青藏高原东北部边缘地区在地理上包括甘肃省大部分区域、青海省东部和北部以及四川省北部, 面积约469,000 km2 , 地貌复杂, 气候各异, 是青藏高原、黄土高原、蒙新高原的交汇地带。水系为青藏高原水系的一部分。由南向北主要分为: 长江流域、黄河流域、青海湖流域、河西内陆河流域。我国三大动物地理区(古北区、东洋区、青藏高原区)也在此交汇(陈宜瑜等, 1996 )。目前认为, 青藏高原东北部边缘可能是一个重要的冰河期避难所和目前一些广泛分布在高原的特有物种遗传多样性较高的地区(赵凯, 2009 )。
高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 )。与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 )。然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究。
青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 )。尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待。朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作。同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作。鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料。
1 材料与方法
1.1 研究区域
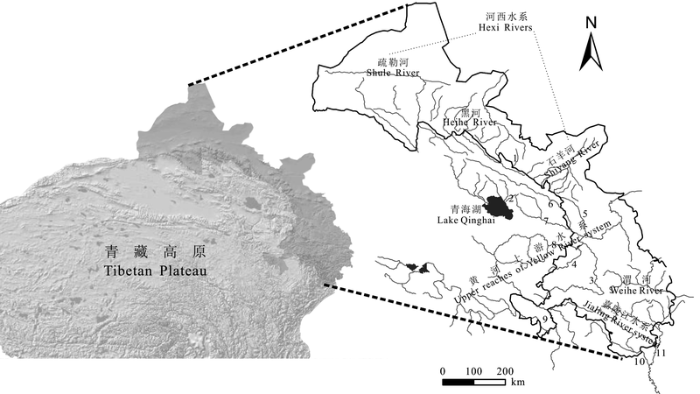
研究区域位于32°14'-40°36' N, 94°19'-106°08' E之间; 跨8个纬度, 12个经度。海拔范围600-4,300 m。调查区域覆盖了青藏高原东北部边缘地区各大水体。依据水系演化关系和水系间鱼类组成的差异, 将研究区域分为嘉陵江水系(JLJ)、黄河上游水系、渭河水系(WH)、青海湖水系(QHH)、河西水系; 其中, 黄河上游水系进一步分为贵德以下兰州段(YH1)和以上段(YH2), 河西水系包括石羊河(SY)、黑河(HH)和疏勒河(SL) 3个水系单元(图1 )。
图1 调查区域水系图示。右图灰色线中, 1: 大马营河; 2: 甘子河; 3: 洮河; 4: 大夏河; 5: 庄浪河; 6: 大通河; 7: 湟水河; 8: 贵德; 9: 玛曲; 10: 白龙江; 11: 嘉陵江上游。
Fig. 1 Drainage systems of investigation area. Gray lines in right part: 1, Damaying River; 2, Ganzi River; 3, Taohe River; 4, Daxia River; 5, Zhuanglang River; 6, Datong River; 7, Huangshui River; 8, Guide; 9, Maqu; 10, Bailong River; 11, Upper reaches of Jialing River.
1.2 调查方法
2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点。主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞。使用GPS定位设备标定采样点的地理信息。捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定。物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 )。
1.3 数据分析
1.3.1 水系单元间物种组成相似性分析
分析所用数据为水系单元间鱼类种类分布的有无, 有分布的赋值为1, 没有分布的赋值为0。相似性采用Jaccard指数(Jaccard, 1901 ), 然后根据相似性系数使用UPGMA法(unweighted pair-group method with arithmetic means)进行聚类分析(Fu et al, 2003 )。
1.3.2 分布热点区分析
运用地理信息系统软件ArcGIS 9.2将调查区域栅格化, 形成25 km × 25 km的地理单元, 对高原鳅属鱼类的分布格局进行热点区域预测。基于调查结果, 并结合历史资料估计高原鳅属每个物种现有的分布区域; 根据物种分布的有无, 确定地理单元内物种的丰富度。
1.3.3 物种多样性指数
使用以下指数进行鱼类群落的物种多样性分析:
物种丰富度(S ) = 群落中的物种数目(王太等, 2015 )
Shannon-Wiener多样性指数(H' ) (Shannon & Weaver, 1949 ):H' = -ΣPi lnPi (1)
Pielou均匀度指数(J' ) (Pielou, 1975 ):J' = H' /lnS (2)
Simpson集中性概率指数(戈峰, 2002 ):λ = ΣPi 2 (3)Pi 代表第i 种个体占总个体数的比例。
1.3.4 物种海拔分布格局分析
根据调查结果并结合大量的早期资料, 尽可能准确地确定每个高原鳅物种的种域(物种的海拔分布范围)。
在分析物种丰富度与海拔高度之间的关系时, 我们以200 m为单位划分海拔带(例如, 600-799 m, 800-999 m)。统计每个海拔带内的高原鳅属物种总数, 并以上文介绍的聚类方法分析各海拔带间的物种组成关系。
在物种多样性的海拔格局分析中, 将每个样点定义为1个群体单元, 然后进行物种多样性分析。对多样性分析结果进行曲线估计(curve estimation), 以探究多样性指数与海拔的关系, 拟合曲线的选择依据刘哲等(2015) 的方法。内容包括Shannon-Wiener指数、Simpson指数和Pielou指数与海拔梯度的关系。全部分析在SPSS 19.0中完成。
2 结果
2.1 高原鳅的种类与分布
2012-2015年间的调查中, 共发现高原鳅属鱼类19种, 其中未定种2种。整体上高原鳅的物种数量有所减少(附录1)。河西内陆水系共发现高原鳅7种; 其中以黑河流域最多, 有6种; 而石羊河与疏勒河各发现4种。河西水系中那些与主流水体隔离的附属河流明显包含较少的高原鳅物种; 除黑河水系的大马营河有4种外, 其余最多仅有2种高原鳅属鱼类(表1 )。青海湖水系发现高原鳅4种, 其中甘子河2种。黄河水系共发现高原鳅12种, 其中洮河的物种数最多, 有8种; 其次为湟水河7种; 大通河与黄河玛曲段的支流均发现6种。渭河水系包含4种高原鳅。嘉陵江水系5种。19种高原鳅中仅斯氏高原鳅(Triplophysa stoliczkae )在各大水系都有分布, 其余物种只在少数水系有分布。各水系间均存在共有种, 物种较多的区域几乎都有邻近水系的物种。
2.2 水系单元间的物种组成相似性
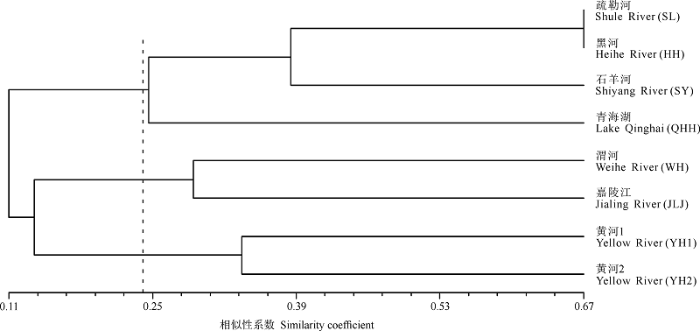
水系单元的物种组成相似性聚类分析结果显示(图2 ), 河西三个水系有最相似的物种组成, 它们首先和青海湖聚在一起; 黄河贵德以上段和以下兰州段聚在一起; 嘉陵江与渭河聚在一起。当相似性系数介于0.29和0.33之间时, 聚类结果分为5组, 与主要水系划分相一致(河西, 青海湖, 渭河, 嘉陵江, 黄河)。此外, 结果也显示, 地理或水系距离越近的水体单元的物种组成越相似。
图2 研究区域各水系单元基于高原鳅属物种组成的UPGMA聚类分析。YH1是黄河上游水系贵德以下段, YH2是黄河上游水系贵德以上段。
Fig. 2 UPGMA cluster of basin units in study area based on component of Triplophysa species. YH1 and YH2 are the sections above and below of Guide in upper reaches of Yellow River.
2.3 高原鳅的分布热点估计
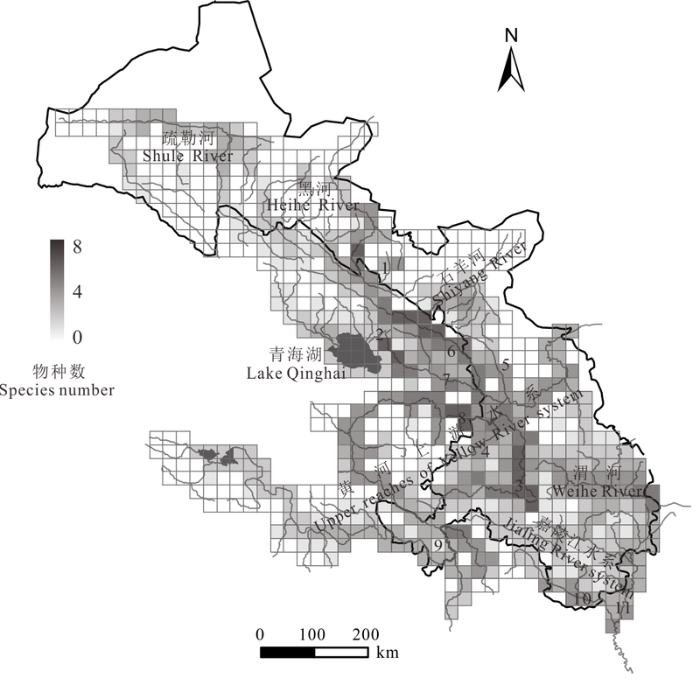
整个调查区域中, 黑河中上游、大通河、洮河的高原鳅属鱼类物种丰富度最高; 河西地区整体的物种丰富度较低(图3 )。而且, 区域内物种分布呈现不均匀现象, 河西地区尤为明显。
图3 青藏高原东北部边缘地区高原鳅属鱼类物种丰富度分布格局, 图中河流编号与图1 中一致。
Fig. 3 The species richness pattern of Triplophysa species in northeastern margin of Tibetan Plateau, serial number in accordance with Fig. 1 .
2.4 物种多样性指数
对各水系的Shannon-Wiener指数、Pielou指数和Simpson指数分析结果(表1 )显示: 大通河的Shannon-Wiener指数最高(1.390 ± 0.014), 其次为洮河(1.198 ± 0.246)、黑河(1.136 ± 0.213); 最低的是黑河流域水体(0.320 ± 0.157)。青海湖流域河流Pielou指数最高(0.973 ± 0.046), 庄浪河最低(0.410); 大通河(0.820 ± 0.057)、洮河(0.848 ± 0.122)、黑河(0.842 ± 0.078)较高。黑河流域的附属水体Simpson指数最高(0.806 ± 0.117), 其次是庄浪河(0.723); 最低的是大通河(0.293 ± 0.019), 其次是洮河(0.337 ± 0.072), 黑河(0.350 ± 0.064)较低。
以上结果表明, 大通河、洮河和黑河是青藏高原东北部边缘地区高原鳅属鱼类多样性水平最高的河流; 黑河流域附属水体(大马营河除外)的多样性最低。黄河水系中, 玛曲至玛多段、庄浪河、大夏河为多样性较低的区域。河西地区三大水系的干流与附属隔离水体间存在明显的多样性差异, 其中黑河水系最为显著。
2.5 物种多样性的海拔分布格局
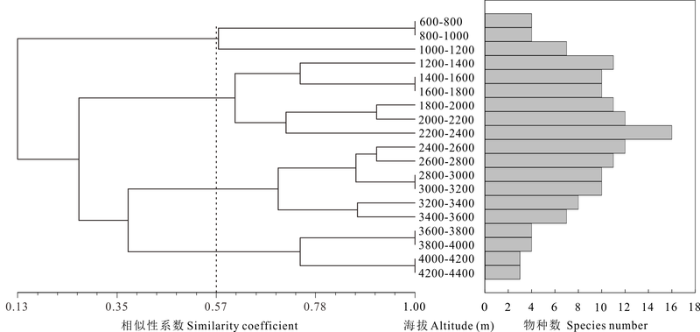
聚类分析显示, 研究区域的海拔根据高原鳅的组成可以分成4段(图4 ): 600-1,200 m、1,200-2,400 m、2,400-3,600 m和3,600-4,400 m。随着海拔的上升, 物种丰富度呈现先增后减的变化模式, 在2,200-2,400 m海拔区间内达到最高。
图4 青藏高原东北部边缘地区高原鳅属鱼类物种丰富度的海拔分布格局
Fig. 4 Elevational distribution pattern of species richness of Triplophysa in northeastern margin of Tibetan Plateau
依据图2 的聚类结果, 相似性系数为0.24时可将全部水系分为3组(河西与青海湖、黄河、嘉陵江与渭河)。据此, 我们也分析了各组内多样性系数与海拔的关系。
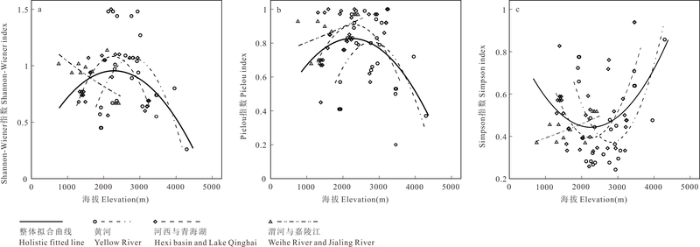
Shannon-Wiener指数与海拔的回归结果(图5 a)显示, 河西与青海湖区域(R 2 = 0.512, P = 0.000)、黄河区域(R 2 = 0.341, P = 0.004)以及整体拟合曲线(R 2 = 0.121, P = 0.009)均呈明显的单峰曲线, 峰值都在2,000-3,000 m的海拔范围内; 其中, 黄河区域的峰值点海拔较河西与青海湖区域的高。渭河与嘉陵江区域(R 2 = 0.477, P = 0.000)的拟合曲线并未呈现单峰型, 而是随着海拔的上升缓缓下降。
图5 生物多样性指数与海拔的关系
Fig. 5 The relationship between biodiversity indices and elevation
图5 b显示的是Pielou指数与海拔的回归分析,结果与Shannon-Wiener指数较相似: 河西与青海湖区域(R 2 = 0.162, P = 0.019)、黄河区域(R 2 = 0.353, P = 0.003)以及整体拟合曲线(R 2 = 0.115, P = 0.008)都为明显的单峰曲线, 峰值都在2,000-3,000 m的海拔范围内; 其中, 黄河区域峰值点的海拔仍较河西与青海湖区域的高。渭河与嘉陵江区域(R 2 = 0.202, P = 0.000)的拟合线随着海拔的上升而上升。
Simpson指数与海拔的回归结果(图5 c)也类似。河西与青海湖区域(R 2 = 0.307, P = 0.001)、黄河区域(R 2 = 0.449, P = 0.000)和整体拟合曲线(R 2 = 0.133, P = 0.004)也均呈明显的单峰曲线, 峰值都在2,000-3,000 m的海拔范围内; 其中黄河区域的峰值点海拔较河西与青海湖区域的高。渭河与嘉陵江区域(R 2 = 0.406, P = 0.000)的拟合曲线随着海拔的上升而上升。
结合以上结果, 该区域整体上在海拔2,000- 3,000 m的范围内物种多样性达到最高。河西与青海湖区域物种多样性最大时的海拔较黄河区域的低, 但都落在2,000-3,000 m的海拔范围内。渭河与嘉陵江区域的Shannon-Wiener指数与Simpson指数显示, 随海拔的上升物种多样性水平下降; 而Pielou指数却显示均匀度随着海拔上升而上升。
3 讨论
3.1 青藏高原东北部边缘区高原鳅属鱼类的分布与多样性现状分析
本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化。高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 )。因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素。在调查过程中, 水电站、灌渠等水利设施随处可见。大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆))。庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段。此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定。
分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区。其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透。黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区。此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件。这些现象也反映了这3个区域的特殊性。现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 )。此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著。造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素。大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 )。
3.2 青藏高原东北部边缘地区高原鳅属鱼类多样性的海拔格局
物种多样性随海拔梯度的变化是生物多样性保护研究的重要内容(Vetaas & Grytnes, 2002 ; Parmesan, 2006 ), 为保护区的选择以及保护方法的提出提供参考。聚类分析结果显示(图4 ), 根据高原鳅的物种组成可以将该区域的海拔分成4段: 600-1,200 m、1,200-2,400 m、2,400-3,600 m和3,600-4,400 m。意味着该区域在海拔梯度上存在着至少4个物种群落带, 即每一段内有着较相似的物种组成而与相邻段在物种成分上差异较大。该结果与Fu等(2004)在长江流域淡水鱼类海拔梯度的研究结果基本吻合。高原鳅物种丰富度呈现随海拔上升先增后降的单峰曲线分布, 在中海拔处最高。物种海拔丰富度与聚类结果的结合很好地印证了Lomolino (2001)的预测, 即“物种密度会在中海拔处达到最大, 且出现在两个并列的有着丰富物种的群落的过渡带”。我们推测, 这种随海拔上升先增后降的单峰模式在一定程度上与物种在海拔方向上的扩散活动有关。
多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致。这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 )。同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔。这种现象可能与该区域中黄河流域有着较高的中点海拔有关。其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高。这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式。对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度。
3.3 青藏高原东北部边缘地区鱼类资源保护与管理的建议
整个青藏高原东北部边缘区域的鱼类几乎都是由中亚高山区系复合体的高原鳅和裂腹鱼组成。在调查的过程中, 我们也发现高原鳅物种数和资源量较大的地区往往其他鱼类也有着较高的资源量。因而我们认为, 高原鳅的群落动态在一定程度上可以反映一定区域水体的渔业资源状况。而且, 高原鳅因其物种分布相对集中且资源量较大, 便于实时地进行群体监测。依据我们的调查与分析结果, 对该区域的鱼类资源保护与管理提出几点建议:
(1)中海拔处(2,000-3,000 m)需要优先保护。中海拔处物种丰富度最高, 且有着较高的多样性水平。本研究中高原鳅的分布热点区也几乎都落在这一范围内。同时, 对于海拔跨度较大的水域, 也应考虑其中间海拔, 这里也往往有着较高的物种丰富度。
(2)合理的水利设施建设。大量的水利建设是影响鱼类种群动态的主要原因之一。水坝的建设在破坏水体生境异质性、阻隔鱼类洄游通道的同时也影响了水坝下游水环境的稳定性; 尤其在河西干旱地区, 水坝下游的河道经常干涸, 有些甚至已经消失, 即便有水的河道也已经渠化, 这些都直接影响鱼类的生存。河道的渠化在破坏土著种生境的同时使得水体的连续性遭破坏, 这样的水体环境更容易遭受扰动。因此, 对于水坝的建设我们认为应尽可能选在物种多样性峰值点以下的海拔位置, 这样可以尽可能多地保存河道中的鱼类物种。
(3)增强河道整治力度。甘肃地区多数河道都存在采石挖沙的现象, 有些河段极为严重。采挖活动破坏河道原有生态环境的同时也改变着水体的理化条件, 直接影响水体中鱼类的组成和数量。需要加大监管和治理力度, 减少生活垃圾对水体的污染。
总之, 该区域鱼类资源的保护和管理应以就地保护为主, 重点在于保护物种栖息地。其次, 保障河流水体的连续性, 减少水体环境受外界因素的波动也是该地区鱼类保护的重要内容。
致谢: 感谢中国科学院西北高原生物研究所李颖、刘哲以及中国农业科学院农田灌溉研究所米兆荣博士在数据分析过程中给予的帮助。甘肃省、青海省、四川省阿坝州各级渔政部门在调查过程中也给予了很大的支持与协助, 在此一并表示感谢。
附录 Supplementary Material
Appendix 1 Species list and distribution of Triplophysa in northeastern margin of Tibetan Plateau
http://www.biodiversity-science.net/fileup/PDF/2016259-1.pdf
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Chen YY Chen YF Liu HZ 1996 ) Studies on the position of the Qinghai-Xizang Plateau region in zoogeographic divisions and its eastern demarcation line.
Acta Hydrobiologica Sinica , 20 , 97 -103 . (in Chinese with English abstract)
[本文引用: 4]
[陈宜瑜 , 陈毅峰 , 刘焕章 (1996 ) 青藏高原动物地理区的地位和东部界线问题
. 水生生物学报 , 20 , 97 -103 .]
[本文引用: 4]
[2]
Ding RH 1994 ) The Fishes of Sichuan . Sichuan Publishing House of Science and Technology , Chengdu . (in Chinese)
[本文引用: 1]
[丁瑞华 (1994 ) 四川鱼类志 . 四川科学技术出版社 , 成都 .]
[本文引用: 1]
[3]
Dong ZR Sun DY Zhao JY Zhang J 2010 ) Holistic conceptual model for the structure and function of river ecosystems.
Advances in Water Science , 21 , 550 -559 . (in Chinese with English abstract)
[本文引用: 1]
[董哲仁 , 孙东亚 , 赵进勇 , 张晶 (2010 ) 河流生态系统结构功能整体性概念模型
. 水科学进展 , 21 , 550 -559 .]
[本文引用: 1]
[4]
Fu CZ Wu JH Chen JK Wu QH Lei GC 2003 ) Freshwater fish biodiversity in the Yangtze River basin of China: patterns, threats and conservation.
Biodiversity and Conservation , 12 , 1649 -1685 .
[本文引用: 1]
[5]
Fu CZ Wu JH Wang XY Lei GC Chen JK 2004 ) Patterns of diversity, altitudinal range and body size among freshwater fishes in the Yangtze River basin, China.
Global Ecology and Biogeography , 13 , 543 -552 .
[6]
Ge F 2002 ) Modern Ecology . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[戈峰 (2002 ) 现代生态学 . 科学出版社 , 北京 .]
[本文引用: 1]
[7]
He CL Song ZB Zhang E 2011 ) Triplophysa fishes in China and the status of its taxonomic studies.
Sichuan Journal of Zoology , 30 , 150 -155 . (in Chinese with English abstract)
[本文引用: 2]
[何春林 , 宋昭彬 , 张鹗 (2011 ) 中国高原鳅属鱼类及其分类研究现状
. 四川动物 , 30 , 150 -155 .]
[本文引用: 2]
[8]
He DK Chen YX Chen YF 2006 ) The molecular phylogeny and biogeography of genus Triplophysa.
Progress in Natural Science , 16 , 1395 -1404 . (in Chinese with English abstract)
[本文引用: 1]
[何德奎 , 陈咏霞 , 陈毅峰 (2006 ) 高原鳅属Triplophysa鱼类的分子系统发育和生物地理学研究
. 自然科学进展 , 16 , 1395 -1404 .]
[本文引用: 1]
[9]
He DK Chen YF 2007 ) Molecular phylogeny and biogeography of the highly specialized grade schizothoracine fishes.
Chinese Science Bulletin , 52 , 303 -312 . (in Chinese with English abstract)
[本文引用: 1]
[何德奎 , 陈毅峰 (2007 ) 高度特化等级裂腹鱼类的分子系统发育与生物地理学
. 科学通报 , 52 , 303 -312 .]
[本文引用: 1]
[10]
Jaccard P 1901 ) Distribution de la flore alpine dans le Bassin des Dranes et dans quelques régions voisines.
Bulletin Société Vaudoise des Science Naturelles , 37 , 241 -272 .
[本文引用: 1]
[11]
Liu Z Li Q Chen DD Zhai WT Zhao L Xu SX Zhao XQ 2015 ) Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground biomass in alpine meadows in Qinghai-Tibet Plateau.
Biodiversity Science , 23 , 451 -462 . (in Chinese with English abstract)
[本文引用: 2]
[刘哲 , 李奇 , 陈懂懂 , 翟文婷 , 赵亮 , 徐世晓 , 赵新全 (2015 ) 青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响
. 生物多样性 , 23 , 451 -462 .]
[本文引用: 2]
[12]
Lomolino MV 2001 ) Elevation gradients of species-density: historical and prospective views.
Global Ecology and Biogeography , 10 , 3 -13 .
[13]
Ma J Wu YJ Xia L Zhang Q Ma Y Yang QS 2010 ) Elevational diversity of small mammals in Luoji Mt. Nature Reserve, Sichuan Province.
Acta Theriologica Sinica , 30 , 400 -410 . (in Chinese with English abstract)
[本文引用: 1]
[马俊 , 吴永杰 , 夏霖 , 张乾 , 马勇 , 杨奇森 (2010 ) 螺髻山自然保护区非飞行小型哺乳动物垂直多样性调查
. 兽类学报 , 30 , 400 -410 .]
[本文引用: 1]
[14]
Myers N Mittermeier RA Mittermeier CG da Fonseca GAB Kent J 2000 ) Biodiversity hotspots for conservation priorities.
Nature , 403 , 853 -858 .
[本文引用: 1]
[15]
Parmesan C 2006 ) Ecological and evolutionary responses to recent climates change. Annual Review of Ecology, Evolution,
and Systematics , 37 , 637 -669 .
[本文引用: 1]
[16]
Pielou EC 1975 ) Ecological Diversity. Wiley, New York.
[本文引用: 1]
[17]
Prendergast JR Quinn RM Lawton JH Eversham BC Gibbons DW 1993 ) Rare species, the coincidence of diversity hotspots and conservation strategies.
Nature , 365 , 335 -337 .
[本文引用: 1]
[18]
Shannon EC Weaver W 1949 ) The Mathematical Theory of Communication . Urbana University of Illinois Press , Illinois .
[本文引用: 1]
[19]
Vetaas OR Grytnes JA 2002 ) Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal.
Global Ecology and Biogeography , 11 , 291 -301 .
[本文引用: 1]
[20]
Wang T Zhang YP Guan LH Du YY Lou ZY Jiao WL 2015 ) Current freshwater fish resources and the application of DNA barcoding in species identification in Gansu Province.
Biodiversity Science , 23 , 306 -313 . (in Chinese with English abstract)
[本文引用: 1]
[王太 , 张艳萍 , 管丽红 , 杜岩岩 , 娄忠玉 , 焦文龙 (2015 ) 甘肃省鱼类资源现状及DNA条形码在鱼类物种鉴定中的应用
. 生物多样性 , 23 , 306 -313 .]
[本文引用: 1]
[21]
Wang XT 1991 ) Vertebrate Fauna of Gansu . Gansu Science and Technology Press , Lanzhou . (in Chinese)
[王香亭 (1991 ) 甘肃省脊椎动物志 . 甘肃科学技术出版社 , 兰州.]
[22]
Williams PH Humphries CJ Vane-Wright RI 1991 ) Measuring biodiversity: taxonomic relatedness for conservation priorities.
Australian Systematic Botany , 4 , 665 -679 .
[本文引用: 1]
[23]
Wu YF Wu CZ 1990 ) A preliminary study of the fishery resources and the countermeasures of fishery development in
[24]
the region of the Karakorum-Kunlun Mountains, China
. Journal of Natural Resources , 5 , 354 -364 . (in Chinese with English abstract)
[本文引用: 1]
[武云飞 , 吴翠珍 (1990 ) 喀喇昆仑山-昆仑山地区渔业资源及渔业发展对策的初步研究
. 自然资源学报 , 5 , 354 -364 .]
[本文引用: 1]
[25]
Wu YF Wu CZ 1992 ) The Fishes of the Qinghai-Xizang Plateau . Sichuan Publishing House of Science & Technology , Chengdu . (in Chinese)
[本文引用: 4]
[武云飞 , 吴翠珍 (1992 ) 青藏高原鱼类 . 四川科学技术出版社 , 成都.]
[本文引用: 4]
[26]
Wu YJ Yang QS Xia L Feng ZJ Zhou HM 2012 ) Species diversity and distribution pattern of non-volant small mammals along the elevational gradient on eastern slope of Gongga Mountain.
Acta Ecologica Sinica , 32 , 4318 -4328 . (in Chinese with English abstract)
[本文引用: 1]
[吴永杰 , 杨奇森 , 夏霖 , 冯祚建 , 周华明 (2012 ) 贡嘎山东坡非飞行小型兽类物种多样性的垂直分布格局
. 生态学报 , 32 , 4318 -4328 .]
[本文引用: 1]
[27]
Yang YT Zhang YM 1991 ) Study on fishes’ fauna and evolution of the Inland River, Hexi Corridor.
Journal of Lanzhou University (Natural Sciences) , 27 , 141 -144 . (in Chinese with English abstract)
[本文引用: 1]
[杨友桃 , 张迎梅 (1991 ) 河西走廊鱼类区系及其演变的研究
. 兰州大学学报(自然科学版) , 27 , 141 -144 .]
[本文引用: 1]
[28]
Zhang YY Zhou LZ Wang QS Wang XJ Xing YJ 2008 ) Distribution pattern and hotspot analysis of breeding birds in Anhui Province.
Biodiversity Science , 16 , 305 -312 . (in Chinese with English abstract)
[本文引用: 1]
[张有瑜 , 周立志 , 王岐山 , 王新建 , 邢雅俊 (2008 ) 安徽省繁殖鸟类分布格局和热点区分析
. 生物多样性 , 16 , 305 -312 .]
[本文引用: 1]
[29]
Zhao K 2009 ) Adaptive Evolution and Biogeography in the Schizothoracine Fish (Cyprinidae) Endemic to the Northeast Tibetan Plateau
. Postdoctoral research report, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining . (in Chinese with English abstract)
[本文引用: 1]
[赵凯 (2009 ) 东北部青藏高原特有裂腹鱼类的适应性进化和生物地理学研究
. 博士后研究工作报告, 中国科学院西北高原生物研究所, 西宁 .]
[本文引用: 1]
[30]
Zheng Z Gong DJ Sun CX Li XJ Li WJ 2014 ) Elevational pattern of amphibian and reptile diversity in Qinling Range and explanation.
Biodiversity Science , 22 , 596 -607 . (in Chinese with English abstract)
[本文引用: 1]
[郑智 , 龚大洁 , 孙呈祥 , 李晓军 , 李万江 (2014 ) 秦岭两栖、爬行动物物种多样性海拔分布格局及其解释
. 生物多样性 , 22 , 596 -607 .]
[本文引用: 1]
[31]
Zhu SQ 1989 ) The Loaches of the Subfamily Nemacheilinae in China . Jiangsu Science and Technology Publishing House, Nanjing . (in Chinese)
[本文引用: 3]
[朱松泉 (1989 ) 中国条鳅志 . 江苏科学技术出版社 , 南京.]
[本文引用: 3]
青藏高原动物地理区的地位和东部界线问题
4
1996
... 青藏高原东北部边缘地区在地理上包括甘肃省大部分区域、青海省东部和北部以及四川省北部, 面积约469,000 km2 , 地貌复杂, 气候各异, 是青藏高原、黄土高原、蒙新高原的交汇地带.水系为青藏高原水系的一部分.由南向北主要分为: 长江流域、黄河流域、青海湖流域、河西内陆河流域.我国三大动物地理区(古北区、东洋区、青藏高原区)也在此交汇(陈宜瑜等, 1996 ).目前认为, 青藏高原东北部边缘可能是一个重要的冰河期避难所和目前一些广泛分布在高原的特有物种遗传多样性较高的地区(赵凯, 2009 ). ...
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
青藏高原动物地理区的地位和东部界线问题
4
1996
... 青藏高原东北部边缘地区在地理上包括甘肃省大部分区域、青海省东部和北部以及四川省北部, 面积约469,000 km2 , 地貌复杂, 气候各异, 是青藏高原、黄土高原、蒙新高原的交汇地带.水系为青藏高原水系的一部分.由南向北主要分为: 长江流域、黄河流域、青海湖流域、河西内陆河流域.我国三大动物地理区(古北区、东洋区、青藏高原区)也在此交汇(陈宜瑜等, 1996 ).目前认为, 青藏高原东北部边缘可能是一个重要的冰河期避难所和目前一些广泛分布在高原的特有物种遗传多样性较高的地区(赵凯, 2009 ). ...
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
1
1994
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
1
1994
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
河流生态系统结构功能整体性概念模型
1
2010
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
河流生态系统结构功能整体性概念模型
1
2010
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
Freshwater fish biodiversity in the Yangtze River basin of China: patterns, threats and conservation.
1
2003
... 分析所用数据为水系单元间鱼类种类分布的有无, 有分布的赋值为1, 没有分布的赋值为0.相似性采用Jaccard指数(Jaccard, 1901 ), 然后根据相似性系数使用UPGMA法(unweighted pair-group method with arithmetic means)进行聚类分析(Fu et al, 2003 ). ...
Patterns of diversity, altitudinal range and body size among freshwater fishes in the Yangtze River basin, China.
2004
1
2002
... Simpson集中性概率指数(戈峰, 2002 ):λ = ΣPi 2 (3)Pi 代表第i 种个体占总个体数的比例. ...
1
2002
... Simpson集中性概率指数(戈峰, 2002 ):λ = ΣPi 2 (3)Pi 代表第i 种个体占总个体数的比例. ...
中国高原鳅属鱼类及其分类研究现状
2
2011
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
中国高原鳅属鱼类及其分类研究现状
2
2011
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
高原鳅属Triplophysa鱼类的分子系统发育和生物地理学研究
1
2006
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
高原鳅属Triplophysa鱼类的分子系统发育和生物地理学研究
1
2006
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
高度特化等级裂腹鱼类的分子系统发育与生物地理学
1
2007
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
高度特化等级裂腹鱼类的分子系统发育与生物地理学
1
2007
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
Distribution de la flore alpine dans le Bassin des Dranes et dans quelques régions voisines.
1
1901
... 分析所用数据为水系单元间鱼类种类分布的有无, 有分布的赋值为1, 没有分布的赋值为0.相似性采用Jaccard指数(Jaccard, 1901 ), 然后根据相似性系数使用UPGMA法(unweighted pair-group method with arithmetic means)进行聚类分析(Fu et al, 2003 ). ...
青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响
2
2015
... 在物种多样性的海拔格局分析中, 将每个样点定义为1个群体单元, 然后进行物种多样性分析.对多样性分析结果进行曲线估计(curve estimation), 以探究多样性指数与海拔的关系, 拟合曲线的选择依据刘哲等(2015) 的方法.内容包括Shannon-Wiener指数、Simpson指数和Pielou指数与海拔梯度的关系.全部分析在SPSS 19.0中完成. ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响
2
2015
... 在物种多样性的海拔格局分析中, 将每个样点定义为1个群体单元, 然后进行物种多样性分析.对多样性分析结果进行曲线估计(curve estimation), 以探究多样性指数与海拔的关系, 拟合曲线的选择依据刘哲等(2015) 的方法.内容包括Shannon-Wiener指数、Simpson指数和Pielou指数与海拔梯度的关系.全部分析在SPSS 19.0中完成. ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
Elevation gradients of species-density: historical and prospective views.
2001
螺髻山自然保护区非飞行小型哺乳动物垂直多样性调查
1
2010
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
螺髻山自然保护区非飞行小型哺乳动物垂直多样性调查
1
2010
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
Biodiversity hotspots for conservation priorities.
1
2000
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
Ecological and evolutionary responses to recent climates change. Annual Review of Ecology, Evolution,
1
2006
... 物种多样性随海拔梯度的变化是生物多样性保护研究的重要内容(Vetaas & Grytnes, 2002 ; Parmesan, 2006 ), 为保护区的选择以及保护方法的提出提供参考.聚类分析结果显示(图4 ), 根据高原鳅的物种组成可以将该区域的海拔分成4段: 600-1,200 m、1,200-2,400 m、2,400-3,600 m和3,600-4,400 m.意味着该区域在海拔梯度上存在着至少4个物种群落带, 即每一段内有着较相似的物种组成而与相邻段在物种成分上差异较大.该结果与Fu等(2004)在长江流域淡水鱼类海拔梯度的研究结果基本吻合.高原鳅物种丰富度呈现随海拔上升先增后降的单峰曲线分布, 在中海拔处最高.物种海拔丰富度与聚类结果的结合很好地印证了Lomolino (2001)的预测, 即“物种密度会在中海拔处达到最大, 且出现在两个并列的有着丰富物种的群落的过渡带”.我们推测, 这种随海拔上升先增后降的单峰模式在一定程度上与物种在海拔方向上的扩散活动有关. ...
Ecological Diversity. Wiley, New York.
1
1975
... Pielou均匀度指数(J' ) (Pielou, 1975 ):J' = H' /lnS (2) ...
Rare species, the coincidence of diversity hotspots and conservation strategies.
1
1993
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
1
1949
... Shannon-Wiener多样性指数(H' ) (Shannon & Weaver, 1949 ):H' = -ΣPi lnPi (1) ...
Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal.
1
2002
... 物种多样性随海拔梯度的变化是生物多样性保护研究的重要内容(Vetaas & Grytnes, 2002 ; Parmesan, 2006 ), 为保护区的选择以及保护方法的提出提供参考.聚类分析结果显示(图4 ), 根据高原鳅的物种组成可以将该区域的海拔分成4段: 600-1,200 m、1,200-2,400 m、2,400-3,600 m和3,600-4,400 m.意味着该区域在海拔梯度上存在着至少4个物种群落带, 即每一段内有着较相似的物种组成而与相邻段在物种成分上差异较大.该结果与Fu等(2004)在长江流域淡水鱼类海拔梯度的研究结果基本吻合.高原鳅物种丰富度呈现随海拔上升先增后降的单峰曲线分布, 在中海拔处最高.物种海拔丰富度与聚类结果的结合很好地印证了Lomolino (2001)的预测, 即“物种密度会在中海拔处达到最大, 且出现在两个并列的有着丰富物种的群落的过渡带”.我们推测, 这种随海拔上升先增后降的单峰模式在一定程度上与物种在海拔方向上的扩散活动有关. ...
甘肃省鱼类资源现状及DNA条形码在鱼类物种鉴定中的应用
1
2015
... 物种丰富度(S ) = 群落中的物种数目(王太等, 2015 ) ...
甘肃省鱼类资源现状及DNA条形码在鱼类物种鉴定中的应用
1
2015
... 物种丰富度(S ) = 群落中的物种数目(王太等, 2015 ) ...
Measuring biodiversity: taxonomic relatedness for conservation priorities.
1
1991
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
A preliminary study of the fishery resources and the countermeasures of fishery development in
1990
喀喇昆仑山-昆仑山地区渔业资源及渔业发展对策的初步研究
1
1990
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
喀喇昆仑山-昆仑山地区渔业资源及渔业发展对策的初步研究
1
1990
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
4
1992
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
4
1992
... 高原鳅属(Triplophysa )隶属于鲤形目条鳅科, 是一类主营底层生活的中小型鱼类; 主要分布于青藏高原及其周边地区, 为该地区鱼类区系的重要组成成分(武云飞和吴翠珍, 1992 ).与裂腹鱼类相似, 高原鳅属鱼类是随着青藏高原的隆升而出现, 由条鳅亚科的原始类群演化而来(陈宜瑜等, 1996 ).然而, 高原鳅作为青藏高原鱼类区系最重要的类群之一, 有关其研究依然主要集中在分类学上(何春林等, 2011 ), 其他方面很少开展研究. ...
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
贡嘎山东坡非飞行小型兽类物种多样性的垂直分布格局
1
2012
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
贡嘎山东坡非飞行小型兽类物种多样性的垂直分布格局
1
2012
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
河西走廊鱼类区系及其演变的研究
1
1991
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
河西走廊鱼类区系及其演变的研究
1
1991
... 本次调查中没有发现部分历史记录的高原鳅属物种, 有少许的物种其分布区域也呈现缩小的趋势, 可能意味着该区域高原鳅属鱼类部分物种的种群已经萎缩或者破碎化.高原鳅是主营底层生活的一类中小型鱼类, 对环境有着广泛的适应性(陈宜瑜等, 1996 ).因此, 相比于环境的波动, 人类活动的过度干扰可能是导致高原鳅种群衰退的最主要因素.在调查过程中, 水电站、灌渠等水利设施随处可见.大规模的水利设施使得水体生境变得单一和不连续, 使部分对生境依赖性强的物种不得不退缩至有限的区域或者消失(杨友桃和张迎梅, 1991 ;青弘(青弘 (2010) 嘉陵江流域鱼类多样性格局及中下游渔业资源变迁分析. 硕士学位论文, 西南大学, 重庆)).庄浪河和大夏河河段水体污染严重, 生活垃圾在河道内较为常见, 这些人为的污染在改变水体理化环境的同时也使得河水由原本适合高原鱼类生存的贫营养型转化为适合鲤鲫鱼类生存的富营养型, 改变着鱼类区系成分(武云飞和吴翠珍, 1990 ), 进而使得这些河段的高原鳅鱼类多样性明显低于同地区其他河段.此外, 河道的挖沙采石、大量外来鱼种的引入等都影响着该地区高原鳅属鱼类种群的稳定. ...
安徽省繁殖鸟类分布格局和热点区分析
1
2008
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
安徽省繁殖鸟类分布格局和热点区分析
1
2008
... 分布热点预测和生物多样性分析显示: 黑河中上游、大通河、洮河是该区域物种丰富度和多样性较大的3个区域, 是生物多样性热点地区.其中, 洮河被认为是青藏区与古北区的过渡(陈宜瑜等, 1996 ), 这里不仅有大量高原鳅属鱼类和裂腹鱼类, 也有古北区的物种, 如北方花鳅(Cobitis granoei )等; 同时洮河与白龙江(青藏区与东洋区的过渡)也可能存在着相互渗透.黑河与大通河的鱼类主要为中亚高山区系复合体, 这里的物种密度和资源量均明显高于其他地区.此外, 武云飞和吴翠珍(1992)曾记录黑河地区有武威高原鳅(Triplophysa wuweiensis )的分布, 而我们调查的结果发现这里分布的其实是黄河广泛分布的拟硬刺高原鳅(T. pseudoscleroptera ); 这也可能意味着黑河与黄河(大通河或湟水河)在历史上曾发生过袭夺或渗透事件.这些现象也反映了这3个区域的特殊性.现有研究认为, 生物多样性热点地区往往是物种丰富度最高, 稀有或分类地位特殊的物种集中分布的地区, 需要优先保护(Williams et al, 1991 ; Prendergast et al, 1993 ; Myers et al, 2000 ; 张有瑜等, 2008 ).此外, 分布热点和多样性分析的结果也显示, 青藏高原东北部边缘地区高原鳅属鱼类的分布很不均匀, 河西地区尤为显著.造成此现象的原因除了地史和人为干扰的因素外, 气候和水体条件也是重要因素.大而连续的水体相比于较小的水体对干旱等环境因素有着更强的抗扰动能力, 同时较大的水体又可以提供更多的生态位和生境类型, 能容纳更多的物种(董哲仁等, 2010 ). ...
东北部青藏高原特有裂腹鱼类的适应性进化和生物地理学研究
1
2009
... 青藏高原东北部边缘地区在地理上包括甘肃省大部分区域、青海省东部和北部以及四川省北部, 面积约469,000 km2 , 地貌复杂, 气候各异, 是青藏高原、黄土高原、蒙新高原的交汇地带.水系为青藏高原水系的一部分.由南向北主要分为: 长江流域、黄河流域、青海湖流域、河西内陆河流域.我国三大动物地理区(古北区、东洋区、青藏高原区)也在此交汇(陈宜瑜等, 1996 ).目前认为, 青藏高原东北部边缘可能是一个重要的冰河期避难所和目前一些广泛分布在高原的特有物种遗传多样性较高的地区(赵凯, 2009 ). ...
东北部青藏高原特有裂腹鱼类的适应性进化和生物地理学研究
1
2009
... 青藏高原东北部边缘地区在地理上包括甘肃省大部分区域、青海省东部和北部以及四川省北部, 面积约469,000 km2 , 地貌复杂, 气候各异, 是青藏高原、黄土高原、蒙新高原的交汇地带.水系为青藏高原水系的一部分.由南向北主要分为: 长江流域、黄河流域、青海湖流域、河西内陆河流域.我国三大动物地理区(古北区、东洋区、青藏高原区)也在此交汇(陈宜瑜等, 1996 ).目前认为, 青藏高原东北部边缘可能是一个重要的冰河期避难所和目前一些广泛分布在高原的特有物种遗传多样性较高的地区(赵凯, 2009 ). ...
秦岭两栖、爬行动物物种多样性海拔分布格局及其解释
1
2014
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
秦岭两栖、爬行动物物种多样性海拔分布格局及其解释
1
2014
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
3
1989
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
3
1989
... 青藏高原的东北部边缘是高原鳅属鱼类分布的一个热点地区(何春林等, 2011 ), 高原鳅属的原始物种也分布于此(何德奎等, 2006 ).尽管该区域在流域上分属不同的水系, 但水系间鱼类组成的相似度显示这些水系的演化在历史上有着极为密切的关系(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ; 何德奎和陈毅峰, 2007 ), 因此可以作为一个整体单元对待.朱松泉(1989)、王香亭(1991)、武云飞和吴翠珍(1992)、丁瑞华(1994)等较详细地整理了该区域高原鳅种类与分布情况; 此后也有不少相关的报道, 但都是一些局部水域的工作.同时, 由于方法的局限性, 早期工作不能全面地反映物种或种群在空间分布上的变异情况, 因而无法最大限度地指导物种多样性的保护工作.鉴于此, 本研究基于2012-2015年间对该区域的调查, 结合现有的资料, 对青藏高原东北部边缘地区的高原鳅属鱼类的多样性与分布格局进行了全面的研究, 旨在为该地区的鱼类保护、生态学和生物地理学研究提供基础资料. ...
... 2012-2015年, 在全区各水系的干支流设置采样点, 较长的河流分上、中、下段进行采样, 每段至少设置2个采样点.主要使用地笼采集标本, 并结合小网目的刺网和捕鱼器进行实地捕捞.使用GPS定位设备标定采样点的地理信息.捕获样品使用95%的酒精固定保存; 对较大的个体, 现场进行鉴定, 测量体长、体重并拍照后, 剪去部分鳍条固定.物种的鉴定及分类依据《中国条鳅志》(朱松泉, 1989 )、《青藏高原鱼类》(武云飞和吴翠珍, 1992 )和《四川鱼类志》(丁瑞华, 1994 ). ...
... 多样性指数的海拔回归分析获得了几乎一致的结果, 反映出多样性水平在中海拔处达到最高, 与物种丰富度的海拔分布相一致.这进一步说明这种中海拔处最大的单峰分布格局可能是青藏高原及其周边山地物种多样性分布的普遍规律(马俊等, 2010 ; 吴永杰等, 2012 ; 郑智等, 2014 ; 刘哲等, 2015 ).同样, 在该区域内海拔跨度较大的水域我们也发现了这种中海拔处最大的多样性分布模式, 并且黄河区域的峰值点海拔明显高于河西与青海湖区域的峰值点海拔.这种现象可能与该区域中黄河流域有着较高的中点海拔有关.其次, 渭河与嘉陵江流域的Shannon-Wiener指数和Simpson指数均显示随海拔降低多样性水平升高.这两条河我们仅对其上游区段进行了详细调查, 而根据文献记录(朱松泉, 1989 ; 武云飞和吴翠珍, 1992 ), 其下游区段的高原鳅减少; 因而我们推断其完整区域的多样性也应该是中间高的分布模式.对于Pielou指数随海拔降低而下降, 我们认为可能是由于随海拔下降, 其他非高原冷水鱼类的增多降低了高原鳅的均匀度. ...
 , 赵凯
, 赵凯

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}