植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 )。其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面。植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成。
植物区系地理学(floristic geography)是主要研究全球或某一区域植物种类的构成、现代和过去的分布规律以及起源进化、演变历史的科学(王荷生, 1992 )。植物区系地理格局的形成建立在明确植物区系分布区类型形成和起源的基础之上。积累植物区系的基本特征等方面的研究资料, 有利于对植物多样性的基础理论问题开展深入研究, 并为生物多样性的保护提供理论依据和实践指导(孙航等, 2017 )。
海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013)。海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 )。整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作。《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属。这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题。此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道。虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 )。
马鞍岭火山口及周边地区是我国唯一的热带海岛城市火山群地质公园, 也是海南省第一家由联合国教科文组织确认具有重要价值的国家4A级旅游景区。但其详尽的植物多样性研究仍是空白。目前, 有关我国火山地区植物区系的研究只有黑龙江五大连池火山自然保护区的植物区系调查研究( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.)和黑龙江省宁安市境内镜泊湖熔岩台地的种子植物多样性研究(蔡赫, 2011 )等, 与本文调查区域在气候条件、人文地理、物种组成和植物区系地理方面均显著不同。为进一步明确马鞍岭火山口及周边维管植物的现状, 我们于2016年12月至2017年3月底开展了本次调查。结合相关文献资料及植物标本, 对马鞍岭火山口地区的植物种类进行鉴定统计, 对不同土地利用类型(人工林、次生林和撂荒地)中的植物多样性及植物区系基本特征、组成和可能的起源与演化历史进行了分析, 拟为该地区植物区系研究提供参考。
1 材料与方法
1.1 研究区域概况
马鞍岭火山口地区指海口市西南石山、永兴镇一带, 西自美造水库, 东至龙桥, 南起罗京盘, 北迄长流的全新世火山岩分布区, 地理坐标为19°48′-20°01′ N, 110°06′-110°27′ E, 面积约500 km² (白志达等, 2003 )。该区域处于热带与南亚热带过渡区, 年均温23.7℃, 年积温8,700℃, 最热月7月均温为29-31℃, 最冷月1月均温17.2℃, 偶尔低至0℃, 全年暖热无冬; 雨量充沛, 年降水量1,685 mm左右, 且多集中于夏秋两季, 雨季常有台风侵袭; 相对湿度约85%, 全年日照时数达2,240 h。气候十分适于热带及亚热带植物生长, 具有我国热带及向南亚热带过渡生物群落的典型特征。
该地具有丰富的与火山相伴的熔岩构造, 为世界罕见的火山地貌景观, 是名副其实的第四纪火山天然博览园。海拔一般在50-100 m, 最高峰为马鞍岭, 有222 m, 超过100 m的还有雷虎岭(130 m)。雷虎和风炉二岭是熔岩台地上的火山锥, 其中部为火山口, 火山锥周围是高低不平的地面或缓坡, 常堆积有火山砾块、火山灰、火山乱石堆(不少为当地群众堆成的石篱)、乱石滩等, 形成典型的石地地形。土壤主要为灰色-黑灰色熔岩块和熔渣覆盖, 土层较浅, 但土质肥沃, 有机质多, 结构疏松( 何平荣 (2008)琼北火山岩地区兰科植物多样性及美花石斛传粉生物学研究. 硕士学位论文, 广西师范大学, 广西桂林.)。
1.2 野外调查
植物种类采用样方调查, 将以马鞍岭火山口为中心的100 km2 的正方形区域划分成100个1 km × 1 km的样地。每个样地随机设置1个20 m × 20 m的样方, 调查样方内出现的乔木种类、数量、高度、胸径、冠幅、物候期、生活力等。在调查乔木的样方内随机设置3个5 m × 5 m的样方, 调查灌木的种类、数量、高度、冠径、物候期、生活力和盖度等。随机设置5个1 m × 1 m的样方, 调查草本植物的种类、株高、物候期、生活力、盖度等。共计调查乔木样方100个, 灌木样方300个, 草本样方500个。
1.3 植物区系组成分析
依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新。按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成。蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法。
非本地植物类别的确定参考杨小波(2016)。逸生种指原为本地和外来引入的栽培种, 而后部分从栽培逃逸为野生状态的植物; 归化种是指外来植物转入本地后直接正常繁育后代, 并大量繁衍成野生状态的植物; 外来入侵种是指外来植物进入本地区后大量繁殖并给本地区的生态系统等造成严重损害的植物, 有部分植物既是归化种也是入侵种。珍稀濒危植物参考《中国生物多样性红色名录——高等植物卷》(环境保护部和中国科学院, 2013)
1.4 生物多样性数据分析
依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数。
$d=\frac{1}{\frac{n_{max}}{N}}$ (1)
$H'=-\sum^{s}_{i=1}p_{i} lnp{i}, p^2_{i}=\frac{n_{i}}{N}$ (2)
$Je=\frac{H'}{H'_{max}}, H'_{max}\ln s$ (3)
式中, N为所有物种的个体数之和, ni 为第i 个种个体数量, s 为群落中的总物种数。
1.5 谱系树的建立及谱系多样性
Phyto谱系树是Qian和Jin (2015)在Zanne等(2014)的谱系树基础上进一步更新、扩展而成。基于样地调查所获的物种数据, 在Qian和Jin (2015)的谱系树基础上, 利用S. Phylo Maker函数以3种不同方式添加物种构建本文的谱系树(如, Scenario 1, Scenario 2, Scenario 3)。我们选用的是第3种(Scenario 3), 该谱系树将Phyto中没有的物种以多分支形态添加到其父母分类单元下, 用BLADJ算法为其分配支长(Webb et al, 2008 ; Qian & Jin, 2015 )。
用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性。PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例。MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标。MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 )。PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算。β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 )。本文数据分析采用SPSS 18.0软件实现。
2 结果
2.1 植物科属组成与物种丰富度
本次在海口马鞍岭火山口及周边地区共记录到维管植物284种(包括种下分类单位, 被子植物参照修订后的APG III系统)(附录1), 隶属于88科241属, 乔木、灌木、草本及藤本4种生活型的物种数分别为77、76、90和41种(表1 )。常见的乔木有荔枝(Litchi chinensis )、龙眼(Dimocarpus longan )、楝(Melia azedarach )、桉(Eucalyptus robusta )等, 灌木有毛柿(Diospyros strigosa )、鹊肾树(Streblus asper )、酒饼簕(Atalantia buxifolia )、马缨丹(Lantana camara )等, 草本有鬼针草(Bidens pilosa )、飞机草(Chromolaena odorata )、吐烟花(Pellionia repens )、斑茅(Saccharum arundinaceum )等, 藤本植物有乌蔹梅(Cayratia japonica )、厚叶崖爬藤(Tetrastigma pachyphyllum )、掌叶鱼黄草(Merremia vitifolia )等。
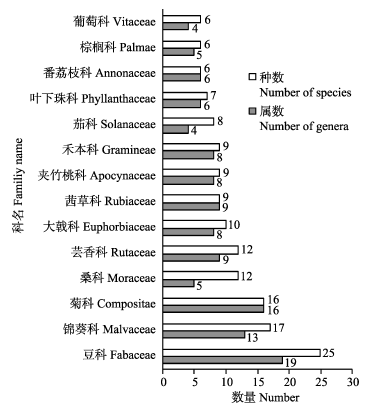
图1 海口马鞍岭火山口地区科属数排列前14位的植物科和属
Fig. 1 The number of families of the top 14 plants and genera at Haikou Maanling volcanic area
马鞍岭火山口及周边地区维管植物主要集中在14个科(每科含有6种以上植物)(图1 ), 占全部维管植物科总数的15.91%, 属总数的49.38%, 种总数的53.17%, 是马鞍岭维管植物的主体。其中豆科物种最为丰富(19属25种), 占全部维管植物物种数的8.80%。
2.2 植物的组成类型
调查结果表明, 海口马鞍岭地区栽培植物40种, 占该地区植物总数的14.08%, 其中大部分是观赏植物, 如黄蝉(Allamanda schottii )、美人蕉(Canna indica )、吊竹梅(Tradescantia zebrina )等, 还有部分园艺作物, 如芋(Colocasia esculenta )、木薯(Manihot esculenta )、菠萝蜜(Artocarpus heterophyllus )、丝瓜(Luffa cylindrica )等; 逸生种19种, 占总物种数6.69%; 归化种31种, 占总物种数的10.92%; 外来入侵种23种, 占总物种数的8.10%; 本地物种71科164属194种, 占总物种数的68.31%。
本地区有珍稀濒危植物4种, 分别为: 土沉香(Aquilaria sinensis )、降香(Dalbergia odorifera )、海南龙血树(Dracaena cambodiana )和荔枝。其中土沉香和降香被《中国物种红色名录》(汪松和解焱, 2004 )列为易危和极危物种, 也被《IUCN红色名录(2013)》列为易危物种, 被《国家重点保护植物名录》列为国家二级保护植物。此外, 土沉香还被《濒危野生动植物种国际贸易公约》附录II收录。《国家重点保护植物名录》将海南龙血树列为易危物种, 将荔枝列为濒危物种。
2.3 植物区系的地理成分
从科的地理成分来看(表2 ), 马鞍岭植物区系中以泛热带分布的科最多, 有38科, 占总科数的45.78%; 其次热带亚洲至热带美洲间断分布科和北温带分布科分别占7.23%, 并列第二; 然后依次是旧世界热带分布科、热带亚洲至大洋洲分布科和热带非洲-热带美洲间断分布科。其中热带分布科(类型2-7)占61.44%, 温带分布科(类型8-14)占7.23%, 热带分布科数量显著高于温带分布科。
从属的地理成分来看, 马鞍岭植物区系中热带分布属(类型2-7)合计205属, 占所统计属总数的87.98%。其中泛热带分布属最多, 占35.62%, 如马缨丹属(Lantana )、榕属(Ficus )、飞机草属(Chromolaena )、含羞草属(Mimosa )等; 其次是旧世界热带分布属, 占16.31%, 如山牵牛属(Thunbergia )、暗罗属(Polyalthia )、八角枫属(Alangium )、红瓜属(Coccinia )等; 再次是热带亚洲至大洋洲分布属, 占11.59%, 如眼树莲属(Dischidia )、厚壳树属(Ehretia )、芭蕉属(Musa )、槟榔属(Areca )等; 热带亚洲至热带美洲间断分布属占8.58%, 如量天尺属(Hylocereus )、番石榴属(Psidium )、紫茉莉属(Mirabilis )、凤梨属(Ananas )等。
温带分布属合计16属, 占所统计属的6.87%, 其中东亚和北美间断分布属最多, 占3.43%, 如络石属(Trachelospermum )、楤木属(Aralia )、柯属(Lithocarpus )、勾儿茶属(Berchemia )等; 北温带分布属位居其次, 占2.15%, 如盐肤木属(Rhus )、忍冬属(Lonicera )、胡颓子属(Elaeagnus )、桑属(Morus )等。其他还有东亚分布属、中亚分布属等。从属的分布区类型看, 热带分布属在马鞍岭植物区系中占绝对优势, 热带性质十分显著。
马鞍岭火山口地区蕨类植物共5科8属8种。从科的地理成分看, 该地蕨类植物区系共有泛热带分布科和世界分布科两类, 其中泛热带分布科占80%, 世界分布科占20%。从属的地理成分看, 泛热带分布属占50.00%, 居于首位, 如海金沙属(Lygodium )、肾蕨属(Nephrolepis )、凤尾蕨属(Pteris )、毛蕨属(Cyclosorus )等; 其次是旧世界热带分布属, 占25.00%, 如鳞盖蕨属(Microlepia )、瘤蕨属(Phymatosorus )等; 世界分布属和东亚分布属分别占12.50%。
2.4 植物物种多样性和谱系多样性
在3种不同土地利用类型(次生林、人工林、撂荒地)中, 不同植被类型的多样性不一样, 其结果显示为: 人工林与撂荒地的乔木优势度差异显著, 而次生林与人工林、撂荒地差异不显著; 灌木和草本优势度在3种土地利用类型间均无显著差异。乔木的Shannon指数在3种不同土地利用类型间差异显著, 表现为人工林 > 次生林 > 撂荒地。 但灌木和草本的Shannon指数在3种土地利用类型间差异不显著。不同土地利用类型中不同生活型水平的Pielou均匀度指数差异均不显著(表3 )。
在谱系多样性方面, 人工林的PD和NTAXA最大, 撂荒地最小, 人工林和次生林差异不显著, 但两者均与撂荒地差异显著。人工林和撂荒地的MPD差异显著, 但两者跟次生林之间的差异都不显著。次生林、撂荒地、人工林的MNTD差异不显著(表4 )。
3 讨论
3.1 马鞍岭火山口地区和其他地区的植物区系组成比较
在我国的火山地区植物区系中, 马鞍岭火山口地区单位面积物种数较多且保护植物的占比较高。在本研究的100 km2 范围内, 我们共发现维管植物284种, 单位陆地面积的种数高达2.84种/km2 。这远高于其他火山植物区系, 如黑龙江五大连池火山自然保护区占地面积1,060 km2 , 有野生植物143科428属l,044种( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.), 单位面积的物种数为0.98种/km2 。黑龙江省宁安市境内镜泊湖熔岩台地占地面积达1,200 km2 , 有种子植物77科265属486种(蔡赫, 2011 ), 单位面积的物种数仅0.41种/km2 。
马鞍岭火山口地区共有非本地植物90种, 占总物种数的31.69%。其中外来入侵物种23种、逸生种19种、归化种31种、栽培种40种, 各占总物种数的8.10%、6.69%、10.92%、14.08%, 而有23种既是入侵种也是归化种。马鞍岭单位面积的外来物种数为0.90种/km2 , 与五大连池火山自然保护区和镜泊湖熔岩台地相比, 马鞍岭的非本地物种较为丰富, 这可能跟马鞍岭地区受人为干扰严重有关。马鞍岭地区本地植物共194种, 单位面积的本地物种数为1.94种/km2 , 相对国内同类火山口地区来说也相当丰富, 这可能跟马鞍岭复杂的地理环境及与大陆相连的历史有关, 它既受到大陆起源的影响, 又一定程度地受到海岛影响, 这样的环境蕴育了丰富多样的野生植物种类。
海口马鞍岭火山口地区植物属的分布区类型与铜鼓岭自然保护区类似, 均以泛热带分布属为主(杨小波等, 2011 ); 跟以热带亚洲成分为主的五指山、尖峰岭热带雨林不同。马鞍岭火山口地区没有中国特有属, 铜鼓岭只有1个, 而吊罗山、尖峰岭、五指山自然保护区分别有10、15、16个。这一方面可能是马鞍岭地区和铜鼓岭自然保护区受人为影响严重, 另一方面它们与另外三者地理及环境存有较大差异。吊罗山、尖峰岭、五指山自然保护区海拔均在1,200 m以上, 海拔跨度较大, 年平均气温比铜鼓岭及马鞍岭火山口偏低, 所以植物的分布区类型有所不同, 但均以热带区系为主。且这3个自然保护区的面积相对较广, 人为干扰较少, 物种保护较好, 而马鞍岭火山口和铜鼓岭海拔低, 年平均气温偏高, 人为干扰严重, 导致物种丰富度及特有性比其他3个保护区低, 且物种存在进一步减少的可能。
Zhu (2016) 将海南岛的植物区系同我国广西、广东以及越南的植物区系进行比较, 发现海南的植物区系与越南的最为相似, 其次是广西。海南植物区系的特异性十分低, 特有属仅7个, 特有种只有10%左右, 表明其大陆起源的特征。
对比越南以及我国广东、广西的植物区系发现, 马鞍岭火山口地区的植物区系跟广东的相似性最大, 其次是越南; 在所比较的164个属的本地植物中, 越南、广东、广西都仅有1个属无分布, 分别是白饭树属(Flueggea )、龙血树属(Dracaena) 、牛筋果属(Harrisonia ), 这在一定程度上显示了马鞍岭火山口植物大陆起源的特征, 和Zhu (2016)的结果类似。但在种的层面上, 马鞍岭火山口地区的194种本地植物中, 仅7种不在广东分布, 12种不在越南分布, 14种不在广西分布, 仅有水仙柯(Lithocarpus naiadarum )在3个地区都不分布, 表明该地植物特有性非常低。这与Zhu (2016) 的结果有一定差异。一方面可能跟马鞍岭火山口植被的形成有关: 马鞍岭火山口最后一次喷发在距今约1万年前, 火山喷发可能彻底毁灭了先前的植被, 再形成了现在的植被形态, 相比整个海南岛的植物区系来说, 马鞍岭火山口地区的植物区系的形成时间更年轻, 并在之后的时间里跟地理位置更近的雷州半岛之间的物种交流更频繁, 导致与广东植物区系更相近; 另一方面本次调查的范围相比整个海南岛而言面积小, 物种数相对少, 且环境一致, 可能导致与Zhu (2016) 的研究结果有偏差。
3.2 物种多样性和谱系多样性
不同土地利用类型中不同生活型的优势度都较高, 这说明优势种较显著, 特别是人工林的乔木优势度与撂荒地之间差异显著; 群落多样性结果显示不同土地利用类型中, 乔木的H' 差异显著, 这可能与该地的投入肥料和虫害管理导致土壤肥力不同(徐远杰等, 2017 )有关。马鞍岭火山口地区的人工林主要是荔枝、龙眼等乔木类的果园或者其他经济林, 这是导致人工林与撂荒地之间乔木的优势度差异显著及人工林、次生林、撂荒地的乔木群落多样性差异显著的主要原因。绝大多数的人工林在建林后的数十年间缺乏管理, 导致灌木和草本的优势度和群落多样性与次生林和撂荒地的差异不显著(仲磊等, 2015 )。
本研究发现, 人工林的PD、NTAXA、MPD、MNTD值最大, 表明人工林群落中的物种最丰富, 但物种相互之间的亲缘关系疏远, 因而谱系结构呈现发散状态。这可能跟人工林(主要是荔枝、龙眼、菠萝蜜和大叶桉林)在前期较长时间的人工干预条件下保留了较多果树等经济作物, 而后在较短的时间内缺乏人为干扰, 较多的草本及灌木大量生长有关。先前的人工干预导致林冠茂密, 而后自然生长的其他乔木、灌木及草本可获取的阳光、矿物质等资源有限, 种间竞争激烈, 物种在资源利用策略上倾向于差异化(Devictor et al, 2010 ; Helmus & Ives, 2012 )。这样先经过人工过度干预而后长期的自然生长导致人工林出现群落物种间亲缘关系疏远而谱系结构发散的特征。次生林的PD、NTAXA、MPD、MNTD值较大, 反映次生林群落中的物种较丰富, 物种间亲缘关系较近, 谱系结构相对聚集。这可能跟次生林的形成过程及环境有关, 马鞍岭的次生林发生在被大面积严重砍伐的区域, 主要由植物在较短的时间范围内自然生长形成。因发育时间短, 加之该地的表土多为火山岩, 土壤较少, 故难以形成发育程度较好的次生林, 多为低矮灌木, 少有小乔木及藤本。这就形成了马鞍岭次生林相对较近的物种间亲缘关系及相对聚集的谱系结构。撂荒地的PD、NTAXA、MPD、MNTD值最小, 这说明其物种相对单一, 群落中物种间的亲缘关系最近和谱系结构最为聚集。可能由于马鞍岭的撂荒地多为废弃火山岩采石场及近些年荒废的土地, 地表岩石众多, 土壤稀薄, 生境较为恶劣, 在环境筛(如水分、养分)的作用下, 导致群落谱系结构聚集(Helmus & Ives, 2012 )。物种丰富度可能影响撂荒地的PD分布格局, 即群落的物种组成较少, PD也较小。
在本研究中, 尚缺乏对该地区物种功能性状和种群变化等数据的长期监测, 未来拟开展相关研究以探索这类生态学问题。同时, 海南岛作为大陆性岛屿, 其起源引起了广泛的研究兴趣(Zhu, 2016 ), 未来将海南岛的物种地理格局、地质历史、植物系统发育等信息整合在一起, 对该地植物区系构成以及地理分布格局成因进行定量分析, 将可能为海南岛的起源和海南岛植物区系的演化提供更加强有力的证据。
附录1 海口马鞍岭火山口地区的维管植物名录
Appendix 1 List of vascular plant in Ma’anling volcano area of Haikou, China
http://www.biodiversity-science.net/fileup/PDF/2017125-1.pdf
致谢: 在野外调查中得到海南大学洪峰、魏亚情、倪苗、占雷雷、孙梦利等同学的协助, 特此致谢!
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
An SQ Wang ZF Zeng FJ Zhang HD Wang BS 1999 a) Biodiversity of tropical mountane rain forest on Diaoluo Mountain, Hainan.
Acta Scientiarum Naturalium Universitatis Sunyatseni , 38 (6 ), 78 -83 . (in Chinese with English abstract)
[安树青 , 王峥峰 , 曾繁敬 , 张宏达 , 王伯荪 (1999 a) 海南吊罗山热带山地雨林植物种类多样性研究
. 中山大学学报自然科学版 , 38 (6 ), 78 -83 .]
[2]
An SQ Zhu XL Wang ZF Campbell DG Li GQ Chen XL 1999 b) The plant species diversity in a tropical mountane rain forest on Wuzhi Mountain, Hainan.
Acta Ecologica Sinica , 19 , 803 -809 . (in Chinese with English abstract)
[本文引用: 2]
[安树青 , 朱学雷 , 王峥峰 , David G. Campbell 李国旗 , 陈兴龙 (1999 b) 海南五指山热带山地雨林植物物种多样性研究
. 生态学报 , 19 , 803 -809 .]
[本文引用: 2]
[3]
Bai ZD Xu DB Wei HA Hu JC 2003 ) Division of the active period of quaternary volcanism in Ma’anling, Northern Hainan Province.
Seismology and Geology , 25 (z1 ), 12 -20 . (in Chinese with English abstract)
[本文引用: 1]
[白志达 , 徐德斌 , 魏海泉 , 胡久常 (2003 ) 琼北马鞍岭地区第四纪火山活动期次划分
. 地震地质 , 25 (z1 ), 12 -20 .]
[本文引用: 1]
[4]
Berger WH Parker FL 1970 ) Diversity of planktonic Foraminifera in deep-sea sediments.
Science , 168 , 1345 -1347 .
[本文引用: 1]
[5]
Cai H 2011 ) Analysis of the diversity of seed plants in the lava platform of Jingbo Lake.
Journal of Anhui Agricultural Sciences , 39 , 11748 -11749 . (in Chinese with English abstract)
[本文引用: 2]
[蔡赫 (2011 ) 镜泊湖火山熔岩台地种子植物多样性分析
. 安徽农业科学 , 39 , 11748 -11749 .]
[本文引用: 2]
[6]
Compilation Group of China’s Biodiversity: A Country Study(1998 )China’s Biodiversity:A Country Study .China Environmental Science Press , Beijing . (in Chinese)
[本文引用: 1]
[《中国生物多样性国情研究报告》编写组) (1998 ) 中国生物多样性国情研究报告 .中国环境科学出版社 , 北京. ]
[本文引用: 1]
[7]
Devictor V Mouillot D Meynard C Jiguet F Thuiller W Mouquet N 2010 ) Spatial mismatch and congruence between taxonomic phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world.
Ecology Letters , 13 , 1030 -1040 .
[本文引用: 1]
[8]
Editorial Committee of Flora Reipublicae Popularis Sinicae(1959 -2004) Flora Reipublicae Popularis Sinicae,Tomus 2-80 . Science Press ,Beijing . (in Chinese)
[中国科学院中国植物志编辑委员会(1959 -2004) 中国植物志(2-80卷) . 科学出版社 , 北京. ]
[9]
Elliott KJ Knoepp JD 2005 ) The effects of three regeneration harvest methods on plant diversity and soil characteristics in the southern Appalachians.
Forest Ecology and Management , 211 , 296 -317 .
[本文引用: 1]
[10]
Faith DP 1992 ) Conservation evaluation and phylogenetic diversity.
Biological Conservation , 61 , 1 -10 .
[本文引用: 1]
[11]
Fang JY Li YD Zhu B Liu GH Zhou GY 2004 ) Community structures and species richness in the mountane rain forest of Jianfengling, Hainan Island, China.
Biodiversity Science , 12 , 29 -34 . (in Chinese with English abstract)
[本文引用: 1]
[方精云 , 李意德 , 朱彪 , 刘国华 , 周光益 (2004 ) 海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位
. 生物多样性 , 12 , 29 -34 .]
[本文引用: 1]
[12]
Graham HG Fine PVA 2008 ) Phylogenetic beta diversity linking ecological and evolutionary processes across space in time.
Ecology Letters , 11 , 1265 -1277 .
[本文引用: 1]
[13]
Helmus MR Ives AR 2012 ) Phylogenetic diversity—area curves.
Ecology , 93 (Suppl. 8 ), S31 -S43 .
[本文引用: 2]
[14]
Hu YJ Ding XQ 2000 ) A study on the plant species diversity of tropical natural forest in Bawangling, Hainan Island.
Chinese Biodiversity , 8 , 370 -377 . (in Chinese with English abstract)
[本文引用: 1]
[胡玉佳 , 丁小球 (2000 ) 海南岛坝王岭热带天然林植物物种多样性研究
. 生物多样性 , 8 , 370 -377 .]
[本文引用: 1]
[15]
International Union for Conservation of Nature (IUCN) (2013 ) The IUCN Red List of Threatened Species, Version 2013. 1.
2013) The IUCN Red List of Threatened Species, Version 2013. 1. . (accessed on 2013-07-02).
URL
[16]
Ishida H Hattori T Takeda Y 2005 ) Comparison of species composition and richness between primary and secondary lucidophyllous forests in two altitudinal zones of Tsushima Island, Japan.
Forest Ecology and Management , 213 , 273 -287 .
[本文引用: 1]
[17]
Keddy PA Smith L Campbell DR Clark M Montz G 2006 ) Patterns of herbaceous plant diversity in southeastern Louisiana pine savannas.
Applied Vegetation Science , 9 , 17 -26 .
[本文引用: 1]
[18]
Kong FZ Yu RC Xu ZJ Zhou MJ 2012 ) Application of excel in calculation of biodiversity indices.
Marine Sciences , 36 (4 ), 57 -62 . (in Chinese with English abstract)
[本文引用: 1]
[孔凡洲 , 于仁成 , 徐子钧 , 周名江 (2012 ) 应用Excel软件计算生物多样性指数
. 海洋科学 , 36 (4 ), 57 -62 .]
[本文引用: 1]
[19]
Lu SG 2007 ) Pteridology . Higher Eduction Press, Beijing . (in Chinese)
[陆树刚 (2007 ) 蕨类植物学. 高等教育出版社 , 北京. ]
[20]
Madsen K 1999 ) Notes on Chinese-American botanical collaboration.
Arnoldia , 58 , 12 -16 .
[本文引用: 1]
[21]
Ministry of Environmental Protection of the People’s Republic of China and Chinese Academy of Sciences (2013 ) The evaluation of China Biodiversity Red List—Higher Plants
.
URL
[环境保护部和中国科学院 (2013)《中国生物多样性红色名录——高等植物卷.
URL
[22]
Ou ZY Yang XB Wu QS 2007 ) Species diversity of natural forests in the enlarged area of Jianfengling National Nature Reserve, Hainan Island.
Biodiversity Science , 15 , 437 -444 . (in Chinese with English abstract)
[本文引用: 1]
[欧芷阳 , 杨小波 , 吴庆书 (2007 ) 尖峰岭自然保护区扩大区域植物多样性研究
. 生物多样性 , 15 , 437 -444 .]
[本文引用: 1]
[23]
Pielou EC 1966 ) The measurement of diversity in different types of biological collections.
Journal of Theoretical Biology , 13 , 131 -144 .]
[本文引用: 1]
[24]
Qian H Jin Y 2015 ) An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure.
Journal of Plant Ecology , 9 , 233 -239 .
[本文引用: 1]
[25]
Shannon CE 1948 ) A mathematical theory of communication.
The Bell System Technical Journal , 27 (3 ), 379 -423 .
[本文引用: 1]
[26]
South China Botanical Garden,Chinese Academy of Sciences (1987 -2011) Flora of Guangdong, Vols. 1-10 . Guangdong Science and Technology Press , Guangdong. (in Chinese)
[中国科学院华南植物园(1987 -2011)广东植物志(1-10卷) . 广东科技出版社 , 广东. ]
[27]
Sun H Deng T Chen YS Zhou Z 2017 ) Current research and development trends in floristic geography.
Biodiversity Science , 25 , 111 -122 . (in Chinese with English abstract)
[本文引用: 2]
[孙航 , 邓涛 , 陈永生 , 周卓 (2017 ) 植物区系地理研究现状及发展趋势
. 生物多样性 , 25 , 111 -122 .]
[本文引用: 2]
[28]
Wang HS 1992 ) Floristic Geography . Science Press, Beijing . (in Chinese)
[本文引用: 1]
[王荷生 (1992 ) 植物区系地理 . 科学出版社 , 北京. ]
[本文引用: 1]
[29]
Wang QQ Long WX Yang XB Xiong MH Kang Y Huang J Wang X Hong XJ Zhou ZL Lu YQ Fang J Li SX 2016 ) Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island.
Chinese Journal of Plant Ecology , 40 , 469 -479 .
[本文引用: 1]
[王茜茜 , 龙文兴 , 杨小波 , 熊梦辉 , 康勇 , 黄瑾 , 王旭 , 洪小江 , 周照骊 , 陆雍泉 , 方精 , 李时兴 (2016 ) 海南岛3个林区热带云雾林植物多样性变化
. 植物生态学报 , 40 , 469 -479 .]
[本文引用: 1]
[30]
Wang S Xie Y 2004 )China Species Red List (Volume 1) . Higher Education Press , Beijing . (in Chinese)
[本文引用: 1]
[汪松 , 解焱 (2004 ) 中国物种红色名录 (第1卷) . 高等教育出版社 , 北京. ]
[本文引用: 1]
[31]
Webb CO 2000 ) Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
The American Naturalist , 156 , 145 -155 .
[本文引用: 1]
[32]
Webb CO Ackerly D Kembel S 2008 ) Phylocom: software for the analysis of phylogenetic community structure and character evolution.
Bioinformatics , 24 , 2098 -2100 .
[本文引用: 2]
[33]
Wu ZY Raven PH Hong DY 1994 -2014) Flora of China,Vols. 2-25. Science Press, Beijing & Missouri Botanical Gardens, St . Louis , Missouri .
[本文引用: 1]
[34]
Wu ZY Sun H Zhou ZK Li DZ Peng H 2011 ) Floristics of Seed Plants from China . Science Press ,Beijing . (in Chinese)
[本文引用: 1]
[吴征镒 , 孙航 , 周浙昆 , 李德铢 , 彭华 (2011 )中国种子植物区系地理 . 科学出版社 , 北京. ]
[本文引用: 1]
[35]
Wu ZY Zhou ZK Li DZ Peng H Sun H 2003 ) The areal-types of the world families of seed plants.
Acta Botanica Yunnanica , 25 , 245 -257 . (in Chinese with English abstract)
[本文引用: 1]
[吴征镒 , 周浙昆 , 李德铢 , 彭华 , 孙航 (2003 ) 世界种子植物科的分布区类型系统
. 云南植物研究 , 25 , 245 -257 .]
[本文引用: 1]
[36]
Wu ZY Zhou ZK Sun H Li DZ Peng H 2006 ) The Areal-types of Seed Plants and Their Origin and Differentiation . Yunnan Science and Technology Press, Kunming . (in Chinese)
[本文引用: 1]
[吴征镒 , 周浙昆 , 孙航 , 李德铢 , 彭华 (2006 ) 种子植物分布区类型及其起源和分化 . 云南科技出版社 , 昆明. ]
[本文引用: 1]
[37]
Xing FW Zhou JS Wang FG Zeng QW Yi QF Liu DM 2012 ) Inventory of Plant Species Diversity of Hainan . Huazhong University of Science and Technology Press, Wuhan . (in Chinese)
[邢福武 , 周劲松 , 王发国 , 曾庆文 , 易绮斐 , 刘东明 (2012 ) 海南植物物种多样性编目 . 华中科技大学出版社 , 武汉. ]
[38]
Xu YJ Lin DM Shi M Xie YJ Wang YZ Guan ZH Xiang JY 2017 ) Spatial heterogeneity and its causes in evergreen broad-leaved forests in the Ailao Mountains, Yunnan Province.
Biodiversity Science , 25 , 23 -33 . (in Chinese with English abstract)
[本文引用: 1]
[徐远杰 , 林敦梅 , 石明 , 谢妍洁 , 王逸之 , 管振华 , 向建英 (2017 ) 云南哀牢山常绿阔叶林的空间分异及其影响因素
. 生物多样性 , 25 , 23 -33 .]
[本文引用: 1]
[39]
Yang XB 2016 )Checklist of Plant Species in Hainan . Science Press ,Beijing . (in Chinese)
[杨小波 (2016 ) 海南植物图志. 科学出版社 , 北京. ]
[40]
Yang XB Lin Y Liang SQ 1994 ) The forest vegetation of Wu-zhi Shan Mountain in Hainan Island. II. The plant population analysis and forest structures analysis of the forest vegetation of Wu-zhi Shan Mountain in Hainan Island.
Natural Science Journal of Hainan University , 12 , 312 -323 . (in Chinese with English abstract)
[本文引用: 2]
[杨小波 , 林英 , 梁淑群 (1994 ) 海南岛五指山的森林植被II. 五指山森林植被的植物种群分析与森林结构分析
. 海南大学学报 , 12 , 311 -323 .]
[本文引用: 2]
[41]
Yang XB Wu QS Li DH 2011 ) Hainan Island The Forest Vegetation of National Forest-ecosystem Nature Reserves , Science Press , Beijing . (in Chinese)
[本文引用: 1]
[杨小波 , 吴庆书 , 李东海 (2011 )海南岛陆域国家级森林生态系统自然保护区森林植被研究 . 科学出版社 , 北京. ]
[本文引用: 1]
[42]
Yu SX Zang RG Jiang YX 2001 ) Species richness-abundance relationships in four tropical forests on altitudinal gradient in Bawangling Nature Reserve, Hainan.
Acta Phytoecologica Sinica , 25 , 291 -297 . (in Chinese with English abstract)
[本文引用: 1]
[余世孝 , 臧润国 , 蒋有绪 (2001 ) 海南岛霸王岭不同热带森林类型的种-个体关系
. 植物生态学报 , 25 , 291 -297 .]
[本文引用: 1]
[43]
Zang RG Ding Y 2009 ) Forest recovery on abandoned logging roads in a tropical montane rain forest of Hainan Island, China.
Acta Oecologica , 35 , 462 -470 .
[本文引用: 1]
[44]
Zanne AE Tank DC Cornwell WK Eastman JM Smith SA FitzJohn RG McGlinn DJ O’Meara BC Moles AT Reich PB Royer DL Soltis DE Stevens PF Westoby M Wright IJ Aarssen L Bertin RI Calaminus A Govaerts R Hemmings F Leishman MR Oleksyn J Soltis PS Swenson NG Warman L Beaulieu JM 2014 ) Three keys to the radiation of angiosperms into freezing environments.
Nature , 506 , 89 -92 .
[45]
Zhang YB Ma KP 2008 ) Geographic distribution patterns and status assessment of threatened plants in China.
Biodiversity and Conservation , 17 , 1783 -1798 .
[本文引用: 1]
[46]
Zhong L Chang-Yang CH Lu P Gu XP Lei ZP Cai YB Zheng FD Sun IF Yu MJ 2015 ) Community structure and species composition of the secondary evergreen broad-leaved forest: the analyses for a 9 ha forest dynamics plot in Wuyanling Nature Reserve, Zhejiang Province.
Biodiversity Science , 23 , 619 -629 .
[本文引用: 1]
(in Chinese with English abstract) [仲磊 , 张杨家豪 , 卢品 , 顾雪萍 , 雷祖培 , 蔡延奔 , 郑方东 , 孙义方 , 于明坚 (2015 ) 次生常绿阔叶林的群落结构与物种组成: 基于浙江乌岩岭9 ha森林动态样地. 生物多样性,
23 , 619 -629 .]
[本文引用: 1]
[47]
Zhu H 2016 ) Biogeographical evidences help revealing the origin of Hainan Island.
PLoS ONE , 11 , e0151941 .
[本文引用: 4]
a) 海南吊罗山热带山地雨林植物种类多样性研究
1999
a) 海南吊罗山热带山地雨林植物种类多样性研究
1999
b) 海南五指山热带山地雨林植物物种多样性研究
2
1999
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
... ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
b) 海南五指山热带山地雨林植物物种多样性研究
2
1999
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
... ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
琼北马鞍岭地区第四纪火山活动期次划分
1
2003
... 马鞍岭火山口地区指海口市西南石山、永兴镇一带, 西自美造水库, 东至龙桥, 南起罗京盘, 北迄长流的全新世火山岩分布区, 地理坐标为19°48′-20°01′ N, 110°06′-110°27′ E, 面积约500 km² (白志达等, 2003 ).该区域处于热带与南亚热带过渡区, 年均温23.7℃, 年积温8,700℃, 最热月7月均温为29-31℃, 最冷月1月均温17.2℃, 偶尔低至0℃, 全年暖热无冬; 雨量充沛, 年降水量1,685 mm左右, 且多集中于夏秋两季, 雨季常有台风侵袭; 相对湿度约85%, 全年日照时数达2,240 h.气候十分适于热带及亚热带植物生长, 具有我国热带及向南亚热带过渡生物群落的典型特征. ...
琼北马鞍岭地区第四纪火山活动期次划分
1
2003
... 马鞍岭火山口地区指海口市西南石山、永兴镇一带, 西自美造水库, 东至龙桥, 南起罗京盘, 北迄长流的全新世火山岩分布区, 地理坐标为19°48′-20°01′ N, 110°06′-110°27′ E, 面积约500 km² (白志达等, 2003 ).该区域处于热带与南亚热带过渡区, 年均温23.7℃, 年积温8,700℃, 最热月7月均温为29-31℃, 最冷月1月均温17.2℃, 偶尔低至0℃, 全年暖热无冬; 雨量充沛, 年降水量1,685 mm左右, 且多集中于夏秋两季, 雨季常有台风侵袭; 相对湿度约85%, 全年日照时数达2,240 h.气候十分适于热带及亚热带植物生长, 具有我国热带及向南亚热带过渡生物群落的典型特征. ...
Diversity of planktonic Foraminifera in deep-sea sediments.
1
1970
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
镜泊湖火山熔岩台地种子植物多样性分析
2
2011
... 马鞍岭火山口及周边地区是我国唯一的热带海岛城市火山群地质公园, 也是海南省第一家由联合国教科文组织确认具有重要价值的国家4A级旅游景区.但其详尽的植物多样性研究仍是空白.目前, 有关我国火山地区植物区系的研究只有黑龙江五大连池火山自然保护区的植物区系调查研究( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.)和黑龙江省宁安市境内镜泊湖熔岩台地的种子植物多样性研究(蔡赫, 2011 )等, 与本文调查区域在气候条件、人文地理、物种组成和植物区系地理方面均显著不同.为进一步明确马鞍岭火山口及周边维管植物的现状, 我们于2016年12月至2017年3月底开展了本次调查.结合相关文献资料及植物标本, 对马鞍岭火山口地区的植物种类进行鉴定统计, 对不同土地利用类型(人工林、次生林和撂荒地)中的植物多样性及植物区系基本特征、组成和可能的起源与演化历史进行了分析, 拟为该地区植物区系研究提供参考. ...
... 在我国的火山地区植物区系中, 马鞍岭火山口地区单位面积物种数较多且保护植物的占比较高.在本研究的100 km2 范围内, 我们共发现维管植物284种, 单位陆地面积的种数高达2.84种/km2 .这远高于其他火山植物区系, 如黑龙江五大连池火山自然保护区占地面积1,060 km2 , 有野生植物143科428属l,044种( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.), 单位面积的物种数为0.98种/km2 .黑龙江省宁安市境内镜泊湖熔岩台地占地面积达1,200 km2 , 有种子植物77科265属486种(蔡赫, 2011 ), 单位面积的物种数仅0.41种/km2 . ...
镜泊湖火山熔岩台地种子植物多样性分析
2
2011
... 马鞍岭火山口及周边地区是我国唯一的热带海岛城市火山群地质公园, 也是海南省第一家由联合国教科文组织确认具有重要价值的国家4A级旅游景区.但其详尽的植物多样性研究仍是空白.目前, 有关我国火山地区植物区系的研究只有黑龙江五大连池火山自然保护区的植物区系调查研究( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.)和黑龙江省宁安市境内镜泊湖熔岩台地的种子植物多样性研究(蔡赫, 2011 )等, 与本文调查区域在气候条件、人文地理、物种组成和植物区系地理方面均显著不同.为进一步明确马鞍岭火山口及周边维管植物的现状, 我们于2016年12月至2017年3月底开展了本次调查.结合相关文献资料及植物标本, 对马鞍岭火山口地区的植物种类进行鉴定统计, 对不同土地利用类型(人工林、次生林和撂荒地)中的植物多样性及植物区系基本特征、组成和可能的起源与演化历史进行了分析, 拟为该地区植物区系研究提供参考. ...
... 在我国的火山地区植物区系中, 马鞍岭火山口地区单位面积物种数较多且保护植物的占比较高.在本研究的100 km2 范围内, 我们共发现维管植物284种, 单位陆地面积的种数高达2.84种/km2 .这远高于其他火山植物区系, 如黑龙江五大连池火山自然保护区占地面积1,060 km2 , 有野生植物143科428属l,044种( 刘艳华 (2007) 五大连池火山保护区野生植物资源调查研究. 硕士学位论文, 延边大学, 吉林延吉.), 单位面积的物种数为0.98种/km2 .黑龙江省宁安市境内镜泊湖熔岩台地占地面积达1,200 km2 , 有种子植物77科265属486种(蔡赫, 2011 ), 单位面积的物种数仅0.41种/km2 . ...
1
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
1
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
Spatial mismatch and congruence between taxonomic phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world.
1
2010
... 本研究发现, 人工林的PD、NTAXA、MPD、MNTD值最大, 表明人工林群落中的物种最丰富, 但物种相互之间的亲缘关系疏远, 因而谱系结构呈现发散状态.这可能跟人工林(主要是荔枝、龙眼、菠萝蜜和大叶桉林)在前期较长时间的人工干预条件下保留了较多果树等经济作物, 而后在较短的时间内缺乏人为干扰, 较多的草本及灌木大量生长有关.先前的人工干预导致林冠茂密, 而后自然生长的其他乔木、灌木及草本可获取的阳光、矿物质等资源有限, 种间竞争激烈, 物种在资源利用策略上倾向于差异化(Devictor et al, 2010 ; Helmus & Ives, 2012 ).这样先经过人工过度干预而后长期的自然生长导致人工林出现群落物种间亲缘关系疏远而谱系结构发散的特征.次生林的PD、NTAXA、MPD、MNTD值较大, 反映次生林群落中的物种较丰富, 物种间亲缘关系较近, 谱系结构相对聚集.这可能跟次生林的形成过程及环境有关, 马鞍岭的次生林发生在被大面积严重砍伐的区域, 主要由植物在较短的时间范围内自然生长形成.因发育时间短, 加之该地的表土多为火山岩, 土壤较少, 故难以形成发育程度较好的次生林, 多为低矮灌木, 少有小乔木及藤本.这就形成了马鞍岭次生林相对较近的物种间亲缘关系及相对聚集的谱系结构.撂荒地的PD、NTAXA、MPD、MNTD值最小, 这说明其物种相对单一, 群落中物种间的亲缘关系最近和谱系结构最为聚集.可能由于马鞍岭的撂荒地多为废弃火山岩采石场及近些年荒废的土地, 地表岩石众多, 土壤稀薄, 生境较为恶劣, 在环境筛(如水分、养分)的作用下, 导致群落谱系结构聚集(Helmus & Ives, 2012 ).物种丰富度可能影响撂荒地的PD分布格局, 即群落的物种组成较少, PD也较小. ...
The effects of three regeneration harvest methods on plant diversity and soil characteristics in the southern Appalachians.
1
2005
... 植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 ).其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面.植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成. ...
Conservation evaluation and phylogenetic diversity.
1
1992
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位
1
2004
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位
1
2004
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
Phylogenetic beta diversity linking ecological and evolutionary processes across space in time.
1
2008
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
Phylogenetic diversity—area curves.
2
2012
... 本研究发现, 人工林的PD、NTAXA、MPD、MNTD值最大, 表明人工林群落中的物种最丰富, 但物种相互之间的亲缘关系疏远, 因而谱系结构呈现发散状态.这可能跟人工林(主要是荔枝、龙眼、菠萝蜜和大叶桉林)在前期较长时间的人工干预条件下保留了较多果树等经济作物, 而后在较短的时间内缺乏人为干扰, 较多的草本及灌木大量生长有关.先前的人工干预导致林冠茂密, 而后自然生长的其他乔木、灌木及草本可获取的阳光、矿物质等资源有限, 种间竞争激烈, 物种在资源利用策略上倾向于差异化(Devictor et al, 2010 ; Helmus & Ives, 2012 ).这样先经过人工过度干预而后长期的自然生长导致人工林出现群落物种间亲缘关系疏远而谱系结构发散的特征.次生林的PD、NTAXA、MPD、MNTD值较大, 反映次生林群落中的物种较丰富, 物种间亲缘关系较近, 谱系结构相对聚集.这可能跟次生林的形成过程及环境有关, 马鞍岭的次生林发生在被大面积严重砍伐的区域, 主要由植物在较短的时间范围内自然生长形成.因发育时间短, 加之该地的表土多为火山岩, 土壤较少, 故难以形成发育程度较好的次生林, 多为低矮灌木, 少有小乔木及藤本.这就形成了马鞍岭次生林相对较近的物种间亲缘关系及相对聚集的谱系结构.撂荒地的PD、NTAXA、MPD、MNTD值最小, 这说明其物种相对单一, 群落中物种间的亲缘关系最近和谱系结构最为聚集.可能由于马鞍岭的撂荒地多为废弃火山岩采石场及近些年荒废的土地, 地表岩石众多, 土壤稀薄, 生境较为恶劣, 在环境筛(如水分、养分)的作用下, 导致群落谱系结构聚集(Helmus & Ives, 2012 ).物种丰富度可能影响撂荒地的PD分布格局, 即群落的物种组成较少, PD也较小. ...
... ).这样先经过人工过度干预而后长期的自然生长导致人工林出现群落物种间亲缘关系疏远而谱系结构发散的特征.次生林的PD、NTAXA、MPD、MNTD值较大, 反映次生林群落中的物种较丰富, 物种间亲缘关系较近, 谱系结构相对聚集.这可能跟次生林的形成过程及环境有关, 马鞍岭的次生林发生在被大面积严重砍伐的区域, 主要由植物在较短的时间范围内自然生长形成.因发育时间短, 加之该地的表土多为火山岩, 土壤较少, 故难以形成发育程度较好的次生林, 多为低矮灌木, 少有小乔木及藤本.这就形成了马鞍岭次生林相对较近的物种间亲缘关系及相对聚集的谱系结构.撂荒地的PD、NTAXA、MPD、MNTD值最小, 这说明其物种相对单一, 群落中物种间的亲缘关系最近和谱系结构最为聚集.可能由于马鞍岭的撂荒地多为废弃火山岩采石场及近些年荒废的土地, 地表岩石众多, 土壤稀薄, 生境较为恶劣, 在环境筛(如水分、养分)的作用下, 导致群落谱系结构聚集(Helmus & Ives, 2012 ).物种丰富度可能影响撂荒地的PD分布格局, 即群落的物种组成较少, PD也较小. ...
海南岛坝王岭热带天然林植物物种多样性研究
1
2000
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
海南岛坝王岭热带天然林植物物种多样性研究
1
2000
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
International Union for Conservation of Nature (IUCN) (2013 ) The IUCN Red List of Threatened Species, Version 2013. 1.
Comparison of species composition and richness between primary and secondary lucidophyllous forests in two altitudinal zones of Tsushima Island, Japan.
1
2005
... 植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 ).其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面.植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成. ...
Patterns of herbaceous plant diversity in southeastern Louisiana pine savannas.
1
2006
... 植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 ).其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面.植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成. ...
应用Excel软件计算生物多样性指数
1
2012
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
应用Excel软件计算生物多样性指数
1
2012
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
Notes on Chinese-American botanical collaboration.
1
1999
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
环境保护部和中国科学院 (2013)《中国生物多样性红色名录——高等植物卷.
环境保护部和中国科学院 (2013)《中国生物多样性红色名录——高等植物卷.
尖峰岭自然保护区扩大区域植物多样性研究
1
2007
... 植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 ).其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面.植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成. ...
尖峰岭自然保护区扩大区域植物多样性研究
1
2007
... 植物多样性常被用来测度某个区域植物资源的丰富程度, 其中物种总数、物种密度和特有种比例是较为重要的3个测量指标(欧芷阳等, 2007 ).其研究内容包括不同植被类型植物多样性的特征(Keddy et al, 2006 )、相邻区域的比较(Ishida et al, 2005 )、不同干扰方式和强度对物种多样性的影响(Elliott & Knoepp, 2005 )等方面.植物多样性和植物区系地理是植物生态学研究中的两个重要方面, 两者相辅相成. ...
The measurement of diversity in different types of biological collections.
1
1966
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure.
1
2015
... Phyto谱系树是Qian和Jin (2015)在Zanne等(2014)的谱系树基础上进一步更新、扩展而成.基于样地调查所获的物种数据, 在Qian和Jin (2015)的谱系树基础上, 利用S. Phylo Maker函数以3种不同方式添加物种构建本文的谱系树(如, Scenario 1, Scenario 2, Scenario 3).我们选用的是第3种(Scenario 3), 该谱系树将Phyto中没有的物种以多分支形态添加到其父母分类单元下, 用BLADJ算法为其分配支长(Webb et al, 2008 ; Qian & Jin, 2015 ). ...
A mathematical theory of communication.
1
1948
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
植物区系地理研究现状及发展趋势
2
2017
... 植物区系地理学(floristic geography)是主要研究全球或某一区域植物种类的构成、现代和过去的分布规律以及起源进化、演变历史的科学(王荷生, 1992 ).植物区系地理格局的形成建立在明确植物区系分布区类型形成和起源的基础之上.积累植物区系的基本特征等方面的研究资料, 有利于对植物多样性的基础理论问题开展深入研究, 并为生物多样性的保护提供理论依据和实践指导(孙航等, 2017 ). ...
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
植物区系地理研究现状及发展趋势
2
2017
... 植物区系地理学(floristic geography)是主要研究全球或某一区域植物种类的构成、现代和过去的分布规律以及起源进化、演变历史的科学(王荷生, 1992 ).植物区系地理格局的形成建立在明确植物区系分布区类型形成和起源的基础之上.积累植物区系的基本特征等方面的研究资料, 有利于对植物多样性的基础理论问题开展深入研究, 并为生物多样性的保护提供理论依据和实践指导(孙航等, 2017 ). ...
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
1
1992
... 植物区系地理学(floristic geography)是主要研究全球或某一区域植物种类的构成、现代和过去的分布规律以及起源进化、演变历史的科学(王荷生, 1992 ).植物区系地理格局的形成建立在明确植物区系分布区类型形成和起源的基础之上.积累植物区系的基本特征等方面的研究资料, 有利于对植物多样性的基础理论问题开展深入研究, 并为生物多样性的保护提供理论依据和实践指导(孙航等, 2017 ). ...
1
1992
... 植物区系地理学(floristic geography)是主要研究全球或某一区域植物种类的构成、现代和过去的分布规律以及起源进化、演变历史的科学(王荷生, 1992 ).植物区系地理格局的形成建立在明确植物区系分布区类型形成和起源的基础之上.积累植物区系的基本特征等方面的研究资料, 有利于对植物多样性的基础理论问题开展深入研究, 并为生物多样性的保护提供理论依据和实践指导(孙航等, 2017 ). ...
海南岛3个林区热带云雾林植物多样性变化
1
2016
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
海南岛3个林区热带云雾林植物多样性变化
1
2016
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
1
2004
... 本地区有珍稀濒危植物4种, 分别为: 土沉香(Aquilaria sinensis )、降香(Dalbergia odorifera )、海南龙血树(Dracaena cambodiana )和荔枝.其中土沉香和降香被《中国物种红色名录》(汪松和解焱, 2004 )列为易危和极危物种, 也被《IUCN红色名录(2013)》列为易危物种, 被《国家重点保护植物名录》列为国家二级保护植物.此外, 土沉香还被《濒危野生动植物种国际贸易公约》附录II收录.《国家重点保护植物名录》将海南龙血树列为易危物种, 将荔枝列为濒危物种. ...
1
2004
... 本地区有珍稀濒危植物4种, 分别为: 土沉香(Aquilaria sinensis )、降香(Dalbergia odorifera )、海南龙血树(Dracaena cambodiana )和荔枝.其中土沉香和降香被《中国物种红色名录》(汪松和解焱, 2004 )列为易危和极危物种, 也被《IUCN红色名录(2013)》列为易危物种, 被《国家重点保护植物名录》列为国家二级保护植物.此外, 土沉香还被《濒危野生动植物种国际贸易公约》附录II收录.《国家重点保护植物名录》将海南龙血树列为易危物种, 将荔枝列为濒危物种. ...
Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
1
2000
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
Phylocom: software for the analysis of phylogenetic community structure and character evolution.
2
2008
... Phyto谱系树是Qian和Jin (2015)在Zanne等(2014)的谱系树基础上进一步更新、扩展而成.基于样地调查所获的物种数据, 在Qian和Jin (2015)的谱系树基础上, 利用S. Phylo Maker函数以3种不同方式添加物种构建本文的谱系树(如, Scenario 1, Scenario 2, Scenario 3).我们选用的是第3种(Scenario 3), 该谱系树将Phyto中没有的物种以多分支形态添加到其父母分类单元下, 用BLADJ算法为其分配支长(Webb et al, 2008 ; Qian & Jin, 2015 ). ...
... 用物种数量(number of taxa, NTAXA)、种间平均成对谱系距离(mean phylogenetic distance, MPD)、平均最近相邻谱系距离(mean nearest phylogenetic taxon distance, MNTD)和Faith谱系多样性(phylogenetic distance, PD) (Faith, 1992 )表示α谱系多样性.PD是某一样地中分类单元谱系分支长度占谱系中所有分支长度之和的比例.MPD是群落中全部物种对间的平均进化距离, 可以很好地表示群落中物种的整体聚集程度(Webb, 2000 ), 是描述群落谱系多样性的基础指标.MNTD是任一物种与群落中和自己亲缘关系最近的一个物种间的平均进化距离, 反映群落中物种的局部聚集程度(王茜茜等, 2016 ).PD由Phylocom 4.2 (Webb et al, 2008 )计算, MPD、MNTD由R 3.2.2 Picante软件包中的mpd、mntd函数计算.β谱系多样性用来反映群落间亲缘关系的相异性, 其描述的是同一时间处于不同位置的两个群落间的谱系距离(Graham & Fine, 2008 ).本文数据分析采用SPSS 18.0软件实现. ...
1
1994
... 依据植物志及Flora of China (Wu et al, 1994- 2014 )对物种进行分类统计, 采用“Biodixcel.xlsx”程序(孔凡洲等, 2012 )计算得到Berger-Parker指数d (Berger & Parker, 1970 )、Shannon指数H' ( Shannon, 1948 )及Pielou均匀度指数Je (Pielou, 1966 ) 等3个物种多样性指数. ...
1
2011
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
1
2011
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
世界种子植物科的分布区类型系统
1
2003
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
世界种子植物科的分布区类型系统
1
2003
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
1
2006
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
1
2006
... 依据修订的APG III系统、Theplantlist (www.theplantlist.org/)、Flora of China (Wu et al, 1994-2014)对马鞍岭种子植物区系的科进行统计及更新.按世界种子植物科的分布区类型(吴征镒等, 2003 )和中国种子植物属分布区类型(吴征镒等, 2006 , 2011 )统计马鞍岭植物区系种子植物科属的分布区类型构成.蕨类植物分布区的界定参照陆树刚(2007)关于蕨类植物区系的划分方法. ...
云南哀牢山常绿阔叶林的空间分异及其影响因素
1
2017
... 不同土地利用类型中不同生活型的优势度都较高, 这说明优势种较显著, 特别是人工林的乔木优势度与撂荒地之间差异显著; 群落多样性结果显示不同土地利用类型中, 乔木的H' 差异显著, 这可能与该地的投入肥料和虫害管理导致土壤肥力不同(徐远杰等, 2017 )有关.马鞍岭火山口地区的人工林主要是荔枝、龙眼等乔木类的果园或者其他经济林, 这是导致人工林与撂荒地之间乔木的优势度差异显著及人工林、次生林、撂荒地的乔木群落多样性差异显著的主要原因.绝大多数的人工林在建林后的数十年间缺乏管理, 导致灌木和草本的优势度和群落多样性与次生林和撂荒地的差异不显著(仲磊等, 2015 ). ...
云南哀牢山常绿阔叶林的空间分异及其影响因素
1
2017
... 不同土地利用类型中不同生活型的优势度都较高, 这说明优势种较显著, 特别是人工林的乔木优势度与撂荒地之间差异显著; 群落多样性结果显示不同土地利用类型中, 乔木的H' 差异显著, 这可能与该地的投入肥料和虫害管理导致土壤肥力不同(徐远杰等, 2017 )有关.马鞍岭火山口地区的人工林主要是荔枝、龙眼等乔木类的果园或者其他经济林, 这是导致人工林与撂荒地之间乔木的优势度差异显著及人工林、次生林、撂荒地的乔木群落多样性差异显著的主要原因.绝大多数的人工林在建林后的数十年间缺乏管理, 导致灌木和草本的优势度和群落多样性与次生林和撂荒地的差异不显著(仲磊等, 2015 ). ...
海南岛五指山的森林植被II. 五指山森林植被的植物种群分析与森林结构分析
2
1994
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
... , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
海南岛五指山的森林植被II. 五指山森林植被的植物种群分析与森林结构分析
2
1994
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
... , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
1
2011
... 海口马鞍岭火山口地区植物属的分布区类型与铜鼓岭自然保护区类似, 均以泛热带分布属为主(杨小波等, 2011 ); 跟以热带亚洲成分为主的五指山、尖峰岭热带雨林不同.马鞍岭火山口地区没有中国特有属, 铜鼓岭只有1个, 而吊罗山、尖峰岭、五指山自然保护区分别有10、15、16个.这一方面可能是马鞍岭地区和铜鼓岭自然保护区受人为影响严重, 另一方面它们与另外三者地理及环境存有较大差异.吊罗山、尖峰岭、五指山自然保护区海拔均在1,200 m以上, 海拔跨度较大, 年平均气温比铜鼓岭及马鞍岭火山口偏低, 所以植物的分布区类型有所不同, 但均以热带区系为主.且这3个自然保护区的面积相对较广, 人为干扰较少, 物种保护较好, 而马鞍岭火山口和铜鼓岭海拔低, 年平均气温偏高, 人为干扰严重, 导致物种丰富度及特有性比其他3个保护区低, 且物种存在进一步减少的可能. ...
1
2011
... 海口马鞍岭火山口地区植物属的分布区类型与铜鼓岭自然保护区类似, 均以泛热带分布属为主(杨小波等, 2011 ); 跟以热带亚洲成分为主的五指山、尖峰岭热带雨林不同.马鞍岭火山口地区没有中国特有属, 铜鼓岭只有1个, 而吊罗山、尖峰岭、五指山自然保护区分别有10、15、16个.这一方面可能是马鞍岭地区和铜鼓岭自然保护区受人为影响严重, 另一方面它们与另外三者地理及环境存有较大差异.吊罗山、尖峰岭、五指山自然保护区海拔均在1,200 m以上, 海拔跨度较大, 年平均气温比铜鼓岭及马鞍岭火山口偏低, 所以植物的分布区类型有所不同, 但均以热带区系为主.且这3个自然保护区的面积相对较广, 人为干扰较少, 物种保护较好, 而马鞍岭火山口和铜鼓岭海拔低, 年平均气温偏高, 人为干扰严重, 导致物种丰富度及特有性比其他3个保护区低, 且物种存在进一步减少的可能. ...
海南岛霸王岭不同热带森林类型的种-个体关系
1
2001
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
海南岛霸王岭不同热带森林类型的种-个体关系
1
2001
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
Forest recovery on abandoned logging roads in a tropical montane rain forest of Hainan Island, China.
1
2009
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
Three keys to the radiation of angiosperms into freezing environments.
2014
Geographic distribution patterns and status assessment of threatened plants in China.
1
2008
... 海南岛地处中国热带地区, 是我国第二大岛屿, 生物多样性丰富, 其中南部地区被列入中国35个生物多样性优先保护区域, 具有较高的保护价值 (《中国生物多样性国情研究报告》编写组, 1998 ; Zhang & Ma, 2008; Zang & Ding, 2009 ; IUCN, 2013 ).海南植物多样性的调查工作开展得较早, 大约230年前, 瑞典人Andrea Dahl已在此地进行植物采集和研究工作; 约100年前, 我国著名植物学家陈焕镛也在海南开始了植物的野外采集和研究(Madsen, 1999 ).整个海南岛及其周边岛礁范围内也进行了很多植物区系的研究工作.《中国植物志》(中国植物志编辑委员会, 1959-2004)记录海南岛的维管植物约3,500种; 《广东植物志》(中国科学院华南植物园, 1987-2011)记录4,196种; 邢福武等(2012)记录了5,108种分布在海南岛及其附属南海岛礁的维管植物; 杨小波(2016)记录的海南岛维管束植物种类达6,036种, 隶属234科1,895属.这些研究基本弄清了我国海南植物区系的组成、地理格局等相关问题.此外, 对海南岛局部区域如尖峰岭(方精云等, 2004 )、霸王岭(胡玉佳和丁小球, 2000 ; 余世孝等, 2001 )、吊罗山(安树青等, 1999a )、五指山(杨小波等, 1994a , b ; 安树青等, 1999b )等地的植被结构及植物多样性也有很多报道.虽然前人对海南植物的研究做了大量调查工作, 但多数研究仍然停留在较为基础的阶段, 研究方法传统, 缺乏整合物种地理格局、环境因子、地质历史、系统发育等方面的信息, 也缺乏定量分析, 未对区系构成以及地理分布格局的成因进行探究, 甚至还有不少区域尚未开展植物区系的调查研究(孙航等, 2017 ). ...
次生常绿阔叶林的群落结构与物种组成: 基于浙江乌岩岭9 ha森林动态样地. 生物多样性,
1
2015
... 不同土地利用类型中不同生活型的优势度都较高, 这说明优势种较显著, 特别是人工林的乔木优势度与撂荒地之间差异显著; 群落多样性结果显示不同土地利用类型中, 乔木的H' 差异显著, 这可能与该地的投入肥料和虫害管理导致土壤肥力不同(徐远杰等, 2017 )有关.马鞍岭火山口地区的人工林主要是荔枝、龙眼等乔木类的果园或者其他经济林, 这是导致人工林与撂荒地之间乔木的优势度差异显著及人工林、次生林、撂荒地的乔木群落多样性差异显著的主要原因.绝大多数的人工林在建林后的数十年间缺乏管理, 导致灌木和草本的优势度和群落多样性与次生林和撂荒地的差异不显著(仲磊等, 2015 ). ...
次生常绿阔叶林的群落结构与物种组成: 基于浙江乌岩岭9 ha森林动态样地. 生物多样性,
1
2015
... 不同土地利用类型中不同生活型的优势度都较高, 这说明优势种较显著, 特别是人工林的乔木优势度与撂荒地之间差异显著; 群落多样性结果显示不同土地利用类型中, 乔木的H' 差异显著, 这可能与该地的投入肥料和虫害管理导致土壤肥力不同(徐远杰等, 2017 )有关.马鞍岭火山口地区的人工林主要是荔枝、龙眼等乔木类的果园或者其他经济林, 这是导致人工林与撂荒地之间乔木的优势度差异显著及人工林、次生林、撂荒地的乔木群落多样性差异显著的主要原因.绝大多数的人工林在建林后的数十年间缺乏管理, 导致灌木和草本的优势度和群落多样性与次生林和撂荒地的差异不显著(仲磊等, 2015 ). ...
Biogeographical evidences help revealing the origin of Hainan Island.
4
2016
... Zhu (2016) 将海南岛的植物区系同我国广西、广东以及越南的植物区系进行比较, 发现海南的植物区系与越南的最为相似, 其次是广西.海南植物区系的特异性十分低, 特有属仅7个, 特有种只有10%左右, 表明其大陆起源的特征. ...
... 对比越南以及我国广东、广西的植物区系发现, 马鞍岭火山口地区的植物区系跟广东的相似性最大, 其次是越南; 在所比较的164个属的本地植物中, 越南、广东、广西都仅有1个属无分布, 分别是白饭树属(Flueggea )、龙血树属(Dracaena) 、牛筋果属(Harrisonia ), 这在一定程度上显示了马鞍岭火山口植物大陆起源的特征, 和Zhu (2016)的结果类似.但在种的层面上, 马鞍岭火山口地区的194种本地植物中, 仅7种不在广东分布, 12种不在越南分布, 14种不在广西分布, 仅有水仙柯(Lithocarpus naiadarum )在3个地区都不分布, 表明该地植物特有性非常低.这与Zhu (2016) 的结果有一定差异.一方面可能跟马鞍岭火山口植被的形成有关: 马鞍岭火山口最后一次喷发在距今约1万年前, 火山喷发可能彻底毁灭了先前的植被, 再形成了现在的植被形态, 相比整个海南岛的植物区系来说, 马鞍岭火山口地区的植物区系的形成时间更年轻, 并在之后的时间里跟地理位置更近的雷州半岛之间的物种交流更频繁, 导致与广东植物区系更相近; 另一方面本次调查的范围相比整个海南岛而言面积小, 物种数相对少, 且环境一致, 可能导致与Zhu (2016) 的研究结果有偏差. ...
... 的结果有一定差异.一方面可能跟马鞍岭火山口植被的形成有关: 马鞍岭火山口最后一次喷发在距今约1万年前, 火山喷发可能彻底毁灭了先前的植被, 再形成了现在的植被形态, 相比整个海南岛的植物区系来说, 马鞍岭火山口地区的植物区系的形成时间更年轻, 并在之后的时间里跟地理位置更近的雷州半岛之间的物种交流更频繁, 导致与广东植物区系更相近; 另一方面本次调查的范围相比整个海南岛而言面积小, 物种数相对少, 且环境一致, 可能导致与Zhu (2016) 的研究结果有偏差. ...
... 在本研究中, 尚缺乏对该地区物种功能性状和种群变化等数据的长期监测, 未来拟开展相关研究以探索这类生态学问题.同时, 海南岛作为大陆性岛屿, 其起源引起了广泛的研究兴趣(Zhu, 2016 ), 未来将海南岛的物种地理格局、地质历史、植物系统发育等信息整合在一起, 对该地植物区系构成以及地理分布格局成因进行定量分析, 将可能为海南岛的起源和海南岛植物区系的演化提供更加强有力的证据. ...



{kind=link}
{kind=link}