物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 )。物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 )。物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义。影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 )。环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响。生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 )。在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种。而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 )。
内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 )。作为我国西北干旱区的重要组成部分, 该地区已有大量研究。如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响。但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少。在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识。本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局。在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响。
1 材料与方法
1.1 研究区概况
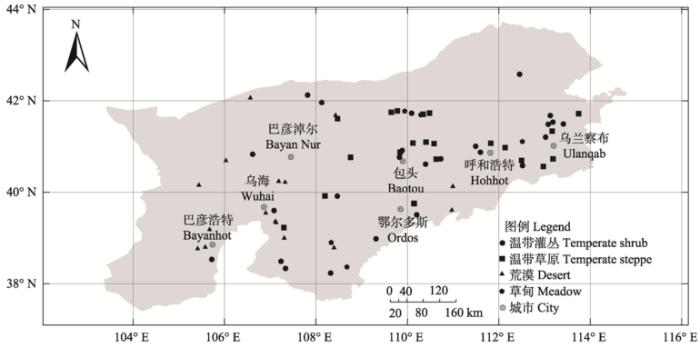
内蒙古西部包括乌兰察布、呼和浩特、包头、鄂尔多斯、巴彦淖尔、乌海和阿拉善7个盟市, 地理坐标为37°30°-42°47′ N, 95°05′-114°49′ E。该区以高原地貌为主, 平均海拔超过1,000 m。气候以温带大陆性季风气候为主, 年均温约-2℃至10℃, 年降水量约29-500 mm, 阿拉善高原的年降水量仅50- 150 mm, 而额济纳旗仅有36 mm (裴浩等, 2000 ), 且蒸发量相对较大。内蒙古西部日照充足, 大部分地区的年日照时数都超过2,700 h, 阿拉善高原西部地区的年日照时数超过3,400 h。该地区以温带荒漠和温带草原为主, 典型植物群落包括克氏针茅(Stipa krylovii )草原、短花针茅(S. breviflora )草原、油蒿(Artemisia ordosica )灌丛、藏锦鸡儿(Caragana tibetica )灌丛、红砂(Reaumuria songarica )和白刺(Nitraria tangutorum )灌丛等。
1.2 数据获取
根据中国植被图和实际考察情况, 分别于2012-2015年的7-8月选择代表性植物群落样地74个进行调查(图1 )。调查采用样方法, 群落类型通过优势种进行确定(吴征镒, 1980 )。样方的选取原则为尽量选择围封或远离城镇公路、人类活动干扰少的开阔地, 面积均为10 m × 10 m。对于灌木样地, 在样方4角设置4个面积为1 m × 1 m的草本样方。对灌木和草本进行分层调查, 调查内容为灌木和草本的种类、高度、盖度。对于草本样地, 在样方4角及中间设置5个面积为1 m × 1 m的小样方, 记录每个小样方中所有物种的种类、高度、盖度(方精云等, 2009 ), 最后统计每个物种在样地的出现次数以计算频度。
图1 内蒙古西部研究区样地分布图
Fig. 1 Distribution of sampling sites in western Inner Mongolia
各样地用土壤取样器采集土壤样品。采样深度为1 m, 按照0-10 cm、10-20 cm、20-30 cm、30- 50 cm、50-70 cm、70-100 cm分为6层取样, 各层样品装入封口聚乙烯塑料袋带回实验室。将样品自然风干, 研磨过筛后, 采用重铬酸钾氧化-外加热法测定有机碳含量(total organic carbon, TOC), 全氮(total nitrogen, TN)用凯氏定氮法测定, 全磷(total phosphorus, TP)采用碱熔-钼锑抗比色法测定; 使用pH计测定pH值。
从WorldClim数据库(http://www.worldclim.org/ )获取月均温和逐月降水量数据, 空间分辨率为30′′ (相当于赤道地区1 km)。年均温(mean annual temperature, MAT, ℃)、年平均降水量(mean annual precipitation, MAP, mm)、潜在蒸散量(potential evapotranspiration, PET, mm) (方精云等, 2009 )、最冷月均温(mean temperature of the coldest month, MTCM, ℃)根据月均温和逐月降水量数据计算 得到。
1.3 数据处理
群落的物种多样性是指群落中植物种类的丰富程度, 而物种多样性(α多样性)指标是度量物种多样性高低及空间分布特征的数值指标。近年来对植物群落物种多样性的研究采用的测度指数主要包括物种丰富度指数、物种多样性指数及均匀度指数, 其中, Patrick丰富度指数、Simpson指数、Shannon-Wiener指数和Pielou均匀度指数在测度物种多样性中较为常见, 与其他研究具有较大可比性(汪殿蓓等, 2001 )。本文也采用这4种多样性指数来表征研究区植物群落的物种多样性。在计算过程中, 首先根据每个样方中各灌木、草本物种的高度、盖度、频度计算每个物种的重要值(p i ) (Zhang et al, 2006 ):
p i = (相对高度 + 相对盖度 + 相对频度)/3 (1)
然后根据每个样方内各物种重要值计算群落的多样性指数。
Patrick丰富度指数 $R=S$ (2)
Simpson指数 $\lambda=-\sum^{s}_{i=1} p^2_{i}$ (3)
Shannon-Wiener指数 $H=-\sum^{s}_{i=1} p_{i} ln(p_i)$ (4)
Pielou均匀度指数 $E=\frac{H'}{ln(S)}$ (5)
其中, S 为每个样方内的物种数, p i 为第i 个物种的重要值(马克平等, 1995 )。
最后, 通过Pearson相关分析确定不同环境因子与群落物种多样性的关系, 采用多元回归分析确定影响群落物种多样性的主导因子。数据的初步处理利用Microsoft Excel 2007完成, 回归分析利用SPSS 20.0完成, 采用R 3.4.1绘制图形。
2 结果
2.1 内蒙古西部自然植被的群落特征
此次调查的植物群落可划分为4种植被型, 即温带灌丛、温带草原、荒漠和草甸(表1 )。内蒙古西部具有丰富的灌木资源, 在此次调查中, 27个灌丛样地被划分为14个灌木群落。该地区的灌丛性质差异明显, 在大青山东段以森林气候下的次生灌丛为主, 如虎榛子(Ostryopsis davidiana )灌丛和蒙古绣线菊(Spiraea mongolica )灌丛。在草原区则广泛分布着旱生的落叶灌丛, 灌木呈斑块状分布在草原基质上, 草本层以草原种类为主, 如典型草原区的小叶锦鸡儿(Caragana microphylla )灌丛和荒漠草原区的藏锦鸡儿灌丛。在荒漠草原和荒漠区的干旱山地上则生长着特有的蒙古扁桃(Amygdalus mongolica )灌丛, 而沙地柏(Juniperus sabina )灌丛、油蒿灌丛和西伯利亚白刺(Nitraria sibirica )灌丛等则在沙地、盐碱地等隐域生境上广泛分布。温带草原调查了10个群落类型共25个样地, 包括典型草原(克氏针茅草原)、荒漠草原(小针茅(Stipa klemenzii )草原、短花针茅草原等)、山地草原(羊茅(Festuca ovina )草原)及因放牧干扰而蒿属植物占优势的蒿类草原(冷蒿(Artemisia frigida )草原)。该地区荒漠类型多样, 西鄂尔多斯-阿拉善荒漠为我国八大特有植物中心之一, 分布着以古地中海孑遗植物为建群种的荒漠植被, 如四合木(Tetraena mongolica )荒漠、半日花(Helianthemum songaricum )荒漠和沙冬青(Ammopiptanthus mongolicus )荒漠(赵一之, 2006 ), 具有重要的保护价值。在该地区的隐域生境上也分布着较大面积的草甸植被, 此次调查到3个类型, 以芨芨草(Achnatherum splendens )草甸最为常见。
2.2 内蒙古西部自然植被的物种多样性特征
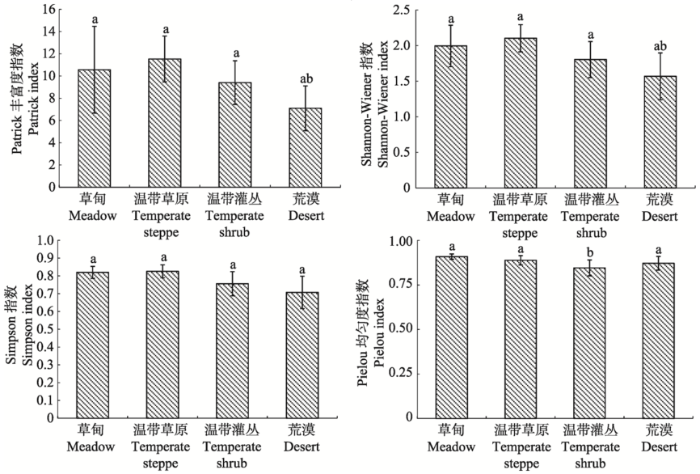
总体来说, 内蒙古西部自然植被的物种数较少,物种多样性偏低, 群落结构较为简单。物种多样性指数的变化趋势与物种数基本一致, 表现为物种数越多, 多样性指数越高(图2 )。温带草原和草甸的物种相对丰富, 但草甸的物种丰富度指数的标准差和变异系数都远大于草原, 这可能主要是因为草甸包含了一个薹草-杂类草草甸样地, 其丰富度指数高达26.20, 而剔除该样地后草甸的丰富度指数仅为7.66, 远低于草原。温带灌丛的物种多样性略低于温带草原和草甸, 但其变异系数高于温带草原, 这说明各灌木群落之间的物种多样性指数差异相对于温带草原来说较大。荒漠的物种多样性最低, 其Partrick丰富度指数(P = 0.003)和Shannon-Wiener指数(P = 0.001)与其他3种植被型均差异显著, 物种丰富度指数均值仅为7.11, 且其变异系数较大, 最大值出现在沙冬青荒漠(Patrick丰富度指数12.667)。而物种数最少且多样性指数最小的沙米(Agriophyllum squarrosum ) + 虫实(Corispermum sp.) + 猪毛菜(Salsola sp.)荒漠, 其Patrick丰富度指数仅2.60。各植被型的Pielou均匀度指数差异均不显著(P = 0.126)。
图2 内蒙古西部不同群落类型多样性指数对比
Fig. 2 Biodiversity indices of different community types in western Inner Mongolia
2.3 影响群落物种多样性的因素
2.3.1 气候因子与群落物种多样性指数的关系
结果表明, 年均温(MAT)、年降水量(MAP)、最冷月均温(MTCM)和潜在蒸散量(PET) 4个气候因子对群落的物种多样性均有影响(表2 )。Patrick丰富度指数与MAT (P < 0.01)、MTCM (P < 0.01)和PET (P < 0.01)均呈极显著的负相关关系, 与MAP呈极显著的正相关关系(P < 0.01)。Simpson指数与PET (P < 0.01)呈极显著的负相关关系, 与MAT (P < 0.05)呈显著的负相关关系, 受MAP和MTCM影响不明显。Shannon-Wiener指数与Patrick丰富度指数相似, 均与MAT (P < 0.01)、MTCM (P < 0.01)和PET (P < 0.01)呈极显著的负相关关系, 而与MAP呈显著的正相关关系(P < 0.05)。Pielou均匀度指数与各气候因子之间的关系均不显著(表2 )。
2.3.2 土壤因子与群落物种多样性的关系
Pearson相关分析显示, 土壤化学性质指标中, 土壤pH值和有机碳含量对群落的物种多样性影响较大(表3 )。Patrick丰富度指数(P < 0.01)、Shannon-Wiener指数(P < 0.01)和Simpson指数(P < 0.05)均与土壤pH值呈显著的负相关关系。4个多样性指数均与有机碳含量呈极显著的正相关关系(P < 0.01)。Patrick丰富度指数(P < 0.05)、Shannon-Wiener指数(P < 0.05)及Pielou均匀度指数(P < 0.01)均与土壤全氮呈显著的正相关关系, 其中对Pielou均匀度指数的影响极显著。土壤全磷对4个多样性指数的影响均不显著。
2.3.3 影响群落物种多样性的主导因子
通过多元逐步回归分析影响各个多样性指数的主导因子, 结果如表4 所示。各环境因子中, 潜在蒸散量和土壤有机碳含量被引入到解释Patrick丰富度指数的方程中, 且潜在蒸散量作为最重要的影响因子解释了丰富度指数的35.8%。而Simpson指数主要受到土壤有机碳含量的影响, 解释率为12.2%。Shannon-Wiener指数与丰富度指数类似, 最重要的两个影响因子也是潜在蒸散量和土壤有机碳含量, 其中, 潜在蒸散量解释了Shannon-Wiener指数的19.6%。潜在蒸散量和土壤有机碳含量是对物种多样性影响最大的因子。而对均匀度指数影响最大的环境因子是土壤全氮含量, 其解释率为10.8%。
3 讨论
3.1 不同植被类型的物种多样性
本研究调查的所有群落中, 温带草原群落的物种多样性最高(Patrick指数11.37, Simpson指数0.83,Shannon-Wiener指数2.09), 这与已有温带草原多样性的研究结果相近, 如鄂尔多斯高原典型草原Patrick丰富度指数为11.13, Simpson指数为0.79, Shannon-Wiener指数为1.99 (黄永梅和张明理, 2006 )。本研究区内草甸的物种多样性与温带草原群落的物种多样性基本一致, 但剔除其中一个丰富度指数(Patrick指数为26.20)高的薹草-杂类草草甸样地后, 草甸群落的丰富度指数变得较低(均值为7.66)。这可能主要是因为此次调查的草甸群落以芨芨草草甸为主, 属于丛生禾草盐生草甸(吴征镒, 1980 ), 为温带干旱、半干旱地区所特有, 生境相对较干旱, 土壤盐碱化明显, 植物种类组成比较匮乏, 群落的多样性也相对较低。结合已有研究表明半干旱区盐生草甸均具有相对较低的物种丰富度, 如宋创业等(2008)在浑善达克沙地的研究表明, 丘间低地及淖尔边缘的拂子茅(Calamagrostis epigejos )群落、芦苇(Phragmites communis )群落、金戴戴(Halerpestes ruthenica )群落Patrick丰富度指数均低于10, 而左小安等(2007)在科尔沁沙地的研究结果表明, 湿草甸和干草甸的Patrick丰富度指数平均值也仅为11.5和14。本研究区灌丛以草原化灌丛、沙地灌丛为主, 均为中生-旱生种(杨明博等, 2005 ), 所以灌丛的多样性也低于森林区的暖温带灌丛(谢晋阳和陈灵芝, 1997 ; 张书理, 2007 ; 周志宇等, 2009 )。荒漠是内蒙古西部的典型群落类型, 该地区约有一半的面积被荒漠覆盖, 荒漠的物种多样性低于其他各类群落。总体而言, 除温带草原群落外, 研究区各群落类型的多样性均偏低。
3.2 影响物种多样性的因素
植物群落物种多样性的主要影响因素有地形因子、气候因子、土壤理化性质、人类活动等(Sander & Wardell-Johnson, 2011 ), 本文综合分析了气候因子和土壤理化性质对该地区植物群落物种多样性的影响。
3.2.1 气候因子
内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系。在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响。此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义。一般来说, 冬季寒冷使物种多样性降低。Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响。本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系。年均温和最冷月均温对物种多样性的影响机制还需要进一步探索。
在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 )。而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 )。潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 )。在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 )。代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一。孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位。本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67。本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系。所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响。
3.2.2 土壤化学性质
土壤理化性质会在局域尺度上影响群落的物种多样性, 本文分析了土壤化学性质对群落物种多样性的影响。多元回归分析结果显示, 土壤有机碳含量是群落物种多样性的主导因子之一, 而全氮则是Pielou均匀度指数的主导因子(表4 )。
土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 )。目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低。但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加。内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性。而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子。研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低。王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响。有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨。
群落的物种多样性是多种环境因子共同作用的结果, 除了前文所述气候条件和土壤理化性质外, 地形因素和人类活动干扰的影响也不容忽视, 未来对内蒙古西部自然植被物种多样性的影响因素的研究还有待进一步加强。
参考文献
文献选项
[1]
Bai YF Li LH Wang QB Zhang LX Zhang Y Chen ZZ 2000 ) Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin, Inner Mongolia.
Acta Phytoecologica Sinica , 24 , 667 -673 . (in Chinese with English abstract)
[白永飞 , 李凌浩 , 王其兵 , 张丽霞 , 张焱 , 陈佐忠 (2000 ) 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
. 植物生态学报 , 24 , 667 -673 .]
[2]
Bai YF Wu JG Chritopher MC Shahid N Pan QM Huang JH Zhang LX Han XG 2010 ) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia Grasslands.
Global Change Biology , 16 , 358 -372 .
[本文引用: 2]
[3]
Bai YF Wu JG Xing Q Pan QM Huang JH Yang DL Han XG 2008 ) Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau.
Ecology , 89 , 2140 -2153 .
[本文引用: 1]
[4]
Bellard C Bertelsmeier C Leadley P Thuiller W Courchamp F 2012 ) Impact of climate change on the future of biodiversity.
Ecology Letters , 15 , 356 -377 .
[本文引用: 1]
[5]
Brown JH Lomolino MV 1998 ) Biogeograph, 2nd edn. Sinauer Associates, Sunderland, Massachusetts.
[本文引用: 1]
[6]
Collins SL Koerner SE Plaut JA Okie JG Brese D Calabrese LB Carvajal A Evansen RJ Nonaka E 2012 ) Stability of tallgrass prairie during a 19-year increase in growing season precipitation.
Functional Ecology , 26 , 1450 -1459 .
[本文引用: 1]
[7]
Dai S Wang XP Liu C Wu X Li QY Wang M 2013 ) Relationship between shrub species richness and climate across central Inner Mongolia, China.
Acta Scientiarum Naturalium Universitatis Pekinensis , 49 , 689 -698 . (in Chinese with English abstract)
[本文引用: 1]
[代爽 , 王襄平 , 刘超 , 武娴 , 李巧燕 , 王敏 (2013 ) 内蒙古灌木群落物种丰富度与气候的关系
. 北京大学学报(自然科学版) , 49 , 689 -698 .]
[本文引用: 1]
[8]
Doran JW Elliott ET Paustian K 1998 ) Soil microbial activity, nitrogen cycling, and long-term changes in organic carbon pools as related to fallow tillage management.
Soil & Tillage Research , 49 , 3 -18 .
[本文引用: 1]
[9]
Fang JY Wang XP Shen ZH Tang ZY He JS Yu D Jiang Y Wang ZH Zheng CY Zhu JL Guo ZD 2009 ) Methods and protocols for plant community inventory.
Biodiversity Science , 17 , 533 -548 . (in Chinese with English abstract)
[本文引用: 1]
[方精云 , 王襄平 , 沈泽昊 , 唐志尧 , 贺金生 , 于丹 , 江源 , 王志恒 , 郑成洋 , 朱江玲 , 郭兆迪 (2009 ) 植物群落清查的主要内容、方法和技术规范
. 生物多样性 , 17 , 533 -548 .]
[本文引用: 1]
[10]
Feng JM 2008 ) Spatial patterns of species diversity of seed plants in China and their climatic explanation.
Biodiversity Science , 16 , 470 -476 . (in Chinese with English abstract)
[冯建孟 (2008 ) 中国种子植物物种多样性的大尺度分布格局及其气候解释
. 生物多样性 , 16 , 470 -476 .]
[11]
Feng XW Wang Y Wu LP 2003 ) Integrated ecological environment evaluation in west region of Inner Mongolia.
Journal of Desert Research , 3 , 322 -327 . (in Chinese with English abstract)
[冯学武 , 王弋 , 吴丽萍 (2003 ) 内蒙古西部生态环境综合评价研究
. 中国沙漠 , 3 , 322 -327 .]
[12]
He JS Chen WL 1997 ) A review of gradient changes in species diversity of land plant communities.
Acta Ecologica Sinica , 17 , 91 -99 . (in Chinese with English abstract)
[本文引用: 2]
[贺金生 , 陈伟烈 (1997 ) 陆地植物群落物种多样性的梯度变化特征
. 生态学报 , 17 , 91 -99 .]
[本文引用: 2]
[13]
Huang YM Zhang ML 2006 ) Temporal and spatial changes of plant community diversity on the Ordos Plateau.
Biodiversity Science , 14 , 13 -20 . (in Chinese with English abstract)
[黄永梅 , 张明理 (2006 ) 鄂尔多斯高原植物群落多样性时空变化特点
. 生物多样性 , 14 , 13 -20 .]
[14]
Liu XH Li LH Chen ZZ 1998 ) Characteristics of vegetation diversity in the Xilin River Basin and their responses to climatic factors.
Acta Phytoecologica Sinica , 22 , 466 -472 . (in Chinese with English abstract)
[本文引用: 1]
[刘先华 , 李凌浩 , 陈佐忠 (1998 ) 内蒙古锡林河流域植被多样性特点及其与气候因子的关系
. 植物生态学报 , 22 , 466 -472 .]
[本文引用: 1]
[15]
Loreau M Naeem S Inchausti P Bengtsson J Grime JP Hector A Hooper DU Huston MA Raffaelli D Schmid B Tilman D Wardle DA 2001 ) Biodiversity and ecosystem functioning: current knowledge and future challenges.
Science , 294 , 804 -808 .
[16]
Ma B Zhou ZY Zhang LL Gao WX Chen SK Zhang BL 2008 ) The spatial distribution characteristics of plant diversity in Alex Left Banner.
Acta Ecologica Sinica , 28 , 6099 -6106 . (in Chinese with English abstract)
[本文引用: 1]
[马斌 , 周志宇 , 张莉丽 , 高文星 , 陈善科 , 张宝林 (2008 ) 阿拉善左旗植物物种多样性空间分布特征
. 生态学报 , 28 , 6099 -6106 .]
[本文引用: 1]
[17]
Ma KP Huang JH Yu SL Chen LZ 1995 ) Plant community diversity in Dongling Mountain, Beijing, China. II. Species richness, evenness and species diversities.
Acta Ecologica Sinica , 15 , 268 -277 . (in Chinese with English abstract)
[马克平 , 黄建辉 , 于顺利 , 陈灵芝 (1995 ) 北京东灵山地区植物群落多样性的研究. II. 丰富度、均匀度和物种多样性指数
. 生态学报 , 15 , 268 -277 .]
[18]
Neff JC Townsend AR Gleixner G Lehman SJ Turnbull J Bowman WD 2002 ) Variable effects of nitrogen additions on the stability and turnover of organic carbon.
Nature , 419 , 915 -917 .
[本文引用: 1]
[19]
O’Brien EM 1993 ) Climatic gradients in woody plant species richness: towards an explanation based on an analysis of southern Africa’s woody flora.
Journal of Biogeography , 20 , 181 -198 .
[20]
Pei H Ao YH Li YP Liu ZL Zhu ZY Wang W Liang CZ 2000 ) The research on the climate division of Alashan League, Inner Mongolia.
Journal of Arid Land Resources and Environment , 14 (3 ), 46 -55 . (in Chinese with English abstract)
[本文引用: 1]
[裴浩 , 敖艳红 , 李云鹏 , 刘钟龄 , 朱宗元 , 王炜 , 梁存柱 (2000 ) 内蒙古阿拉善地区气候区划研究
. 干旱区资源与环境 , 14 (3 ), 46 -55 .]
[本文引用: 1]
[21]
Phillips OL Vargas PN Monteagudo AL Cruz AP Zans MEC Sánchez WG Yli-Halla M Rose S 2003 ) Habitat association among Amazonian tree species: a landscape-sc¬ale approach.
Journal of Ecology , 91 , 757 -775 .
[22]
Qian H Klinka K Kajahara GJ 1998 ) Longitudinal patterns of plant diversity in the North American boreal forest.
Plant Ecology , 138 , 161 -178 .
[本文引用: 2]
[23]
Sander J Wardell-Johnson G 2011 ) Impacts of soil fertility on species and phylogenetic turnover in the high-rainfall zone of the Southwest Australian global biodiversity hotspot.
Plant Soil , 345 , 103 -124 .
[本文引用: 1]
[24]
Sha W Dong SK Liu SL Liu QR Shi JB Li XW Su XK Wu Y 2016 ) Spatial patterns of plant community biomass and species diversity in Aerjin Mountain Nature Reserve and their influencing factors.
Chinese Journal of Ecology , 35 , 330 -337 . (in Chinese with English abstract)
[沙威 , 董世魁 , 刘世梁 , 刘全儒 , 石建斌 , 李晓文 , 苏旭坤 , 吴娱 (2016 ) 阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素
. 生态学杂志 , 35 , 330 -337 .]
[25]
Song CY Guo K Liu GH 2008 ) Relationships between plant community’s species diversity and soil factors on Otingdag sandy land.
Chinese Journal of Ecology , 27 , 8 -13 . (in Chinese with English abstract)
[本文引用: 1]
[宋创业 , 郭柯 , 刘高焕 (2008 ) 浑善达克沙地植物群落物种多样性与土壤因子的关系
. 生态学杂志 , 27 , 8 -13 .]
[本文引用: 1]
[26]
Suding KN Gross KL Houseman GR 2004 ) Alternative states and positive feedbacks in restoration ecology.
Trends in Ecology & Evolution , 19 , 46 -53 .
[本文引用: 1]
[27]
Sun XL Kang S Zhang Q Chang CM Ma WJ Niu JM 2015 ) Relationship between species diversity, productivity, climatic factors and soil nutrients in the desert steppe.
Acta Prataculturae Sinica , 24 (12 ), 10 -19 . (in Chinese with English abstract)
[本文引用: 1]
[孙小丽 , 康萨如拉 , 张庆 , 常昌明 , 马文静 , 牛建明 (2015 ) 荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
. 草业学报 , 24 (12 ), 10 -19 .]
[本文引用: 1]
[28]
Tang ZY Fang JY 2004 ) A review on the elevational patterns of plant species diversity.
Biodiversity Science , 12 , 20 -28 . (in Chinese with English abstract)
[本文引用: 1]
[唐志尧 , 方精云 (2004 ) 植物物种多样性的垂直分布格局
. 生物多样性 , 12 , 20 -28 .]
[本文引用: 1]
[29]
van Bavel CHM 1966 ) Potential evaporation: the combination concept and its experimental verification.
Water Resources Research , 2 , 455 -467 .
[本文引用: 1]
[30]
Vitousek PM Howarth RW 1991 ) Nitrogen limitation on land and in the sea: how can it occur?
Biogeochemistry , 13 , 87 -115 .
[31]
Volder A Briske DD Tjoelker MG 2013 ) Climate warming and precipitation redistribution modify tree-grass interactions and tree species establishment in a warm-temperate savanna.
Global Change Biology , 19 , 843 -857 .
[本文引用: 1]
[32]
Wang CL Guo QS Tan DY Shi ZM Ma C 2005 ) Haloxylon ammodendron community patterns in different habitats along southeastern edge of Zhunger Basin.
Chinese Journal of Applied Ecology , 16 , 1224 -1229 . (in Chinese with English abstract)
[本文引用: 1]
[王春玲 , 郭泉水 , 谭德远 , 史作民 , 马超 (2005 ) 准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究
. 应用生态学报 , 16 , 1224 -1229 .]
[本文引用: 1]
[33]
Wang CT Wang QJ Long RJ Jing ZC Shi HL 2004 ) Changes in plant species diversity and productivity along an elevation gradient in an alpine meadow.
Acta Phytoecol¬ogica Sinica , 28 , 240 -245 . (in Chinese with English abst¬ract)
[本文引用: 1]
[王长庭 , 王启基 , 龙瑞军 , 景增春 , 史惠兰 (2004 ) 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究
. 植物生态学报 , 28 , 240 -245 .]
[本文引用: 1]
[34]
Wang DB Ji SY Chen FP 2001 ) A review on the species diversity of plant community.
Chinese Journal of Ecology , 20 (4 ), 55 -60 . (in Chinese with English abstract)
[本文引用: 2]
[汪殿蓓 , 暨淑仪 , 陈飞鹏 (2001 ) 植物群落物种多样性研究综述
. 生态学杂志 , 20 (4 ), 55 -60 .]
[本文引用: 2]
[35]
Wang M Dong ZB Luo WY Lu JF Li JY Cui XJ Zhang Y 2015 ) Species diversity of vegetation and its relationship with soil characteristics in the southern marginal zone of the Badain Jaran Desert. Acta Botanica Boreali-Occidentalia Sinica
, 35 , 379 -388 . (in Chinese with English abstract)
[王蒙 , 董治宝 , 罗万银 , 逯军锋 , 李继彦 , 崔徐甲 , 张玉 (2015 ) 巴丹吉林沙漠南缘植被物种多样性及其与土壤特性的关系
. 西北植物学报 , 35 , 379 -388 .]
[36]
Wang YF Chen ZZ Tieszen LT 1998 ) Distribution of soil organic carbon in the major grasslands of Xilinguole, Inner Mongolia, China.
Acta Phytoecologica Sinica , 22 , 66 -72 . (in Chinese with English abstract)
[本文引用: 1]
[王艳芬 , 陈佐忠 , Tieszen LT 1998 ) 人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响
. 植物生态学报 , 22 , 66 -72 .]
[本文引用: 1]
[37]
Wu ZY 1980 ) Vegetation of China . Science Press , Beijing . (in Chinese)
[吴征镒 (1980 ) 中国植被 . 科学出版社 , 北京 .]
[38]
Xie JY Chen LZ 1997 ) The studies of some aspects of biodiversity on scrubs in the warm temperate zone in China.
Acta Phytoecologica Sinica , 21 , 197 -207 . (in Chinese with English abstract)
[本文引用: 1]
[谢晋阳 , 陈灵芝 (1997 ) 中国暖温带若干灌丛群落多样性问题的研究
. 植物生态学报 , 21 , 197 -207 .]
[本文引用: 1]
[39]
Xu YJ Chen YN Li WH Fu AH Ma XD Gui DW Chen YP 2010 ) Distribution pattern and environmental interpretation of plant species diversity in the mountainous region of Ili River Valley, Xinjiang, China.
Chinese Journal of Plant Ecology , 34 , 1142 -1154 . (in Chinese with English abstract)
[徐远杰 , 陈亚宁 , 李卫红 , 付爱红 , 马晓东 , 桂东伟 , 陈亚鹏 (2010 ) 伊犁河谷山地植物群落物种多样性分布格局及环境解释
. 植物生态学报 , 34 , 1142 -1154 .]
[40]
Yang MB Yang J Yang JY 2005 ) Physiological and biochemical analysis of the leaves of seven species of Caragana. Acta Botanica Boreali-Occidentalia Sinica
, 25 , 2272 -2276 . (in Chinese with English abstract)
[杨明博 , 杨劼 , 杨九艳 (2005 ) 锦鸡儿属7种植物叶的生理生化分析
. 西北植物学报 , 25 , 2272 -2276 .]
[41]
Zhang JN Lai X Li G Zhao JN Zhang YS Yang DL 2010 ) Response of plant diversity and soil nutrient condition to grazing disturbance in Stipa baicalensis Roshev grassland.
Acta Agrestia Sinica , 18 , 177 -182 . (in Chinese with English abstract)
[本文引用: 2]
[张静妮 , 赖欣 , 李刚 , 赵建宁 , 张永生 , 杨殿林 (2010 ) 贝加尔针茅草原植物多样性及土壤养分对放牧干扰的响应
. 草地学报 , 18 , 177 -182 .]
[本文引用: 2]
[42]
Zhang Q Hou XY Li YH Niu JM Zhou YL Ding Y Zhao LQ Li X Ma WJ Kang S 2014 a) Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia grassland.
PLoS ONE , 9 , e93518 .
[本文引用: 1]
[43]
Zhang Q Wu JG Buyantuev A Niu JM Zhou YL Ding Y Kang S Ma W 2014 b) Plant species diversity is correlated with climatic factors differently at the community and the functional group levels: a case study of desert steppe in Inner Mongolia, China.
Plant Biosystems , 150 (1 ), 121 -123 .
[本文引用: 3]
[44]
Zhang SL 2007 ) Study on Plants Diversity and Conservation in Saihanwula Nature Reserve, Inner Mongolia
. PhD dissertation, Beijing Forestry University, Beijing . (in Chinese with English abstract)
[本文引用: 1]
[张书理 (2007 ) 内蒙古赛罕乌拉自然保护区植物多样性及其保护研究
. 博士学位论文, 北京林业大学, 北京 .]
[本文引用: 1]
[45]
Zhang XP Wang MB She B Xiao Y 2006 ) Quantitative classification and ordination of forest communities in Pangquangou National Nature Reserve.
Acta Ecologica Sinica , 26 , 754 -761 .
[本文引用: 2]
[46]
Zhang XS 1989 ) The potential evapotranspriration (PE) index for vegetation and vegetation-climatic classification (1)—an introduction of main methods and PEP program.
Acta Phytoecologica et Geobotanica Sinica , 13 , 1 -9 . (in Chinese with English abstract)
[本文引用: 1]
[张新时 (1989 ) 植被的PE (可能蒸散)指标与植被-气候分类(一)——几种主要方法与PEP程序介绍
. 植物生态学与地植物学学报 , 13 , 1 -9 .]
[本文引用: 1]
[47]
Zhao YZ 2006 ) Vascular Plants of Plateau Ordos . Inner Mongolia University Press, Hohhot . (in Chinese)
[赵一之 (2006 ) 鄂尔多斯高原维管植物. 内蒙古大学出版社 , 呼和浩特 .]
[48]
Zheng JG Wu GX He MZ Li XR Gan HJ 2009 ) Correlation between plant diversity and soil properties in Alax Desert.
Journal of Arid Land Resources and Environment , 23 , 151 -155 . (in Chinese with English abstract)
[本文引用: 1]
[郑敬刚 , 吴国玺 , 何明珠 , 李新荣 , 甘红军 (2009 ) 阿拉善荒漠区植物多样性与土壤理化性质相关性研究
. 干旱区资源与环境 , 23 , 151 -155 .]
[本文引用: 1]
[49]
Zheng JK Wei TX Zheng LK Dalin Z Chen ZF Zhao J Zhu WD Sun H 2009 ) Effects of landforms on α biodiversity in slope scale.
Ecology and Environmental Sciences , 18 , 2254 -2259 . (in Chinese with English abstract)
[郑江坤 , 魏天兴 , 郑路坤 , 大林直 , 陈致富 , 赵健 , 朱文德 , 孙慧 (2009 ) 坡面尺度上地貌对α生物多样性的影响
. 生态环境学报 , 18 , 2254 -2259 .]
[50]
Zhou GS Zhang XS 1996 ) Study on Chinese climate-vegeta¬tion relationship.
Acta Phytoecologica Sinica , 20 , 113 -119 . (in Chinese with English abstract)
[本文引用: 1]
[周广胜 , 张新时 (1996 ) 中国气候-植被关系初探
. 植物生态学报 , 20 , 113 -119 .]
[本文引用: 1]
[51]
Zhou ZY Yan SY Qin Y Zou LN 2009 ) The characters of shrubby diversity of Alax arid desert region.
Journal of Arid Land Resources and Environment , 23 , 146 -150 . (in Chinese with English abstract)
[周志宇 , 颜淑云 , 秦彧 , 邹丽娜 (2009 ) 阿拉善干旱荒漠区灌木多样性的特点
. 干旱区资源与环境 , 23 , 146 -150 .]
[52]
Zhu Y Kang MY Liu QR Su Y Jiang Y He KJ Xu GC Wang GR Tao Y Zhu HF 2007 ) Relationship between biodiversity and aboveground biomass in alpine meadow on Mt. Helan, China.
Chinese Journal of Applied and Environmental Biology , 13 , 771 -776 . (in Chinese with English abstract)
[本文引用: 4]
[朱源 , 康慕谊 , 刘全儒 , 苏云 , 江源 , 和克俭 , 徐广才 , 王耿锐 , 陶岩 , 朱恒峰 (2007 ) 贺兰山高山草甸生物多样性和地上生物量的关系
. 应用与环境生物学报 , 13 , 771 -776 .]
[本文引用: 4]
[53]
Zuo XA Zhao XY Zhao HL Li YQ Guo YR Zhao YP 2007 ) Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin sand land.
Environmental Science , 28 , 945 -951 . (in Chinese with English abstract)
[左小安 , 赵学勇 , 赵哈林 , 李玉强 , 郭轶瑞 , 赵玉萍 (2007 ) 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
. 环境科学 , 28 , 945 -951 .]
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
2000
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
2000
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia Grasslands.
2
2010
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau.
1
2008
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
Impact of climate change on the future of biodiversity.
1
2012
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
Biogeograph, 2nd edn. Sinauer Associates, Sunderland, Massachusetts.
1
1998
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
Stability of tallgrass prairie during a 19-year increase in growing season precipitation.
1
2012
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
内蒙古灌木群落物种丰富度与气候的关系
1
2013
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
内蒙古灌木群落物种丰富度与气候的关系
1
2013
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
Soil microbial activity, nitrogen cycling, and long-term changes in organic carbon pools as related to fallow tillage management.
1
1998
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
中国种子植物物种多样性的大尺度分布格局及其气候解释
2008
中国种子植物物种多样性的大尺度分布格局及其气候解释
2008
陆地植物群落物种多样性的梯度变化特征
2
1997
... 根据中国植被图和实际考察情况, 分别于2012-2015年的7-8月选择代表性植物群落样地74个进行调查(图1 ).调查采用样方法, 群落类型通过优势种进行确定(吴征镒, 1980 ).样方的选取原则为尽量选择围封或远离城镇公路、人类活动干扰少的开阔地, 面积均为10 m × 10 m.对于灌木样地, 在样方4角设置4个面积为1 m × 1 m的草本样方.对灌木和草本进行分层调查, 调查内容为灌木和草本的种类、高度、盖度.对于草本样地, 在样方4角及中间设置5个面积为1 m × 1 m的小样方, 记录每个小样方中所有物种的种类、高度、盖度(方精云等, 2009 ), 最后统计每个物种在样地的出现次数以计算频度. ...
... 从WorldClim数据库(http://www.worldclim.org/ )获取月均温和逐月降水量数据, 空间分辨率为30′′ (相当于赤道地区1 km).年均温(mean annual temperature, MAT, ℃)、年平均降水量(mean annual precipitation, MAP, mm)、潜在蒸散量(potential evapotranspiration, PET, mm) (方精云等, 2009 )、最冷月均温(mean temperature of the coldest month, MTCM, ℃)根据月均温和逐月降水量数据计算 得到. ...
陆地植物群落物种多样性的梯度变化特征
2
1997
... 根据中国植被图和实际考察情况, 分别于2012-2015年的7-8月选择代表性植物群落样地74个进行调查(图1 ).调查采用样方法, 群落类型通过优势种进行确定(吴征镒, 1980 ).样方的选取原则为尽量选择围封或远离城镇公路、人类活动干扰少的开阔地, 面积均为10 m × 10 m.对于灌木样地, 在样方4角设置4个面积为1 m × 1 m的草本样方.对灌木和草本进行分层调查, 调查内容为灌木和草本的种类、高度、盖度.对于草本样地, 在样方4角及中间设置5个面积为1 m × 1 m的小样方, 记录每个小样方中所有物种的种类、高度、盖度(方精云等, 2009 ), 最后统计每个物种在样地的出现次数以计算频度. ...
... 从WorldClim数据库(http://www.worldclim.org/ )获取月均温和逐月降水量数据, 空间分辨率为30′′ (相当于赤道地区1 km).年均温(mean annual temperature, MAT, ℃)、年平均降水量(mean annual precipitation, MAP, mm)、潜在蒸散量(potential evapotranspiration, PET, mm) (方精云等, 2009 )、最冷月均温(mean temperature of the coldest month, MTCM, ℃)根据月均温和逐月降水量数据计算 得到. ...
内蒙古锡林河流域植被多样性特点及其与气候因子的关系
1
1998
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
内蒙古锡林河流域植被多样性特点及其与气候因子的关系
1
1998
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
Biodiversity and ecosystem functioning: current knowledge and future challenges.
2001
阿拉善左旗植物物种多样性空间分布特征
1
2008
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
阿拉善左旗植物物种多样性空间分布特征
1
2008
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
北京东灵山地区植物群落多样性的研究. II. 丰富度、均匀度和物种多样性指数
1995
北京东灵山地区植物群落多样性的研究. II. 丰富度、均匀度和物种多样性指数
1995
Variable effects of nitrogen additions on the stability and turnover of organic carbon.
1
2002
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
Climatic gradients in woody plant species richness: towards an explanation based on an analysis of southern Africa’s woody flora.
1993
内蒙古阿拉善地区气候区划研究
1
2000
... 本研究调查的所有群落中, 温带草原群落的物种多样性最高(Patrick指数11.37, Simpson指数0.83,Shannon-Wiener指数2.09), 这与已有温带草原多样性的研究结果相近, 如鄂尔多斯高原典型草原Patrick丰富度指数为11.13, Simpson指数为0.79, Shannon-Wiener指数为1.99 (黄永梅和张明理, 2006 ).本研究区内草甸的物种多样性与温带草原群落的物种多样性基本一致, 但剔除其中一个丰富度指数(Patrick指数为26.20)高的薹草-杂类草草甸样地后, 草甸群落的丰富度指数变得较低(均值为7.66).这可能主要是因为此次调查的草甸群落以芨芨草草甸为主, 属于丛生禾草盐生草甸(吴征镒, 1980 ), 为温带干旱、半干旱地区所特有, 生境相对较干旱, 土壤盐碱化明显, 植物种类组成比较匮乏, 群落的多样性也相对较低.结合已有研究表明半干旱区盐生草甸均具有相对较低的物种丰富度, 如宋创业等(2008)在浑善达克沙地的研究表明, 丘间低地及淖尔边缘的拂子茅(Calamagrostis epigejos )群落、芦苇(Phragmites communis )群落、金戴戴(Halerpestes ruthenica )群落Patrick丰富度指数均低于10, 而左小安等(2007)在科尔沁沙地的研究结果表明, 湿草甸和干草甸的Patrick丰富度指数平均值也仅为11.5和14.本研究区灌丛以草原化灌丛、沙地灌丛为主, 均为中生-旱生种(杨明博等, 2005 ), 所以灌丛的多样性也低于森林区的暖温带灌丛(谢晋阳和陈灵芝, 1997 ; 张书理, 2007 ; 周志宇等, 2009 ).荒漠是内蒙古西部的典型群落类型, 该地区约有一半的面积被荒漠覆盖, 荒漠的物种多样性低于其他各类群落.总体而言, 除温带草原群落外, 研究区各群落类型的多样性均偏低. ...
内蒙古阿拉善地区气候区划研究
1
2000
... 本研究调查的所有群落中, 温带草原群落的物种多样性最高(Patrick指数11.37, Simpson指数0.83,Shannon-Wiener指数2.09), 这与已有温带草原多样性的研究结果相近, 如鄂尔多斯高原典型草原Patrick丰富度指数为11.13, Simpson指数为0.79, Shannon-Wiener指数为1.99 (黄永梅和张明理, 2006 ).本研究区内草甸的物种多样性与温带草原群落的物种多样性基本一致, 但剔除其中一个丰富度指数(Patrick指数为26.20)高的薹草-杂类草草甸样地后, 草甸群落的丰富度指数变得较低(均值为7.66).这可能主要是因为此次调查的草甸群落以芨芨草草甸为主, 属于丛生禾草盐生草甸(吴征镒, 1980 ), 为温带干旱、半干旱地区所特有, 生境相对较干旱, 土壤盐碱化明显, 植物种类组成比较匮乏, 群落的多样性也相对较低.结合已有研究表明半干旱区盐生草甸均具有相对较低的物种丰富度, 如宋创业等(2008)在浑善达克沙地的研究表明, 丘间低地及淖尔边缘的拂子茅(Calamagrostis epigejos )群落、芦苇(Phragmites communis )群落、金戴戴(Halerpestes ruthenica )群落Patrick丰富度指数均低于10, 而左小安等(2007)在科尔沁沙地的研究结果表明, 湿草甸和干草甸的Patrick丰富度指数平均值也仅为11.5和14.本研究区灌丛以草原化灌丛、沙地灌丛为主, 均为中生-旱生种(杨明博等, 2005 ), 所以灌丛的多样性也低于森林区的暖温带灌丛(谢晋阳和陈灵芝, 1997 ; 张书理, 2007 ; 周志宇等, 2009 ).荒漠是内蒙古西部的典型群落类型, 该地区约有一半的面积被荒漠覆盖, 荒漠的物种多样性低于其他各类群落.总体而言, 除温带草原群落外, 研究区各群落类型的多样性均偏低. ...
Habitat association among Amazonian tree species: a landscape-sc¬ale approach.
2003
Longitudinal patterns of plant diversity in the North American boreal forest.
2
1998
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
Impacts of soil fertility on species and phylogenetic turnover in the high-rainfall zone of the Southwest Australian global biodiversity hotspot.
1
2011
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素
2016
阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素
2016
浑善达克沙地植物群落物种多样性与土壤因子的关系
1
2008
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
浑善达克沙地植物群落物种多样性与土壤因子的关系
1
2008
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
Alternative states and positive feedbacks in restoration ecology.
1
2004
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
1
2015
... 其中, S 为每个样方内的物种数, p i 为第i 个物种的重要值(马克平等, 1995 ). ...
荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
1
2015
... 其中, S 为每个样方内的物种数, p i 为第i 个物种的重要值(马克平等, 1995 ). ...
植物物种多样性的垂直分布格局
1
2004
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
植物物种多样性的垂直分布格局
1
2004
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
Potential evaporation: the combination concept and its experimental verification.
1
1966
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
Nitrogen limitation on land and in the sea: how can it occur?
1991
Climate warming and precipitation redistribution modify tree-grass interactions and tree species establishment in a warm-temperate savanna.
1
2013
... 内蒙古西部包括乌兰察布、呼和浩特、包头、鄂尔多斯、巴彦淖尔、乌海和阿拉善7个盟市, 地理坐标为37°30°-42°47′ N, 95°05′-114°49′ E.该区以高原地貌为主, 平均海拔超过1,000 m.气候以温带大陆性季风气候为主, 年均温约-2℃至10℃, 年降水量约29-500 mm, 阿拉善高原的年降水量仅50- 150 mm, 而额济纳旗仅有36 mm (裴浩等, 2000 ), 且蒸发量相对较大.内蒙古西部日照充足, 大部分地区的年日照时数都超过2,700 h, 阿拉善高原西部地区的年日照时数超过3,400 h.该地区以温带荒漠和温带草原为主, 典型植物群落包括克氏针茅(Stipa krylovii )草原、短花针茅(S. breviflora )草原、油蒿(Artemisia ordosica )灌丛、藏锦鸡儿(Caragana tibetica )灌丛、红砂(Reaumuria songarica )和白刺(Nitraria tangutorum )灌丛等. ...
准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究
1
2005
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究
1
2005
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究
1
2004
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究
1
2004
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
植物群落物种多样性研究综述
2
2001
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... 植物群落物种多样性的主要影响因素有地形因子、气候因子、土壤理化性质、人类活动等(Sander & Wardell-Johnson, 2011 ), 本文综合分析了气候因子和土壤理化性质对该地区植物群落物种多样性的影响. ...
植物群落物种多样性研究综述
2
2001
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... 植物群落物种多样性的主要影响因素有地形因子、气候因子、土壤理化性质、人类活动等(Sander & Wardell-Johnson, 2011 ), 本文综合分析了气候因子和土壤理化性质对该地区植物群落物种多样性的影响. ...
巴丹吉林沙漠南缘植被物种多样性及其与土壤特性的关系
2015
巴丹吉林沙漠南缘植被物种多样性及其与土壤特性的关系
2015
人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响
1
1998
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响
1
1998
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
中国暖温带若干灌丛群落多样性问题的研究
1
1997
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
中国暖温带若干灌丛群落多样性问题的研究
1
1997
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
伊犁河谷山地植物群落物种多样性分布格局及环境解释
2010
伊犁河谷山地植物群落物种多样性分布格局及环境解释
2010
贝加尔针茅草原植物多样性及土壤养分对放牧干扰的响应
2
2010
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
贝加尔针茅草原植物多样性及土壤养分对放牧干扰的响应
2
2010
... 内蒙古西部横跨半干旱、干旱两个气候带, 植被和土壤都具有明显的地带性分布差异, 植物群落类型丰富(冯学武等, 2003 ).作为我国西北干旱区的重要组成部分, 该地区已有大量研究.如孙小丽等(2015)对荒漠草原物种多样性和生产力的研究, 代爽等(2013)探讨了灌木群落物种丰富度与气候的关系, 张静妮等(2010)通过对贝加尔针茅(Stipa baicalensis )草原的研究, 阐述了土壤化学性质对草地群落植物组分、结构和多样性格局的影响.但这些研究多集中在单一群落类型(周志宇等, 2009 ; 张静妮等, 2010 ; 代爽等, 2013 ; 孙小丽等, 2015 ), 而对区域尺度的群落物种多样性特征及其空间分布格局的研究较少.在生物多样性形成机制的环境解释方面, 有很多研究只针对气候或土壤单一因子对物种多样性的影响(刘先华等, 1998 ; 白永飞等, 2000 ; 左小安等, 2007 ; 宋创业等, 2008 ), 将气候和土壤等环境因子相结合的综合分析研究尚不多见, 这不利于对干旱半干旱地区生物多样性形成机制的深入认识.本文在华北植物群落综合调查项目的支持下, 在2012-2015年通过实地调查获取了内蒙古西部4个植被类型共74个样地的数据, 分析了该区自然植被的物种多样性特征及其空间分布格局.在此基础上, 结合降水、气温、土壤等环境数据, 进一步探讨了干旱半干旱地区自然环境因素对物种多样性的影响. ...
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
a) Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia grassland.
1
2014
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
b) Plant species diversity is correlated with climatic factors differently at the community and the functional group levels: a case study of desert steppe in Inner Mongolia, China.
3
2014
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
内蒙古赛罕乌拉自然保护区植物多样性及其保护研究
1
2007
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
内蒙古赛罕乌拉自然保护区植物多样性及其保护研究
1
2007
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
Quantitative classification and ordination of forest communities in Pangquangou National Nature Reserve.
2
2006
... 内蒙古西部处于干旱、半干旱区, 一般认为水热因子是影响该区域植物群落物种多样性的主要因素, 如锡林河流域(刘先华等, 1998 ; 白永飞等, 2000 )、内蒙古荒漠草原(Zhang et al, 2014b ; 孙小丽等, 2015 )、内蒙古草原(Zhang et al, 2014a )的研究表明物种丰富度和多样性均与年降水量呈正相关, 而与年均温呈负相关关系.在本研究中, Patrick丰富度指数、Simpson指数和Shannon-Wiener指数均与年均温呈显著的负相关关系, 而Patrick丰富度指数和Simpson指数与年降水量呈显著的正相关关系, 这与已有研究结果基本一致, 说明水热因子对该区域植物群落的物种多样性存在较大的影响.此外, 冬季寒冷会影响多年生植物的生活史策略, 从而影响群落的物种多样性(Zhang et al, 2016 ), 因此, 最冷月均温对群落的物种多样性也具有重要意义.一般来说, 冬季寒冷使物种多样性降低.Zhang等(2014b)的研究也表明, 最冷月均温对灌木-半灌木群落的多样性具有显著的影响.本研究中Patrick丰富度指数和Shannon-Wiener指数均与最冷月均温具有显著的负相关关系.年均温和最冷月均温对物种多样性的影响机制还需要进一步探索. ...
... 土壤有机碳含量及其动态平衡是反映土壤质量的一个重要指标, 直接影响着土壤肥力, 从而影响植物营养的生物有效性(Doran et al, 1998 )氮的可利用性是陆地生态系统的主要限制因素之一, 影响着生态系统的物种多样性(Vitousek & Howarth, 1991 ).目前广泛开展的氮添加实验结果表明, 氮的增加会使群落的物种多样性降低.但Suding等(2004) 的研究发现, 在氮限制的地区增加氮素会使群落的物种多样性增加.内蒙古西部地区属于氮限制地区(Bai et al, 2010 ), 在不添加外源氮素的情况下, 土壤全氮含量较高的群落, 氮限制的程度相对较轻, 可维持较高的物种多样性.而较高的氮素也使植物对氮素的竞争减弱, 这也可以解释为何在研究区内全氮含量成为Pielou均匀度指数的主导因子.研究区中土壤全氮含量较低的柽柳灌丛、红砂荒漠及四合木荒漠的物种多样性均较低, Patrick丰富度指数均低于8, Pielou均匀度指数也较其他群落低.王蒙等(2015)对巴丹吉林沙漠南缘自然植被的研究结果也显示, 均匀度指数与土壤全氮含量显著相关, 而丰富度指数则主要受有机质含量影响.有研究表明, 全球氮沉降水平预计在未来25年内会加倍(Neff et al, 2002 ), 在这样的背景下, 内蒙古西部自然植被物种多样性将如何变化也值得进一步探讨. ...
植被的PE (可能蒸散)指标与植被-气候分类(一)——几种主要方法与PEP程序介绍
1
1989
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
植被的PE (可能蒸散)指标与植被-气候分类(一)——几种主要方法与PEP程序介绍
1
1989
... 在北美、非洲以及欧亚草原的许多研究表明, 降水是群落物种多样性最主要的影响因子(Bai et al, 2008 ; Collins et al, 2012 ; Volder et al, 2013 ).而在本研究中, 基于年均温和年降水量计算的潜在蒸散量对群落的物种多样性影响显著, 多元逐步回归结果也表明, 潜在蒸散量是内蒙古西部群落物种多样性的主导因素中唯一的气候因子(表4 ).潜在蒸散量作为综合热量和水分两个最重要的生态因子的参数, 对于植被的分布和群落的组成具有重要意义(van Bavel, 1966 ; 张新时, 1989 ; 周广胜和张新时, 1996 ).在非洲南部的研究表明, 与温度相比, 潜在蒸散量可能更适合用于反映热量或能量对植被的影响(O’Brien, 1993 ).代爽等(2013)对内蒙古中西部灌木群落的研究发现, 物种丰富度的地理格局受水分和能量共同限制的水分能量动态假说也能较好地解释本研究区灌木群落多样性的地理格局, 而潜在蒸散量正是该假说的主要参数之一.孙小丽等(2015)对内蒙古荒漠草原的研究也发现, 反映水分和温度综合作用的干燥指数在对群落物种多样性的作用中占主导地位.本研究区内潜在蒸散量最低的薹草-杂类草草甸群落中, Patrick丰富度指数高达26.20, 而潜在蒸散量最高的梭梭(Haloxylon ammodendron )荒漠群落, 其Patrick丰富度指数仅为3.67.本研究中, Patrick丰富度指数、Shannon-Wiener指数和Simpson指数均与潜在蒸散量呈极显著的负相关关系.所以, 未来的群落物种多样性的影响因素研究中, 应更重视潜在蒸散量的影响. ...
阿拉善荒漠区植物多样性与土壤理化性质相关性研究
1
2009
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
阿拉善荒漠区植物多样性与土壤理化性质相关性研究
1
2009
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
中国气候-植被关系初探
1
1996
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
中国气候-植被关系初探
1
1996
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
贺兰山高山草甸生物多样性和地上生物量的关系
4
2007
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... 群落的物种多样性是指群落中植物种类的丰富程度, 而物种多样性(α多样性)指标是度量物种多样性高低及空间分布特征的数值指标.近年来对植物群落物种多样性的研究采用的测度指数主要包括物种丰富度指数、物种多样性指数及均匀度指数, 其中, Patrick丰富度指数、Simpson指数、Shannon-Wiener指数和Pielou均匀度指数在测度物种多样性中较为常见, 与其他研究具有较大可比性(汪殿蓓等, 2001 ).本文也采用这4种多样性指数来表征研究区植物群落的物种多样性.在计算过程中, 首先根据每个样方中各灌木、草本物种的高度、盖度、频度计算每个物种的重要值(p i ) (Zhang et al, 2006 ): ...
贺兰山高山草甸生物多样性和地上生物量的关系
4
2007
... 物种组成及其多样性是群落最基本的特征, 是形成群落的基础(王春玲等, 2005 ).物种多样性对于维持群落功能, 特别是环境条件多变的群落的功能具有至关重要的意义(Loreau et al, 2001 ).物种多样性的变化与生境紧密相关(汪殿蓓等, 2001 ), 生物多样性沿环境梯度的变化规律已成为生物多样性研究的一个重要议题(唐志尧和方精云, 2004 ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... ), 了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义.影响植物群落物种多样性的因素可分为环境因子和生物因子两类(汪殿蓓等, 2001 ).环境因子中, 经度(Qian et al, 1998 ; 马斌等, 2008 )、纬度(贺金生和陈伟烈, 1997 )、海拔(唐志尧和方精云, 2004 ; 王长庭等, 2004 ; 朱源等, 2007 ; 徐远杰等, 2010 )、气候(冯建孟, 2008 ; Bellard et al, 2012 )、地质地貌(郑江坤等, 2009 )、土壤的物理化学性质(郑敬刚等, 2009 ; Sander & Wardell-Johnson, 2011 )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... )等都会对群落的物种多样性产生影响.生物因子尤其是人为活动干扰对植物群落物种多样性的改变影响也较大(汪殿蓓等, 2001 ).在大尺度范围, 群落物种多样性主要受纬度和海拔影响, 一般认为物种多样性随纬度增加而下降(Brown & Lomolino, 1998 ), 而随海拔梯度的变化有多种形式, 最常见的为负相关和中间高度“膨胀现象” (沙威等, 2016 )两种.而在区域尺度上, 地形、气候、土壤则成为主要影响因素, 这3种要素的不同组合产生了复杂的群落物种多样性格局(Phillips et al, 2003 ). ...
... 群落的物种多样性是指群落中植物种类的丰富程度, 而物种多样性(α多样性)指标是度量物种多样性高低及空间分布特征的数值指标.近年来对植物群落物种多样性的研究采用的测度指数主要包括物种丰富度指数、物种多样性指数及均匀度指数, 其中, Patrick丰富度指数、Simpson指数、Shannon-Wiener指数和Pielou均匀度指数在测度物种多样性中较为常见, 与其他研究具有较大可比性(汪殿蓓等, 2001 ).本文也采用这4种多样性指数来表征研究区植物群落的物种多样性.在计算过程中, 首先根据每个样方中各灌木、草本物种的高度、盖度、频度计算每个物种的重要值(p i ) (Zhang et al, 2006 ): ...
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
2007
科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系
2007



{kind=link}
{kind=link}
{kind=link}
{kind=link}