(Neofelis nebulosa )、穿山甲(Manis pentadactyla )等, 在本区域内可能已局域灭绝. ...
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
6
2003
... 戴云山脉为福建第二大山脉, 中段位于德化县境内, 以戴云山为主峰, 向四周倾斜, 呈层状逐级下降, 最高海拔1,856 m, 最低海拔650 m, 垂直海拔梯度大, 气候与植被垂直变化明显.戴云山区主峰周边建有福建戴云山国家级自然保护区和德化石牛山森林公园两个自然保护地, 拥有丰富的野生动物资源, 是该区重要的生物多样性基因库.自1985年戴云山国家级自然保护区建立以来, 保护区先后多次对区内野生动物资源进行调查, 2003年出版的《福建戴云山自然保护区综合科学考察报告》中记录该区共有哺乳动物8目21科58种, 鸟类17目45科194种(林鹏, 2003 ), 为戴云山山脉闽中地区野生动物监测奠定了基础.2016年保护区夏季鸟类样线调查共发现鸟类29科22属72种, 其中14种为保护区新记录种(林开淼等, 2017 ).然而, 受到调查手段等限制, 一些物种记录并不十分准确, 历史记录数据不能完全反映该区域当前动物资源的现状.尤其是随着人为活动对动物栖息环境的破坏以及气候变化, 一些20世纪80年代以前记录的物种如金钱豹(Panthera pardus )、华南虎(P. tigris huananensis ), 及2003年科考报告中记录的物种如云豹(Neofelis nebulosa )、穿山甲(Manis pentadactyla )等, 在本区域内可能已局域灭绝. ...
... 在戴云山山脉中段戴云山区内, 选择戴云山国家级自然保护区(以下简称戴云山保护区)和德化石牛山森林公园两个区域开展野外红外相机监测.戴云山保护区总面积为134.72 km2 , 范围在118°05°22°°- 118°20°15°° E, 25°38°07°°-25°43°40°° N之间, 海拔跨度650-1,856 m; 石牛山森林公园面积86.82 km2 , 主峰位于水口镇, 海拔跨度300-1,782 m, 地理位置介于118°24°3.3°°-118°28°40°° E, 25°36°59°°-25°40°50°° N之间.戴云山区地处亚热带海洋性季风气候带, 是东南沿海典型的山地森林生态系统, 植被类型为亚热带常绿硬叶林、亚热带常绿阔叶林和大陆东南沿海分布最南端、面积最大、保存最完好的黄山松(Pinus taiwanensis )林, 是福建省重要的生物多样性基因库、水源涵养区、生态安全屏障和珍稀野生动植物的生物安全岛屿, 对于福建省中部生物多样性保护、生态安全保障、水源涵养、调节气候、净化空气、科研教学实习等方面具有重要意义(林鹏, 2003 ). ...
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
... 对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 ).鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低.松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
... )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
6
2003
... 戴云山脉为福建第二大山脉, 中段位于德化县境内, 以戴云山为主峰, 向四周倾斜, 呈层状逐级下降, 最高海拔1,856 m, 最低海拔650 m, 垂直海拔梯度大, 气候与植被垂直变化明显.戴云山区主峰周边建有福建戴云山国家级自然保护区和德化石牛山森林公园两个自然保护地, 拥有丰富的野生动物资源, 是该区重要的生物多样性基因库.自1985年戴云山国家级自然保护区建立以来, 保护区先后多次对区内野生动物资源进行调查, 2003年出版的《福建戴云山自然保护区综合科学考察报告》中记录该区共有哺乳动物8目21科58种, 鸟类17目45科194种(林鹏, 2003 ), 为戴云山山脉闽中地区野生动物监测奠定了基础.2016年保护区夏季鸟类样线调查共发现鸟类29科22属72种, 其中14种为保护区新记录种(林开淼等, 2017 ).然而, 受到调查手段等限制, 一些物种记录并不十分准确, 历史记录数据不能完全反映该区域当前动物资源的现状.尤其是随着人为活动对动物栖息环境的破坏以及气候变化, 一些20世纪80年代以前记录的物种如金钱豹(Panthera pardus )、华南虎(P. tigris huananensis ), 及2003年科考报告中记录的物种如云豹(Neofelis nebulosa )、穿山甲(Manis pentadactyla )等, 在本区域内可能已局域灭绝. ...
... 在戴云山山脉中段戴云山区内, 选择戴云山国家级自然保护区(以下简称戴云山保护区)和德化石牛山森林公园两个区域开展野外红外相机监测.戴云山保护区总面积为134.72 km2 , 范围在118°05°22°°- 118°20°15°° E, 25°38°07°°-25°43°40°° N之间, 海拔跨度650-1,856 m; 石牛山森林公园面积86.82 km2 , 主峰位于水口镇, 海拔跨度300-1,782 m, 地理位置介于118°24°3.3°°-118°28°40°° E, 25°36°59°°-25°40°50°° N之间.戴云山区地处亚热带海洋性季风气候带, 是东南沿海典型的山地森林生态系统, 植被类型为亚热带常绿硬叶林、亚热带常绿阔叶林和大陆东南沿海分布最南端、面积最大、保存最完好的黄山松(Pinus taiwanensis )林, 是福建省重要的生物多样性基因库、水源涵养区、生态安全屏障和珍稀野生动植物的生物安全岛屿, 对于福建省中部生物多样性保护、生态安全保障、水源涵养、调节气候、净化空气、科研教学实习等方面具有重要意义(林鹏, 2003 ). ...
... 通过一年多的红外相机监测, 拍摄到大量的野生动物照片和视频(附录3), 可鉴定哺乳动物18种、鸟类24种, 分别占戴云山保护区及周边地区已记录哺乳动物的31.03% (林鹏, 2003 )和鸟类的11.53% (林开淼等, 2017 ).红外相机拍摄到大量小型啮齿类动物, 如鼠类, 它们主要在夜间活动, 且动作迅速, 体型较小, 外形相似, 红外相机很难捕捉到可辨认物种的清晰照片, 本次调查能鉴别的只有板齿鼠(Bandicota indica )和白腹巨鼠(Leopoldmys edwardsi )两种.红外相机监测适合于拍摄地面活动的哺乳动物和林下鸟类, 对营洞穴生活的小型哺乳动物和体型较小的林冠活动鸟类来说则不是一种有效的调查手段(李佳等, 2018 ).与保护区历史记录鸟类名录相比, 本次调查拍摄到的白颈长尾雉是保护区新记录种.开展鸟类多样性专项调查, 应该采用多种调查方法如样线法来补充红外相机调查技术的不足. ...
... 对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 ).鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低.松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
... )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
冬季清凉峰山区小麂和野猪的生境选择及差异
1
2007
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
冬季清凉峰山区小麂和野猪的生境选择及差异
1
2007
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
小麂、赤麂、黑麂mtDNA序列变异性及反映的进化关系
1
2004
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
小麂、赤麂、黑麂mtDNA序列变异性及反映的进化关系
1
2004
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
毛冠鹿的分布、资源和习性
2
1982
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
... ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
毛冠鹿的分布、资源和习性
2
1982
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
... ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
利用红外相机调查四川卧龙国家级自然保护区鸟兽多样性
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
利用红外相机调查四川卧龙国家级自然保护区鸟兽多样性
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
1
1990
... 对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 ).鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低.松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
1
1990
... 对比戴云山保护区以往的记录(林鹏, 2003 )和现存标本, 本次调查拍摄到的灵猫科动物中, 果子狸(Paguma larvata )较为常见, 小灵猫拍到6次, 历史有记录的大灵猫(Viverra zibetha )没有拍到, 大灵猫在20世纪80年代初已经十分稀少(王岐山, 1990 ).鼬科有猪獾、黄喉貂、鼬獾(Melogale moschata )、黄鼬(Mustela sibirica ) 4种, 黄喉貂相对多度最低.松鼠科有赤腹松鼠和倭花鼠(Tamiops maritimus ), 历史记录的泊氏长吻松鼠(Dremomys pernyi )(林鹏, 2003 )没有拍到.该地区历史上分布的一些重要珍稀物种如豹猫(Prionailurus bengalensis )、金猫(Catopuma temminckii )、云豹、金钱豹和华南虎等也未拍摄到.结合保护区内当地居民的观察和保护区近年来的样线调查记录, 这些物种目前种群状况不明, 可能种群密度极低, 或有些物种如华南虎可能已局域绝灭, 其主要原因可归结于该地区栖息地片断化、丧失以及近期历史上的过度猎杀.另外, 由于本次调查主要集中在戴云山区的南部和东部, 北部和西部的红外相机监测点较少, 且监测时间较短, 是否仍然存在云豹等大型哺乳动物还需要更多的调查验证.在本次调查中, 有蹄类哺乳动物如小麂、赤麂和野猪等在较多位点被监测到, 可能间接佐证该区缺乏大型食肉哺乳动物来调控食草动物种群数量(Gordon et al, 2015 ). ...
微生境尺度下有蹄类动物空间季节利用互作关系研究
1
2018
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
微生境尺度下有蹄类动物空间季节利用互作关系研究
1
2018
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
1
2017
... 整理红外相机所拍摄到的哺乳动物和鸟类照片并进行物种鉴定.哺乳动物的分类体系参考《中国哺乳动物多样性(第2版)》(蒋志刚等, 2017 ), 鸟类的分类体系参考《中国鸟类分类与分布名录(第三版)》(郑光美, 2017 ).夜晚拍到的鼠科动物由于个体较小, 不易辨识, 在计算相对多度指数(relative abundance index, RAI)时统一归为鼠类.定义同一只(群)动物在同一相机位点间隔30 min内连续拍摄的照片作为1张独立有效照片.根据独立有效照片计算相对多度指数(李勤等, 2013 ; 施小刚等, 2017 ): ...
福建哺乳动物种类名录
1
1997
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...
福建哺乳动物种类名录
1
1997
... 本次在戴云山区拍摄到3种鹿科动物: 小麂、赤麂和毛冠鹿(Elaphodus cephalophus ), 这与《福建哺乳动物种类名录》(周冬良, 1997 )和戴云山保护区以往的记录(林鹏, 2003 )相一致.同时本次调查发现3种鹿科动物相对多度指数大小差异较大, 依次为小麂 > 赤麂 > 毛冠鹿, 在福建省其他地区还未有三种动物多度指数大小差异的相关报道, 是否别的地区也呈现类似规律还有待进一步研究.但是从全国红外相机哺乳动物调查结果来看, 三者同时出现在同一地区的情况较少, 仅见广东南岭国家级自然保护区的报道(蔡玉生等, 2016 ), 且毛冠鹿数量稀少.毛冠鹿在中国分布范围较为广泛, 最适生存区在亚热带范围内(盛和林和陆厚基, 1982 ), 但最北可以到宁夏六盘山(高惠等, 2017 ); 而赤麂主要分布于印度、东南亚和我国热带及南亚热带地区(单祥年等, 2004 ); 小麂主要分布在我国秦岭以南、云南哀牢山以北地区, 属于中亚热带至北亚热带范围(鲁庆彬等, 2007 ).同域内鹿科动物稳定共存可能受到栖息地环境、食性、生理、活动时间和体型以及种间关系等因素的影响(杨帆等, 2018 ).戴云山位于中亚热带和南亚热带的交界线, 植物多样性丰富、海拔高度差较大, 为3种动物同域共存提供了基础.赤麂和小麂的栖息地生境选择具有一定的重叠性, 一般选择在低丘乔木密度适中、灌丛密度大和郁闭度适中的生境(白德凤等, 2018 ).两者在本区空间分布上的大量重叠表明它们可能在空间和时间尺度上对食性、生境利用的细微区别.尽管毛冠鹿的食性和小麂甚为相似(盛和林和陆厚基, 1982 ), 但毛冠鹿主要栖息于较高的丘陵, 在空间上与赤麂和小麂重叠性较小(Chen & Hu, 2012 ), 戴云山较大的海拔差能够为毛冠鹿提供较为广泛的活动空间和食物来源, 降低与赤麂和小麂在食物和空间上的竞争.但是受食物资源、温度和环境等影响, 毛冠鹿季节性的海拔梯度迁移可能造成与赤麂和小麂生态位的重叠; 而且戴云山低海拔人为活动的影响造成的小麂、赤麂栖息地破坏和片断化有可能使其向更高海拔迁移, 这将加剧3种动物的种间竞争, 造成种群数量的波动.随着全球气候变化和人为活动等多因素的影响, 需要进一步开展小麂、赤麂和毛冠鹿的种群分布、变化和动态迁移等多方面的研究(徐海根, 2018)① (①徐海根 (2018) 2017年全国生物多样性观测报告. 见: 生物多样性绿皮书, 52-53页. 南京环境科学研究所, 南京.). ...



{kind=link}
{kind=link}