植物区系是植物类群(科、属、种)分布式基本一致的再现, 影响其组成的因素不仅包括水、肥、气、热等生态因子, 还包括影响分布区形成的地质事件及所属分布区发展的历史植物的地理因素(吴征镒等, 2003 )。因为海岛生态系统具有独立性、完整性、脆弱性, 形成了特殊的海岛植被类型, 研究海岛的植物区系分化, 是进行海岛植被资源合理利用和保护的基础, 对于目前国家提倡的海岛整治修复尤其是海岛植被保护和修复具重要意义。
早在达尔文时期, 人们以海岛为对象的科学研究就已经开始。岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 )。物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 )。此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 )。种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 )。但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 )。
生物入侵对环境造成了重大破坏, 改变了生态系统的结构和组分, 甚至改变了生物群落的组分和过程, 而且生物入侵造成的生态系统的改变往往难以恢复, 这些改变也是全球生物多样性丧失的主要原因之一(鞠瑞亭等, 2012 )。根据IUCN红色名录, 全球高度受威胁的脊椎动物分布在约465,000个岛屿上, 同时这些岛屿上均存在入侵物种, 它们是岛屿上物种灭绝的主要驱动因素。全球有61%的已知灭绝种和37%的极度濒危物种发生在海岛上(Spatz, 2017 )。岛屿作为孤立的陆地, 生态系统脆弱, 更容易遭受人为影响, 因此需要紧急行动起来保护海岛生物多样性。
近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来。如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性。这些研究多停留在小区域范围内少数海岛植物群落的研究。
厦门岛周边岛屿较多, 以无居民海岛为多, 鲜少人为活动, 海岛植物资源信息较少。根据全国海岛地名普查数据可知, 厦门周边共有53个海岛, 4个为有居民海岛, 其余49个为无居民海岛。其中28个海岛面积小于1,000 m2 (27个岛上无植被), 面积为1,000-5,000 m2 的海岛有9个(其中2个岛上无植被, 5个主要植被是草本及少数灌木, 仅2个岛上有草本和乔木)。面积大于5,000 m2 的海岛12个(1个是军事用岛)。厦门周边岛屿大小和隔离程度不一, 这些海岛的主要植物区系组成如何? 其物种丰富度主要受何种岛屿空间特征参数影响? 外来植物入侵状况如何? 为回答上述问题, 我们于2017年10月中旬选择1个1,000-5,000 m2 的海岛(有草本及乔木)、11个大于5,000 m2 的海岛(军事用岛除外)进行了植被资源调查。通过分析岛屿植物区系组成、种-面积关系以及岛屿间物种相似性, 探讨了厦门近岸12个无居民海岛面积、岛屿距大陆距离等岛屿空间特征参数对物种丰富度的影响。
1 调查区概况和调查方法
1.1 调查区概况
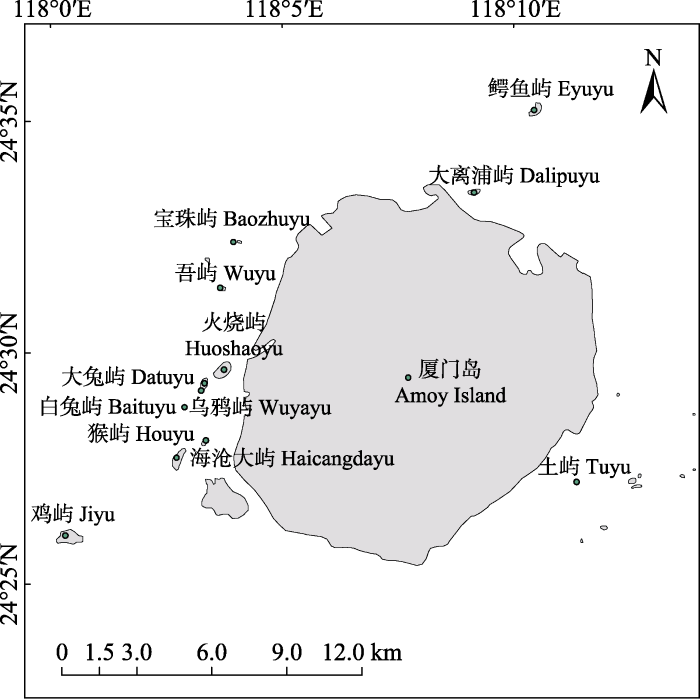
厦门市辖区内海岛南北跨距19.08 km (24°25°59.21''-24°35°16.45'' N), 东西跨距16.55 km (118°0°18.83''-118°11°21.48'' E); 属南亚热带海洋性季风气候, 冬无严寒, 夏无酷暑, 年均温20.7℃, 1月和7月均温分别为12.8℃和28.0℃; 年、日温差都较小; 极端最低温2℃, 极端最高温39.8℃; 月均降水量111.03 mm, 7-9月为台风季节; 降雨集中在5-9月(http://data.cma.cn/data/weatherBk.html )。本次调查的12个无居民海岛大部分为基岩岛, 部分岛屿以变质砂岩为主, 夹粉砂岩、泥岩及火山碎屑岩, 主要集中在厦门本岛和海沧区的近海海域, 均属于近岸岛, 除土屿外, 其余11个均为沿岸岛(图1 )。近年来, 火烧屿、宝珠屿、乌鸦屿、大离浦屿、鳄鱼屿等岛屿经历了部分人工开发, 海沧大屿和鸡屿位于厦门珍稀海洋物种国家级自然保护区核心区内, 人为干扰痕迹较少, 其余岛屿面积较小, 开发利用价值不大, 均保持了较为原生的海岛原貌。
图1 厦门12个调查岛屿区域分布图
Fig. 1 Geographical distribution of the investigated 12 islands at Amoy Island
1.2 调查方法
以整岛踏查为主, 样线和样方调查为辅。详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本。乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个。现场记录植被类型、株高、盖度等信息, 形成解译标志。利用GPS确定样方位置。按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 )。并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分。
1.3 数据处理
1.3.1 岛屿植物物种多样性分析
本研究采用以下指数度量群落物种多样性:
物种丰富度S = 出现在样地的物种数 (1)
Shannon-Wiener多样性指数(Shannon, 1938 )
${H}'=-\sum\limits_{i=1}^{s}{{{P}_{i}}\ln {{P}_{i}}}$(2)
Pielou均匀度指数(田家怡等, 2011 )
$J=-\sum\limits_{i=1}^{s}{({{P}_{i}}\ln {{P}_{i}})/\ln S}$ (3)
式中, P i 为第i种的个体数Ni 占所有个体总数N 的比例, 即P i = Ni /N , i = 1, 2, 3, …, S , S 为样地中物种数, 所有的计算在Excel中完成。
1.3.2 岛屿空间特征参数
根据国家海洋信息中心海岛监视监测系统内的海岛资料(表1 ), 利用岛屿面积(A )、岸线长度(Per )、距大陆距离(I m )、岛屿间距离(I i )、岛屿高程(E )、周长/面积比(PAR )、形状指数(SI )等参数来描述调查岛屿的空间特征, 其中I m 表示岛屿边缘至大陆的最小直线距离, PAR 为岛屿边缘的相对长度, 表征岛屿的边界效应(Wu, 2004 )。SI 反映岛屿形状的复杂程度, 由公式SI = Per /[2×(π×A )×0.5]计算。
1.3.3 种-面积关系的拟合
选择线性函数、二次函数、三次函数、复合函数y = abx 、幂函数y = axb 、S型曲线y = ea+b/x y = ea+bx y = a ebx y = a + b ln(x ), Logistic函数y = 1/(1/u + abx )和逆函数y = a + b /x 等11种方程对物种丰富度和面积的关系进行曲线拟合, 选取拟合较好的曲线进行分析, 所有统计分析均在SPSS 20统计软件中完成。
1.3.4 Jaccard相似性系数
采用Jaccard相似性系数C j = j /(a + b - j )计算不同岛屿间的相似性(Whittaker, 1972 ), 其中j 为不同岛屿间共有的物种数, a 和b 分别为不同岛屿自身包含的物种数目。
采用SPSS 20对12个无居民岛屿上述空间特征参数进行相关性分析, 同时与岛屿物种丰富度进行相关性和偏相关分析。此外, 构建了适合草本和木本植物丰富度的种-面积关系模型。数据处理使用Excel 2013和SPSS 20完成。
2 结果
2.1 厦门12个无居民海岛植被概况
12个无居民海岛共记录维管束植物360种, 隶属于102科286属, 其中蕨类植物8科8属13种, 裸子植物6科6属7种, 被子植物88科272属340种(附录1); 乔木78种, 小乔木或灌木109种, 藤本23种, 草本150种。岛屿间物种丰富度差异较大, 最多的岛屿有178种, 最少的仅19种, 物种数40-60的岛屿数量占50.0%, 物种数90-120的岛屿数量占33.3% (表1 )。
从属种系数(一个地区植物属总数与种总数的比值, 田家怡等, 2011 )来看, 厦门12个无居民海岛的属种系数最大为100%, 分别为吾屿和宝珠屿, 反映出该二岛全为单属种, 生境条件较为恶劣, 也可能由于岛屿面积较小, 一定程度上存在的小岛屿效应导致该二岛全为单属种。属种系数较小的是海沧大屿, 属种系数为88.0%。总体来看, 本次调查的12个海岛属种系数均较高, 平均为94.17%, 表明该地区的单种属或少种属较多, 也反映出海岛生境条件较为恶劣, 相较大陆种群, 适宜生长的植物种类有限。厦门周边海域无居民海岛平均Shannon-Wiener多样性指数为0.92, Pielou均匀度指数为0.52 (表2 )。其中鸡屿的维管束植物多样性水平最高, 其次为海沧大屿、白兔屿, 较低的为鳄鱼屿、猴屿、土屿、吾屿和乌鸦屿。
2.2 种子植物区系特点
12个无居民海岛共记录种子植物278属347种, 可划分为13个分布区类型及其变型(表3 ), 主要是泛热带分布及其变型, 其次为热带亚洲和热带美洲间断分布和世界分布型, 不存在温带亚洲分布和中亚分布及其变型。热带性质的属占73.02%, 温带性质的属占17.27%, 以热带、亚热带类型占主导地位, 符合其亚热带地理分布特点。
2.3 岛屿物种丰富度和种-面积拟合分析
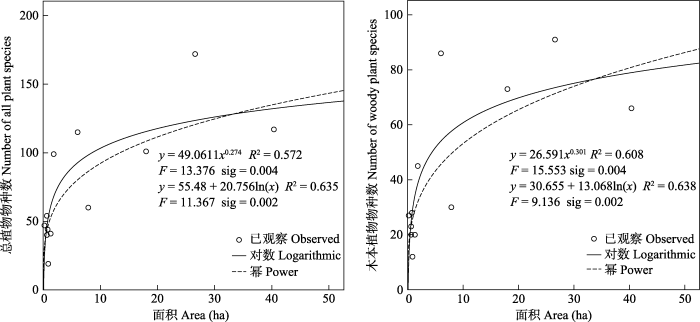
从本文选定的11种函数的拟合结果来看, 幂函数和对数函数对总植物物种数和木本植物种数与岛屿面积关系的拟合效果较好(图2 ), 而草本物种数与岛屿面积的关系拟合效果不佳, 未列出。总体上, 随着岛屿面积的增大, 物种数目增多, 但面积增大到一定程度后, 植物物种数达到饱和。
图2 物种数和岛屿面积的关系
Fig. 2 Relationship between species number and island area
2.4 岛屿空间特征对物种丰富度的影响
由表4 可以看出, 岛屿面积与岸线长度、高程均呈显著正相关(P < 0.01), 与形状指数显著相关(P < 0.05), 与周长/面积比呈显著负相关(P < 0.01)。距大陆距离与周长/面积比显著正相关(P < 0.05), 岸线长度和高程、形状指数也呈显著正相关(P < 0.01),岸线长度、高程、形状指数均和周长/面积比呈现显著负相关(P < 0.01)。高程和形状指数呈显著正相关(P < 0.05)。
岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 )。随着面积、岸线长度和高程的增加, 形状越复杂的海岛拥有的植物丰富度越高; 周长/面积比与物种丰富度显著负相关, 即边缘效应越显著, 丰富度越低。
2.5 不同岛屿间物种组成相似性系数
从表6 可知, 大兔屿和白兔屿的物种组成相似性最高, 相似性系数为0.78, 其次为鳄鱼屿和白兔屿, 相似性系数为0.65。总体上, 相似性系数大于0.4的岛屿有6组。物种相似性最低的是宝珠屿和鳄鱼屿、火烧屿和吾屿。吾屿由于面积较小, 物种数少, 与大多数岛屿的植被相似性均较低。
物种相似性系数C j 与岛屿空间特征的相关分析显示, δA = 0.058, δH = 0.243* , δI = -0.183, δPer = 0.101, δE = 0.056, δPAR = -0.264* , δSI = 0.093。其中, δA 、δH 、δI 、δPer 、δE 、δPAR 、δSI 分别表示两个岛屿之间的面积比、生境类型数量比、距大陆距离比(主要根据2015年全国海岛地名普查数据)、岸线长度比、最大高程比、周长/面积比的比值、形状指数比。结果表明, C j 与生境类型数目显著正相关(P < 0.05), 与周长面积比(PAR)显著负相关(P < 0.05), 与其他参数无显著相关性。
2.6 外来植物分析
厦门近岸海域海岛合计有入侵植物49种, 占总物种数的13.6%。根据中国外来入侵植物信息系统(http://ias.iplant.cn/protlist)公布的中国入侵植物名录及入侵等级(闫小玲等, 2014 ), 12个无居民海岛均有恶性入侵植物, 共10种, 分别是土荆芥(Chenopodium ambrosioides )、银胶菊(Parthenium hysterophorus )、钻叶紫菀(Aster subulatus )、光荚含羞草(Mimosa sepiaria )、互花米草(Spartina alterniflora )、藿香蓟(Ageratum conyzoides )、蒺藜草(Cenchrus echinatus )、马缨丹(Lantana camara )、白花鬼针草(Bidens pilosa var. radiata )、假臭草(Eupatorium catarium )。其中鸡屿和大兔屿恶性入侵植物最多, 分别多达7种和6种, 占比分别为5.98%和5.22%。其次是严重入侵物种, 12个海岛共计14种, 除吾屿和宝珠屿以外, 其他10个海岛上或多或少存在严重入侵物种。此外, 12个海岛局部入侵类5种、一般入侵类5种、有待观察类15种。总体来看, 厦门周边的海岛维管束植物以乡土植物为主。
3 讨论
3.1 海岛植物区系组成分析
厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点。张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘。泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 )。福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源。
厦门近岸海域12个海岛与厦门本岛(厦门同安区内田村)相比, 相同点为热带性质的属占大多数, 不同之处在于12个海岛热带至亚热带性质属的比例更高, 占73.02%, 而内田村的为64.95%。该结果与林文俊(2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)对比闽台植物区系所得结论相一致, 台湾岛的热带属性质强于邻近大陆福建。福建热带分布属占总属数的60.34%, 温带分布属占总属数的24.10%, 表现出明显的热带性质, 厦门近岸海域12个海岛热带性质的属占总属数的73.02%, 温带性质的属占17.27%, 热带属性质高于福建整体水平(林文俊, 2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)。同台湾植物区系相比, 厦门周边海域12个海岛热带属性质也高于台湾(热带分布属占总属数的66.21%), 温带分布属性质低于台湾(温带分布属占22.47%)(应俊生等, 2002 ; 林文俊, 2013① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.))。这与海岛的气候环境和物理性质有关, 厦门近岸海域海岛相对而言环境更为恶劣, 海岛面积较小, 受到大风、阳光直射的影响较大, 温带植物分布较少, 热带至亚热带性质的植物更适宜生长。
同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 )。根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 )。第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 )。厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 )。本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属。其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 )。本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等。这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 )。位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似。本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛。面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型。总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息。结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2)。
3.2 影响岛屿维管束植物物种丰富度和岛屿相似性的空间特征参数
从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致。此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 )。一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 )。而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大。同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高。由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著。周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长。岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散。另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著。
物种相似性系数C j 与生境类型数目显著正相关, 与周长/面积比(PAR )显著负相关, 与其他参数无显著相关性。厦门周边12个无居民海岛中, 物种组成相似性较高的有: 大兔屿和白兔屿, 鳄鱼屿和白兔屿, 大兔屿和鳄鱼屿, 鸡屿和大兔屿, 猴屿和鳄鱼屿, 白兔屿和海沧大屿。如上文所提, 生境异质性是决定物种丰富度的主要因素。周长/面积比越大, 即边缘越大, 相同面积内可供植物生长的实际面积减小, 因此物种丰富度越低, 相应的物种相似性也较低。总体上, 不同岛屿间维管束植物物种相似性受生境多样性和岛屿边缘效应影响, 该结果与王金旺等(2017) 的研究一致。此外, 我们发现吾屿与大多数岛屿的物种相似性均较低, 可能是由于其面积较小, 导致维管束植物种类数少, 一定程度上反映吾屿可能出现小岛屿效应。
3.3 外来入侵植物分析
总体来看, 厦门周边的海岛维管束植物以乡土植物为主, 但岛上的入侵物种不容忽视。例如猴屿上的马缨丹纯群丛: 由于早年建设高压铁塔时, 岛上植被遭到严重破坏, 建设单位引入马缨丹进行植被修复, 现今几乎占据了整个灌木层。马缨丹具有强烈的化感作用, 在岛上形成单优群落, 压制或排挤本地种, 严重影响了海岛物种多样性。岛屿作为孤立的大陆, 经常维持着种群规模小、繁殖力低、防御力弱的简单生态系统(Spatz, 2017 ), 岛屿物种更容易遭受各种外来因素的影响。入侵物种还可通过杂交和基因渗入的方式入侵到土著植物的基因库中, 造成土著植物基因型的丧失, 并有可能导致土著植物的灭绝, 这种杂交很有可能使产生的后代比亲本具有更强的入侵性(万宣伍等, 2015 ), 从而导致更为严重的生态后果, 需要引起重视。
(责任编委: 沈泽昊 责任编辑: 黄祥忠、周玉荣)
致谢:
感谢厦门大学杨盛昌课题组的黄晓敏、彭建、黄含玥、杨慧等师弟、师妹帮忙野外调查以及后期数据的整理。
附录 Supplementary Material
附录1 厦门近岸12个无居民岛屿植物名录
Appendix 1 Plant list of 12 uninhabited islands in the nearshore Xiamen
http://www.biodiversity-science.net/fileup/PDF/2018124-1.pdf
附录2 2017年度福建省厦门11个海岛植被类型图
Appendix 2 Vegetation maps of 11 islands in the nearshore Xiamen, Fujian Province in 2017
http://www.biodiversity-science.net/fileup/PDF/2018124-2.pdf
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Chen ZS Xia DX Wang JW Zhou PX Guo DX Jiang F Wang DZ Bao YE Dou ZX Lu YX Wang WH Song CB Chen WL Xie QC Feng YJ Liu WK Chen RX Lin YX Guo F Li JF Chen F Xiong SL Wang JL Qiu ZG 1993 ) Survey of China Bays, Vol. 8 , pp. 165 -191 . China Ocean Press , Beijing . (in Chinese)
[本文引用: 1]
[陈则实 , 夏东兴 , 王建文 , 周培兴 , 郭德喜 , 姜富 , 王德正 , 鲍永恩 , 窦振兴 . 路应贤 , 王文海 , 宋昌斌 , 陈伟伦 , 谢钦春 , 冯应俊 , 刘维坤 , 陈瑞祥 , 林应信 , 郭郛 , 李家芳 , 陈峰 , 熊仕林 , 王加林 , 邱志高 (1993 ) 中国海湾志第八册 , 165 -191 页. 海洋出版社 , 北京 .]
[本文引用: 1]
[2]
Fang JY Wang XP Shen ZH Tang ZY He JS Yu D Jiang Y Wang ZH Zheng CY Zhu JL Guo ZD 2009 ) Methods and protocols for plant community inventory
. Biodiversity Science , 17 , 533 -548 . (in Chinese with English abstract)
[本文引用: 1]
[方精云 , 王襄平 , 沈泽昊 , 唐志尧 , 贺金生 , 于丹 , 江源 , 王志恒 , 郑成洋 , 朱江玲 , 郭兆迪 (2009 ) 植物群落清查的主要内容、方法和技术规范
. 生物多样性 , 17 , 533 -548 .]
[本文引用: 1]
[3]
Hoffmeister TS Lem V Biere A Holsinger K Fisher J 2005 ) Ecological and evolutionary consequences of biological invasion and habitat fragmentation
. Ecosystems , 8 , 657 -667 .
[4]
Invasive Alien Species of China (IASC) (2016 ) Authority Platform for China’s Invasive Alien Species of China
. (in Chinese)
URL
[中国外来入侵物种信息系统 (2016 ) 权威的中国外来入侵物种信息系统
.]. (accessed on 2018-05-31
URL
[5]
Ju RT Li H Shi ZR Li B 2012 ) Progress of biological invasions research in China over the last decade
. Biodiversity Science , 20 , 581 -611 . (in Chinese with English abstract)
[本文引用: 1]
[鞠瑞亭 , 李慧 , 石正人 , 李博 (2012 ) 近十年中国生物入侵研究进展
. 生物多样性 , 20 , 581 -611 .]
[本文引用: 1]
[6]
Kerr JT Packer L 1997 ) Habitat heterogeneity as a determinant of mammal species richness in high-energy regions
. Nature , 385 , 252 -254 .
[本文引用: 1]
[7]
Lomolino MV 2000 ) Ecology’s most general, yet protean pattern: The species-area relationship
. Journal of Biogeography , 27 , 17 -26 .
[本文引用: 1]
[8]
Morrison LW 2011 ) Why do some small islands lack vegetation? Evidence from long-term introduction experiments
. Ecography , 34 , 384 -391 .
[本文引用: 1]
[9]
Pinheiro HT Bernardi G Simon T Joyeux JC Mecieira RM Gasparini JL Rocha C Rocha LA 2017 ) Island biogeography of marine organisms
. Nature , 549 , 82 -85 .
[本文引用: 1]
[10]
Qian H Ricklefs RE White PS 2005 ) Beta diversity of angiosperms in temperate floras of eastern Asia and eastern North America
. Ecology Letters , 8 , 15 -22 .
[本文引用: 1]
[11]
Shannon CE 1938 ) A mathematical theory of communication
. The Bell System Technical Journal , 196 , 519 -520 .
[本文引用: 1]
[12]
Spatz DR Zilliacus KM Holmes ND Butchart SHM Genovesi P Ceballos G Tershy BR Croll DA 2017 ) Globally threatened vertebrates on islands with invasive species
. Science Advances , 3 , e1603080 .
[本文引用: 2]
[13]
Sun YM Tang KX Chen HY Chen YZ Huang HP Cai LC 2017 ) Preliminary study on the characteristics of species diversity and the soil in non-resident island vegetation restoration
. Journal of Applied Oceanography , 36 , 1 -5 . (in Chinese with English abstract)
[本文引用: 1]
[孙元敏 , 汤坤贤 , 陈慧英 , 陈玉珍 , 黄海萍 , 蔡鹭春 (2017 ) 无居民海岛植被恢复过程中物种多样性及土壤特征初步研究
. 应用海洋学学报 , 36 , 1 -5 .]
[本文引用: 1]
[14]
Sun Q Lu JB Zhang FF Xu GF 2009 ) Plant species diversity in relation to island size
. Acta Ecologica Sinica , 29 , 2195 -2202 . (in Chinese with English abstract)
[本文引用: 1]
[孙雀 , 卢剑波 , 张凤凤 , 徐高福 (2009 ) 植物物种多样性与岛屿面积的关系
. 生态学报 , 29 , 2195 -2202 .]
[本文引用: 1]
[15]
Su XF Yuan JF Hu G Xu GF Yu MJ 2014 ) Edge effect of the plant community structure on land-bridge islands in the Thousand Island Lake
. Chinese Journal of Applied Ecology , 25 , 77 -84 . (in Chinese with English abstract)
[本文引用: 1]
[苏晓飞 , 袁金凤 , 胡广 , 徐高福 , 于明坚 (2014 ) 千岛湖陆桥岛屿植物群落结构的边缘效应
. 应用生态学报 , 25 , 77 -84 .]
[本文引用: 1]
[16]
Tan ZQ Zeng WB Zhang RT 1996 ) A study on the flora of Neitian in Tong’an, Xiamen
. Journal of Xiamen University (Natural Science) , 35 , 428 -433 . (in Chinese with English abstract)
[本文引用: 1]
[谭忠奇 , 曾文彬 , 张娆挺 (1996 ) 厦门同安县内田植物区系成分分析,
厦门大学学报(自然科学版) , 35 , 428 -433 .]
[本文引用: 1]
[17]
Tian JY Xia JB Sun JK 2011 ) Ecological Protection and Restoration of Shell Island in the Yellow Sea Delta . Chemical Industry Press , Beijing . (in Chinese)
[本文引用: 2]
[田家怡 , 夏江宝 , 孙景宽 (2011 ) 黄海三角洲贝壳堤岛生态保护与修复 . 化学工业出版社 , 北京 .]
[本文引用: 2]
[18]
Wan XW Liu YH Luo LM Feng CH Wang S Ma L 2015 ) Inference on the invasion history of invasive alien species based on molecular methods
. Acta Ecologica Sinica , 35 , 1296 -1309 . (in Chinese with English abstract)
[本文引用: 1]
[万宣伍 , 刘映红 , 罗林明 , 封传红 , 王胜 , 马利 (2015 ) 基于分子生物学方法的外来入侵物种入侵历史重构
. 生态学报 , 35 , 1296 -1309 .]
[本文引用: 1]
[19]
Wang JW Wei X Chen QX Li XW Yang S 2017 ) Factors affecting species richness and beta diversity of vascular plants on small islands in the Wenzhou region of eastern China
. Acta Ecologica Sinica , 37 , 523 -540 . (in Chinese with English abstract)
[本文引用: 2]
[王金旺 , 魏馨 , 陈秋夏 , 李效文 , 杨升 (2017 ) 温州沿海小型海岛植物丰富度和β多样性及其影响因子
. 生态学报 , 37 , 523 -540 .]
[本文引用: 2]
[20]
Wang RS Shi ZQ Huang CQ Yu ZH Zheng WW Chi JW 2015 ) Research report on the development of islands geological environment in Fujian Province
. Strait Science , (1 ), 10 -16 . (in Chinese)
[本文引用: 1]
[王仁山 , 施志强 , 黄昌旗 , 余芝华 , 郑慰雯 , 池际武 (2015 ) 福建省岛屿地质环境学科发展研究报告
. 海峡科学 , (1 ), 10 -16 .]
[本文引用: 1]
[21]
Wu JG 2004 ) Effects of changing scale on landscape pattern analysis: Scaling relations
. Landscape Ecology , 19 , 125 -138 .
[本文引用: 1]
[22]
Whittaker RH 1972 ) Evolution and measurement of species diversity
. Taxon , 21 , 213 -251 .
[本文引用: 1]
[23]
Wu ZY 1991 ) The areal-types of Chinese genera of seed plants
. Acta Botanica Yunnanica , 13 (Suppl. 4 ), 1 -139 . (in Chinese with English abstract)
[本文引用: 1]
[吴征镒 (1991 ) 中国种子植物属的分布区类型
. 云南植物研究 , 13 (Suppl. 4 ), 1 -139 .]
[本文引用: 1]
[24]
Wu ZY Zhou ZK Li DZ Peng H Sun H 2003 ) The areal- types of the world families of seed plants
. Acta Botanica Yunnanica , 25 , 245 -257 . (in Chinese with English abstract)
[本文引用: 2]
[吴征镒 , 周浙昆 , 李德铢 , 彭华 , 孙航 (2003 ) 世界种子植物科的分布区类型系统
. 云南植物研究 , 25 , 245 -257 .]
[本文引用: 2]
[25]
Xiao L Yang SC Hou R Liu CA Lu WZ Chen PF 2018 ) Ecological niche characteristics of vascular plant species in the Natural Chenier Island Reserve of Wudi, Shandong Province
. Journal of Xiamen University (Natural Science) , 57 , 79 -84 . (in Chinese with English abstract)
[本文引用: 1]
[肖兰 , 杨盛昌 , 侯蕊 , 刘长安 , 卢伟志 , 陈鹏飞 (2018 ) 山东无棣贝壳堤保护区维管束植物的生态位研究
. 厦门大学学报(自然科学版) , 57 , 79 -84 .]
[本文引用: 1]
[26]
Xu HS Ma DW 2016 ) The seed plants areal types in Gulangyu Island of Fujian
. Journal of Sichuan Normal University (Natural Science) , 39 , 588 -592 . (in Chinese with English abstract)
[本文引用: 2]
[徐河山 , 马丹炜 (2016 ) 鼓浪屿岛种子植物分布区类型初步研究
. 四川师范大学学报(自然科学版) , 39 , 588 -592 .]
[本文引用: 2]
[27]
Yan XL Liu QR Shou HY Zeng XF Zhang Y Chen L Liu Y Ma HY Qi SY Ma JS 2014 ) The categorization and analysis on the geographic distribution patterns of Chinese alien invasive plants
. Biodiversity Science , 22 , 667 -676 . (in Chinese with English abstract)
[本文引用: 1]
[闫小玲 , 刘全儒 , 寿海洋 , 曾宪锋 , 张勇 , 陈丽 , 刘演 , 马海英 , 齐淑艳 , 马金双 (2014 ) 中国外来入侵植物的等级划分与地理分布格局分析
. 生物多样性 , 22 , 667 -676 .]
[本文引用: 1]
[28]
Ying JS Xu GS 2002 ) An analysis of the flora of seed plants of Taiwan, China: Its nature, characteristics, and relations with the flora of the mainland
. Acta Phytotaxonomica Sinica , 40 , 1 -51 . (in Chinese with English abstract)
[本文引用: 2]
[应俊生 , 徐国士 (2002 ) 中国台湾种子植物区系的性质、特点及其与大陆植物区系的关系
. 植物分类学报 , 40 , 1 -51 .]
[本文引用: 2]
[29]
Zeng WB 1981 ) The origin of Fujian flora, China
. Journal of Xiamen University (Natural Science) , 20 , 487 -495 . (in Chinese with English abstract)
[本文引用: 3]
[曾文彬 (1981 ) 福建植物区系的由来
. 厦门大学学报(自然科学版) , 20 , 487 -495 .]
[本文引用: 3]
[30]
Zeng WB 1983 ) The flora and phytogeographical subdivision of Fujian
. Journal of Xiamen University (Natural Science) , 22 , 217 -226 . (in Chinese with English abstract)
[本文引用: 1]
[曾文彬 (1983 ) 福建植物区系与植物地理区域
. 厦门大学学报(自然科学版) , 22 , 217 -226 .]
[本文引用: 1]
[31]
Zhang LT Wang WQ Jiang DG 2017 ) Analysis of seed flora and its remediation strategies on Xiamen Datu Island
. Ocean Development and Management , 34 (3 ), 81 -87 . (in Chinese with English abstract)
[本文引用: 1]
[张琳婷 , 王文卿 , 姜德刚 (2017 ) 厦门大兔屿种子植物区系分析及其植被修复对策研究
. 海洋开发与管理 , 34 (3 ), 81 -87 .]
[本文引用: 1]
[32]
Zhang YW 2012 ) Island Development and Protection in China: Geographical Perspective. China Ocean Press , Beijing . (in Chinese)
[本文引用: 1]
[张耀文 (2012 ) 中国海岛开发与保护——地理学视角 . 海洋出版社 , 北京 .]
[本文引用: 1]
[33]
Zheng JM Fang X Zhu XP Zhu DD Deng CY 2016 ) Wild plant germplasm and biodiversity in Dayu Island, Pingtan
. Journal of Anhui Agricultural University , 43 , 640 -645 . (in Chinese with English abstract)
[本文引用: 1]
[郑俊鸣 , 方笑 , 朱雪平 , 朱丹丹 , 邓传远 (2016 ) 平潭大屿岛植物资源及其多样性研究
. 安徽农业大学学报 , 43 , 640 -645 .]
[本文引用: 1]
[34]
Zheng JM Fang X Zhu XP Zhu DD Deng CY Huang LJ 2017 ) Vegetation characteristics and plant diversity of Waimalangshan Island, Zhoushan
. Guihaia , 37 , 271 -279 . (in Chinese with English abstract)
[本文引用: 1]
[郑俊鸣 , 方笑 , 朱雪平 , 朱丹丹 , 邓传远 , 黄柳菁 (2017 ) 外马廊山岛植被特性与植物多样性
. 广西植物 , 37 , 271 -279 .]
[本文引用: 1]
[35]
Zheng LT Su T Liu XY Yin F Guo C Tuo B Yan ER 2018 ) Species, functional, structural diversity of typical plant communities and their responses to environmental factors in Miao Archipelago, China
. Chinese Journal of Applied Ecology , 29 , 343 -351 . (in Chinese with English abstract)
[本文引用: 1]
[郑丽婷 , 苏田 , 刘翔宇 , 尹芳 , 郭超 , 妥彬 , 阎恩荣 (2018 ) 庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应
. 应用生态学报 , 29 , 343 -351 .]
[本文引用: 1]
1
1993
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
1
1993
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
Ecological and evolutionary consequences of biological invasion and habitat fragmentation
2005
近十年中国生物入侵研究进展
1
2012
... 生物入侵对环境造成了重大破坏, 改变了生态系统的结构和组分, 甚至改变了生物群落的组分和过程, 而且生物入侵造成的生态系统的改变往往难以恢复, 这些改变也是全球生物多样性丧失的主要原因之一(鞠瑞亭等, 2012 ).根据IUCN红色名录, 全球高度受威胁的脊椎动物分布在约465,000个岛屿上, 同时这些岛屿上均存在入侵物种, 它们是岛屿上物种灭绝的主要驱动因素.全球有61%的已知灭绝种和37%的极度濒危物种发生在海岛上(Spatz, 2017 ).岛屿作为孤立的陆地, 生态系统脆弱, 更容易遭受人为影响, 因此需要紧急行动起来保护海岛生物多样性. ...
近十年中国生物入侵研究进展
1
2012
... 生物入侵对环境造成了重大破坏, 改变了生态系统的结构和组分, 甚至改变了生物群落的组分和过程, 而且生物入侵造成的生态系统的改变往往难以恢复, 这些改变也是全球生物多样性丧失的主要原因之一(鞠瑞亭等, 2012 ).根据IUCN红色名录, 全球高度受威胁的脊椎动物分布在约465,000个岛屿上, 同时这些岛屿上均存在入侵物种, 它们是岛屿上物种灭绝的主要驱动因素.全球有61%的已知灭绝种和37%的极度濒危物种发生在海岛上(Spatz, 2017 ).岛屿作为孤立的陆地, 生态系统脆弱, 更容易遭受人为影响, 因此需要紧急行动起来保护海岛生物多样性. ...
Habitat heterogeneity as a determinant of mammal species richness in high-energy regions
1
1997
... 从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致.此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 ).一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 ).而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大.同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高.由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著.周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长.岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散.另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著. ...
Ecology’s most general, yet protean pattern: The species-area relationship
1
2000
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
Why do some small islands lack vegetation? Evidence from long-term introduction experiments
1
2011
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
Island biogeography of marine organisms
1
2017
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
Beta diversity of angiosperms in temperate floras of eastern Asia and eastern North America
1
2005
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
A mathematical theory of communication
1
1938
... Shannon-Wiener多样性指数(Shannon, 1938 ) ...
Globally threatened vertebrates on islands with invasive species
2
2017
... 生物入侵对环境造成了重大破坏, 改变了生态系统的结构和组分, 甚至改变了生物群落的组分和过程, 而且生物入侵造成的生态系统的改变往往难以恢复, 这些改变也是全球生物多样性丧失的主要原因之一(鞠瑞亭等, 2012 ).根据IUCN红色名录, 全球高度受威胁的脊椎动物分布在约465,000个岛屿上, 同时这些岛屿上均存在入侵物种, 它们是岛屿上物种灭绝的主要驱动因素.全球有61%的已知灭绝种和37%的极度濒危物种发生在海岛上(Spatz, 2017 ).岛屿作为孤立的陆地, 生态系统脆弱, 更容易遭受人为影响, 因此需要紧急行动起来保护海岛生物多样性. ...
... 总体来看, 厦门周边的海岛维管束植物以乡土植物为主, 但岛上的入侵物种不容忽视.例如猴屿上的马缨丹纯群丛: 由于早年建设高压铁塔时, 岛上植被遭到严重破坏, 建设单位引入马缨丹进行植被修复, 现今几乎占据了整个灌木层.马缨丹具有强烈的化感作用, 在岛上形成单优群落, 压制或排挤本地种, 严重影响了海岛物种多样性.岛屿作为孤立的大陆, 经常维持着种群规模小、繁殖力低、防御力弱的简单生态系统(Spatz, 2017 ), 岛屿物种更容易遭受各种外来因素的影响.入侵物种还可通过杂交和基因渗入的方式入侵到土著植物的基因库中, 造成土著植物基因型的丧失, 并有可能导致土著植物的灭绝, 这种杂交很有可能使产生的后代比亲本具有更强的入侵性(万宣伍等, 2015 ), 从而导致更为严重的生态后果, 需要引起重视. ...
无居民海岛植被恢复过程中物种多样性及土壤特征初步研究
1
2017
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
无居民海岛植被恢复过程中物种多样性及土壤特征初步研究
1
2017
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
植物物种多样性与岛屿面积的关系
1
2009
... 从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致.此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 ).一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 ).而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大.同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高.由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著.周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长.岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散.另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著. ...
植物物种多样性与岛屿面积的关系
1
2009
... 从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致.此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 ).一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 ).而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大.同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高.由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著.周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长.岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散.另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著. ...
千岛湖陆桥岛屿植物群落结构的边缘效应
1
2014
... 从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致.此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 ).一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 ).而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大.同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高.由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著.周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长.岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散.另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著. ...
千岛湖陆桥岛屿植物群落结构的边缘效应
1
2014
... 从种-面积关系拟合效果来看, 12个无居民海岛的物种丰富度与面积的关系遵循对数函数和幂函数的规律, 在一定范围内, 物种丰富度随着面积的增大而增加, 增大到一定范围后物种丰富度保持不变, 该结果符合岛屿地理学理论, 与孙雀等(2009) 的研究结果一致.此次调查的12个岛屿物种丰富度不但受面积影响, 还受其他岛屿空间特征影响, 岛屿面积、岸线长度、高程、周长/面积比、形状指数显著影响维管束植物丰富度(表5 ).一般地, 随着面积的增大, 生境类型也越多, 尤其在低纬度地区(Kerr et al, 1997 ).而物种多样性依赖于生境异质性, 生境多样性越高, 物种丰富度越大.同面积相类似, 随岸线长度和高程的增加, 海岛拥有的物种丰富度越高.由于面积和岸线长度、高程存在显著相关性, 因此岸线的长度和高程对岛屿物种丰富度的影响一定程度是受面积的影响; 形状越复杂, 岛屿物种丰富度越高, 但不显著.周长/面积比即岛屿边缘效应与物种丰富度显著负相关, 不同于传统的岛屿边缘效应的研究, 处于边缘的物种由于具有相邻生境的特点其物种丰富度高于内部生态系统(袁金凤, 2011① (① 袁金凤 (2011) 边缘效应对千岛湖陆桥岛屿植物群落结构的影响. 硕士学位论文, 浙江大学, 杭州); 苏晓飞等, 2014 ), 主要是由于海岛岸线周围都是礁石亦或滩涂, 并不适宜海岛植物生长.岛屿植物丰富度与离岸距离不存在显著相关性, 一方面可能是由于这些岛屿(除土屿外)均为沿岸岛, 物种间的扩散受到的阻碍较小, 再加上厦门近岸海域的无居民海岛是鸟类栖息的重要场所, 通常情况下, 近距离的海岛为鸟类传播扩散物种提供了良好的廊道, 有利于物种间的扩散.另一方面, 上文的推测表明这些岛原先均是同一大陆区域, 海岛植物类群有着共同的生长史, 因此隔离程度对物种丰富度的影响不显著. ...
厦门同安县内田植物区系成分分析,
1
1996
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
厦门同安县内田植物区系成分分析,
1
1996
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
2
2011
... Pielou均匀度指数(田家怡等, 2011 ) ...
... 从属种系数(一个地区植物属总数与种总数的比值, 田家怡等, 2011 )来看, 厦门12个无居民海岛的属种系数最大为100%, 分别为吾屿和宝珠屿, 反映出该二岛全为单属种, 生境条件较为恶劣, 也可能由于岛屿面积较小, 一定程度上存在的小岛屿效应导致该二岛全为单属种.属种系数较小的是海沧大屿, 属种系数为88.0%.总体来看, 本次调查的12个海岛属种系数均较高, 平均为94.17%, 表明该地区的单种属或少种属较多, 也反映出海岛生境条件较为恶劣, 相较大陆种群, 适宜生长的植物种类有限.厦门周边海域无居民海岛平均Shannon-Wiener多样性指数为0.92, Pielou均匀度指数为0.52 (表2 ).其中鸡屿的维管束植物多样性水平最高, 其次为海沧大屿、白兔屿, 较低的为鳄鱼屿、猴屿、土屿、吾屿和乌鸦屿. ...
2
2011
... Pielou均匀度指数(田家怡等, 2011 ) ...
... 从属种系数(一个地区植物属总数与种总数的比值, 田家怡等, 2011 )来看, 厦门12个无居民海岛的属种系数最大为100%, 分别为吾屿和宝珠屿, 反映出该二岛全为单属种, 生境条件较为恶劣, 也可能由于岛屿面积较小, 一定程度上存在的小岛屿效应导致该二岛全为单属种.属种系数较小的是海沧大屿, 属种系数为88.0%.总体来看, 本次调查的12个海岛属种系数均较高, 平均为94.17%, 表明该地区的单种属或少种属较多, 也反映出海岛生境条件较为恶劣, 相较大陆种群, 适宜生长的植物种类有限.厦门周边海域无居民海岛平均Shannon-Wiener多样性指数为0.92, Pielou均匀度指数为0.52 (表2 ).其中鸡屿的维管束植物多样性水平最高, 其次为海沧大屿、白兔屿, 较低的为鳄鱼屿、猴屿、土屿、吾屿和乌鸦屿. ...
基于分子生物学方法的外来入侵物种入侵历史重构
1
2015
... 总体来看, 厦门周边的海岛维管束植物以乡土植物为主, 但岛上的入侵物种不容忽视.例如猴屿上的马缨丹纯群丛: 由于早年建设高压铁塔时, 岛上植被遭到严重破坏, 建设单位引入马缨丹进行植被修复, 现今几乎占据了整个灌木层.马缨丹具有强烈的化感作用, 在岛上形成单优群落, 压制或排挤本地种, 严重影响了海岛物种多样性.岛屿作为孤立的大陆, 经常维持着种群规模小、繁殖力低、防御力弱的简单生态系统(Spatz, 2017 ), 岛屿物种更容易遭受各种外来因素的影响.入侵物种还可通过杂交和基因渗入的方式入侵到土著植物的基因库中, 造成土著植物基因型的丧失, 并有可能导致土著植物的灭绝, 这种杂交很有可能使产生的后代比亲本具有更强的入侵性(万宣伍等, 2015 ), 从而导致更为严重的生态后果, 需要引起重视. ...
基于分子生物学方法的外来入侵物种入侵历史重构
1
2015
... 总体来看, 厦门周边的海岛维管束植物以乡土植物为主, 但岛上的入侵物种不容忽视.例如猴屿上的马缨丹纯群丛: 由于早年建设高压铁塔时, 岛上植被遭到严重破坏, 建设单位引入马缨丹进行植被修复, 现今几乎占据了整个灌木层.马缨丹具有强烈的化感作用, 在岛上形成单优群落, 压制或排挤本地种, 严重影响了海岛物种多样性.岛屿作为孤立的大陆, 经常维持着种群规模小、繁殖力低、防御力弱的简单生态系统(Spatz, 2017 ), 岛屿物种更容易遭受各种外来因素的影响.入侵物种还可通过杂交和基因渗入的方式入侵到土著植物的基因库中, 造成土著植物基因型的丧失, 并有可能导致土著植物的灭绝, 这种杂交很有可能使产生的后代比亲本具有更强的入侵性(万宣伍等, 2015 ), 从而导致更为严重的生态后果, 需要引起重视. ...
温州沿海小型海岛植物丰富度和β多样性及其影响因子
2
2017
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
... 物种相似性系数C j 与生境类型数目显著正相关, 与周长/面积比(PAR )显著负相关, 与其他参数无显著相关性.厦门周边12个无居民海岛中, 物种组成相似性较高的有: 大兔屿和白兔屿, 鳄鱼屿和白兔屿, 大兔屿和鳄鱼屿, 鸡屿和大兔屿, 猴屿和鳄鱼屿, 白兔屿和海沧大屿.如上文所提, 生境异质性是决定物种丰富度的主要因素.周长/面积比越大, 即边缘越大, 相同面积内可供植物生长的实际面积减小, 因此物种丰富度越低, 相应的物种相似性也较低.总体上, 不同岛屿间维管束植物物种相似性受生境多样性和岛屿边缘效应影响, 该结果与王金旺等(2017) 的研究一致.此外, 我们发现吾屿与大多数岛屿的物种相似性均较低, 可能是由于其面积较小, 导致维管束植物种类数少, 一定程度上反映吾屿可能出现小岛屿效应. ...
温州沿海小型海岛植物丰富度和β多样性及其影响因子
2
2017
... 早在达尔文时期, 人们以海岛为对象的科学研究就已经开始.岛屿生物地理学理论认为, 物种组成的变化会导致岛屿物种丰富度出现波动, 而岛屿物种丰富度主要取决于岛屿面积和隔离程度(Pinheiro et al, 2017 ).物种相似性系数常用于分析不同区域群落间物种组成的差异(Qian et al, 2005 ).此外, 有研究表明, 岛屿面积大小、形状、隔离程度等空间特征参数对物种多样性均有显著影响(王金旺等, 2017 ).种-面积关系(species-area relationship, SAR)定量表达了物种丰富度随取样面积增大而不断增加的变化趋势, 从逻辑上可推出, 岛屿面积小到一定程度将仅能够支持极少数物种存在或者由于动态波动某些时刻这些小型岛屿上将不存在物种(Morrison, 2011 ).但越来越多的生物地理和物种多样性研究案例表明, 当岛屿面积小于某个特定阈值时, 岛屿物种丰富度将不再符合种-面积关系, 即存在小岛屿效应(small island effect, SIE) (Lomolino, 2000 ). ...
... 物种相似性系数C j 与生境类型数目显著正相关, 与周长/面积比(PAR )显著负相关, 与其他参数无显著相关性.厦门周边12个无居民海岛中, 物种组成相似性较高的有: 大兔屿和白兔屿, 鳄鱼屿和白兔屿, 大兔屿和鳄鱼屿, 鸡屿和大兔屿, 猴屿和鳄鱼屿, 白兔屿和海沧大屿.如上文所提, 生境异质性是决定物种丰富度的主要因素.周长/面积比越大, 即边缘越大, 相同面积内可供植物生长的实际面积减小, 因此物种丰富度越低, 相应的物种相似性也较低.总体上, 不同岛屿间维管束植物物种相似性受生境多样性和岛屿边缘效应影响, 该结果与王金旺等(2017) 的研究一致.此外, 我们发现吾屿与大多数岛屿的物种相似性均较低, 可能是由于其面积较小, 导致维管束植物种类数少, 一定程度上反映吾屿可能出现小岛屿效应. ...
福建省岛屿地质环境学科发展研究报告
1
2015
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
福建省岛屿地质环境学科发展研究报告
1
2015
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
Effects of changing scale on landscape pattern analysis: Scaling relations
1
2004
... 根据国家海洋信息中心海岛监视监测系统内的海岛资料(表1 ), 利用岛屿面积(A )、岸线长度(Per )、距大陆距离(I m )、岛屿间距离(I i )、岛屿高程(E )、周长/面积比(PAR )、形状指数(SI )等参数来描述调查岛屿的空间特征, 其中I m 表示岛屿边缘至大陆的最小直线距离, PAR 为岛屿边缘的相对长度, 表征岛屿的边界效应(Wu, 2004 ).SI 反映岛屿形状的复杂程度, 由公式SI = Per /[2×(π×A )×0.5]计算. ...
Evolution and measurement of species diversity
1
1972
... 采用Jaccard相似性系数C j = j /(a + b - j )计算不同岛屿间的相似性(Whittaker, 1972 ), 其中j 为不同岛屿间共有的物种数, a 和b 分别为不同岛屿自身包含的物种数目. ...
中国种子植物属的分布区类型
1
1991
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
中国种子植物属的分布区类型
1
1991
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
世界种子植物科的分布区类型系统
2
2003
... 植物区系是植物类群(科、属、种)分布式基本一致的再现, 影响其组成的因素不仅包括水、肥、气、热等生态因子, 还包括影响分布区形成的地质事件及所属分布区发展的历史植物的地理因素(吴征镒等, 2003 ).因为海岛生态系统具有独立性、完整性、脆弱性, 形成了特殊的海岛植被类型, 研究海岛的植物区系分化, 是进行海岛植被资源合理利用和保护的基础, 对于目前国家提倡的海岛整治修复尤其是海岛植被保护和修复具重要意义. ...
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
世界种子植物科的分布区类型系统
2
2003
... 植物区系是植物类群(科、属、种)分布式基本一致的再现, 影响其组成的因素不仅包括水、肥、气、热等生态因子, 还包括影响分布区形成的地质事件及所属分布区发展的历史植物的地理因素(吴征镒等, 2003 ).因为海岛生态系统具有独立性、完整性、脆弱性, 形成了特殊的海岛植被类型, 研究海岛的植物区系分化, 是进行海岛植被资源合理利用和保护的基础, 对于目前国家提倡的海岛整治修复尤其是海岛植被保护和修复具重要意义. ...
... 以整岛踏查为主, 样线和样方调查为辅.详细记录12个岛上的植物群落类型、优势种、种类组成并拍照, 疑难种类采集标本.乔木群落样方的面积为10 m × 10 m, 灌木样方为5 m × 5 m, 草本样方为1 m × 1 m, 样方设置囊括不同的植物群落, 每类样方数量不少于2个.现场记录植被类型、株高、盖度等信息, 形成解译标志.利用GPS确定样方位置.按照植物群落调查的常规方法调查种类组成、优势种、乔木树种的数量和大小(方精云等, 2009 ).并根据《世界种子植物科的分布区类型系统》(吴征镒等, 2003 )、《中国种子植物属的分布区类型》(吴征镒, 1991 ), 对12个无居民海岛分布区类型进行划分. ...
山东无棣贝壳堤保护区维管束植物的生态位研究
1
2018
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
山东无棣贝壳堤保护区维管束植物的生态位研究
1
2018
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
鼓浪屿岛种子植物分布区类型初步研究
2
2016
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
鼓浪屿岛种子植物分布区类型初步研究
2
2016
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
中国外来入侵植物的等级划分与地理分布格局分析
1
2014
... 厦门近岸海域海岛合计有入侵植物49种, 占总物种数的13.6%.根据中国外来入侵植物信息系统(http://ias.iplant.cn/protlist)公布的中国入侵植物名录及入侵等级(闫小玲等, 2014 ), 12个无居民海岛均有恶性入侵植物, 共10种, 分别是土荆芥(Chenopodium ambrosioides )、银胶菊(Parthenium hysterophorus )、钻叶紫菀(Aster subulatus )、光荚含羞草(Mimosa sepiaria )、互花米草(Spartina alterniflora )、藿香蓟(Ageratum conyzoides )、蒺藜草(Cenchrus echinatus )、马缨丹(Lantana camara )、白花鬼针草(Bidens pilosa var. radiata )、假臭草(Eupatorium catarium ).其中鸡屿和大兔屿恶性入侵植物最多, 分别多达7种和6种, 占比分别为5.98%和5.22%.其次是严重入侵物种, 12个海岛共计14种, 除吾屿和宝珠屿以外, 其他10个海岛上或多或少存在严重入侵物种.此外, 12个海岛局部入侵类5种、一般入侵类5种、有待观察类15种.总体来看, 厦门周边的海岛维管束植物以乡土植物为主. ...
中国外来入侵植物的等级划分与地理分布格局分析
1
2014
... 厦门近岸海域海岛合计有入侵植物49种, 占总物种数的13.6%.根据中国外来入侵植物信息系统(http://ias.iplant.cn/protlist)公布的中国入侵植物名录及入侵等级(闫小玲等, 2014 ), 12个无居民海岛均有恶性入侵植物, 共10种, 分别是土荆芥(Chenopodium ambrosioides )、银胶菊(Parthenium hysterophorus )、钻叶紫菀(Aster subulatus )、光荚含羞草(Mimosa sepiaria )、互花米草(Spartina alterniflora )、藿香蓟(Ageratum conyzoides )、蒺藜草(Cenchrus echinatus )、马缨丹(Lantana camara )、白花鬼针草(Bidens pilosa var. radiata )、假臭草(Eupatorium catarium ).其中鸡屿和大兔屿恶性入侵植物最多, 分别多达7种和6种, 占比分别为5.98%和5.22%.其次是严重入侵物种, 12个海岛共计14种, 除吾屿和宝珠屿以外, 其他10个海岛上或多或少存在严重入侵物种.此外, 12个海岛局部入侵类5种、一般入侵类5种、有待观察类15种.总体来看, 厦门周边的海岛维管束植物以乡土植物为主. ...
中国台湾种子植物区系的性质、特点及其与大陆植物区系的关系
2
2002
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... 厦门近岸海域12个海岛与厦门本岛(厦门同安区内田村)相比, 相同点为热带性质的属占大多数, 不同之处在于12个海岛热带至亚热带性质属的比例更高, 占73.02%, 而内田村的为64.95%.该结果与林文俊(2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)对比闽台植物区系所得结论相一致, 台湾岛的热带属性质强于邻近大陆福建.福建热带分布属占总属数的60.34%, 温带分布属占总属数的24.10%, 表现出明显的热带性质, 厦门近岸海域12个海岛热带性质的属占总属数的73.02%, 温带性质的属占17.27%, 热带属性质高于福建整体水平(林文俊, 2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.).同台湾植物区系相比, 厦门周边海域12个海岛热带属性质也高于台湾(热带分布属占总属数的66.21%), 温带分布属性质低于台湾(温带分布属占22.47%)(应俊生等, 2002 ; 林文俊, 2013① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)).这与海岛的气候环境和物理性质有关, 厦门近岸海域海岛相对而言环境更为恶劣, 海岛面积较小, 受到大风、阳光直射的影响较大, 温带植物分布较少, 热带至亚热带性质的植物更适宜生长. ...
中国台湾种子植物区系的性质、特点及其与大陆植物区系的关系
2
2002
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... 厦门近岸海域12个海岛与厦门本岛(厦门同安区内田村)相比, 相同点为热带性质的属占大多数, 不同之处在于12个海岛热带至亚热带性质属的比例更高, 占73.02%, 而内田村的为64.95%.该结果与林文俊(2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)对比闽台植物区系所得结论相一致, 台湾岛的热带属性质强于邻近大陆福建.福建热带分布属占总属数的60.34%, 温带分布属占总属数的24.10%, 表现出明显的热带性质, 厦门近岸海域12个海岛热带性质的属占总属数的73.02%, 温带性质的属占17.27%, 热带属性质高于福建整体水平(林文俊, 2013)① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.).同台湾植物区系相比, 厦门周边海域12个海岛热带属性质也高于台湾(热带分布属占总属数的66.21%), 温带分布属性质低于台湾(温带分布属占22.47%)(应俊生等, 2002 ; 林文俊, 2013① (① 林文俊 (2013) 闽台种子植物区系比较研究. 硕士学位论文, 福建农林大学, 福州.)).这与海岛的气候环境和物理性质有关, 厦门近岸海域海岛相对而言环境更为恶劣, 海岛面积较小, 受到大风、阳光直射的影响较大, 温带植物分布较少, 热带至亚热带性质的植物更适宜生长. ...
福建植物区系的由来
3
1981
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
... ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
福建植物区系的由来
3
1981
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
... ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
福建植物区系与植物地理区域
1
1983
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
福建植物区系与植物地理区域
1
1983
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
厦门大兔屿种子植物区系分析及其植被修复对策研究
1
2017
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
厦门大兔屿种子植物区系分析及其植被修复对策研究
1
2017
... 厦门近岸海域12个海岛植物区系组成基本符合其亚热带地理分布特点.张琳婷等(2017) 、徐河山等(2016) 分析了厦门大兔屿和鼓浪屿岛的植物区系特征, 发现均以热带成分为主, 但属于热带分布属的北缘.泛热带分布及其变型在厦门同安县内田(今厦门同安区内田村) (谭忠奇等, 1996 )和台湾也占最大比例, 并以热带至亚热带性质居多(应俊生等, 2002 ), 鼓浪屿植物区系具有热带性亲缘, 但因其地处热带向温带过渡的南亚热带地区, 所以温带成分也占有一定比例(徐河山等, 2016 ).福建现代的地理位置是处于泛北极植物区最南端, 是泛北极植物区与古热带植物区的过渡带, 往南紧接着是古热带植物区, 是我国亚热带植物区系的摇篮和发源地之一(曾文彬等, 1981 ), 因此厦门周边海岛主要植物区系与泛热带分布及其变型有一定的历史渊源. ...
1
2012
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
1
2012
... 同山东贝壳堤岛新生土地上植物种类不同(肖兰等, 2018 ), 福建沿海岛屿多数为近岸岛, 基本架构受大陆地质地貌控制(张耀光, 2012 ).根据《中国海湾志第八分册(福建省南部海湾)》对厦门港的记载, 早新生代以来厦门港区内受喜马拉雅运动影响, 表现为断块升降, 使厦门港区沿海岛屿与大陆分离(陈则实, 1993 ).第三纪末至第四纪初, 发生的喜马拉雅运动导致台湾海峡发生断裂, 东南沿海地区自第四纪以来还遭过海浸(曾文彬等, 1981 ).厦门周边海域海岛属于第四纪地质的新构造分区中的闽中轻微上升区(王仁山等, 2015 ).本次调查发现, 厦门周边海域12个无居民海岛, 除海沧乌鸦屿和土屿由于面积太小外, 其余10个均保留有福建生长的古老植物的直接后裔, 例如蕨类中的海金沙属(Lygodium )、裸子植物中的杉木属(Cunninghamia )、松属(Pinus ), 它们都是新生代或中生代的古老科属.其中芒萁(Dicranopteris dichotoma )是中生代三叠纪就已出现的, 海金沙(Lygodium japonicum )是白垩纪已存在的古老孑遗植物(曾文彬等, 1983 ).本次12个海岛调查到的福建古老植物有: 中生代白垩纪出现的樟科、木兰科、桑科、豆科、夹竹桃科、蔷薇科、毛茛科、五加科、冬青科、芸香科、漆树科、大戟科、蓼科等, 新生代第三纪以后出现的有黎科、榆科、葡萄科、鼠李科、卫矛科、山茶科、桃金娘科、唇形科、菊科、禾本科等.这些古老科属占福建现代植物区系成分中的大部分, 福建现代植物区系是福建第三纪古老植物区系的发展和直接延伸(曾文彬等, 1981 ).位于厦门白鹭自然保护区核心区的鸡屿, 受人为活动干扰少, 较好地保持了原生植被类型, 其岛上潺槁木姜子(Litsea glutinosa )群落、马尾松(Pinus massoniana )群落、林下的蕨类群落都和福建大陆植物群落类型相似.本次调查的12个海岛除土屿外均为沿岸岛, 为此我们进一步猜测, 这12个海岛原与大陆相连, 由于遭受到海浸和不断活动上升的板块运动, 导致与大陆板块分离, 逐渐形成海岛.面积较大的海岛仍保留着原大陆的植物类群, 而面积较小的海岛上因长期遭受海上大风、盐雾、风暴潮等的影响, 原陆生植物在恶劣的环境中经历漫长的自然选择, 不适应的种群先后被淘汰, 适应性强的留存下来, 植物群落类型逐渐发生变化, 形成典型的海岛植被类型.总体上, 本次调查的无居民海岛资源数据完整, 能够代表厦门周边海域无居民海岛的植物资源信息.结合岛屿地理资料和现场调查结果, 摸清了厦门近岸无居民海岛的主要群落类型(附录2). ...
平潭大屿岛植物资源及其多样性研究
1
2016
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
平潭大屿岛植物资源及其多样性研究
1
2016
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
外马廊山岛植被特性与植物多样性
1
2017
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
外马廊山岛植被特性与植物多样性
1
2017
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应
1
2018
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应
1
2018
... 近年来, 国内针对海岛植物区系以及植物多样性的研究开始丰富起来.如郑俊鸣等(2016 , 2017 )对平潭大屿岛的植物资源及其群落多样性、舟山无居民海岛外马廊山岛的植被特征与植物多样性进行了调查, 发现两个岛屿植物种类偏少, 以高位芽植物为主, 并且群落结构简单, 多样性偏低, 还存在一定的生态干扰; 孙元敏等(2017) 调查比较了厦门无居民海岛猴屿人工恢复林与次生林的物种多样性和肥力, 次生林肥力显著高于人工恢复林, 二者多样性差异不显著; 郑丽婷等(2018) 认为庙岛群岛的4种典型植物群落多样性特征既有与大陆植被相似的特征, 也有海岛特殊性.这些研究多停留在小区域范围内少数海岛植物群落的研究. ...
 , 杨盛昌
, 杨盛昌

{kind=link}
{kind=link}
{kind=link}
{kind=link}