自然杂交指的是已发生遗传分化的种群之间的交配(Arnold, 1992 ; Allendorf et al, 2001 ), 一般更容易发生在亲缘关系较近的类群间。然而, 蕨类植物的一个案例表明, 分化六千万年的物种之间也可以发生杂交(Rothfels et al, 2015 )。大部分自然杂交后代(hybrid)由于生殖隔离造成的不育而被淘汰, 或存在于杂交带中成为亲本间基因流的桥梁; 但仍有一部分杂交后代通过各种机制得以留存而形成自然杂交种(hybrid species), 如同倍杂交后代由于适应新的环境与父母本产生生殖隔离, 或通过异源染色体加倍的方式直接和父母本产生生殖隔离(生物种概念)(Mallet, 2007 ); 而一些奇数倍染色体的杂交后代通过无性繁殖得以延续, 由于其形态的可辨识性, 也可能被命名为杂交种(形态种)。
自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议。传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象。另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a )。
随着自然杂交研究的增多, 人们越来越认识到物种杂交是自然界的普遍现象, 而且是植物物种形成的主要驱动力。杂交有害的情况更多地发生在同倍体杂交中, 而多倍化杂交物种往往比其亲本更利于生态位建成和适应性增强。由此带来的混合基因库具备更高的遗传多样性和快速适应新环境的潜力, 对生物多样性的保持具有一定的积极作用。所以, 对其保护价值的讨论具有重要意义(Anderson & Stebbins Jr, 1954 ; Nolte & Tautz, 2010 ; Piett et al, 2015 ; Kasari et al, 2016 )。
Jackiw等(2015)对北美的物种保护条例进行了研究, 发现虽然人们已经开始关注到自然杂交的保护, 但是条例中涉及的条款很少, 并且往往语焉不详, 执行情况也不理想。世界自然保护联盟(IUCN)的物种红色名录里, 甚至明确说明要把自然杂交种剔除, 完全忽略其价值(IUCN, 2013 , 2014 )。为此, Jackiw等(2015)针对北美的情况制定了一个评估杂交是否值得保护的原则框架, 他建议保育工作者在制定法规和具体的保护工作中, 系统地考虑杂交的成因、对父母本的威胁程度, 以及对生态环境的影响等。这些评估原则对中国的保护工作也有很好的借鉴意义。
在中国, 自然杂交的保护问题还没有引起人们的足够关注, 1996年颁布的《野生植物保护条例》和2016年颁布的《野生动物保护法》都没有明确提及自然杂交种是否需要保护的问题。但在已有的保护名录里实际上已经包括了自然杂交种, 例如《国家重点保护野生植物名录》(1999)中的国家二级保护植物半枫荷(Semiliquidambar cathayensis )是杂交起源(吴伟, 2009 ; Wu et al, 2010 )。相信随着保护研究的进展, 类似的例子以及相关的讨论会越来越多, 而这也是保护生物学无法回避的问题。
本文结合近些年有关自然杂交保护生物学的研究进展, 特别是Jackiw等(2015) 提出的自然杂交保护的策略框架, 拟探讨: (1)保护自然杂交的意义; (2)结合中国现在的自然杂交保护现状, 我们应该如何保护自然杂交?
1 自然杂交的保护价值
人们对自然杂交的顾虑往往在于其可能对父母本以及生态系统产生潜在威胁。但很多情况下, 自然杂交带来的影响并不都是有害的, 需要结合实际情况来进行科学的判断。保护生物学最主要的目的就是保护生物多样性, 而自然杂交是否值得保护也要从多样性的角度去考虑。
1.1 遗传多样性
不完全生殖隔离导致的杂交渐渗现象中, 物种间的基因流对物种演化和生态适应起到非常重要的作用。由于父母本双方基因库的混合而产生新的等位基因组合, 杂交带往往比父母本种群具有更高的遗传多样性以及适应性(Yakimowski & Rieseberg, 2014 )。当渗透的等位基因受到选择时, 适度的渗透可以使物种快速获得适应性。这种基因交流可以给父母本物种提供一个快速适应新环境、提高适合度的机会。从这个角度来说, 适度的自然杂交显然是有利的。只有当渗透过度时, 才可能造成生殖隔离的突破而造成物种逆转。
另外, 当父母本有一方或双方由于其自身适合度低下而濒临灭绝时, 杂交种可能会因杂种优势具有较高适合度, 成为保存其遗传多样性的有效方式。例如Hori等(2014)利用核基因研究日本的变异鳞毛蕨(Dryopteris varia )复合群网状进化时, 发现了一个不属于任何现存物种的单倍型, 推测其来源于一个已灭绝的物种。
1.2 物种多样性
自然杂交不仅可以增加遗传多样性和物种的适应性, 而且可以导致物种的辐射式演化, 促进大量物种在相对较短的时期内形成(Seehausen, 2004 )。
首先, 大量现存物种是通过自然杂交起源的, 或者处于自然杂交物种形成的演化途中(Abbott et al, 2013 ), 网状进化是一种重要的物种演化模式。一方面, 杂交后代可以通过染色体加倍形成异源多倍化物种(Soltis et al, 2014 ); 另一方面, 由于同倍性的杂交后代与父母本间的生殖隔离不如异倍杂交那样容易产生, 所以同倍性杂交物种比较少见, 但仍然是不可忽视的物种形成方式之一。例如, 生长在粘性土壤中的向日葵(Helianthus annuus )和其生长在砂质土壤中的近亲植物草原太阳花(H. petiolaris ), 产生了3种适应不同环境的自然杂交种(H. anomalus、H. deserticola 和H. paradoxus ) (Rieseberg, 1991 )。
其次, 在同域物种形成的过程中, 当新物种和旧物种的生殖隔离尚未完全建立时, 杂交渗透所形成的物种间基因流可能会导致适应不同生态位的基因被选择作用固定在不同的物种中, 促进物种的分化(Coyne & Orr, 2004 ; Rolán-Alvarez, 2007 )。
总之, 自然杂交是自然物种进化历程中非常重要的组成部分, 其价值理应受到重视。
1.3 生态系统多样性
一般认为, 生态系统组成的多样性与稳定性存在某种程度的相关性, 虽然May (1976) 从数学上论证了多样性有时会导致稳定性下降的可能性, 但在多数情况下, 通过增加生态系统内的多样性, 可促进系统的稳定性。自然杂交在生态系统中的存在, 则可能通过影响遗传多样性和物种多样性进而影响生态系统的稳定性和可恢复性。
杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 )。实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 )。如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 )。此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 )。总之, 自然杂交对维持生态系统的稳定性起到很重要的作用。
2 自然杂交的保护原则
保护生物学家在自然杂交的保护领域已经开始了理论探讨和实践尝试。除Jackiw等(2015)的工作以外, Richards和Hobbs (2015) 针对珊瑚礁群落中的杂交保护问题进行了讨论, 首先综述了珊瑚虫以及岩礁鱼类杂交研究进展, 并据此评估了杂交对珊瑚礁群落多样性的影响。分析结果表明, 杂交可以造成多种进化生物学后果, 既有可能是增加遗传多样性, 产生新的类群, 也有可能造成物种逆转。由于对自然杂交保护立法的忽视, 可能使得有价值的自然杂交没有得到有效的保护而导致其灭绝。针对这种情况, 他们提出, 应逐个案例(case-by-case)评估杂交的原因和后果, 有针对性地决定是保护还是清除杂交。
综合这些意见, 对自然杂交的成因以及后果分别进行科学评估是采取适宜保护措施的前提。由于自然杂交对其父母本物种、物种多样性乃至生态系统具有重要意义, 因此在制定野生动植物保护法律法规以及编写红色名录的时候就要根据具体的情况进行科学的评估。
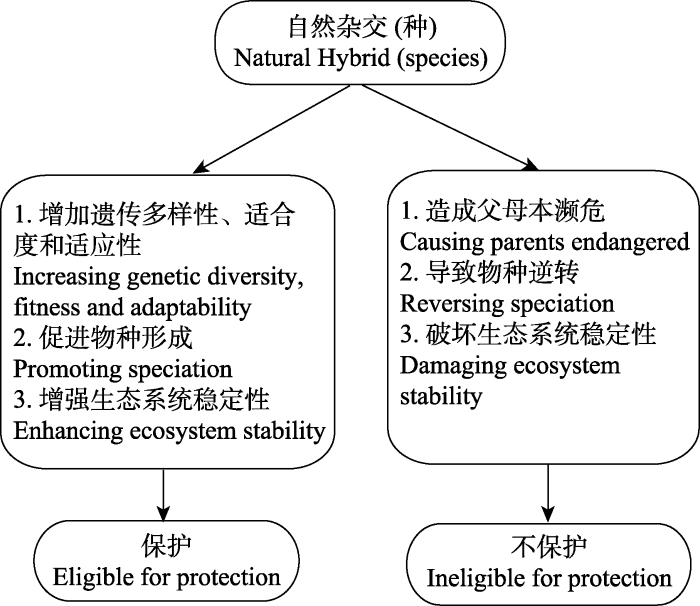
有鉴于此, 我们参考国际同行的观点, 并考虑到中国自然杂交保护的现状, 提出有关中国自然杂交的保护原则(图1 )。首先应对自然杂交保护的对象进行明确的规定: (1)该自然杂交后代已独立成种并能在自然环境中独立建立自我繁殖体系(具有稳定基因交流的自然杂交带除外); (2)该自然杂交后代或自然杂交种受到了生存威胁或在IUCN等级评估标准中处于受威胁状态, 或者对维持或保存父母本遗传多样性、对群落物种多样性、生态系统稳定性以及对人类社会具有显著的意义。涉及濒危的自然杂交种可以考虑列入物种红色名录; 涉及生态保护价值或社会经济价值、科学研究价值的则列入相应类型的保护名录。据此, 将关于自然杂交的保护策略归纳为以下4个方面:
图1 自然杂交种的保护原则
Fig. 1 Principle for the conservation of natural hybridization
(1)自然杂交指的是近期或者历史上自然发生的没有受到人为因素干扰的物种杂交事件, 包括自然杂交种、基因渐渗和杂交带等。这里需要指出的是, 用于可操作的自然保护的自然杂交对象一般是指具有相对稳定的遗传特征的杂交事件, 可以是能够自我繁殖的自然杂交种, 如孟连铁线蕨(Adiantum menglianense )与假鞭叶铁线蕨(A. malesianum )和苍山铁线蕨(A. sinense )分别产生的自然杂交后代: 梅山铁线蕨(A. meishanianum )和哀牢山铁线蕨(A. ailaoshanense ), 均可以通过无性繁殖在自然界自我维系自然种群(Wang et al, 2015b ; Shang et al, 2016 ); 也可以是由于杂交渐渗在自然杂交带中产生的杂交群体, 如分布于丽江玉龙雪山的橘红灯台报春(Primula bulleyana )和霞红灯台报春(P. beesiana )可以在其分布重叠区内高频率发生自然杂交并形成杂交后代。虽然其杂交后代不育, 但杂交后代通过回交可能对亲本的遗传多样性产生重要的影响(Hayashi et al, 2009 )。因此可以视其濒危程度及其亲本的生存状况等因素, 酌情考虑保护措施。但是, 对自然界中偶尔产生的一些自然杂交后代, 由于遗传上的不稳定性和发生频率的随机性, 一般不需要列入保护对象, 如粤紫萁(Osmunda mildei )是华南紫萁(O. vachellii )和紫萁(O. japonica )的自然杂交后代(Tsutsumi et al, 2012 ), 其杂交后代不具可育性, 无法在野外独立建立种群, 而是仅在亲本的重叠分布区产生极少量的随机个体, 因此也很难对该自然杂交个体进行有效的保护。
(2)当父母本一方或双方濒危, 但杂交后代或杂交种没有威胁到其父母本时, 自然杂交应该被保护。日本西部特有分布的稀子蕨属植物Monachosorum arakii 是亚洲大陆分布的稀子蕨(M. henryi )和日本分布的日本稀子蕨(M. nipponicum )的自然杂交产物, 该杂交种可以通过无性繁殖芽孢自行在野外建立种群。由于稀子蕨现仅分布于中国大陆以及热带亚洲地区, 在日本已经灭绝, 稀子蕨和日本稀子蕨的杂交可能是过去地质时代的孑遗产物(Ebihara et al, 2016 )。在此, 由于杂交没有威胁到父母本的生存, 自然杂交种Monachosorum arakii 在日本的分布具有重要科学研究价值并可维系已灭绝的亲本遗传信息, 因此该自然杂交种应该受到必要的保护。
(3)当父母本双方都不濒危, 并且杂交后代或杂交种没有威胁到其父母本时, 自然杂交应该被保护。高山松(Pinus densata )是油松(P. tabuliformis )和云南松(P. yunnanensis )的自然杂交种(Wang et al, 2001 ), 是乔木物种二倍体自然杂交同倍体成种的范例, 广泛分布于云南、四川、西藏、青海等地, 生长于海拔1,500-4,500 m的地区。但其父母本均不濒危, 且高山松没有威胁到父母本的生存; 由于高山松具有较大的社会经济价值, 因此可以列入林业系统的珍稀林木保护名录; 然而, 由于高山松分布广泛, 其野外种群没有受到明显的威胁, 目前尚不需要将该种列入物种红色名录。
(4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定。如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护。如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛。大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁。鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护。相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护。厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀。由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 )。同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 )。对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护。
3 结语
评估自然杂交的保护价值需要综合考虑各方面的因素, 不能一概而论。保护生物学的目的并不是保护物种纯正的血统, 而是应该更多地从生物多样性和生物多样化过程的角度去考虑问题并评估保护价值。我们提出的保护策略可以为实际物种保护工作以及相关法律法规的制定提供参考, 而中国现行野生植物保护政策中, 将部分类群如水韭属(Isoetes )、桫椤科、红豆杉属(Taxus )、兰科等作为一个整体加以保护, 对保护该类群的自然杂交无疑具有积极意义。然而, 目前需要更多的关于自然杂交的科学研究以及更广泛的科普宣传, 使人们进一步了解自然杂交在生物进化历程以及生态系统中的重要作用。
致谢:感谢马永鹏研究员、邓敏研究员、金冬梅博士、刘子玥、刘逸在本文写作过程中给予的帮助和建议; 感谢审稿专家提出的修改意见, 极大地提升了本文的专业性和逻辑的严谨性。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Abbott R Albach D Ansell S Arntzen JW Baird SJ Bierne N Boughman J Brelsford A Buerkle CA Buggs R 2013 ) Hybridization and speciation.
Journal of Evolutionary Biology , 26 , 229 -246 .
[本文引用: 1]
[2]
Allendorf FW Leary RF Spruell P Wenburg JK 2001 ) The problems with hybrids: setting conservation guidelines.
Trends in Ecology & Evolution , 16 , 613 -622 .
[本文引用: 1]
[3]
Anderson E Stebbins G Jr 1954 ) Hybridization as an evolutionary stimulus.
Evolution , 378 -388 .
[本文引用: 1]
[4]
Arnold ML 1992 ) Natural hybridization as an evolutionary process.
Annual Review of Ecology and Systematics , 23 , 237 -261 .
[本文引用: 1]
[5]
Bohling JH 2016 ) Strategies to address the conservation threats posed by hybridization and genetic introgression.
Biological Conservation , 203 , 321 -327 .
[本文引用: 1]
[6]
Coyne JA Orr HA 2004 ) Speciation
. Sinauer Associates Inc., Sunderland, MA .
[本文引用: 1]
[7]
Ebihara A Nakato N Amoroso VB Hidayat A Kuo LY 2016 ) Monachosorum arakii Tagawa (Dennstaedtiaceae) is a relict “international” hybrid: a reassessment of the Monachosorum species.
Systematic Botany , 41 , 586 -595 .
[本文引用: 1]
[8]
Fitzpatrick BM Shaffer HB 2007 ) Hybrid vigor between native and introduced salamanders raises new challenges for conservation
. Proceedings of the National Academy of Sciences, USA , 104 , 15793 -15798 .
[本文引用: 1]
[9]
Gow JL Peichel CL Taylor EB 2006 ) Contrasting hybridization rates between sympatric three-spined sticklebacks highlight the fragility of reproductive barriers between evolutionarily young species.
Molecular Ecology , 15 , 739 -752 .
[本文引用: 1]
[10]
Grant PR Grant BR 2014 ) Evolutionary biology: speciation undone.
Nature , 507 , 178 -179 .
[本文引用: 1]
[11]
Gross B Rieseberg L 2005 ) The ecological genetics of homoploid hybrid speciation.
Journal of Heredity , 96 , 241 -252 .
[本文引用: 1]
[12]
Hayashi M Kato J Ohashi H Mii M 2009 ) Unreduced 3x gamete formation of allotriploid hybrid derived from the cross of Primula denticulata (4x) × P. rosea (2x) as a causal factor for producing pentaploid hybrids in the backcross with pollen of tetraploid P. denticulata.
Euphytica , 169 , 123 .
[本文引用: 1]
[13]
Hill KD 1993 ) The endangered species act: what do we mean by species.
Boston College Environmental Affairs Law Review , 20 , 239 .
[本文引用: 1]
[14]
Hori K Tono A Fujimoto K Kato J Ebihara A Watano Y Murakami N 2014 ) Reticulate evolution in the apogamous Dryopteris varia complex (Dryopteridaceae, subg. Erythrovariae, sect. Variae) and its related sexual species in Japan.
Journal of Plant Research , 127 , 661 -684 .
[15]
IUCN (2013 ) The IUCN Red List of Threatened Species
. IUCN, Gland, Switzerland .
[本文引用: 1]
[16]
IUCN (2014 ) Guidelines for Using the IUCN Red List Categories and Criteria, Version 11
. IUCN, Gland, Switzerland .
[本文引用: 1]
[17]
Jackiw RN Mandil G Hager HA 2015 ) A framework to guide the conservation of species hybrids based on ethical and ecological considerations.
Conservation Biology , 29 , 1040 -1051 .
[本文引用: 1]
[18]
Kasari L Saar L de Bello F Takkis K Helm A 2016 ) Hybrid ecosystems can contribute to local biodiversity conservation.
Biodiversity and Conservation , 25 , 3023 -3041 .
[本文引用: 1]
[19]
Kleindorfer S O’Connor JA Dudaniec RY Myers SA Robertson J Sulloway FJ 2014 ) Species collapse via hybridization in Darwin’s tree finches.
The American Naturalist , 183 , 325 -341 .
[本文引用: 1]
[20]
Mallet J 2007 ) Hybrid speciation.
Nature , 446 , 279 .
[本文引用: 1]
[21]
May R 1976 ) Theoretical Ecology
. Saunders, Philadelphia .
[本文引用: 1]
[22]
Nolte AW Tautz D 2010 ) Understanding the onset of hybrid speciation.
Trends in Genetics , 26 , 54 -58 .
[本文引用: 1]
[23]
Piett S Hager HA Gerrard C 2015 ) Characteristics for evaluating the conservation value of species hybrids.
Biodiversity and Conservation , 24 , 1931 -1955 .
[本文引用: 1]
[24]
Richards ZT Hobbs JPA 2015 ) Hybridisation on coral reefs and the conservation of evolutionary novelty.
Current Zoology , 61 , 132 -145 .
[本文引用: 1]
[25]
Rieseberg LH 1991 ) Homoploid reticulate evolution in Helianthus (Asteraceae): evidence from ribosomal genes.
American Journal of Botany , 1218 -1237 .
[本文引用: 1]
[26]
Rolán-Alvarez E 2007 ) Sympatric speciation as a by-product of ecological adaptation in the Galician Littorina saxatilis hybrid zone.
Journal of Molluscan Studies , 73 , 1 -10 .
[本文引用: 1]
[27]
Rothfels CJ Johnson AK Hovenkamp PH Swofford DL Roskam HC Fraser-Jenkins CR Windham MD Pryer KM 2015 ) Natural hybridization between genera that diverged from each other approximately 60 million years ago.
The American Naturalist , 185 , 433 -442 .
[本文引用: 1]
[28]
Seehausen O 2004 ) Hybridization and adaptive radiation.
Trends in Ecology & Evolution , 19 , 198 -207 .
[本文引用: 1]
[29]
Seehausen O 2006 ) Conservation: losing biodiversity by reverse speciation.
Current Biology , 16 , R334 -R337 .
[本文引用: 1]
[30]
Shang H Wang Y Zhu XF Zhao GH Wang FH Lu JM Yan YH 2016 ) Likely allopatric origins of Adiantum× meishanianum (Pteridaceae) through multiple hybridizations.
Journal of Systematics and Evolution , 54 , 528 -534 .
[本文引用: 1]
[31]
Soltis DE Visger CJ Soltis PS 2014 ) The polyploidy revolution then and now: Stebbins revisited.
American Journal of Botany , 101 , 1057 -1078 .
[本文引用: 1]
[32]
Stelkens RB Brockhurst MA Hurst GD Greig D 2014 ) Hybridization facilitates evolutionary rescue.
Evolutionary Applications , 7 , 1209 -1217 .
[本文引用: 1]
[33]
Taylor E Boughman J Groenenboom M Sniatynski M Schluter D Gow J 2006 ) Speciation in reverse: morphological and genetic evidence of the collapse of a three- spined stickleback (Gasterosteus aculeatus) species pair.
Molecular Ecology , 15 , 343 -355 .
[本文引用: 1]
[34]
Tsutsumi C Hirayama Y Kato M Yatabe-Kakugawa Y Zhang S 2012 ) Molecular evidence on the origin of Osmunda × mildei (Osmundaceae).
American Fern Journal , 102 , 55 -68 .
[本文引用: 1]
[35]
Vilà M Weber E Antonio CM 2000 ) Conservation implications of invasion by plant hybridization.
Biological Invasions , 2 , 207 -217 .
[本文引用: 2]
[36]
Wang XR Szmidt AE Savolainen O 2001 ) Genetic composition and diploid hybrid speciation of a high mountain pine, Pinus densata, native to the Tibetan Plateau.
Genetics , 159 , 337 -346 .
[本文引用: 1]
[37]
Wang Y Shang H Gu YF Wei HJ Zhao GH Dai XL Yan YH 2015 a) A new cryptic hybrid species of Adiantum L. (Pteridaceae) identified by nuclear and chloroplast DNA sequences.
Chinese Science Bulletin , 60 , 922 -932 .
[本文引用: 1]
[38]
Wang Y Shang H Zhou XL Zhao GH Dai XL Yan YH 2015 b) Adiantum× ailaoshanense (Pteridaceae), a new natural hybrid from Yunnan, China.
Phytotaxa , 236 , 266 -272 .
[本文引用: 1]
[39]
Whitham TG Martinsen GD Keim P Floate KD Dungey HS Potts BM 1999 ) Plant hybrid zones affect biodiversity: tools for a genetic-based understanding of community structure.
Ecology , 80 , 416 -428 .
[本文引用: 1]
[40]
Wu W 2009 ) Natural Hybridization, Phylogeography and Speciation Patterns of Altingiaceae
. PhD dissertaion, Sun Yat-sen University, Guangzhou. (in Chinese with English abstract)
[本文引用: 1]
[吴伟 (2009 ) 阿丁枫科的自然杂交、亲缘地理学与物种形成模式
. 中山大学博士学位论文, 广州 .]
[本文引用: 1]
[41]
Wu W Zhou R Huang Y Boufford DE Shi S 2010 ) Molecular evidence for natural intergeneric hybridization between Liquidambar and Altingia.
Journal of Plant Research , 123 , 231 -239 .
[本文引用: 1]
[42]
Wyk AM Dalton DL Hoban S Bruford MW Russo IRM Birss C Grobler P Vuuren BJ Kotzé A 2017 ) Quantitative evaluation of hybridization and the impact on biodiversity conservation.
Ecology and Evolution , 7 , 320 -330 .
[本文引用: 1]
[43]
Yakimowski SB Rieseberg LH 2014 ) The role of homoploid hybridization in evolution: a century of studies synthesizing genetics and ecology.
American Journal of Botany , 101 , 1247 -1258 .
[本文引用: 1]
[44]
Zhang JL 2007 ) Natural Hybridization Origin of Rhododendron agastum (Ericaceae) in Yunnan, China
. PhD dissertaion, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming . (in Chinese with English abstract)
[本文引用: 1]
[张敬丽 (2007 ) 杜鹃花属迷人杜鹃的自然杂交起源研究
. 中国科学院昆明植物研究所博士学位论文, 昆明 .]
[本文引用: 1]
[45]
Zhou QJ Cai YC Ng WL Wu W Dai SP Wang F Zhou RC 2017 ) Molecular evidence for natural hybridization between two Melastoma species endemic to Hainan and their widespread congeners.
Biodiversity Science , 25 , 638 -646 . (in Chinese with English abstract)
[本文引用: 1]
[周秋杰 , 蔡亚城 , 黄伟伦 , 吴伟 , 代色平 , 王峰 , 周仁超 (2017 ) 野牡丹属两个海南特有种与同属广布种自然杂交的分子证据
. 生物多样性 , 25 , 638 -646 .]
[本文引用: 1]
Hybridization and speciation.
1
2013
... 首先, 大量现存物种是通过自然杂交起源的, 或者处于自然杂交物种形成的演化途中(Abbott et al, 2013 ), 网状进化是一种重要的物种演化模式.一方面, 杂交后代可以通过染色体加倍形成异源多倍化物种(Soltis et al, 2014 ); 另一方面, 由于同倍性的杂交后代与父母本间的生殖隔离不如异倍杂交那样容易产生, 所以同倍性杂交物种比较少见, 但仍然是不可忽视的物种形成方式之一.例如, 生长在粘性土壤中的向日葵(Helianthus annuus )和其生长在砂质土壤中的近亲植物草原太阳花(H. petiolaris ), 产生了3种适应不同环境的自然杂交种(H. anomalus、H. deserticola 和H. paradoxus ) (Rieseberg, 1991 ). ...
The problems with hybrids: setting conservation guidelines.
1
2001
... 自然杂交指的是已发生遗传分化的种群之间的交配(Arnold, 1992 ; Allendorf et al, 2001 ), 一般更容易发生在亲缘关系较近的类群间.然而, 蕨类植物的一个案例表明, 分化六千万年的物种之间也可以发生杂交(Rothfels et al, 2015 ).大部分自然杂交后代(hybrid)由于生殖隔离造成的不育而被淘汰, 或存在于杂交带中成为亲本间基因流的桥梁; 但仍有一部分杂交后代通过各种机制得以留存而形成自然杂交种(hybrid species), 如同倍杂交后代由于适应新的环境与父母本产生生殖隔离, 或通过异源染色体加倍的方式直接和父母本产生生殖隔离(生物种概念)(Mallet, 2007 ); 而一些奇数倍染色体的杂交后代通过无性繁殖得以延续, 由于其形态的可辨识性, 也可能被命名为杂交种(形态种). ...
Hybridization as an evolutionary stimulus.
1
1954
... 随着自然杂交研究的增多, 人们越来越认识到物种杂交是自然界的普遍现象, 而且是植物物种形成的主要驱动力.杂交有害的情况更多地发生在同倍体杂交中, 而多倍化杂交物种往往比其亲本更利于生态位建成和适应性增强.由此带来的混合基因库具备更高的遗传多样性和快速适应新环境的潜力, 对生物多样性的保持具有一定的积极作用.所以, 对其保护价值的讨论具有重要意义(Anderson & Stebbins Jr, 1954 ; Nolte & Tautz, 2010 ; Piett et al, 2015 ; Kasari et al, 2016 ). ...
Natural hybridization as an evolutionary process.
1
1992
... 自然杂交指的是已发生遗传分化的种群之间的交配(Arnold, 1992 ; Allendorf et al, 2001 ), 一般更容易发生在亲缘关系较近的类群间.然而, 蕨类植物的一个案例表明, 分化六千万年的物种之间也可以发生杂交(Rothfels et al, 2015 ).大部分自然杂交后代(hybrid)由于生殖隔离造成的不育而被淘汰, 或存在于杂交带中成为亲本间基因流的桥梁; 但仍有一部分杂交后代通过各种机制得以留存而形成自然杂交种(hybrid species), 如同倍杂交后代由于适应新的环境与父母本产生生殖隔离, 或通过异源染色体加倍的方式直接和父母本产生生殖隔离(生物种概念)(Mallet, 2007 ); 而一些奇数倍染色体的杂交后代通过无性繁殖得以延续, 由于其形态的可辨识性, 也可能被命名为杂交种(形态种). ...
Strategies to address the conservation threats posed by hybridization and genetic introgression.
1
2016
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
Speciation
1
2004
... 其次, 在同域物种形成的过程中, 当新物种和旧物种的生殖隔离尚未完全建立时, 杂交渗透所形成的物种间基因流可能会导致适应不同生态位的基因被选择作用固定在不同的物种中, 促进物种的分化(Coyne & Orr, 2004 ; Rolán-Alvarez, 2007 ). ...
Monachosorum arakii Tagawa (Dennstaedtiaceae) is a relict “international” hybrid: a reassessment of the Monachosorum species.
1
2016
... (2)当父母本一方或双方濒危, 但杂交后代或杂交种没有威胁到其父母本时, 自然杂交应该被保护.日本西部特有分布的稀子蕨属植物Monachosorum arakii 是亚洲大陆分布的稀子蕨(M. henryi )和日本分布的日本稀子蕨(M. nipponicum )的自然杂交产物, 该杂交种可以通过无性繁殖芽孢自行在野外建立种群.由于稀子蕨现仅分布于中国大陆以及热带亚洲地区, 在日本已经灭绝, 稀子蕨和日本稀子蕨的杂交可能是过去地质时代的孑遗产物(Ebihara et al, 2016 ).在此, 由于杂交没有威胁到父母本的生存, 自然杂交种Monachosorum arakii 在日本的分布具有重要科学研究价值并可维系已灭绝的亲本遗传信息, 因此该自然杂交种应该受到必要的保护. ...
Hybrid vigor between native and introduced salamanders raises new challenges for conservation
1
2007
... 杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 ).实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 ).如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 ).此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 ).总之, 自然杂交对维持生态系统的稳定性起到很重要的作用. ...
Contrasting hybridization rates between sympatric three-spined sticklebacks highlight the fragility of reproductive barriers between evolutionarily young species.
1
2006
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
Evolutionary biology: speciation undone.
1
2014
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
The ecological genetics of homoploid hybrid speciation.
1
2005
... 杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 ).实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 ).如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 ).此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 ).总之, 自然杂交对维持生态系统的稳定性起到很重要的作用. ...
Unreduced 3x gamete formation of allotriploid hybrid derived from the cross of Primula denticulata (4x) × P. rosea (2x) as a causal factor for producing pentaploid hybrids in the backcross with pollen of tetraploid P. denticulata.
1
2009
... (1)自然杂交指的是近期或者历史上自然发生的没有受到人为因素干扰的物种杂交事件, 包括自然杂交种、基因渐渗和杂交带等.这里需要指出的是, 用于可操作的自然保护的自然杂交对象一般是指具有相对稳定的遗传特征的杂交事件, 可以是能够自我繁殖的自然杂交种, 如孟连铁线蕨(Adiantum menglianense )与假鞭叶铁线蕨(A. malesianum )和苍山铁线蕨(A. sinense )分别产生的自然杂交后代: 梅山铁线蕨(A. meishanianum )和哀牢山铁线蕨(A. ailaoshanense ), 均可以通过无性繁殖在自然界自我维系自然种群(Wang et al, 2015b ; Shang et al, 2016 ); 也可以是由于杂交渐渗在自然杂交带中产生的杂交群体, 如分布于丽江玉龙雪山的橘红灯台报春(Primula bulleyana )和霞红灯台报春(P. beesiana )可以在其分布重叠区内高频率发生自然杂交并形成杂交后代.虽然其杂交后代不育, 但杂交后代通过回交可能对亲本的遗传多样性产生重要的影响(Hayashi et al, 2009 ).因此可以视其濒危程度及其亲本的生存状况等因素, 酌情考虑保护措施.但是, 对自然界中偶尔产生的一些自然杂交后代, 由于遗传上的不稳定性和发生频率的随机性, 一般不需要列入保护对象, 如粤紫萁(Osmunda mildei )是华南紫萁(O. vachellii )和紫萁(O. japonica )的自然杂交后代(Tsutsumi et al, 2012 ), 其杂交后代不具可育性, 无法在野外独立建立种群, 而是仅在亲本的重叠分布区产生极少量的随机个体, 因此也很难对该自然杂交个体进行有效的保护. ...
The endangered species act: what do we mean by species.
1
1993
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
Reticulate evolution in the apogamous Dryopteris varia complex (Dryopteridaceae, subg. Erythrovariae, sect. Variae) and its related sexual species in Japan.
2014
The IUCN Red List of Threatened Species
1
2013
... Jackiw等(2015)对北美的物种保护条例进行了研究, 发现虽然人们已经开始关注到自然杂交的保护, 但是条例中涉及的条款很少, 并且往往语焉不详, 执行情况也不理想.世界自然保护联盟(IUCN)的物种红色名录里, 甚至明确说明要把自然杂交种剔除, 完全忽略其价值(IUCN, 2013 , 2014 ).为此, Jackiw等(2015)针对北美的情况制定了一个评估杂交是否值得保护的原则框架, 他建议保育工作者在制定法规和具体的保护工作中, 系统地考虑杂交的成因、对父母本的威胁程度, 以及对生态环境的影响等.这些评估原则对中国的保护工作也有很好的借鉴意义. ...
Guidelines for Using the IUCN Red List Categories and Criteria, Version 11
1
2014
... Jackiw等(2015)对北美的物种保护条例进行了研究, 发现虽然人们已经开始关注到自然杂交的保护, 但是条例中涉及的条款很少, 并且往往语焉不详, 执行情况也不理想.世界自然保护联盟(IUCN)的物种红色名录里, 甚至明确说明要把自然杂交种剔除, 完全忽略其价值(IUCN, 2013 , 2014 ).为此, Jackiw等(2015)针对北美的情况制定了一个评估杂交是否值得保护的原则框架, 他建议保育工作者在制定法规和具体的保护工作中, 系统地考虑杂交的成因、对父母本的威胁程度, 以及对生态环境的影响等.这些评估原则对中国的保护工作也有很好的借鉴意义. ...
A framework to guide the conservation of species hybrids based on ethical and ecological considerations.
1
2015
... 本文结合近些年有关自然杂交保护生物学的研究进展, 特别是Jackiw等(2015) 提出的自然杂交保护的策略框架, 拟探讨: (1)保护自然杂交的意义; (2)结合中国现在的自然杂交保护现状, 我们应该如何保护自然杂交? ...
Hybrid ecosystems can contribute to local biodiversity conservation.
1
2016
... 随着自然杂交研究的增多, 人们越来越认识到物种杂交是自然界的普遍现象, 而且是植物物种形成的主要驱动力.杂交有害的情况更多地发生在同倍体杂交中, 而多倍化杂交物种往往比其亲本更利于生态位建成和适应性增强.由此带来的混合基因库具备更高的遗传多样性和快速适应新环境的潜力, 对生物多样性的保持具有一定的积极作用.所以, 对其保护价值的讨论具有重要意义(Anderson & Stebbins Jr, 1954 ; Nolte & Tautz, 2010 ; Piett et al, 2015 ; Kasari et al, 2016 ). ...
Species collapse via hybridization in Darwin’s tree finches.
1
2014
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
Hybrid speciation.
1
2007
... 自然杂交指的是已发生遗传分化的种群之间的交配(Arnold, 1992 ; Allendorf et al, 2001 ), 一般更容易发生在亲缘关系较近的类群间.然而, 蕨类植物的一个案例表明, 分化六千万年的物种之间也可以发生杂交(Rothfels et al, 2015 ).大部分自然杂交后代(hybrid)由于生殖隔离造成的不育而被淘汰, 或存在于杂交带中成为亲本间基因流的桥梁; 但仍有一部分杂交后代通过各种机制得以留存而形成自然杂交种(hybrid species), 如同倍杂交后代由于适应新的环境与父母本产生生殖隔离, 或通过异源染色体加倍的方式直接和父母本产生生殖隔离(生物种概念)(Mallet, 2007 ); 而一些奇数倍染色体的杂交后代通过无性繁殖得以延续, 由于其形态的可辨识性, 也可能被命名为杂交种(形态种). ...
Theoretical Ecology
1
1976
... 一般认为, 生态系统组成的多样性与稳定性存在某种程度的相关性, 虽然May (1976) 从数学上论证了多样性有时会导致稳定性下降的可能性, 但在多数情况下, 通过增加生态系统内的多样性, 可促进系统的稳定性.自然杂交在生态系统中的存在, 则可能通过影响遗传多样性和物种多样性进而影响生态系统的稳定性和可恢复性. ...
Understanding the onset of hybrid speciation.
1
2010
... 随着自然杂交研究的增多, 人们越来越认识到物种杂交是自然界的普遍现象, 而且是植物物种形成的主要驱动力.杂交有害的情况更多地发生在同倍体杂交中, 而多倍化杂交物种往往比其亲本更利于生态位建成和适应性增强.由此带来的混合基因库具备更高的遗传多样性和快速适应新环境的潜力, 对生物多样性的保持具有一定的积极作用.所以, 对其保护价值的讨论具有重要意义(Anderson & Stebbins Jr, 1954 ; Nolte & Tautz, 2010 ; Piett et al, 2015 ; Kasari et al, 2016 ). ...
Characteristics for evaluating the conservation value of species hybrids.
1
2015
... 随着自然杂交研究的增多, 人们越来越认识到物种杂交是自然界的普遍现象, 而且是植物物种形成的主要驱动力.杂交有害的情况更多地发生在同倍体杂交中, 而多倍化杂交物种往往比其亲本更利于生态位建成和适应性增强.由此带来的混合基因库具备更高的遗传多样性和快速适应新环境的潜力, 对生物多样性的保持具有一定的积极作用.所以, 对其保护价值的讨论具有重要意义(Anderson & Stebbins Jr, 1954 ; Nolte & Tautz, 2010 ; Piett et al, 2015 ; Kasari et al, 2016 ). ...
Hybridisation on coral reefs and the conservation of evolutionary novelty.
1
2015
... 保护生物学家在自然杂交的保护领域已经开始了理论探讨和实践尝试.除Jackiw等(2015)的工作以外, Richards和Hobbs (2015) 针对珊瑚礁群落中的杂交保护问题进行了讨论, 首先综述了珊瑚虫以及岩礁鱼类杂交研究进展, 并据此评估了杂交对珊瑚礁群落多样性的影响.分析结果表明, 杂交可以造成多种进化生物学后果, 既有可能是增加遗传多样性, 产生新的类群, 也有可能造成物种逆转.由于对自然杂交保护立法的忽视, 可能使得有价值的自然杂交没有得到有效的保护而导致其灭绝.针对这种情况, 他们提出, 应逐个案例(case-by-case)评估杂交的原因和后果, 有针对性地决定是保护还是清除杂交. ...
Homoploid reticulate evolution in Helianthus (Asteraceae): evidence from ribosomal genes.
1
1991
... 首先, 大量现存物种是通过自然杂交起源的, 或者处于自然杂交物种形成的演化途中(Abbott et al, 2013 ), 网状进化是一种重要的物种演化模式.一方面, 杂交后代可以通过染色体加倍形成异源多倍化物种(Soltis et al, 2014 ); 另一方面, 由于同倍性的杂交后代与父母本间的生殖隔离不如异倍杂交那样容易产生, 所以同倍性杂交物种比较少见, 但仍然是不可忽视的物种形成方式之一.例如, 生长在粘性土壤中的向日葵(Helianthus annuus )和其生长在砂质土壤中的近亲植物草原太阳花(H. petiolaris ), 产生了3种适应不同环境的自然杂交种(H. anomalus、H. deserticola 和H. paradoxus ) (Rieseberg, 1991 ). ...
Sympatric speciation as a by-product of ecological adaptation in the Galician Littorina saxatilis hybrid zone.
1
2007
... 其次, 在同域物种形成的过程中, 当新物种和旧物种的生殖隔离尚未完全建立时, 杂交渗透所形成的物种间基因流可能会导致适应不同生态位的基因被选择作用固定在不同的物种中, 促进物种的分化(Coyne & Orr, 2004 ; Rolán-Alvarez, 2007 ). ...
Natural hybridization between genera that diverged from each other approximately 60 million years ago.
1
2015
... 自然杂交指的是已发生遗传分化的种群之间的交配(Arnold, 1992 ; Allendorf et al, 2001 ), 一般更容易发生在亲缘关系较近的类群间.然而, 蕨类植物的一个案例表明, 分化六千万年的物种之间也可以发生杂交(Rothfels et al, 2015 ).大部分自然杂交后代(hybrid)由于生殖隔离造成的不育而被淘汰, 或存在于杂交带中成为亲本间基因流的桥梁; 但仍有一部分杂交后代通过各种机制得以留存而形成自然杂交种(hybrid species), 如同倍杂交后代由于适应新的环境与父母本产生生殖隔离, 或通过异源染色体加倍的方式直接和父母本产生生殖隔离(生物种概念)(Mallet, 2007 ); 而一些奇数倍染色体的杂交后代通过无性繁殖得以延续, 由于其形态的可辨识性, 也可能被命名为杂交种(形态种). ...
Hybridization and adaptive radiation.
1
2004
... 自然杂交不仅可以增加遗传多样性和物种的适应性, 而且可以导致物种的辐射式演化, 促进大量物种在相对较短的时期内形成(Seehausen, 2004 ). ...
Conservation: losing biodiversity by reverse speciation.
1
2006
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
Likely allopatric origins of Adiantum× meishanianum (Pteridaceae) through multiple hybridizations.
1
2016
... (1)自然杂交指的是近期或者历史上自然发生的没有受到人为因素干扰的物种杂交事件, 包括自然杂交种、基因渐渗和杂交带等.这里需要指出的是, 用于可操作的自然保护的自然杂交对象一般是指具有相对稳定的遗传特征的杂交事件, 可以是能够自我繁殖的自然杂交种, 如孟连铁线蕨(Adiantum menglianense )与假鞭叶铁线蕨(A. malesianum )和苍山铁线蕨(A. sinense )分别产生的自然杂交后代: 梅山铁线蕨(A. meishanianum )和哀牢山铁线蕨(A. ailaoshanense ), 均可以通过无性繁殖在自然界自我维系自然种群(Wang et al, 2015b ; Shang et al, 2016 ); 也可以是由于杂交渐渗在自然杂交带中产生的杂交群体, 如分布于丽江玉龙雪山的橘红灯台报春(Primula bulleyana )和霞红灯台报春(P. beesiana )可以在其分布重叠区内高频率发生自然杂交并形成杂交后代.虽然其杂交后代不育, 但杂交后代通过回交可能对亲本的遗传多样性产生重要的影响(Hayashi et al, 2009 ).因此可以视其濒危程度及其亲本的生存状况等因素, 酌情考虑保护措施.但是, 对自然界中偶尔产生的一些自然杂交后代, 由于遗传上的不稳定性和发生频率的随机性, 一般不需要列入保护对象, 如粤紫萁(Osmunda mildei )是华南紫萁(O. vachellii )和紫萁(O. japonica )的自然杂交后代(Tsutsumi et al, 2012 ), 其杂交后代不具可育性, 无法在野外独立建立种群, 而是仅在亲本的重叠分布区产生极少量的随机个体, 因此也很难对该自然杂交个体进行有效的保护. ...
The polyploidy revolution then and now: Stebbins revisited.
1
2014
... 首先, 大量现存物种是通过自然杂交起源的, 或者处于自然杂交物种形成的演化途中(Abbott et al, 2013 ), 网状进化是一种重要的物种演化模式.一方面, 杂交后代可以通过染色体加倍形成异源多倍化物种(Soltis et al, 2014 ); 另一方面, 由于同倍性的杂交后代与父母本间的生殖隔离不如异倍杂交那样容易产生, 所以同倍性杂交物种比较少见, 但仍然是不可忽视的物种形成方式之一.例如, 生长在粘性土壤中的向日葵(Helianthus annuus )和其生长在砂质土壤中的近亲植物草原太阳花(H. petiolaris ), 产生了3种适应不同环境的自然杂交种(H. anomalus、H. deserticola 和H. paradoxus ) (Rieseberg, 1991 ). ...
Hybridization facilitates evolutionary rescue.
1
2014
... 杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 ).实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 ).如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 ).此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 ).总之, 自然杂交对维持生态系统的稳定性起到很重要的作用. ...
Speciation in reverse: morphological and genetic evidence of the collapse of a three- spined stickleback (Gasterosteus aculeatus) species pair.
1
2006
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
Molecular evidence on the origin of Osmunda × mildei (Osmundaceae).
1
2012
... (1)自然杂交指的是近期或者历史上自然发生的没有受到人为因素干扰的物种杂交事件, 包括自然杂交种、基因渐渗和杂交带等.这里需要指出的是, 用于可操作的自然保护的自然杂交对象一般是指具有相对稳定的遗传特征的杂交事件, 可以是能够自我繁殖的自然杂交种, 如孟连铁线蕨(Adiantum menglianense )与假鞭叶铁线蕨(A. malesianum )和苍山铁线蕨(A. sinense )分别产生的自然杂交后代: 梅山铁线蕨(A. meishanianum )和哀牢山铁线蕨(A. ailaoshanense ), 均可以通过无性繁殖在自然界自我维系自然种群(Wang et al, 2015b ; Shang et al, 2016 ); 也可以是由于杂交渐渗在自然杂交带中产生的杂交群体, 如分布于丽江玉龙雪山的橘红灯台报春(Primula bulleyana )和霞红灯台报春(P. beesiana )可以在其分布重叠区内高频率发生自然杂交并形成杂交后代.虽然其杂交后代不育, 但杂交后代通过回交可能对亲本的遗传多样性产生重要的影响(Hayashi et al, 2009 ).因此可以视其濒危程度及其亲本的生存状况等因素, 酌情考虑保护措施.但是, 对自然界中偶尔产生的一些自然杂交后代, 由于遗传上的不稳定性和发生频率的随机性, 一般不需要列入保护对象, 如粤紫萁(Osmunda mildei )是华南紫萁(O. vachellii )和紫萁(O. japonica )的自然杂交后代(Tsutsumi et al, 2012 ), 其杂交后代不具可育性, 无法在野外独立建立种群, 而是仅在亲本的重叠分布区产生极少量的随机个体, 因此也很难对该自然杂交个体进行有效的保护. ...
Conservation implications of invasion by plant hybridization.
2
2000
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
... 杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 ).实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 ).如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 ).此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 ).总之, 自然杂交对维持生态系统的稳定性起到很重要的作用. ...
Genetic composition and diploid hybrid speciation of a high mountain pine, Pinus densata, native to the Tibetan Plateau.
1
2001
... (3)当父母本双方都不濒危, 并且杂交后代或杂交种没有威胁到其父母本时, 自然杂交应该被保护.高山松(Pinus densata )是油松(P. tabuliformis )和云南松(P. yunnanensis )的自然杂交种(Wang et al, 2001 ), 是乔木物种二倍体自然杂交同倍体成种的范例, 广泛分布于云南、四川、西藏、青海等地, 生长于海拔1,500-4,500 m的地区.但其父母本均不濒危, 且高山松没有威胁到父母本的生存; 由于高山松具有较大的社会经济价值, 因此可以列入林业系统的珍稀林木保护名录; 然而, 由于高山松分布广泛, 其野外种群没有受到明显的威胁, 目前尚不需要将该种列入物种红色名录. ...
A new cryptic hybrid species of Adiantum L. (Pteridaceae) identified by nuclear and chloroplast DNA sequences.
1
2015
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
Adiantum× ailaoshanense (Pteridaceae), a new natural hybrid from Yunnan, China.
1
2015
... (1)自然杂交指的是近期或者历史上自然发生的没有受到人为因素干扰的物种杂交事件, 包括自然杂交种、基因渐渗和杂交带等.这里需要指出的是, 用于可操作的自然保护的自然杂交对象一般是指具有相对稳定的遗传特征的杂交事件, 可以是能够自我繁殖的自然杂交种, 如孟连铁线蕨(Adiantum menglianense )与假鞭叶铁线蕨(A. malesianum )和苍山铁线蕨(A. sinense )分别产生的自然杂交后代: 梅山铁线蕨(A. meishanianum )和哀牢山铁线蕨(A. ailaoshanense ), 均可以通过无性繁殖在自然界自我维系自然种群(Wang et al, 2015b ; Shang et al, 2016 ); 也可以是由于杂交渐渗在自然杂交带中产生的杂交群体, 如分布于丽江玉龙雪山的橘红灯台报春(Primula bulleyana )和霞红灯台报春(P. beesiana )可以在其分布重叠区内高频率发生自然杂交并形成杂交后代.虽然其杂交后代不育, 但杂交后代通过回交可能对亲本的遗传多样性产生重要的影响(Hayashi et al, 2009 ).因此可以视其濒危程度及其亲本的生存状况等因素, 酌情考虑保护措施.但是, 对自然界中偶尔产生的一些自然杂交后代, 由于遗传上的不稳定性和发生频率的随机性, 一般不需要列入保护对象, 如粤紫萁(Osmunda mildei )是华南紫萁(O. vachellii )和紫萁(O. japonica )的自然杂交后代(Tsutsumi et al, 2012 ), 其杂交后代不具可育性, 无法在野外独立建立种群, 而是仅在亲本的重叠分布区产生极少量的随机个体, 因此也很难对该自然杂交个体进行有效的保护. ...
Plant hybrid zones affect biodiversity: tools for a genetic-based understanding of community structure.
1
1999
... 杂交可增加物种对植食作用、病原体和寄生虫压力的抗性(Vilà et al, 2000 ), 在应对气候剧变、虫害爆发、病原菌等环境压力时, 可能会表现更出色(Fitzpatrick & Shaffer, 2007 ).实验表明, 杂交的酵母属(Saccharomyce )种群在逆境下表现得比其父母本更加出色, 当环境剧变时, 通过基因交换, 可以使物种快速获得适应新环境的能力而得以存活(Stelkens et al, 2014 ).如果自然杂交后代适应了极端环境或新的栖息地, 可能会增加生态系统对环境压力的恢复能力(Gross & Rieseberg, 2005 ).此外, 杂交可能改变现有的群落结构, 整合分析表明, 植物杂交带使栖息环境更多样化, 促进了昆虫和真菌, 甚至鸟类的物种多样性(Whitham et al, 1999 ).总之, 自然杂交对维持生态系统的稳定性起到很重要的作用. ...
阿丁枫科的自然杂交、亲缘地理学与物种形成模式
1
2009
... 在中国, 自然杂交的保护问题还没有引起人们的足够关注, 1996年颁布的《野生植物保护条例》和2016年颁布的《野生动物保护法》都没有明确提及自然杂交种是否需要保护的问题.但在已有的保护名录里实际上已经包括了自然杂交种, 例如《国家重点保护野生植物名录》(1999)中的国家二级保护植物半枫荷(Semiliquidambar cathayensis )是杂交起源(吴伟, 2009 ; Wu et al, 2010 ).相信随着保护研究的进展, 类似的例子以及相关的讨论会越来越多, 而这也是保护生物学无法回避的问题. ...
阿丁枫科的自然杂交、亲缘地理学与物种形成模式
1
2009
... 在中国, 自然杂交的保护问题还没有引起人们的足够关注, 1996年颁布的《野生植物保护条例》和2016年颁布的《野生动物保护法》都没有明确提及自然杂交种是否需要保护的问题.但在已有的保护名录里实际上已经包括了自然杂交种, 例如《国家重点保护野生植物名录》(1999)中的国家二级保护植物半枫荷(Semiliquidambar cathayensis )是杂交起源(吴伟, 2009 ; Wu et al, 2010 ).相信随着保护研究的进展, 类似的例子以及相关的讨论会越来越多, 而这也是保护生物学无法回避的问题. ...
Molecular evidence for natural intergeneric hybridization between Liquidambar and Altingia.
1
2010
... 在中国, 自然杂交的保护问题还没有引起人们的足够关注, 1996年颁布的《野生植物保护条例》和2016年颁布的《野生动物保护法》都没有明确提及自然杂交种是否需要保护的问题.但在已有的保护名录里实际上已经包括了自然杂交种, 例如《国家重点保护野生植物名录》(1999)中的国家二级保护植物半枫荷(Semiliquidambar cathayensis )是杂交起源(吴伟, 2009 ; Wu et al, 2010 ).相信随着保护研究的进展, 类似的例子以及相关的讨论会越来越多, 而这也是保护生物学无法回避的问题. ...
Quantitative evaluation of hybridization and the impact on biodiversity conservation.
1
2017
... 自然杂交包括杂交过程和杂交产物(未成种的杂交后代和自然杂交种), 它们是否需要保护一直备受争议.传统观点认为杂交会对纯种的父母本造成威胁, 特别是在均质化环境中过度的杂交渐渗会造成两物种融合, 即发生物种逆转(reverse speciation), 从而导致物种多样性的丢失(Seehausen, 2006 ; Grant & Grant, 2014 ), 所以主张杂交有害不予保护(Hill, 1993 ; Vilà et al, 2000 ), 应采取积极措施防范其威胁(Bohling, 2016 ), 甚至应通过分子标记准确地鉴别后人为清除(Wyk et al, 2017 ); 而没有形成自然杂交种的个别杂交后代则迟早被自然所淘汰, 更无须作为保护对象.另外, 一些隐性杂交种往往难以通过形态鉴定, 或者无法确定其起源, 因此容易被忽视(Wang et al, 2015a ). ...
The role of homoploid hybridization in evolution: a century of studies synthesizing genetics and ecology.
1
2014
... 不完全生殖隔离导致的杂交渐渗现象中, 物种间的基因流对物种演化和生态适应起到非常重要的作用.由于父母本双方基因库的混合而产生新的等位基因组合, 杂交带往往比父母本种群具有更高的遗传多样性以及适应性(Yakimowski & Rieseberg, 2014 ).当渗透的等位基因受到选择时, 适度的渗透可以使物种快速获得适应性.这种基因交流可以给父母本物种提供一个快速适应新环境、提高适合度的机会.从这个角度来说, 适度的自然杂交显然是有利的.只有当渗透过度时, 才可能造成生殖隔离的突破而造成物种逆转. ...
杜鹃花属迷人杜鹃的自然杂交起源研究
1
2007
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
杜鹃花属迷人杜鹃的自然杂交起源研究
1
2007
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
野牡丹属两个海南特有种与同属广布种自然杂交的分子证据
1
2017
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...
野牡丹属两个海南特有种与同属广布种自然杂交的分子证据
1
2017
... (4)无论父母本双方濒危或不濒危, 当杂交后代威胁到其父母本时, 自然杂交是否被保护就需要视具体情况确定.如果杂交后代或杂交种比其父母本具有更高的适合度、具有更好的环境可塑性、有助于群落结构稳定、增加遗传多样性或促进成种进程(hybrid speciation)而不是导致物种逆转时, 应考虑予以保护.如迷人杜鹃(Rododendron agastum )和露珠杜鹃(R. irroratum )均为马缨花(R. delavayi )和大白花杜鹃(R. decorum )的自然杂交种, 且两个自然杂交种的分布区和马缨花的分布区一样广泛.大白花杜鹃分布在低海拔地区, 马缨花的分布区则高中低海拔均有分布, 而自然杂交种迷人杜鹃和露珠杜鹃分布在高海拔地区, 核基因序列分析显示杂交后代与亲本之间具有广泛的基因交流和回交历史(张敬丽, 2007 ), 但杂交后代对父母本的生存没有明显的威胁.鉴于全球变化对高山植物的威胁、近年来中国高山生态环境的破坏, 以及该种重要的观赏价值, 该自然杂交种应该得到保护.相反, 在本专辑中另一篇论文(周杰秋等, 2017 )中, 枝毛野牡丹(Melastoma dendrisetosum )和毛菍(M. sanguineum )的自然杂交后代同样出现了威胁亲本之一海南特有濒危物种枝毛野牡丹生存的问题, 对这样的自然杂交种则不应该保护.厄瓜多尔的科隆群岛中弗雷里安纳岛上原本分布着大树雀(Camarhynchus psittacula )、中树雀(C. pauper )和小树雀(C. parvulus ) 3种树雀.由于种间杂交, 大树雀已经在该岛消失, 剩余2个种也面临着融合的危险(Kleindorfer et al, 2014 ).同样的情况已经发生在加拿大温哥华岛恩诺斯湖, 一个三刺鱼复合群(Gasterosteus aculeatus complex)的种对原本包含了区别显著的2个物种, 但经过几十年的杂交渗透(1977-2002年), 两个物种迅速混合为一个杂种群(hybrid swarm) (Gow et al, 2006 ; Taylor et al, 2006 ).对这类由于杂交渗透造成的物种逆转的情况, 也不应予以保护. ...



{kind=link}
{kind=link}