植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 )。植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 )。在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 )。植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说。然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究。
高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 )。由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 )。高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 )。“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群。这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在。
山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 )。如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 )。大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 )。然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 )。目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义。
中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点。因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏。尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题。但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观。北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 )。这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见。
本文针对中国主要高大山地的高山带种子植物区系, 在大量野外调查和数据集成的基础上, 进行高山带植物区系之间植物多样性与区系地理成分结构的比较研究, 以期探讨高山植物区系成分的地理分异格局和相关因子, 拟回答以下问题: (1) 中国高山植物区系的多样性、科属构成与地理成分存在怎样的宏观格局?(2)高山种子植物区系的地理分异与业已建立的中国种子植物区系分区系统是否吻合?(3)空间距离和地理环境因素对中国高山植物区系结构的宏观分异有怎样的影响?
1 数据与方法
1.1 物种数据、地理数据和气候数据
选择国内具有高山环境的14座主要山地, 见图1 。青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程。结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 )。台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 )。本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据。
图1 本研究选择的中国14座山地的高山带分布地点
Fig. 1 Locations of alpine zone of 14 main mountain ranges in China
参照Flora of China 的植物分类系统, 对所研究的14座山地的高山植物物种名录进行校核, 得到中国高山植物区系的总物种名录。本文分析的高山植物物种数据部分来自各合作者的野外植物调查结果, 另有部分山地的数据来自公开出版物, 具体说明见表1 。物种属性数据包括物种的拉丁科属种名、生活型及所在属的地理分布区类型, 分布区类型依据吴征镒(1991)。根据定义, 高山带植物应分布在高山树线以上, 但很多高山植物可以向下分布到中山森林带。在第四纪冰期, 青藏高原腹地以外的中国植被-气候带海拔普遍下移500-1,500 m(郑卓, 1999 ; 施雅风等, 2006 )。随着冰后期气候变暖, 物种分布范围上升, 但根据野外调查观测, 大量物种的分布下限当前仍滞留在高山树线以下。因此, 本文中采取统一标准选择高山物种, 即其分布的上限高度显著高于当前树线的海拔(>100 m), 其下限不低于亚高山针叶林, 即所在山地冷杉(Abies spp.)林和落叶松(Larix spp.)林的下限海拔, 建立的物种名录不包括树线树种和少数海拔分布极宽的植物种, 如车前(Plantago spp.)等。因为前者本来就属于亚高山分布, 后者作为广布种对高山环境完全没有指示意义和山体之间的区分作用。
根据上述14座山地的高山树线和永久雪线, 在Google Earth上运用“添加-多边形”工具勾画各个山地高山带分布范围的多边形(polygon), 即高山树线以上、永久雪线以下的地带范围(图1 )。将数据存为.kmz文件, 在地理信息系统软件ArcGIS 10.2 (www.esri.com/software/arcgis/)中转换成.shp格式。利用美国国家航空航天局发布的3″分辨率全球数字高程模型(SRTM v4.1, http://datamirror.csdb.cn/)截取各山地高山带范围, 应用Asia North Albers Equal Area Conic投影后重采样成分辨率100 m的研究区数字高程模型(Farr & Kobrick, 2000 ; van Zyl, 2001 ), 据此提取每座高山的最高海拔, 并计算了各山地高山区域的投影面积。
同时, 用各山地高山区的数字高程模型数据, 从世界气候数据库(www.worldclim.org/, WorldClim v1.4)中, 提取每个高山区的不同气候指标数据(BIO1-19)。该数据库利用全球各地气象站记录的气象观测资料, 采用薄片样条函数模型插值生成全球气候栅格数据(1950-2000年平均值), 在WPS1984投影下的空间分辨率为1 km (Hijmans et al, 2005 )。
图2 中国14座山地的高山植物区系地理成分构成。T1: 世界分布型; T2-7: 热带分布型; T8-11: 北温带分布型; T12-13: 古地中海分布型; T14: 东亚分布型; T14SH: 中国-喜马拉雅分布型; T15: 中国特有分布型。
Fig. 2 Percentage compositions of alpine flora of 14 main mountains in China. T1, Cosmopolitan; T2-7, Tropic types; T8-11, North temperate types; T12-13, Mediterranean types; T14, East Asia types; T14SH, Sino-Himalayan type; T15, Endemic to China.
1.2 分析方法
分别在科、属、种水平评估和比较各山地高山带的植物多样性。采用中国种子植物属地理分布区类型系统(吴征镒, 1991 )划分各山地高山带植物区系属的地理成分类型, 并分别统计不同分布型的属数及其所包含的种数。参照吴征镒等(2010), 根据不同地理成分之间的联系将世界分布型之外的类型进一步分为热带成分(T2-7)、北温带成分(T8- 11)、古地中海成分(T12-13)和东亚成分(T14-15)。采用Pearson相关系数分析不同山地高山带植物区系之间的相似性, 包括分类群构成和区系成分构成的相似性。
用一元线性或非线性回归模型分别拟合上述五大区系地理成分的属数百分比和种数百分比沿经度和纬度梯度的空间格局, 并用全部15个地理分布型的属数百分比结合t 检验来分析不同山地高山带的种子植物区系之间关联性的显著性水平。
采用Mantel检验和偏Mantel检验来评估地理和环境距离对植物区系构成差异的影响(韩杰等, 2016 )。选择各山地高山带分布范围的几何中心经纬度坐标代表该区地理位置, 选择高山树线海拔(m)、山体最大海拔(m)和地形投影面积(km2 )作为地理距离相关变量; 选用高山树线附近的平均气候状况, 包括年均温(mean annual temperature, MAT, ℃)、最冷月最低温(minimum temperature of coldest month, MTCM, ℃)、最热月最高温(maximum temperature of coldest month, MTWM, ℃)、年降水量(mean annual precipitation, MAP, mm)来代表各山地高山带的气候条件。用欧几里得距离计算各高山带之间的地理空间距离和环境距离, 用Jaccard相异性系数计算各高山植物区系之间的相异性, 用偏Mantel检验来评估空间距离和环境差异的独立贡献。
上述数量分析在统计软件R (Version 3.2.5 ; R Development Core Team 2015 )中完成。
2 结果
2.1 中国高山植物区系的物种丰富度、科属及区系成分构成
根据统计, 全部14座山地的高山带植物区系包含维管束植物共计77科507属3,417种63亚种282变种; 其中, 蕨类植物12科18属86种5变种, 裸子植物1科2属10种2变种, 单子叶植物9科96属579种19亚种33变种, 双子叶植物55科391属2,742种44亚种242变种。
根据14座山地高山带的植物记录, 高山植物区系种类最多的前10科依次为: 菊科58属484种, 禾本科35属233种, 毛茛科24属233种, 蔷薇科20属201种, 杜鹃花科8属191种, 玄参科13属181种, 十字花科40属161种, 龙胆科12属152种, 石竹科15属151种, 莎草科7属147种。而种类最丰富的前10个属依次是: 杜鹃属(Rhododendron ) 168种, 马先蒿属(Pedicularis ) 120种, 报春花属(Primula ) 96种, 龙胆属(Gentiana ) 96种, 苔草属(Carex ) 95种, 风毛菊属(Saussurea ) 92种, 虎耳草属(Saxifraga ) 86种, 委陵菜属(Potentilla ) 62种, 葶苈属(Draba ) 58种, 早熟禾属(Poa ) 54种。
这些高山植物全部489个种子植物属包含15个地理分布区类型18个亚类。其中世界分布型47属, 占总数的近10%。在其余442属中, 北温带分布(T8及其亚型)是最大的组分(180属), 占属级地理成分(除世界分布)的40.7%,包括南、北温带间断分布(T8-4)的30属和北极-高山分布型(T8-2)的11属。其次是旧世界温带分布型(T10)和东亚-北美间断分布型(T9), 合计占15.4%; 古地中海分布型(T12和T13及各自的亚型)合计有54属, 占12.2%; 东亚特有分布型合计有66属, 其中中国-喜马拉雅分布亚型(T14SH)有47属, 占总数的10.6%, 而中国-日本分布亚型(T14SJ)完全未见于高山植物区系。中国特有分布型有23个属, 占5.2%。而热带分布诸型(T2-7)合计仅15属, 占3.4%。
2.2 高山植物区系成分及优势科属的地理格局
各山地高山种子植物区系地理成分的构成与其总体表现出类似的结构(附录1), 五大主要地理成分在14座山地中表现出不一致的地理格局特征(图3 )。其中,与基于种数统计的区系成分百分比相比, 基于属数统计的区系成分百分比沿经向和纬向地理梯度的变化更为显著。具体情况如下: (1)世界分布成分占全部种、属的比例沿纬度并无显著的梯度变化, 而分别在种(0.05 < P < 0.1)和属(P < 0.05)的层面呈现出自西向东增加的趋势。(2)热带成分自南向北在属和种数比例上都呈现接近显著(0.05 < P <0.1)的递减趋势, 自西向东在属的比例上也有接近显著的降低趋势。(3)作为高山植物区系主导性成分的北温带分布类型则出现与热带成分大致相反的趋势, 在属和种的比例上自南向北有接近显著的增加趋势, 且在经向上自西向东增加的趋势更为显著(P < 0.001), 在东部高山植物区系中几乎占到100%的比例。(4)古地中海成分的比例沿纬度梯度的变化并不显著, 仅在种水平有接近显著的自南向北增加趋势, 而在经向上属和种的比例从西向东均显著递减(P < 0.001)。(5)包括东亚分布、中国-喜马拉雅分布和中国特有分布在内的东亚成分所占的比例, 在属和种水平都呈现出最为显著的沿纬度递减趋势(P < 0.001), 而在经向上则表现为中间高、东西两边低的格局(P < 0.05)。
图3 不同区系地理成分百分比的经、纬度格局
Fig. 3 Latitudinal and longitudinal patterns of the geographical elements of alpine floras
在14座山地高山种子植物区系物种数最多的前10科中, 除杜鹃花科以外, 其余9科同时也是一半以上的山地(>7座)高山种子植物区系前10位的优势科(附录2)。在天山、祁连山、阿尔金山和珠峰蝶形花科成为优势科之一, 在高黎贡山(杜鹃花科第一)和台湾山脉(兰科第二)也分别有独特的重要科。在高山种子植物区系前10位的属中, 除早熟禾被嵩草(Kobresia )替代之外, 其余9属也正好是超过一半山地(>7座)前10位的优势属(附录2)。但各山地之间的差异较大, 例如: 台湾区系与区系整体的前10属仅有2个是相同的, 而珠峰区系与区系整体的前10属仅有1属不同。图4 显示了各山地高山种子植物区系之间地理成分(以属数计)百分比的相似性空间格局。
图4 14座山地高山种子植物区系属的地理分布区类型百分比构成的相似性格局
Fig. 4 The similarity patterns of the percentage composition of alpine floristic elements in the 14 main mountains in China
从14座山地高山植物区系属的地理成分百分比可以看出, 各山地之间都存在不同程度的相似性, 表现出以下特点: (1)天山与高黎贡山、台湾山脉之间, 以及高黎贡山与贺兰山之间的高山植物区系地理成分相关不显著。(2)天山与其他山地的高山植物区系的关联相对最为疏远。(3)高黎贡山和珠峰的高山植物区系与青藏高原以外的北方山地(如长白山、五台山、太白山、天山和贺兰山)联系较弱, 而与同在高原周边的贡嘎山、摩天岭、祁连山的联系明显更强; 除天山以外, 北方山地之间的高山植物区系也有较密切的联系。(4)台湾海峡的存在并未造成台湾高山植物区系地理成分与大陆山地之间区系成分的更大差异, 但台湾的高山植物区系与北方山地(尤其是东部)的联系明显强于纬度接近的喜马拉雅-横断山区。
2.3 各山地高山植物区系联系的环境与空间关联
Mantel检验表明, 不同山地高山带之间的地理空间距离(R = 0.601, P = 0.001)比环境距离(R = 0.551, P = 0.002)对植物区系地理成分构成的差异性影响更大。尤其是偏Mantel检验表明, 在控制环境差异(全部显著因子)影响的情况下, 单纯的地理空间距离效应仍然显著(R = 0.445, P = 0.001), 而在控制距离效应的情况下, 影响显著的环境因子距离矩阵对区系地理成分构成差异的解释力较低(R = 0.354, P = 0.008)。
把高山带环境因子分为空间形态特征(包括高山带面积的log值和山体最大高度)和气候特征两部分。针对高山带空间特征差异的Mantel检验及控制地理距离的偏Mantel检验表明, 高山带形态特征差异对区系地理成分构成差异的影响接近环境因子整体的影响, R 值分别为0.546 (P = 0.002)和0.353 (P = 0.017)。另一方面,气温、年降水量、最热月最高温、最冷月最低温4个高山带气候因子的差异影响中, 最热月最高温差异对高山带区系地理成分构成的差异有最显著的影响, Mantel检验和控制地理距离的偏Mantel检验的R 值分别为0.213(P = 0.046)和0.201 (P = 0.064)。
各山地高山带之间地理距离对区系地理成分差异的影响在分别控制全部环境因子、空间形态因子和最热月最高温的效应下, 偏Mantel检验的R 值分别为0.440 (P = 0.004)、0.445 (P = 0.004)和0.598 (P = 0.001), 也表明高山带空间特征的效应强于最热月最高温差异的效应。
3 讨论
根据统计, 本研究涉及的14座主要山地的高山带至少分布有种子植物65科489属3,670种(含种下单位)。根据中国植物多样性的最新统计资料(王利松等, 2015 ), 这些高山植物大约占中国分布的种子植物总科数的24.9%、总属数的16.3%和总种数的12.3%。尽管高山带环境极端恶劣, 植被稀疏、结构简单, 但在这仅占全国陆地面积0.13% (投影面积约12,747.85 km2 ), 分散于14座高山的区域内所分布的植物多样性是非常高的。
《中国种子植物区系地理》(吴征镒等, 2010 ) 一书已经对中国的种子植物区系起源、成分结构、全球地位进行了全面总结, 并提出了全国尺度的植物区系区划方案。已有的关于植物区系地理成分的分布格局的研究主要是以行政区域(一般为省)或特定自然地理区划单元(如自然地理区)为对象进行分析的, 尽管区域间植物区系成分构成的比较是植物区系地理研究的一种重要途径(沈泽昊和张新时, 2000 ), 但针对山地、平原、高原、湖泊等自然景观单元的植物区系比较往往局限在有限的区域范围内, 并不能有效地检验中国植物区系区划系统的合理性。本研究首次在全国尺度上对高山植物区系的地理成分进行了区域比较和宏观格局分析。
3.1 高山植物区系的区域归属与联系
在中国种子植物区系分区系统中, 天山属于泛北极植物区, 阿尔金山、祁连山和贺兰山则属于古地中海植物区; 台湾的高山带被明确划归古热带植物区, 本文所涉及的其他山地均属于东亚植物区, 其中长白山、五台山、太白山属于中国-日本森林亚区,西南山地包括摩天岭、贡嘎山、高黎贡山、玉龙雪山和轿子雪山均属于中国-喜马拉雅植物亚区, 珠峰地区则属于青藏高原亚区(吴征镒等, 2010 )。
本文结果表明, 位于泛北极植物区中的天山高山带种子植物区系的确表现出与其余山地高山植物区系最大的平均差异; 青藏高原周缘山地的高山植物区系之间存在着较为密切的联系, 同处中国-日本植物区的北方山地如长白山、五台山、太白山之间也存在很高的区系相似性。此外, 古地中海区系成分随经度增加而迅速降低并在贺兰山以东的北方山地不再出现。这些格局特征从高山植物区系的角度显示了不同区系地理成分的区域分异及其边界特征, 为中国植物区系区划系统提供了支持。另外, 本文数据还表明, 在散布全国的14座山地高山植物区系中, 中国特有分布属的比例达到5.2%, 相比于中国植物区系地理成分整体构成而言(沈泽昊和张新时, 2000 )是一个相当高的比例, 表明高山带局部发生的物种形成十分旺盛。包括中国特有分布在内的东亚特有成分百分比在高山带呈现出自南向北递减、东西方向上中部隆起的分布格局, 与全国整体和低海拔地区的格局完全吻合。显示出中国西南山地对于中国特有植物分布和东亚植物区系发育的重要意义。
但是, 针对高山植物区系的分析也揭示了新的区系地理特征。首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分。这也显示了中国-日本分布成分不属于高山分布型植物。其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系。第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 )。第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系。尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系。这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 )。
值得指出的是, 目前基于植物区系地理成分构成进行的区系分区方案主要是针对较大区域进行的研究结果(吴征镒等, 2010 )。本文数据针对高山区特定范围的数据, 所得结果应是对既有成果的一种补充, 也是在不同尺度上的验证。我们发现台湾的高山带植物区系仍保留有鲜明而深远的泛北极和东亚北部区系成分的影响, 不应归于古热带植物区, 这一结果与Zhu (2016)的观点一致; 喜马拉雅山脉中段的珠峰地区高山带(尤其是南坡)植物区系也明显不同于青藏高原生物区; 天山而不是隔海相望的台湾高山带的植物区系与中国其他高山区的植物区系更加隔离。这些与既有认识不一致之处值得进一步探索验证。
3.2 高山带范围和气候条件对区系成分构成的 影响
岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 )。但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 )。
本文结果表明, 中国高山植物区系之间的地理成分构成变化受地理距离的隔离比环境差异(主要是气候)的影响更大。同时, 在各气候因子中, 能量差异的影响高于寒冷限制和水分差异所带来的影响, 表现为单一环境因子的Mantel检验结果中, 暖季高温差异(R = 0.213, P = 0.046)的相关性大于年均温差异(R = 0.189, P = 0.066)、冷季低温差异(R = 0.185, P = 0.075)和年降水量差异(R = 0.007, P = 0.469)。另外, 区系相似性空间格局表明, 台湾海峡并没有显示出对高山植物区系产生空间距离之外的隔离效应。
高山带是同时受到山地之间的几何距离、沿海拔梯度的非生物环境距离以及宽阔的山地森林带造成的生物竞争性距离等多重隔离的岛屿型非森林生境, 彼此之间物种迁移受隔离的程度应不亚于真正的岛屿。同时高山带是极端气候下形成的高度特化的生态系统, 彼此之间在气候环境上的相似性更为明显, 包括巨大的日温差、高辐射、稀薄空气和大风等(Körner, 2003 )。本文分析的14座山地高山带之间的两两平均几何距离在16.836个经纬度, 即 >1,000 km, 由此带来的隔离效应强于环境距离是可以理解的。
另一方面, 面积大小和生境的异质性是岛屿承载不同生态属性物种能力的决定因子, 并决定了岛屿物种更替的灭绝速率(Russell et al, 2006 ; Si et al, 2014 )。因此, 岛屿面积和海拔范围的差异与岛屿物种构成的差异具有正相关性的结果同样适用于高山带。全球高山树线的海拔高度非常一致地对应于生长季均温(约6.7℃)等值线(Körner & Paulsen, 2004 ), 表明生长季的能量积累是山地高山带植物分布的主要限制因子。本文数据表明, 最热月最高温比其他气候指标对高山带植物区系相似性的影响更大, 主要的原因可能正是特定山地高山带的平均最热月最高温与高山带范围大小之间存在内在的关联。
3.3 研究结果的不确定性分析
受不同山地高山带植物区系调查强度和调查范围的影响, 本文分析的14座山地的高山植物区系名录会存在不同程度的不确定性, 主要包括调查不充分导致的物种名录不完整、物种鉴定不准确导致的种类误差、不同物种名录因采用不同分类系统而造成的异名以及新近发表的物种或物种分类位置调整所带来的误差等等。为了降低数据的不确定性对结果可靠性的影响, 我们在数据分析阶段将不同名录的分类系统均调整为Flora of China 的系统, 以消除异名产生的影响; 同时, 尽管统计了各高山植物区系全部维管束植物的丰富度, 但重点分析种子植物属的分布区类型, 这不仅可以降低物种鉴定误差带来的影响(主要出现在种和种下水平), 也避免了蕨类植物分布区类型划分没有直接依据的困难。需要说明的是, 目前的中国植物区系地理成分的分布区类型系统是基于种子植物属而建立的, 如果直接套用于种水平会存在一定的误差, 且这个误差从未得到评估。考虑到建立全国甚至仅仅高山带植物种的分布区类型都仍是一时难以完成的工作, 本文中不同区系地理成分的种数统计仅仅是对属水平分析的一种附加统计, 而非种的地理成分分析。
综上, 基于种子植物属的分布区类型的数据分析能较好地消减因取样不足造成的不同数据之间的可比性, 同时基于不同区系成分百分比构成的分析也消减了不同名录样本量大小差异的影响, 因此可以认为本文结果具有较高的可靠性。当然, 由于样本量和各山地数据质量的差异, 对高山植物多样性格局更加深入的分析会因而受限。
4 小结
中国山地的高山带分布着物种丰富的植物区系, 主要由北温带分布及其亚型、世界分布、旧世界温带分布和东亚分布及中国-喜马拉雅分布等地理成分构成, 缺少中国-日本分布成分, 中国特有属的比例较高。中国14座主要山地高山植物区系构成的地理分异显示了自南向北热带和东亚成分减少、北温带成分增加, 自西向东古地中海成分减少、北温带成分增加、而东亚成分在中部达到最大值的格局特征; 14座山地的高山植物区系之间在属级地理成分构成上具有显著的相似性, 其中北方山地和青藏高原周缘山地构成了区系成分近似的两大群组, 台湾与大陆东部北方山地的高山植物区系之间联系更深。上述结果对我国植物区系地理分区系统提供了一个独特的验证。此外, 地理距离的隔离是高山植物区系分异的首要因素, 高山带的面积大小也影响到其区系成分的构成, 而夏季热量是显著影响中国高山植物区系地理分异的关键气候因子。
(责任编辑: 周玉荣)
附录 Supplementary Material
附录1 中国14座主要山地高山种子植物区系属的地理成分构成
Appendix 1 The percentage composition of floristic elements of alpine seed plants in 14 main mountain ranges in China
http:// www.biodiversity-science.net/fileup/PDF/2017014-1.pdf
附录2 中国14座主要山地高山种子植物区系前10科和前10个属及其物种数
Appendix 2 The top 10 families and top 10 genera of seed plant species number of alpine flora in the 14 main mountains in China
http:// www.biodiversity-science.net/fileup/PDF/2017014-2.pdf
致谢: 两位匿名审稿人对本文初稿提出宝贵意见和建议。谨致谢忱!
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bliss LC 1971 ) Arctic and alpine plant life cycles.
Annual Review of Ecology and Systematics , 2 , 405 -438 .
[本文引用: 2]
[2]
Cao W Li JY 2003 ) Natural Distribution of Plants in Changbai Mountain . Northeast University Press, Shenyang .
[本文引用: 2]
(in Chinese) [曹伟 , 李冀云 (2003 ) 长白山植物自然分布. 东北大学出版社 , 沈阳.]
[本文引用: 2]
[3]
Caplat P Edelaar P Dudaniec RY Green AJ Okamura B Cote J Ekroos J Jonsson PR Löndahl J Tesson SVM Petit EJ 2016 ) Looking beyond the mountain, dispersal barriers in a changing world.
Frontiers in Ecology and the Environment , 14 , 261 -268 .
[本文引用: 2]
[4]
Chapin III FS Körner C 1994 ) Arctic and alpine biodiversity, patterns, causes and ecosystem consequences.
Trends in Ecology & Evolution , 9 , 45 -47 .
[本文引用: 2]
[5]
Chen LZ 2014 ) Geography of Flora and Vegetation in China . Science Press, Beijing .
[本文引用: 2]
(in Chinese) [陈灵芝 (2014 ) 中国植物区系与植被地理. 科学出版社 , 北京.]
[本文引用: 2]
[6]
Chen S Jiang G Zhang J Li Y Qian H 2011 ) Species turn- over of amphibians and reptiles in eastern China, disentangling the relative effects of geographic distance and environmental difference.
Ecological Research , 26 , 949 -956 .
[本文引用: 2]
[7]
Chen ZD Ying JS Lu AM 2012 ) Disjunct distribution of seed plants between Southwest China and Taiwan Island of China.
Chinese Bulletin of Botany , 47 , 551 -570 .
[本文引用: 2]
(in Chinese with English abstract) [陈之端 , 应俊生 , 路安民 (2012 ) 中国西南地区与台湾种子植物间断分布现象. 植物学报,
47 , 551 -570 .]
[本文引用: 2]
[8]
Cui NR Akefu P 1993 ) The formation and the basic nature of the flora in the Argion Mountain Natural Reserve, China.
Journal of Xinjiang Normal University (Natural Sciences Edition) ,1 , 47 -53 .
[本文引用: 1]
(in Chinese with English abstract) [崔乃然 , 彼得·爱克夫 (1993 ) 中国阿尔金山自然保护区植物区系组成及基本特征. 新疆师范大学学报(自然科学版),
1 , 47 -53 .]
[本文引用: 1]
[9]
Cui ZJ Chen YX Zhang W Zhou SZ Zhou LP Zhang M Li CC 2011 ) Research history, glacial chronology and origins of Quaternary glaciations in China.
Quaternary Sciences , 31 , 749 -764 .
[本文引用: 2]
(in Chinese with English abstract) [崔之久 , 陈艺鑫 , 张威 , 周尚哲 , 周力平 , 张梅 , 李川川 (2011 ) 中国第四纪冰期历史、特征及成因探讨. 第四纪研究,
31 , 749 -764 .]
[本文引用: 2]
[10]
Farr TG Kobrick M 2000 ) Shuttle radar topography mission produces a wealth of data. Eos,
Transactions American Geophysical Union , 81 , 583 -585 .
[本文引用: 2]
[11]
Feng JM Wang XP Xu CD Yang YH Fang JY 2006 ) Altitudinal patterns of plant species diversity and community structure on Yulong Mountains, Yunnan, China.
Journal of Mountain Science , 24 , 110 -116 .
[本文引用: 1]
(in Chinese with English abstract) [冯建孟 , 王襄平 , 徐成东 , 杨元合 , 方精云 (2006 ) 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报,
24 , 110 -116 .]
[本文引用: 1]
[12]
Han J Shen ZH Shi SL Peng PH 2016 ) Comparison of plant species diversity and composition in the dry valleys of Yalong River and Dadu River: evaluating the effects of climate, topography and space.
Biodiversity Science , 24 , 421 -430 .
[本文引用: 2]
(in Chinese with English abstract) [韩杰 , 沈泽昊 , 石松林 , 彭培好 (2016 ) 雅砻江和大渡河干旱河谷植被物种多样性比较: 气候、地形与空间的影响. 生物多样性,
24 , 421 -430 .]
[本文引用: 2]
[13]
Hijmans RJ Cameron SE Parra JL Jones PG Jarvis A 2005 ) Very high resolution interpolated climate surfaces for global land areas.
International Journal of Climatology , 25 , 1965 -1978 .
[本文引用: 2]
[14]
Keil P Schweiger O Kühn I Kunin WE Kuussaari M Settele J Henle K Brotons L Pe’er G Lengyel S Moustakas A Steinicke H Storch D 2012 ) Patterns of beta diversity in Europe, the role of climate, land cover and distance across scales.
Journal of Biogeography , 39 , 1473 -1486 .
[本文引用: 1]
[15]
Körner C Paulsen J 2004 ) A world-wide study of high altitude treeline temperatures.
Journal of Biogeography , 31 , 713 -732 .
[本文引用: 1]
[16]
Körner C 2003 ) Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Springer, Berlin.
[本文引用: 2]
[17]
Kropf M Comes HP Kadereit JW 2006 ) Long-distance dispersal vs vicariance, the origin and genetic diversity of alpine plants in the Spanish Sierra Nevada.
New Phytologist , 172 , 169 -184 .
[本文引用: 1]
[18]
Li CF Chytry M Zelený D Hsieh CF 2013 ) Classification of Taiwan forest vegetation.
Applied Vegetation Science , 16 , 698 -719 .
[本文引用: 1]
[19]
Li H Guo HJ Dao ZL 2000 ) Flora of Gaoligong Mountains . Science Press, Beijing .
[本文引用: 1]
(in Chinese) [李恒 , 郭辉军 , 刀志灵 (2000 ) 高黎贡山植物. 科学出版社 , 北京.]
[本文引用: 1]
[20]
Li LP Yin LK Tang ZY 2011 ) Distribution patterns of the species richness of plants and animals in Xinjiang, China.
Arid Zone Research , 28 , 1 -9 .
[本文引用: 1]
(in Chinese with English abstract) [李利平 , 尹林克 , 唐志尧 (2011 ) 新疆野生动植物物种丰富度的分布格局. 干旱区研究,
28 , 1 -9 .]
[本文引用: 1]
[21]
Liang CZ 2012 ) Vegetation of Helan Mountain . Sunshine Press, Yinchuan .
[本文引用: 1]
(in Chinese) [梁存柱 (2012 ) 贺兰山植被. 阳光出版社 , 银川.]
[本文引用: 1]
[22]
López-Pujol J Zhang FM Sun HQ Ying TS Ge S 2011 ) Mountains of Southern China as “plant museums” and “plant cradles”, evolutionary and conservation insights.
Mountain Research and Development , 31 , 261 -269 .
[本文引用: 1]
[23]
Lu AM 1999 ) The Geography of Spermatophytic Families and Genera . Science Press, Beijing .
[本文引用: 2]
(in Chinese) [路安民 (1999 ) 种子植物科属地理. 科学出版社 , 北京.]
[本文引用: 2]
[24]
Luo D Yue JP Sun WG Xu B Li ZM Comes HP Sun H 2016 ) Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains, first insights from comparative phylogeography of four perennial herbs.
Journal of Biogeography , 43 , 31 -43 .
[本文引用: 1]
[25]
Luo J Zheng WL Pan G Wang JS 2006 ) Study on spermatophyte flora of the alpine frigid zone in Shergyla Mountain of Tibet.
Journal of Wuhan Botanical Research , 24 , 215 -219 .
(in Chinese with English abstract) [罗建 , 郑维列 , 潘刚 , 王景升 (2006 ) 色季拉山区高山寒带种子植物区系研究. 武汉植物研究,
24 , 215 -219 .]
[26]
MacArthur RH Wilson EO 1967 ) The Theory of Island Biogeography . Princeton University Press, Princeton .
[本文引用: 1]
[27]
Marshall J 1957 ) The Birds of the Pine-oak Woodland in Southern Arizona and Adjacent Mexico.
Pacific Coast Avifauna , 32 , 5 -125 .
[本文引用: 1]
[28]
Mary E Diamond JM 1976 ) Birds on islands in the sky, origin of the montane avifauna of Northern Melanesia. Proceedings of the National Academy of Sciences,
USA , 73 , 1765 -1769 .
[本文引用: 1]
[29]
Packer JG 1974 ) Differentiation and dispersal in alpine floras.
Arctic and Alpine Research , 6 , 117 -128 .
[本文引用: 1]
[30]
Peng H Liu ED 2015 ) Yunnan Jiaozi Snow Mountain National Nature Reserve . China Forestry Publishing House, Beijing .
(in Chinese) [彭华 , 刘恩德 (2015 ) 云南轿子雪山国家级自然保护区. 中国林业出版社 , 北京.]
[31]
Qian H Ricklefs RE 2012 ) Disentangling the effects of geographic distance and environmental dissimilarity on global patterns of species turnover.
Global Ecology and Biogeography , 21 , 341 -351 .
[本文引用: 1]
[32]
Ren Y 2006 ) Study and Management of Biodiversity in Taibai Mountain Nature Reserve . China Forestry Publishing House, Beijing .
[本文引用: 1]
(in Chinese) [任毅 (2006 ) 太白山自然保护区生物多样性研究与管理. 中国林业出版社 , 北京.]
[本文引用: 1]
[33]
Russell GJ Diamond JM Reed TM Pimm SL 2006 ) Breeding birds on small islands, island biogeography or optimal foraging?
Journal of Animal Ecology , 75 , 324 -339 .
[本文引用: 1]
[34]
Sandel B Arge L Dalsgaard B 2011 ) The influence of late Quaternary climate-change velocity on species endemism.
Science , 334 , 660 -664 .
[本文引用: 1]
[35]
Shen ZH Liu ZL Wu J 2004 ) Altitudinal pattern of flora on the eastern slope of Mt. Gongga.
Biodiversity Science , 12 , 89 -98 .
[本文引用: 1]
(in Chinese with English abstract) [沈泽昊 , 刘增力 , 伍杰 (2004 ) 贡嘎山东坡植物区系的垂直分布格局. 生物多样性,
12 , 89 -98 .]
[本文引用: 1]
[36]
Shen ZH Zhang XS 2000 ) A quantitative analysis on the floristic elements of the Chinese subtropical region and their spatial patterns.
Acta Phytotaxonomica Sinica , 38 , 366 -380 .
[本文引用: 4]
(in Chinese with English abstract) [沈泽昊 , 张新时 (2000 ) 中国亚热带地区植物区系地理成分及其空间格局的数量分析. 植物分类学报,
38 , 366 -380 .]
[本文引用: 4]
[37]
Shen ZF 1997 ) Biogeography of Taiwan. I. Some preliminary thinking and studies.
Annuals of Taiwan Provincial Museum , 40 , 361 -450 .
[本文引用: 1]
(in Chinese) [沈中桴 (1997 ) 台湾的生物地理. 1. 一些初步思考和研究. 台湾省立博物馆年刊,
40 , 361 -450 .]
[本文引用: 1]
[38]
Shi YF Cui ZJ Su Z 2006 ) The Quaternary Glaciations and Environmental Variations in China . Hebei Science and Technology Publishing House, Shijiazhuang .
[本文引用: 1]
(in Chinese) [施雅风 , 崔之久 , 苏珍 (2006 ) 中国第四纪冰川与环境变化.河北科学技术出版社 , 石家庄.]
[本文引用: 1]
[39]
Si X Pimm SL Russell GJ Ding P 2014 ) Turnover of breeding bird communities on islands in an inundated lake.
Journal of Biogeography , 41 , 2283 -2292 .
[本文引用: 2]
[40]
Sun H 2002 ) Tethys retreat and Himalayas-Hengduanshan Mountains uplift and their significance on the origin and development of the Sino-Himalayan elements and alpine flora.
Acta Botanica Yunnanica , 24 , 273 -288 .
[本文引用: 2]
(in Chinese with English abstract) [孙航 (2002 ) 古地中海退却与喜马拉雅-横断山的隆起在中国喜马拉雅成分及高山植物区系的形成与发展上的意义. 云南植物研究,
24 , 273 -288 .]
[本文引用: 2]
[41]
Tackenberg O Stöcklin J 2009 ) Wind dispersal of alpine plant species, a comparison with lowland species.
Journal of Vegetation Science , 19 , 109 -118 .
[本文引用: 1]
[42]
Tao JR 1992 ) The Tertiary vegetation and flora and floristic regions in China.
Acta Phytotaxonomica Sinica , 30 , 25 -43 .
[本文引用: 1]
(in Chinese with English abstract) [陶君容 (1992 ) 中国第三纪植被与植物区系历史及分区. 植物分类学报,
30 , 25 -43 .]
[本文引用: 1]
[43]
van Zyl JJV 2001 ) The shuttle radar topography mission (SRTM), a breakthrough in remote sensing of topography.
Acta Astronautica , 48 , 559 -565 .
[本文引用: 1]
[44]
Wang HS 1992 ) Floristic Geography . Science Press, Beijing .
[本文引用: 1]
(in Chinese) [王荷生 (1992 ) 植物区系地理. 科学出版社 , 北京.]
[本文引用: 1]
[45]
Wang J 2012 ) Study on the flora of spermatophyte in Xinluhai Nature Reserve of the Queer Mountain.
Journal of Anhui Agricultural Science , 40 , 11020 -11021 .
[本文引用: 1]
(in Chinese with English abstract) [王静 (2012 ) 雀儿山新路海自然保护区高山种子植物区系研究. 安徽农业科学,
40 , 11020 -11021 .]
[本文引用: 1]
[46]
Wang LS Jia Y Zhang XC Qin HN 2015 ) Overview of higher plant diversity in China.
Biodiversity Science , 23 , 217 -224 .
[本文引用: 1]
(in Chinese with English abstract) [王利松 , 贾渝 , 张宪春 , 覃海宁 (2015 ) 中国高等植物多样性. 生物多样性,
23 , 217 -224 .]
[本文引用: 1]
[47]
Wang WC 1992 ) On some distribution patterns and some migration routes found in the eastern Asiatic region. Acta Phytotaxonomic Sinica, 30, 1-24
; 30 , 97 -117 .
[本文引用: 3]
(in English with Chinese abstract) [王文采 (1992 ) 东亚植物区系的一些分布式样和迁移路线. 植物分类学报, 30, 1-24;
30 , 97 -117 .]
[本文引用: 3]
[48]
Wondimu T Gizaw A Tusiime FM Masao CA Abdi AA Gussarova G Popp M Nemomissa S Brochmann C 2014 ) Crossing barriers in an extremely fragmented system, two case studies in the afro-alpine sky island flora.
Plant Systematics and Evolution , 300 , 415 -430 .
[本文引用: 1]
[49]
Wu ZY 1991 ) The areal-types of Chinese genera of seed plants.
Acta Botanica Yunnanica , 13 (Suppl. IV ), 1 -139 .
[本文引用: 1]
(in Chinese with English abstract) [吴征镒 (1991 ) 中国种子植物属的分布区类型. 云南植物研究,
13 (增刊IV ), 1 -139 .]
[本文引用: 1]
[50]
Wu ZY Sun H Zhou ZK Li DZ Peng H 2010 ) Floristic Geography of Seed Plants in China . Science Press, Beijing .
[本文引用: 4]
(in Chinese) [吴征镒 , 孙航 , 周浙昆 , 李德铢 , 彭华 (2010 ) 中国种子植物区系地理. 科学出版社 , 北京.]
[本文引用: 4]
[51]
Wurster CM Bird MI 2014 ) Barriers and bridges, early human dispersals in equatorial SE Asia. Geology and Archaeology, Submerged Landscapes of the Continental Shelf (eds Harff J, Bailey G, Lüth F),
Special Publications, pp . 411 . Geological Society, London.
[本文引用: 1]
[52]
Yan YH Zhang XC Ma KP 2013 ) Pteridophytes in China : Diversity and Distribution. Science Press, Beijing .
[本文引用: 1]
(in Chinese) [严岳鸿 , 张宪春 , 马克平 (2013 ) 中国蕨类植物多样性与地理分布. 科学出版社 , 北京.]
[本文引用: 1]
[53]
Yang QS Liu JQ Wang YK 2008 ) The Integrated Scientific Investigation Report of Gansu Qilian Mountain National Nature Reserve . Gansu Science and Technology Press, Lanzhou .
(in Chinese) [杨全生 , 刘建泉 , 汪有奎 (2008 ) 甘肃祁连山国家级自然保护区综合科学考察报告. 甘肃科学技术出版社 , 兰州.]
[54]
Ye JF Chen ZD Liu B Qin HN Yang Y 2012 ) Disjunct distribution of vascular plants between southwestern area and Taiwan area in China.
Biodiversity Science , 20 , 482 -494 .
[本文引用: 1]
(in Chinese with English abstract) [叶建飞 , 陈之端 , 刘冰 , 覃海宁 , 杨永 (2012 ) 中国西南与台湾地区维管植物的间断分布格局及形成机制. 生物多样性,
20 , 482 -494 .]
[本文引用: 1]
[55]
Ying LX Liu H Chiu QA Chen TY Chen XY Luo SJ Shen ZH 2016 ) The phylogeography of Fagus hayatae (Fagaceae) in China: genetic isolation among populations.
Ecology & Evolution , 6 , 2805 -2816 .
[本文引用: 1]
[56]
Yu HB Zhang YL 2013 ) Advance in phylogeography of alpine plants in the Tibetan Plateau and adjacent region. Acta Botanica Boreali-Occidentalia Sinica
, 33 , 1268 -1278 .
[本文引用: 1]
(in Chinese with English abstract) [于海彬 , 张镱锂 (2013 ) 青藏高原及其周边地区高山植物谱系地理学研究进展. 西北植物学报,
33 , 1268 -1278 .]
[本文引用: 1]
[57]
Zhang HD 1980 ) Origin and development of flora in China.
Acta Scientiarum Naturalium Universitatis Sunyatseni , 1 , 89 -98 .
[本文引用: 1]
(in Chinese with English abstract) [张宏达 (1980 ) 华夏植物区系的起源与发展. 中山大学学报(自然科学版),
1 , 89 -98 .]
[本文引用: 1]
[58]
Zhang JQ Meng SY Allen GA Wen J Rao GY 2014 ) Rapid radiation and dispersal out of the Qinghai-Tibetan Plateau of an alpine plant lineage Rhodiola (Crassulaceae).
Molecular Phylogenetics and Evolution , 77 , 147 -158 .
[本文引用: 1]
[59]
Zhang WJ Lu QY Liang J Shen ZH 2010 ) Altitudinal gradients of species richness and range size of vascular plants in Taiwan, a test of Rapoport’s rule.
Biodiversity Science , 18 , 312 -322 .
(in Chinese with English abstract) [张婉君 , 卢绮妍 , 梁军 , 沈泽昊 (2010 ) 台湾维管束植物物种丰富度和种域宽度的海拔格局及Rapoport法则检验. 生物多样性,
18 , 312 -322 .]
[60]
Zhang XS 1994 ) The basic ecological geographical types of altitudinal spectrum of mountain vegetation in China. In: Studies of Vegetation Ecology, Memorial of Professor Xueyu Hou, the Famous Ecologist (eds Jiang S, Chen CD) , pp. 250 -270. Science Press, Beijing.
[本文引用: 1]
(in Chinese) [张新时 (1994 ) 中国山地植被垂直带的基本生态地理类型. 植被生态学研究——纪念著名生态学家侯学煜教授(姜恕 , 陈昌笃主编), 250-270页, 科学出版社, 北京.]
[本文引用: 1]
[61]
Zheng Z Gong DJ Zhang Q Zhao HB 2014 ) Vertical patterns of plant species diversity in the Baishuijiang Nature Reserve, explanation of area, climate and boundary constraint.
Chinese Journal of Applied Ecology , 25 , 3390 -3398 .
(in Chinese with English abstract) [郑智 , 龚大洁 , 张乾 , 赵海斌 (2014 ) 白水江自然保护区植物物种多样性的垂直格局、面积、气候、边界限制的解释. 应用生态学报,
25 , 3390 -3398 .]
[62]
Zheng Z 1999 ) Response of altitudinal belts of vegetation to the late Quaternary climate changes in tropic Asia.
Geographic Research , 18 , 96 -102 .
[本文引用: 1]
(in Chinese with English abstract) [郑卓 (1999 ) 亚洲热带山地植被垂直带对晚第四纪气候变化的响应. 地理研究,
18 , 96 -102 .]
[本文引用: 1]
[63]
Zhu H 2016 ) A biogeographical comparison between Yunnan, Southwest China, and Taiwan, Southeast China, with impli- cations for the evolutionary history of the East Asian Flora.
Annals of the Missouri Botanical Garden , 101 , 750 -771 .
Arctic and alpine plant life cycles.
2
1971
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
2
2003
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
2
2003
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
Looking beyond the mountain, dispersal barriers in a changing world.
2
2016
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
Arctic and alpine biodiversity, patterns, causes and ecosystem consequences.
2
1994
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
2
2014
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
2
2014
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
Species turn- over of amphibians and reptiles in eastern China, disentangling the relative effects of geographic distance and environmental difference.
2
2011
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 ).但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 ). ...
中国西南地区与台湾种子植物间断分布现象. 植物学报,
2
2012
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
中国西南地区与台湾种子植物间断分布现象. 植物学报,
2
2012
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
中国阿尔金山自然保护区植物区系组成及基本特征. 新疆师范大学学报(自然科学版),
1
1993
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
中国阿尔金山自然保护区植物区系组成及基本特征. 新疆师范大学学报(自然科学版),
1
1993
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
中国第四纪冰期历史、特征及成因探讨. 第四纪研究,
2
2011
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
中国第四纪冰期历史、特征及成因探讨. 第四纪研究,
2
2011
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
Shuttle radar topography mission produces a wealth of data. Eos,
2
2000
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 根据上述14座山地的高山树线和永久雪线, 在Google Earth上运用“添加-多边形”工具勾画各个山地高山带分布范围的多边形(polygon), 即高山树线以上、永久雪线以下的地带范围(图1 ).将数据存为.kmz文件, 在地理信息系统软件ArcGIS 10.2 (www.esri.com/software/arcgis/)中转换成.shp格式.利用美国国家航空航天局发布的3″分辨率全球数字高程模型(SRTM v4.1, http://datamirror.csdb.cn/)截取各山地高山带范围, 应用Asia North Albers Equal Area Conic投影后重采样成分辨率100 m的研究区数字高程模型(Farr & Kobrick, 2000 ; van Zyl, 2001 ), 据此提取每座高山的最高海拔, 并计算了各山地高山区域的投影面积. ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报,
1
2006
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报,
1
2006
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
雅砻江和大渡河干旱河谷植被物种多样性比较: 气候、地形与空间的影响. 生物多样性,
2
2016
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 采用Mantel检验和偏Mantel检验来评估地理和环境距离对植物区系构成差异的影响(韩杰等, 2016 ).选择各山地高山带分布范围的几何中心经纬度坐标代表该区地理位置, 选择高山树线海拔(m)、山体最大海拔(m)和地形投影面积(km2 )作为地理距离相关变量; 选用高山树线附近的平均气候状况, 包括年均温(mean annual temperature, MAT, ℃)、最冷月最低温(minimum temperature of coldest month, MTCM, ℃)、最热月最高温(maximum temperature of coldest month, MTWM, ℃)、年降水量(mean annual precipitation, MAP, mm)来代表各山地高山带的气候条件.用欧几里得距离计算各高山带之间的地理空间距离和环境距离, 用Jaccard相异性系数计算各高山植物区系之间的相异性, 用偏Mantel检验来评估空间距离和环境差异的独立贡献. ...
雅砻江和大渡河干旱河谷植被物种多样性比较: 气候、地形与空间的影响. 生物多样性,
2
2016
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 采用Mantel检验和偏Mantel检验来评估地理和环境距离对植物区系构成差异的影响(韩杰等, 2016 ).选择各山地高山带分布范围的几何中心经纬度坐标代表该区地理位置, 选择高山树线海拔(m)、山体最大海拔(m)和地形投影面积(km2 )作为地理距离相关变量; 选用高山树线附近的平均气候状况, 包括年均温(mean annual temperature, MAT, ℃)、最冷月最低温(minimum temperature of coldest month, MTCM, ℃)、最热月最高温(maximum temperature of coldest month, MTWM, ℃)、年降水量(mean annual precipitation, MAP, mm)来代表各山地高山带的气候条件.用欧几里得距离计算各高山带之间的地理空间距离和环境距离, 用Jaccard相异性系数计算各高山植物区系之间的相异性, 用偏Mantel检验来评估空间距离和环境差异的独立贡献. ...
Very high resolution interpolated climate surfaces for global land areas.
2
2005
... The basic characteristics of the 14 mountains studied
山名 省份 最高峰 纬度范围 树线海拔Alpine 林线树种下限 高山林线树种 高山带面积 天山 Mt. Tianshan[1 ] 新疆 Xinjiang 7,439 42°18'-44°15' 2,800 1,800 雪岭云杉 Picea schrenkiana 832,930.0 阿尔金山 Mt. Arjin[2 ] 新疆 Xinjiang 6,973 37°55'-39°35' 3,500 — 无 3,137.6 祁连山 Mt. Qilian[3 ] 青海 Qinghai 5,808 37°16'-39°19' 3,600 2,600 祁连圆柏 Sabina przewalskii 16,345.1 贺兰山 Mt. Helan[4 ] 宁夏 Ningxia 3,556 38°07'-39°30' 3,200 2,600 青海云杉 P. erassifolia 755.5 长白山 Mt. Changbai[5 ] 吉林 Jilin 2,750 41°42'-42°51' 2,100 1,800 岳桦 Betula ermanii 9,063.4 五台山 Mt. Wutai[6 ] 河北 Hebei 3,061 38°50'-39°05' 2,800 2,300 华北落叶松 Larix principis-rupprechtii 923.7 太白山 Mt. Taibai[7 ] 陕西 Shaanxi 3,767 33°49'-34°08' 3,400 2,800 太白红杉 L. chinensis 2,678.7 摩天岭 Mt. Motianling[8 ] 甘肃 Gansu 3,837 32°39'-32°49' 3,450 2,900 秦岭冷杉 Abies chensiensis 5,698.9 贡嘎山 Mt. Gongga[9 ] 四川 Sichuan 7,556 29°20'-30°20' 3,800 2,700 峨眉冷杉 A. fabri 68,413.9 高黎贡山 Mt. Gaoligong[10 ] 云南 Yunnan 5,128 24°40'-28°30' 4,000 3,000 急尖长苞冷杉 A. georgei var. smithii 93,695.2 玉龙雪山 Mt. Yulong[11 ] 云南 Yunnan 5,596 27°10'-27°15' 3,900 3,200 长苞冷杉 A. georgei 5,628.1 轿子雪山 Mt. Jiaozi [12 ] 云南 Yunnan 4,247 26°00'-26°10' 4,000 3,200 急尖长苞冷杉 A. georgei var. smithii 294.3 台湾山脉 Mts. Taiwan[13 ] 台湾 Taiwan 3,998 22°36'-24°26' >3,998 3,000 台湾冷杉 A. kawakamii 3,277.2 珠峰 Mt. Zhumulangma# 西藏 Tibet 8,848 27°48'-29°19' 4,300 3,600 糙皮桦 B. albo-sinensis var. septentrionalis 231,943.0
[1]李利平等(2011); [2]崔乃然, 彼得·爱克夫(1993); [3]杨全生等(2008); [4]梁存柱(2012); [5]曹伟和李冀云(2003); [6]曹艳丽(1999); [7]任毅(2006); [8]郑智等(2014); [9]沈泽昊等(2004); [10]李恒等(2000); [11]冯建孟等(2006); [12]彭华和刘恩德(2015); [13]张婉君等(2010); # 未发表数据. ...
... 同时, 用各山地高山区的数字高程模型数据, 从世界气候数据库(www.worldclim.org/, WorldClim v1.4)中, 提取每个高山区的不同气候指标数据(BIO1-19).该数据库利用全球各地气象站记录的气象观测资料, 采用薄片样条函数模型插值生成全球气候栅格数据(1950-2000年平均值), 在WPS1984投影下的空间分辨率为1 km (Hijmans et al, 2005 ). ...
Patterns of beta diversity in Europe, the role of climate, land cover and distance across scales.
1
2012
... 岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 ).但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 ). ...
A world-wide study of high altitude treeline temperatures.
1
2004
... 另一方面, 面积大小和生境的异质性是岛屿承载不同生态属性物种能力的决定因子, 并决定了岛屿物种更替的灭绝速率(Russell et al, 2006 ; Si et al, 2014 ).因此, 岛屿面积和海拔范围的差异与岛屿物种构成的差异具有正相关性的结果同样适用于高山带.全球高山树线的海拔高度非常一致地对应于生长季均温(约6.7℃)等值线(Körner & Paulsen, 2004 ), 表明生长季的能量积累是山地高山带植物分布的主要限制因子.本文数据表明, 最热月最高温比其他气候指标对高山带植物区系相似性的影响更大, 主要的原因可能正是特定山地高山带的平均最热月最高温与高山带范围大小之间存在内在的关联. ...
Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Springer, Berlin.
2
2003
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
... 高山带是同时受到山地之间的几何距离、沿海拔梯度的非生物环境距离以及宽阔的山地森林带造成的生物竞争性距离等多重隔离的岛屿型非森林生境, 彼此之间物种迁移受隔离的程度应不亚于真正的岛屿.同时高山带是极端气候下形成的高度特化的生态系统, 彼此之间在气候环境上的相似性更为明显, 包括巨大的日温差、高辐射、稀薄空气和大风等(Körner, 2003 ).本文分析的14座山地高山带之间的两两平均几何距离在16.836个经纬度, 即 >1,000 km, 由此带来的隔离效应强于环境距离是可以理解的. ...
Long-distance dispersal vs vicariance, the origin and genetic diversity of alpine plants in the Spanish Sierra Nevada.
1
2006
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
Classification of Taiwan forest vegetation.
1
2013
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
1
2000
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
1
2000
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
新疆野生动植物物种丰富度的分布格局. 干旱区研究,
1
2011
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
新疆野生动植物物种丰富度的分布格局. 干旱区研究,
1
2011
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
1
2012
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
1
2012
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
Mountains of Southern China as “plant museums” and “plant cradles”, evolutionary and conservation insights.
1
2011
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
2
1999
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
2
1999
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
Evolutionary history of the subnival flora of the Himalaya-Hengduan Mountains, first insights from comparative phylogeography of four perennial herbs.
1
2016
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
色季拉山区高山寒带种子植物区系研究. 武汉植物研究,
2006
色季拉山区高山寒带种子植物区系研究. 武汉植物研究,
2006
1
1967
... 岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 ).但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 ). ...
The Birds of the Pine-oak Woodland in Southern Arizona and Adjacent Mexico.
1
1957
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
Birds on islands in the sky, origin of the montane avifauna of Northern Melanesia. Proceedings of the National Academy of Sciences,
1
1976
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
Differentiation and dispersal in alpine floras.
1
1974
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
Disentangling the effects of geographic distance and environmental dissimilarity on global patterns of species turnover.
1
2012
... 岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 ).但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 ). ...
1
2006
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
1
2006
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
Breeding birds on small islands, island biogeography or optimal foraging?
1
2006
... 另一方面, 面积大小和生境的异质性是岛屿承载不同生态属性物种能力的决定因子, 并决定了岛屿物种更替的灭绝速率(Russell et al, 2006 ; Si et al, 2014 ).因此, 岛屿面积和海拔范围的差异与岛屿物种构成的差异具有正相关性的结果同样适用于高山带.全球高山树线的海拔高度非常一致地对应于生长季均温(约6.7℃)等值线(Körner & Paulsen, 2004 ), 表明生长季的能量积累是山地高山带植物分布的主要限制因子.本文数据表明, 最热月最高温比其他气候指标对高山带植物区系相似性的影响更大, 主要的原因可能正是特定山地高山带的平均最热月最高温与高山带范围大小之间存在内在的关联. ...
The influence of late Quaternary climate-change velocity on species endemism.
1
2011
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
贡嘎山东坡植物区系的垂直分布格局. 生物多样性,
1
2004
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
贡嘎山东坡植物区系的垂直分布格局. 生物多样性,
1
2004
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
中国亚热带地区植物区系地理成分及其空间格局的数量分析. 植物分类学报,
4
2000
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 《中国种子植物区系地理》(吴征镒等, 2010 ) 一书已经对中国的种子植物区系起源、成分结构、全球地位进行了全面总结, 并提出了全国尺度的植物区系区划方案.已有的关于植物区系地理成分的分布格局的研究主要是以行政区域(一般为省)或特定自然地理区划单元(如自然地理区)为对象进行分析的, 尽管区域间植物区系成分构成的比较是植物区系地理研究的一种重要途径(沈泽昊和张新时, 2000 ), 但针对山地、平原、高原、湖泊等自然景观单元的植物区系比较往往局限在有限的区域范围内, 并不能有效地检验中国植物区系区划系统的合理性.本研究首次在全国尺度上对高山植物区系的地理成分进行了区域比较和宏观格局分析. ...
... 本文结果表明, 位于泛北极植物区中的天山高山带种子植物区系的确表现出与其余山地高山植物区系最大的平均差异; 青藏高原周缘山地的高山植物区系之间存在着较为密切的联系, 同处中国-日本植物区的北方山地如长白山、五台山、太白山之间也存在很高的区系相似性.此外, 古地中海区系成分随经度增加而迅速降低并在贺兰山以东的北方山地不再出现.这些格局特征从高山植物区系的角度显示了不同区系地理成分的区域分异及其边界特征, 为中国植物区系区划系统提供了支持.另外, 本文数据还表明, 在散布全国的14座山地高山植物区系中, 中国特有分布属的比例达到5.2%, 相比于中国植物区系地理成分整体构成而言(沈泽昊和张新时, 2000 )是一个相当高的比例, 表明高山带局部发生的物种形成十分旺盛.包括中国特有分布在内的东亚特有成分百分比在高山带呈现出自南向北递减、东西方向上中部隆起的分布格局, 与全国整体和低海拔地区的格局完全吻合.显示出中国西南山地对于中国特有植物分布和东亚植物区系发育的重要意义. ...
中国亚热带地区植物区系地理成分及其空间格局的数量分析. 植物分类学报,
4
2000
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 《中国种子植物区系地理》(吴征镒等, 2010 ) 一书已经对中国的种子植物区系起源、成分结构、全球地位进行了全面总结, 并提出了全国尺度的植物区系区划方案.已有的关于植物区系地理成分的分布格局的研究主要是以行政区域(一般为省)或特定自然地理区划单元(如自然地理区)为对象进行分析的, 尽管区域间植物区系成分构成的比较是植物区系地理研究的一种重要途径(沈泽昊和张新时, 2000 ), 但针对山地、平原、高原、湖泊等自然景观单元的植物区系比较往往局限在有限的区域范围内, 并不能有效地检验中国植物区系区划系统的合理性.本研究首次在全国尺度上对高山植物区系的地理成分进行了区域比较和宏观格局分析. ...
... 本文结果表明, 位于泛北极植物区中的天山高山带种子植物区系的确表现出与其余山地高山植物区系最大的平均差异; 青藏高原周缘山地的高山植物区系之间存在着较为密切的联系, 同处中国-日本植物区的北方山地如长白山、五台山、太白山之间也存在很高的区系相似性.此外, 古地中海区系成分随经度增加而迅速降低并在贺兰山以东的北方山地不再出现.这些格局特征从高山植物区系的角度显示了不同区系地理成分的区域分异及其边界特征, 为中国植物区系区划系统提供了支持.另外, 本文数据还表明, 在散布全国的14座山地高山植物区系中, 中国特有分布属的比例达到5.2%, 相比于中国植物区系地理成分整体构成而言(沈泽昊和张新时, 2000 )是一个相当高的比例, 表明高山带局部发生的物种形成十分旺盛.包括中国特有分布在内的东亚特有成分百分比在高山带呈现出自南向北递减、东西方向上中部隆起的分布格局, 与全国整体和低海拔地区的格局完全吻合.显示出中国西南山地对于中国特有植物分布和东亚植物区系发育的重要意义. ...
台湾的生物地理. 1. 一些初步思考和研究. 台湾省立博物馆年刊,
1
1997
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
台湾的生物地理. 1. 一些初步思考和研究. 台湾省立博物馆年刊,
1
1997
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
1
2006
... 参照Flora of China 的植物分类系统, 对所研究的14座山地的高山植物物种名录进行校核, 得到中国高山植物区系的总物种名录.本文分析的高山植物物种数据部分来自各合作者的野外植物调查结果, 另有部分山地的数据来自公开出版物, 具体说明见表1 .物种属性数据包括物种的拉丁科属种名、生活型及所在属的地理分布区类型, 分布区类型依据吴征镒(1991).根据定义, 高山带植物应分布在高山树线以上, 但很多高山植物可以向下分布到中山森林带.在第四纪冰期, 青藏高原腹地以外的中国植被-气候带海拔普遍下移500-1,500 m(郑卓, 1999 ; 施雅风等, 2006 ).随着冰后期气候变暖, 物种分布范围上升, 但根据野外调查观测, 大量物种的分布下限当前仍滞留在高山树线以下.因此, 本文中采取统一标准选择高山物种, 即其分布的上限高度显著高于当前树线的海拔(>100 m), 其下限不低于亚高山针叶林, 即所在山地冷杉(Abies spp.)林和落叶松(Larix spp.)林的下限海拔, 建立的物种名录不包括树线树种和少数海拔分布极宽的植物种, 如车前(Plantago spp.)等.因为前者本来就属于亚高山分布, 后者作为广布种对高山环境完全没有指示意义和山体之间的区分作用. ...
1
2006
... 参照Flora of China 的植物分类系统, 对所研究的14座山地的高山植物物种名录进行校核, 得到中国高山植物区系的总物种名录.本文分析的高山植物物种数据部分来自各合作者的野外植物调查结果, 另有部分山地的数据来自公开出版物, 具体说明见表1 .物种属性数据包括物种的拉丁科属种名、生活型及所在属的地理分布区类型, 分布区类型依据吴征镒(1991).根据定义, 高山带植物应分布在高山树线以上, 但很多高山植物可以向下分布到中山森林带.在第四纪冰期, 青藏高原腹地以外的中国植被-气候带海拔普遍下移500-1,500 m(郑卓, 1999 ; 施雅风等, 2006 ).随着冰后期气候变暖, 物种分布范围上升, 但根据野外调查观测, 大量物种的分布下限当前仍滞留在高山树线以下.因此, 本文中采取统一标准选择高山物种, 即其分布的上限高度显著高于当前树线的海拔(>100 m), 其下限不低于亚高山针叶林, 即所在山地冷杉(Abies spp.)林和落叶松(Larix spp.)林的下限海拔, 建立的物种名录不包括树线树种和少数海拔分布极宽的植物种, 如车前(Plantago spp.)等.因为前者本来就属于亚高山分布, 后者作为广布种对高山环境完全没有指示意义和山体之间的区分作用. ...
Turnover of breeding bird communities on islands in an inundated lake.
2
2014
... 岛屿面积、生境异质性和距离隔离是岛屿物种丰富度的决定因子(MacArthur & Wilson, 1967 ), 并且通过物种的迁移拓殖和局部灭绝过程来影响岛屿之间物种构成的更替(Qian & Ricklefs, 2012 ).但是, 不同研究对这几个因子影响作用的相对大小有不同的认识, 并发现研究范围的尺度、岛屿生境之间的连通性、对象的生活史特征等都是潜在的影响因素(Chen et al, 2011 ; Keil et al, 2012 ; Si et al, 2014 ). ...
... 另一方面, 面积大小和生境的异质性是岛屿承载不同生态属性物种能力的决定因子, 并决定了岛屿物种更替的灭绝速率(Russell et al, 2006 ; Si et al, 2014 ).因此, 岛屿面积和海拔范围的差异与岛屿物种构成的差异具有正相关性的结果同样适用于高山带.全球高山树线的海拔高度非常一致地对应于生长季均温(约6.7℃)等值线(Körner & Paulsen, 2004 ), 表明生长季的能量积累是山地高山带植物分布的主要限制因子.本文数据表明, 最热月最高温比其他气候指标对高山带植物区系相似性的影响更大, 主要的原因可能正是特定山地高山带的平均最热月最高温与高山带范围大小之间存在内在的关联. ...
古地中海退却与喜马拉雅-横断山的隆起在中国喜马拉雅成分及高山植物区系的形成与发展上的意义. 云南植物研究,
2
2002
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
古地中海退却与喜马拉雅-横断山的隆起在中国喜马拉雅成分及高山植物区系的形成与发展上的意义. 云南植物研究,
2
2002
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
Wind dispersal of alpine plant species, a comparison with lowland species.
1
2009
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
中国第三纪植被与植物区系历史及分区. 植物分类学报,
1
1992
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
中国第三纪植被与植物区系历史及分区. 植物分类学报,
1
1992
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
The shuttle radar topography mission (SRTM), a breakthrough in remote sensing of topography.
1
2001
... 根据上述14座山地的高山树线和永久雪线, 在Google Earth上运用“添加-多边形”工具勾画各个山地高山带分布范围的多边形(polygon), 即高山树线以上、永久雪线以下的地带范围(图1 ).将数据存为.kmz文件, 在地理信息系统软件ArcGIS 10.2 (www.esri.com/software/arcgis/)中转换成.shp格式.利用美国国家航空航天局发布的3″分辨率全球数字高程模型(SRTM v4.1, http://datamirror.csdb.cn/)截取各山地高山带范围, 应用Asia North Albers Equal Area Conic投影后重采样成分辨率100 m的研究区数字高程模型(Farr & Kobrick, 2000 ; van Zyl, 2001 ), 据此提取每座高山的最高海拔, 并计算了各山地高山区域的投影面积. ...
1
1992
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
1
1992
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
雀儿山新路海自然保护区高山种子植物区系研究. 安徽农业科学,
1
2012
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
雀儿山新路海自然保护区高山种子植物区系研究. 安徽农业科学,
1
2012
... 中国的高山带主要分布在青藏高原及其周边, 以及北方和台湾主要山系, 具有气候寒冷、环境极端、交通不便等特点.因此, 相比其他区域, 针对高山带的植物区系地理研究仍相对缺乏.尤其是需要掌握足够数量的物种分布信息以获得相对可靠的植物分布范围数据, 这对于高山带植物区系比较研究更是一个极具挑战性的问题.但在中国, 已经有了一些针对特定山体或山脉的植物区系结构, 特别是区系地理成分的分析工作, 包括对天山(李利平等, 2011 )、贺兰山(梁存柱, 2012 )、秦岭太白山(任毅, 2006 )、五台山(曹艳丽, 1999(曹艳丽 (1999) 山西五台山高山林线的植被景观.北京大学硕士研究生毕业论文 ))、长白山(曹伟和李冀云, 2003 )及青藏高原周缘高山的研究(如罗建等, 2006 ; 李恒等, 2000 ; 王静, 2012 ).这些工作为开展山脉或山体为单元的宏观区域格局或比较研究提供了一定的基础, 但迄今不同山地之间高山带植物区系的比较却十分少见. ...
中国高等植物多样性. 生物多样性,
1
2015
... 根据统计, 本研究涉及的14座主要山地的高山带至少分布有种子植物65科489属3,670种(含种下单位).根据中国植物多样性的最新统计资料(王利松等, 2015 ), 这些高山植物大约占中国分布的种子植物总科数的24.9%、总属数的16.3%和总种数的12.3%.尽管高山带环境极端恶劣, 植被稀疏、结构简单, 但在这仅占全国陆地面积0.13% (投影面积约12,747.85 km2 ), 分散于14座高山的区域内所分布的植物多样性是非常高的. ...
中国高等植物多样性. 生物多样性,
1
2015
... 根据统计, 本研究涉及的14座主要山地的高山带至少分布有种子植物65科489属3,670种(含种下单位).根据中国植物多样性的最新统计资料(王利松等, 2015 ), 这些高山植物大约占中国分布的种子植物总科数的24.9%、总属数的16.3%和总种数的12.3%.尽管高山带环境极端恶劣, 植被稀疏、结构简单, 但在这仅占全国陆地面积0.13% (投影面积约12,747.85 km2 ), 分散于14座高山的区域内所分布的植物多样性是非常高的. ...
东亚植物区系的一些分布式样和迁移路线. 植物分类学报, 30, 1-24;
3
1992
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
... ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
东亚植物区系的一些分布式样和迁移路线. 植物分类学报, 30, 1-24;
3
1992
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
... ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
Crossing barriers in an extremely fragmented system, two case studies in the afro-alpine sky island flora.
1
2014
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
中国种子植物属的分布区类型. 云南植物研究,
1
1991
... 分别在科、属、种水平评估和比较各山地高山带的植物多样性.采用中国种子植物属地理分布区类型系统(吴征镒, 1991 )划分各山地高山带植物区系属的地理成分类型, 并分别统计不同分布型的属数及其所包含的种数.参照吴征镒等(2010), 根据不同地理成分之间的联系将世界分布型之外的类型进一步分为热带成分(T2-7)、北温带成分(T8- 11)、古地中海成分(T12-13)和东亚成分(T14-15).采用Pearson相关系数分析不同山地高山带植物区系之间的相似性, 包括分类群构成和区系成分构成的相似性. ...
中国种子植物属的分布区类型. 云南植物研究,
1
1991
... 分别在科、属、种水平评估和比较各山地高山带的植物多样性.采用中国种子植物属地理分布区类型系统(吴征镒, 1991 )划分各山地高山带植物区系属的地理成分类型, 并分别统计不同分布型的属数及其所包含的种数.参照吴征镒等(2010), 根据不同地理成分之间的联系将世界分布型之外的类型进一步分为热带成分(T2-7)、北温带成分(T8- 11)、古地中海成分(T12-13)和东亚成分(T14-15).采用Pearson相关系数分析不同山地高山带植物区系之间的相似性, 包括分类群构成和区系成分构成的相似性. ...
4
2010
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
... 《中国种子植物区系地理》(吴征镒等, 2010 ) 一书已经对中国的种子植物区系起源、成分结构、全球地位进行了全面总结, 并提出了全国尺度的植物区系区划方案.已有的关于植物区系地理成分的分布格局的研究主要是以行政区域(一般为省)或特定自然地理区划单元(如自然地理区)为对象进行分析的, 尽管区域间植物区系成分构成的比较是植物区系地理研究的一种重要途径(沈泽昊和张新时, 2000 ), 但针对山地、平原、高原、湖泊等自然景观单元的植物区系比较往往局限在有限的区域范围内, 并不能有效地检验中国植物区系区划系统的合理性.本研究首次在全国尺度上对高山植物区系的地理成分进行了区域比较和宏观格局分析. ...
... 在中国种子植物区系分区系统中, 天山属于泛北极植物区, 阿尔金山、祁连山和贺兰山则属于古地中海植物区; 台湾的高山带被明确划归古热带植物区, 本文所涉及的其他山地均属于东亚植物区, 其中长白山、五台山、太白山属于中国-日本森林亚区,西南山地包括摩天岭、贡嘎山、高黎贡山、玉龙雪山和轿子雪山均属于中国-喜马拉雅植物亚区, 珠峰地区则属于青藏高原亚区(吴征镒等, 2010 ). ...
... 值得指出的是, 目前基于植物区系地理成分构成进行的区系分区方案主要是针对较大区域进行的研究结果(吴征镒等, 2010 ).本文数据针对高山区特定范围的数据, 所得结果应是对既有成果的一种补充, 也是在不同尺度上的验证.我们发现台湾的高山带植物区系仍保留有鲜明而深远的泛北极和东亚北部区系成分的影响, 不应归于古热带植物区, 这一结果与Zhu (2016)的观点一致; 喜马拉雅山脉中段的珠峰地区高山带(尤其是南坡)植物区系也明显不同于青藏高原生物区; 天山而不是隔海相望的台湾高山带的植物区系与中国其他高山区的植物区系更加隔离.这些与既有认识不一致之处值得进一步探索验证. ...
4
2010
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
... 《中国种子植物区系地理》(吴征镒等, 2010 ) 一书已经对中国的种子植物区系起源、成分结构、全球地位进行了全面总结, 并提出了全国尺度的植物区系区划方案.已有的关于植物区系地理成分的分布格局的研究主要是以行政区域(一般为省)或特定自然地理区划单元(如自然地理区)为对象进行分析的, 尽管区域间植物区系成分构成的比较是植物区系地理研究的一种重要途径(沈泽昊和张新时, 2000 ), 但针对山地、平原、高原、湖泊等自然景观单元的植物区系比较往往局限在有限的区域范围内, 并不能有效地检验中国植物区系区划系统的合理性.本研究首次在全国尺度上对高山植物区系的地理成分进行了区域比较和宏观格局分析. ...
... 在中国种子植物区系分区系统中, 天山属于泛北极植物区, 阿尔金山、祁连山和贺兰山则属于古地中海植物区; 台湾的高山带被明确划归古热带植物区, 本文所涉及的其他山地均属于东亚植物区, 其中长白山、五台山、太白山属于中国-日本森林亚区,西南山地包括摩天岭、贡嘎山、高黎贡山、玉龙雪山和轿子雪山均属于中国-喜马拉雅植物亚区, 珠峰地区则属于青藏高原亚区(吴征镒等, 2010 ). ...
... 值得指出的是, 目前基于植物区系地理成分构成进行的区系分区方案主要是针对较大区域进行的研究结果(吴征镒等, 2010 ).本文数据针对高山区特定范围的数据, 所得结果应是对既有成果的一种补充, 也是在不同尺度上的验证.我们发现台湾的高山带植物区系仍保留有鲜明而深远的泛北极和东亚北部区系成分的影响, 不应归于古热带植物区, 这一结果与Zhu (2016)的观点一致; 喜马拉雅山脉中段的珠峰地区高山带(尤其是南坡)植物区系也明显不同于青藏高原生物区; 天山而不是隔海相望的台湾高山带的植物区系与中国其他高山区的植物区系更加隔离.这些与既有认识不一致之处值得进一步探索验证. ...
Barriers and bridges, early human dispersals in equatorial SE Asia. Geology and Archaeology, Submerged Landscapes of the Continental Shelf (eds Harff J, Bailey G, Lüth F),
1
2014
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
1
2013
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
1
2013
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
中国西南与台湾地区维管植物的间断分布格局及形成机制. 生物多样性,
1
2012
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
中国西南与台湾地区维管植物的间断分布格局及形成机制. 生物多样性,
1
2012
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
The phylogeography of Fagus hayatae (Fagaceae) in China: genetic isolation among populations.
1
2016
... 但是, 针对高山植物区系的分析也揭示了新的区系地理特征.首先, 在14座山地高山植物区系的属级分类单元中均缺乏中国-日本植物区系成分.这也显示了中国-日本分布成分不属于高山分布型植物.其次, 属于中国-日本植物区的长白山、五台山和太白山以及古地中海植物区东部边界的贺兰山在其高山植物区系中全部缺少属级的热带分布成分, 而中国-喜马拉雅植物区和古地中海植物区的诸山地的高山植物区系中却还能找到与热带植物区系的微弱联系.第三, 古地中海植物区系的成分普遍出现于中国-喜马拉雅植物区的山地中, 有力支持了古地中海植物区系与中国-喜马拉雅植物区系的深远联系(孙航, 2002 ).第四, 也是最为突出的一点, 划归古热带的台湾高山植物区系与中国北方山地的高山植物区系联系反而强于其与处在近似纬度的西南山地的高山植物区系.尽管基于特定山地植物区系种级地理成分构成的研究(沈泽昊等, 2004 )、区域植物区系比较研究(陈之端等, 2012 )及特定物种谱系地理研究的结果(Ying et al, 2016 )都证实, 存在着从西南山地经秦岭和大巴山-大别山向东的区系联系通道(王文采, 1992 ), 但本文结果表明对于高山植物区系而言, 中国东部地区南北方向的植物区系联系强度大于东西向的区系联系.这一结果为第四纪冰期/间冰期循环驱动中国东部南北向植物区系联系, 包括实现岛屿与大陆之间物种扩散交流的机制性假说提供了新的佐证(王文采, 1992 ; 沈中桴, 1997 ; 叶建飞等, 2012 ). ...
青藏高原及其周边地区高山植物谱系地理学研究进展. 西北植物学报,
1
2013
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
青藏高原及其周边地区高山植物谱系地理学研究进展. 西北植物学报,
1
2013
... 山地被称为生物迁移的障碍或桥梁, 这取决于生物扩散方向与山脉延伸方向之间的关系(Wurster & Bird, 2014 ; Caplat et al, 2016 ).如中国青藏高原-华北山地-白令海峡-北美西部的落基山脉就被认为是东亚与北美之间植物区系联系的桥梁(王文采, 1992 ; 路安民, 1999 ), 而横断山区的平行山脉对很多植物种分布产生了明显的隔离作用(如Luo et al, 2016 ).大量研究表明, 山地是生物多样性分布的热点区域, 其复杂多样的环境提供了丰富且相对稳定的生态位, 成为变化环境中的生物避难所和博物馆(López-Pujol et al, 2011 ; Sandel et al, 2011 ); 同时, 山地特别是相对隔离的高山带特殊环境中具有较为强烈的隔离分化和成种过程(孙航, 2002 ; 于海彬和张镱锂, 2013 ; Luo et al, 2016 ).然而, 在宏观区域尺度上, 山地之间的空间距离和延伸走向构成的空间格局对植物分布的总体影响, 以及这种影响在区域植物区系构成中的反映还很少有研究(沈泽昊和张新时, 2000 ).目前, 中国高山植物区系的宏观分异格局尚不清楚, 研究空间距离的区系隔离效应和山脉延伸走向的区系联系效应, 对于理解高山植物扩散的宏观途径和区域山地格局对植物区系发育过程的影响具有重要意义. ...
华夏植物区系的起源与发展. 中山大学学报(自然科学版),
1
1980
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
华夏植物区系的起源与发展. 中山大学学报(自然科学版),
1
1980
... 植物区系地理是研究全球或某一区域所有植物的种类组成及其现代与过去的分布, 并探讨其起源和演化历史的科学(王荷生, 1992 ).植物区系地理通过系统分析不同植物类群的地理分布规律和类群间差异, 进行植物区系成分分类和区系分区, 为进一步从历史、环境和进化的途径探索植物区系演变的时空格局与机制奠定了宏观基础(张宏达, 1980 ; 陶君容, 1992 ).在总结中国3,116个种子植物属的地理分布范围基础上, 吴征镒(1991)把中国种子植物区系归纳为15个地理分布区类型和31个亚型, 首次为中国植物区系地理的量化和比较研究提供了一个分析模板, 在过去20年里极大地推动了中国境内的植物区系地理研究(路安民, 1999 ; 沈泽昊和张新时, 2000 ; 严岳鸿等, 2013 ; 陈灵芝, 2014 ).植物区系地理成分分类体系的建立为植物多样性的构成分析提供了一个独特的框架与途径, 也建立了一个基于自然环境时空分异的宏观机制性假说.然而, 正如吴征镒等(2010)指出, 针对这一业已建立的中国植物区系地理分类与分区体系, 还有待针对特定的类群或区域, 从历史、环境和进化角度进行验证及不同尺度的深入研究. ...
Rapid radiation and dispersal out of the Qinghai-Tibetan Plateau of an alpine plant lineage Rhodiola (Crassulaceae).
1
2014
... 高山带(alpine zone)是全球气候和植被分类体系中具有鲜明判识标志的分类单元, 分布在亚高山森林边界(即高山树线过渡带, alpine treeline ecotone)以上、永久雪线以下, 以非乔木生活型为特征的带状植物分布区域(Chapin III & Körner, 1994 ).由于低温、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响, 高山植物普遍表现了与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性(Bliss, 1971 ; Körner, 2003 ).高山带植被在众多海拔数千米的高山上部呈岛屿状分布, 被称为“天岛” (sky islands) (Marshall, 1957 ).“天岛”之间的空间距离和海拔梯度上陡峭的、气候主导的环境变化, 构成了高山之间动、植物物种隔离分布、遗传分化成种以至区系分化的强烈驱动力(Mary & Diamond, 1976 ; Kropf et al, 2006 ; Zhang et al, 2014 ); 另一方面, 适应高山环境的植物也通过各种传播途径实现了“天岛”之间的迁移扩散和基因交流(Packer, 1974 ; Tackenberg & Stöcklin, 2009 ; Wondimu et al, 2014 ), 并形成了具有独特地理分布的高山带植物群.这群植物的地理分布特征在吴征镒(1991)提出的中国种子植物属分布区类型体系中得到印证, 即北极-高山分布型(8-2型)的独特存在. ...
台湾维管束植物物种丰富度和种域宽度的海拔格局及Rapoport法则检验. 生物多样性,
2010
台湾维管束植物物种丰富度和种域宽度的海拔格局及Rapoport法则检验. 生物多样性,
2010
1
1994
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
1
1994
... 选择国内具有高山环境的14座主要山地, 见图1 .青藏高原及周缘山地以东的地区地势相对低平, 仅有少数孤立的山峰达到海拔3,000 m以上高程.结合第四纪冰期气候的变化历史, 一般认为除长白山、五台山和太白山以外中国大陆东部缺少典型的高山环境(张新时, 1994 ; 崔之久等, 2011 ).台湾山脉最高峰达到3,997 m, 但现代森林的分布上限达到海拔3,800 m以上(Li et al, 2013 ), 故台湾高山带的存在颇具争议(赖明洲, 2003 ), 中国种子植物区系也将台湾的高山植被带明确划归古热带植物区(吴征镒等, 2010 ).本研究将台湾高山植物树线以上的植物区系数据纳入分析, 通过与其他高山带植物区系的比较, 以期获得新的证据. ...
白水江自然保护区植物物种多样性的垂直格局、面积、气候、边界限制的解释. 应用生态学报,
2014
白水江自然保护区植物物种多样性的垂直格局、面积、气候、边界限制的解释. 应用生态学报,
2014
亚洲热带山地植被垂直带对晚第四纪气候变化的响应. 地理研究,
1
1999
... 参照Flora of China 的植物分类系统, 对所研究的14座山地的高山植物物种名录进行校核, 得到中国高山植物区系的总物种名录.本文分析的高山植物物种数据部分来自各合作者的野外植物调查结果, 另有部分山地的数据来自公开出版物, 具体说明见表1 .物种属性数据包括物种的拉丁科属种名、生活型及所在属的地理分布区类型, 分布区类型依据吴征镒(1991).根据定义, 高山带植物应分布在高山树线以上, 但很多高山植物可以向下分布到中山森林带.在第四纪冰期, 青藏高原腹地以外的中国植被-气候带海拔普遍下移500-1,500 m(郑卓, 1999 ; 施雅风等, 2006 ).随着冰后期气候变暖, 物种分布范围上升, 但根据野外调查观测, 大量物种的分布下限当前仍滞留在高山树线以下.因此, 本文中采取统一标准选择高山物种, 即其分布的上限高度显著高于当前树线的海拔(>100 m), 其下限不低于亚高山针叶林, 即所在山地冷杉(Abies spp.)林和落叶松(Larix spp.)林的下限海拔, 建立的物种名录不包括树线树种和少数海拔分布极宽的植物种, 如车前(Plantago spp.)等.因为前者本来就属于亚高山分布, 后者作为广布种对高山环境完全没有指示意义和山体之间的区分作用. ...
亚洲热带山地植被垂直带对晚第四纪气候变化的响应. 地理研究,
1
1999
... 参照Flora of China 的植物分类系统, 对所研究的14座山地的高山植物物种名录进行校核, 得到中国高山植物区系的总物种名录.本文分析的高山植物物种数据部分来自各合作者的野外植物调查结果, 另有部分山地的数据来自公开出版物, 具体说明见表1 .物种属性数据包括物种的拉丁科属种名、生活型及所在属的地理分布区类型, 分布区类型依据吴征镒(1991).根据定义, 高山带植物应分布在高山树线以上, 但很多高山植物可以向下分布到中山森林带.在第四纪冰期, 青藏高原腹地以外的中国植被-气候带海拔普遍下移500-1,500 m(郑卓, 1999 ; 施雅风等, 2006 ).随着冰后期气候变暖, 物种分布范围上升, 但根据野外调查观测, 大量物种的分布下限当前仍滞留在高山树线以下.因此, 本文中采取统一标准选择高山物种, 即其分布的上限高度显著高于当前树线的海拔(>100 m), 其下限不低于亚高山针叶林, 即所在山地冷杉(Abies spp.)林和落叶松(Larix spp.)林的下限海拔, 建立的物种名录不包括树线树种和少数海拔分布极宽的植物种, 如车前(Plantago spp.)等.因为前者本来就属于亚高山分布, 后者作为广布种对高山环境完全没有指示意义和山体之间的区分作用. ...
A biogeographical comparison between Yunnan, Southwest China, and Taiwan, Southeast China, with impli- cations for the evolutionary history of the East Asian Flora.
2016
 , 杨明正
, 杨明正

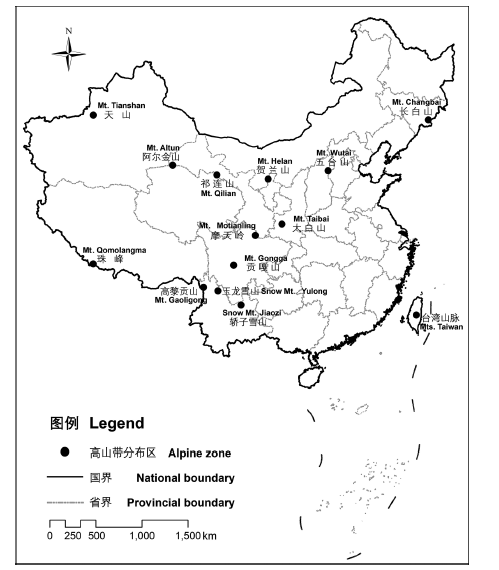
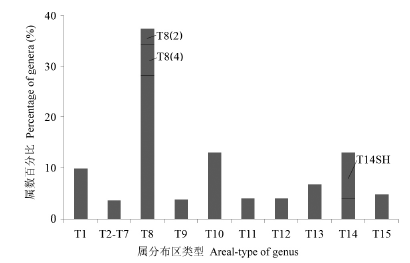
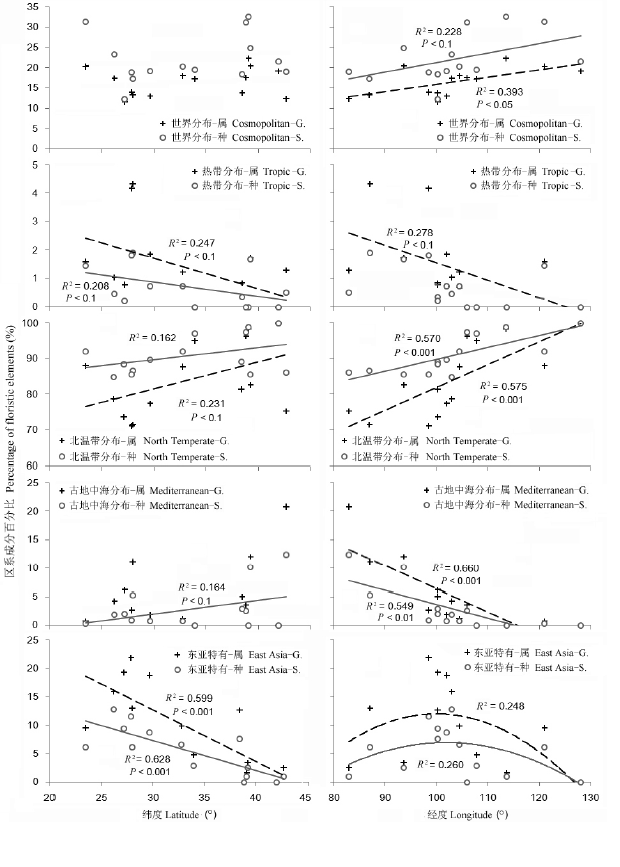
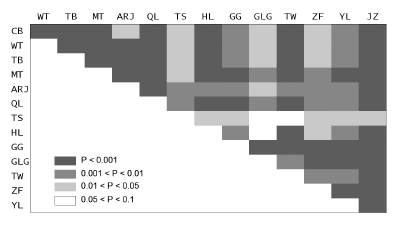
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}