1 State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093 2 University of the Chinese Academy of Sciences, Beijing 1000491 State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093 2 University of the Chinese Academy of Sciences, Beijing 100049
With the rise of plant molecular systematics, tremendous progress has been made in understanding phylogenetic relationships within angiosperms. With the basic phylogenetic framework of angiosperms established, a DNA phylogeny-based angiosperm classification system at the order and familial levels was proposed by the Angiosperm Phylogeny Group (APG) in 1998 and has been updated three times. In this paper, we summarize the major achievements of the APG system as follows: (1) testing the repeatability and predictability of the APG system for angiosperms; (2) resolving the systematic positions of some segregate taxa which were not placed based on morphological characters; (3) proving that it is not reasonable to first divide angiosperms based on cotyledon character; (4) demonstrating the importance of tricolpate/tricolporate pollen and derivatives for angiosperm classification; (5) finding that the centrifugal development of stamens in polyandrous groups have evolved independently many times and should not be used to delimit class or subclass of angiosperms; (6) supporting that most of the families delimited by broad morphological characters are natural; and (7) separating some families which are traditionally regarded as natural. We then point out potential problems that need to be resolved in the future, including: (1) how to harmonize the APG system and the morphology-based systems; (2) establishing new morphological evolution theories on the basis of the APG system; (3) determining whether it is enough to only use “monophyly” as a criterion to circumscribe orders and families; (4) determining morphological synapormorphies of those orders in the APG system; and (5) how to best compile a key to distinguish the orders and families of the APG system and to list their diagnostic characters for orders and families. In addition, we propose suggestions for the phylogenetic relationships and taxonomic status of some taxa mainly distributed in Asia, specifically East Asia, including Illiciaceae, Acanthochlamydaceae, Tetracentraceae, Leeaceae, Rhoipteiaceae, Hippocastenaceae, Aceraceae, Bretschneideraceae as familial status, and dividing Cornaceae sensu lato into Cornaceae sensu stricto and Nyssaceae sensu lato.
WeiWang, XiaoxiaZhang, ZhiduanChen, AnmingLu. Comments on the APG’s classification of angiosperms[J]. Biodiversity Science, 2017, 25(4): 418-426 https://doi.org/10.17520/biods.2017015
随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics)。在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993)。这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义。从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟。在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统。被子植物由此成为了第一个基于分子数据建立分类系统的大类群。之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016)。该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统。可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009)。但并不是说APG系统已经完美, 不存在需要继续研究的问题了。经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题。
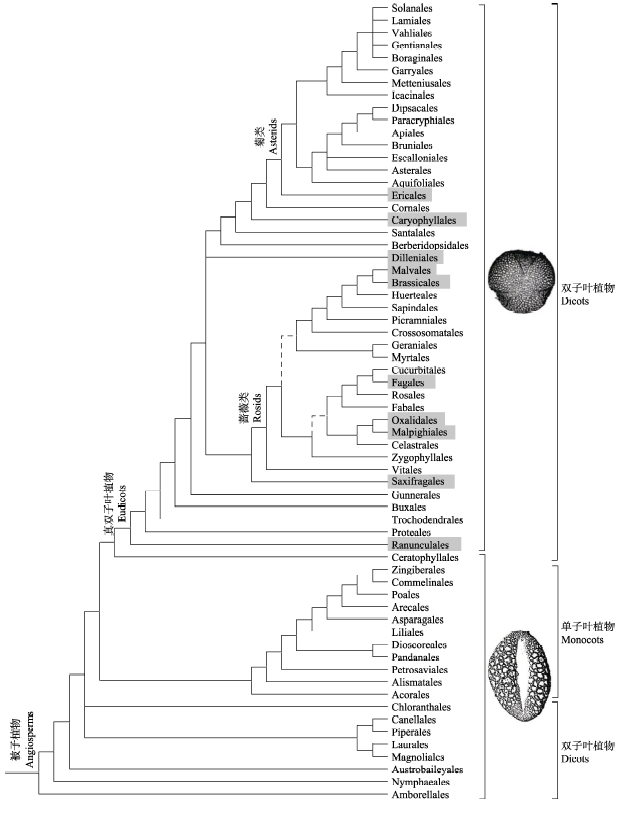
图1 被子植物APG IV (2016)系统的目间系统关系。虚线表示核/线粒体树与叶绿体树冲突; 标灰色的目含有多雄蕊离心发育的类群; 花粉和子叶性状标在系统树的右边。
Fig. 1 Interrelationships of the APG IV (2016) orders of angiosperms. The dotted lines indicate the conflicting placements between nuclear/mitochondrial and chloroplast trees. The orders with the gray contain at least one taxon with multiple centrifugal development stamens. Pollen and cotyledonal characters are labeled on the right.
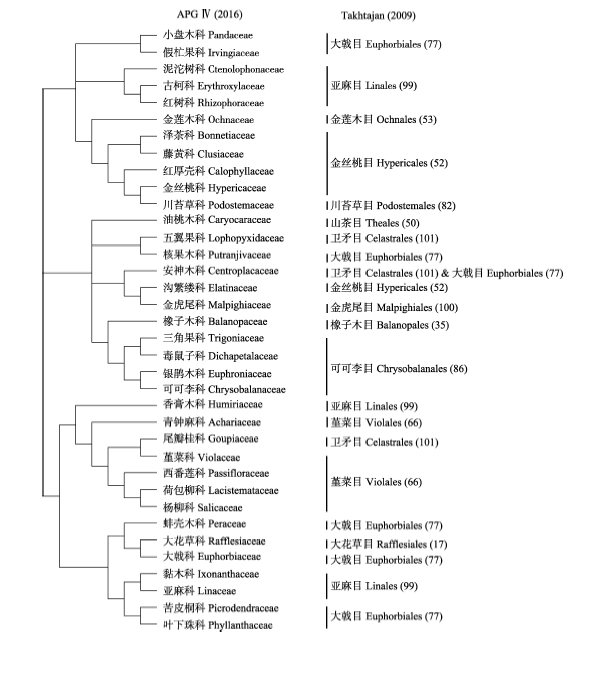
图2 比较APG IV (2016)系统与Takhtajan (2009)系统中金虎尾目的范围。括号中的数字示Takhtajan (2009)系统中目的序号。
Fig. 2 Comparison of the circumscription of Malpighiales between the APG IV (2016) and Takhtajan (2009) systems. The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009).
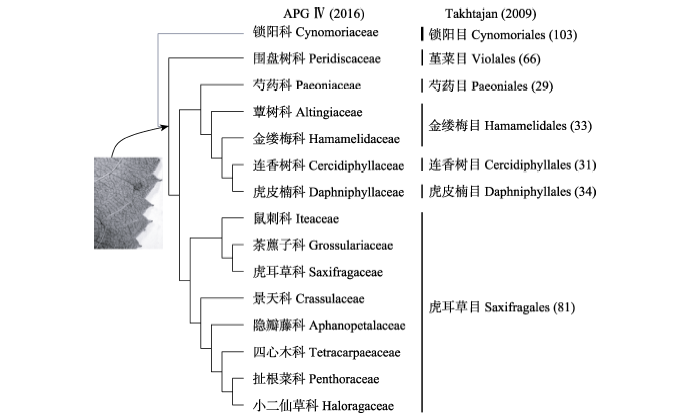
Fig. 3 Comparison of the circumscription of Saxifragales between the APG IV (2016) and Takhtajan (2009) systems. The placement of Cynomoriaceae is based on the result of Bellot et al (2016). The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009).
Results from an online survey of family delimitation in angiosperms and ferns: recommendations to the Angiosperm Phylogeny Group for thorny problems in plant classification.
Botanical Journal of the Linnean Society, 178, 501-528.
DonoghueMJ, DoyleJA (1989) Phylogenetic analysis of angiosperms and the relationships of Hamamelidae. In: Evolution, Systematics, and Fossil History of the Hamamelidae, vol. 1 (eds Crane PR, Blackmore S), pp. 17-45. Clarendon Press, Oxford, UK.
DoyleJA, HottonCL (1991) Diversification of early angiosperm pollen in a cladistic context. In: Pollen and Spores: Patterns of Diversification (eds Blackmore S, Barnes SH), pp. 165-195. Clarendon Press, Oxford, UK.
[17]
FengM, FuDZ, LiangHX, LuAM (1995)
Floral morphogenesis of Aquilegia L. (Ranunculaceae).
Acta Botanica Sinica, 37, 791-794. (in Chinese with English abstract)
Nonflowers near the base of extant angiosperms? Spatiotemporal arrangement of organs in reproductive units of Hydatellaceae and its bearing on the origin of the flower.
WuZY, LuAM, TangYC, ChenZD, LiDZ (2003) The families and genera of angiosperms in China: a comprehensive analysis. Science Press, Beijing. (in Chinese)
Large-scale phylogenetic analyses reveal fagalean diversification promoted by the interplay of diaspores and environments in the Paleogene. Perspectives in Plant Ecology,
An ordinal classification for the families of flowering plants.
1998
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II.
1
2003
... 随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics).在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993).这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义.从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟.在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统.被子植物由此成为了第一个基于分子数据建立分类系统的大类群.之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016).该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统.可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009).但并不是说APG系统已经完美, 不存在需要继续研究的问题了.经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题. ...
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
2
2009
... 随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics).在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993).这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义.从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟.在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统.被子植物由此成为了第一个基于分子数据建立分类系统的大类群.之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016).该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统.可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009).但并不是说APG系统已经完美, 不存在需要继续研究的问题了.经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题. ...
... 由于严格强调单系性, APG系统中一些目、科比较异质, 其共衍征很难确定.例如, 由莲科(Nelumbonaceae)、悬铃木科(Platanaceae)、山龙眼科(Proteaceae)组成的山龙眼目(Proteales) (APG III, 2009), 很难找到它们的共衍征, 而APG IV (2016)又把清风藤科(Sabiaceae)归入到山龙眼目, 这导致该目更为异质.又如, APG系统的金虎尾目是一个大拼盘, 包含的36个科分布在Takhtajan (2009)系统中不同位置的13个目中(图2).分化时间分析显示金虎尾目的28个主要分支在白垩纪中期(112-94 Ma)由爆发式的快速辐射所产生(Davis et al, 2005).此外, APG III (2009)界定的虎耳草目(Saxifragales)包括了Takhtajan (2009)的29、31、33、34、66和81目(图3).通过仔细研究形态性状的进化, Carlsward等(2011)发现虎耳草目的共衍征为叶具堇菜型(violoid)或山茶型(theoid)叶齿(图3).APG IV (2016)依据S. Bellot和S. Renner未发表的数据将寄生植物科——锁阳科(Cynomoriaceae)放在虎耳草目中, 而锁阳科在Takhtajan系统中独立成目——锁阳目(Cynomoriales) (第103目).近来Bellot等(2016)根据叶绿体、线粒体和核基因组的数据将锁阳科放在虎耳草目的最基部, 这导致Carlsward等(2011)所发现的叶的性状不能用于界定扩大的虎耳草目(图3).上述分析说明APG系统中一些目, 如睡莲目、山龙眼目、金虎尾目、虎耳草目以及蔷薇目(Rosales)等的划分仍需研究. ...
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV.
5
2016
... 随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics).在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993).这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义.从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟.在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统.被子植物由此成为了第一个基于分子数据建立分类系统的大类群.之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016).该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统.可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009).但并不是说APG系统已经完美, 不存在需要继续研究的问题了.经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题. ...
... Interrelationships of the APG IV (2016) orders of angiosperms. The dotted lines indicate the conflicting placements between nuclear/mitochondrial and chloroplast trees. The orders with the gray contain at least one taxon with multiple centrifugal development stamens. Pollen and cotyledonal characters are labeled on the right. ...
... Comparison of the circumscription of Malpighiales between the APG IV (2016) and Takhtajan (2009) systems. The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
... Comparison of the circumscription of Saxifragales between the APG IV (2016) and Takhtajan (2009) systems. The placement of Cynomoriaceae is based on the result of Bellot et al (2016). The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
Assembled plastid and mitochondrial genomes, as well as nuclear genes, place the parasite family Cynomoriaceae in the Saxifragales.
1
2016
... Comparison of the circumscription of Saxifragales between the APG IV (2016) and Takhtajan (2009) systems. The placement of Cynomoriaceae is based on the result of Bellot et al (2016). The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
Biogeography and diversification of Brassicales: a 103 million year tale.
Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL.
1
1993
... 随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics).在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993).这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义.从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟.在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统.被子植物由此成为了第一个基于分子数据建立分类系统的大类群.之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016).该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统.可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009).但并不是说APG系统已经完美, 不存在需要继续研究的问题了.经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题. ...
Tree of life for the genera of Chinese vascular plants.
2016
Results from an online survey of family delimitation in angiosperms and ferns: recommendations to the Angiosperm Phylogeny Group for thorny problems in plant classification.
2
2015
... 在Cronquist (1981)系统中, 被子植物共包括383科, 其中双子叶318科, 单子叶65科.在APG III (2009)中, 约87%的科的单系性得到支持, 仅有13%的科(约50个科)的界限和范围有大问题(Christenhusz et al, 2015), 如大戟科(Euphobiaceae)、大风子科(Flacourtiaceae)、百合科(Liliaceae)、马齿苋科(Portulacaceae)和檀香科(Santalaceae).而这些科多数在传统分类上也被认为是异质的, 如广义百合科是一个大杂烩, 后来曾被划分为30多个科(Dahlgren, 1983). ...
Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proceedings of the National Academy of Sciences,
1
2007
... 在传统分类系统中(如Cronquist, 1981; Dahlgren, 1983; Thorne, 1992; Takhtajan, 2009), 子叶数目(1个或2个)被作为被子植物一级分类的性状.然而, 分子系统学研究发现具双子叶的“ANITA进化阶”(grade)构成被子植物最早分化的3个分支, 即无油樟目(Amborellales)、睡莲目和木兰藤目(Austrobaileyales), 而单子叶植物与另外4个具双子叶的类群——金粟兰目(Chloranthales)、木兰类(Magnolids)、金鱼藻目(Ceratophyllales)以及真双子叶植物(Eudicots)构成被子植物早期快速辐射的五大分支(图1; Moore et al, 2007). ...
Nonflowers near the base of extant angiosperms? Spatiotemporal arrangement of organs in reproductive units of Hydatellaceae and its bearing on the origin of the flower.
Hydatellaceae identified as a new branch near the base of the angiosperm.
2007
Molecular markers and concepts of plant evolutionary relationships: Progress, promise and future prospects.
1
2009
... 随着DNA测序和生物信息技术的发展, 自20世纪90年代兴起利用分子数据研究生物类群间的系统发育关系称为分子系统发育学(Molecular Phylogenetics).在1993年, Mark Chase等42位作者合作发表了“Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL”一文(Chase et al, 1993).这是由世界几十个实验室共同完成的当时规模最大的系统发育分析, 在被子植物系统学研究中具有划时代的意义.从那之后, 人们逐渐广泛地利用DNA序列来重建植物类群间的系统发育关系, 于是植物分子系统学作为植物系统学的一个分支逐渐走向成熟.在1998年, 被子植物系统发育组(Angiosperm Phylogeny Group, APG)综合多个大尺度的系统发育分析结果, 为被子植物提出了一个目、科分类阶元上的分类系统, 简称APG系统.被子植物由此成为了第一个基于分子数据建立分类系统的大类群.之后, 随着分子数据的增加, APG系统经历了3次修订(APG, 1998; APG II, 2003; APG III, 2009; APG IV, 2016).该系统对被子植物系统学和分类学研究产生了重大影响, 大大改变了两百多年来植物学家们以形态学(广义)性状为根据提出的分类系统.可以说, 最近20年在理解植物进化历史上所取得的进展比过去200年都要大(Soltis et al, 2009).但并不是说APG系统已经完美, 不存在需要继续研究的问题了.经历了两个多世纪的发展和沉积, 人们积累了极为丰富的形态性状, 包括形态学、解剖学、胚胎学、孢粉学、细胞学和个体发生等, 这些性状在植物系统学和分类学研究中如何利用和评价仍是当前亟待解决的课题. ...
onwards) Angiosperm Phylogeny Website. Version 12, July
2012
onwards) Angiosperm Phylogeny Website. Version 12, July
2012
Paraphyly and the origin and classification of angiosperms.
... Comparison of the circumscription of Malpighiales between the APG IV (2016) and Takhtajan (2009) systems. The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
... systems. The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
... Comparison of the circumscription of Saxifragales between the APG IV (2016) and Takhtajan (2009) systems. The placement of Cynomoriaceae is based on the result of Bellot et al (2016). The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
... . The numbers in brackets indicate the serial numbers of the orders of Takhtajan (2009). ...
Large-scale phylogenetic analyses reveal fagalean diversification promoted by the interplay of diaspores and environments in the Paleogene. Perspectives in Plant Ecology,



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}