物种是生物学研究的基石, 任何生物学研究都离不开物种, 即以一个或多个物种为研究对象开展各种不同的研究(Darwin, 1859 ; Mallet, 1995 ; Coyne & Orr, 2004 )。但是, 物种又是生物学中争论最大的定义或概念(Mallet, 1995 )。如何定义物种?物种概念如何指导物种界定和分类修订以及其他研究?下面是我对这些问题的思考。
1 “整合物种概念”
到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义。这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 )。其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来。但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状。在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要。一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准。
生物学物种是所有教科书介绍“物种”时都要介绍的经典概念。生殖隔离是生物学物种概念的核心(Mayr, 1963 ); 但在定义和阐述生物学物种概念时, 人们往往忽略了在群体水平上两个物种的生殖隔离存在一定的“度”, 从100% (所有的个体都存在生殖隔离)到0% (所有的个体之间都不存在生殖隔离)。合子后的生殖隔离是生物学物种中最常见的支持证据; 但是两个近缘物种杂交后代雄性不育、雌性可育(Haldane’s rule)在很多动物类群中都十分常见(Coyne & Orr, 2004 ), 从而说明这些所谓的“生物学物种”也不是100%的生殖隔离, 因为雌性可育也可能导致种间界限打破, 使得基因在种间转移(Mallet, 1995 )。植物以及部分动物属中还广泛存在多倍化现象, 多倍化形成的物种一般认为是比较理想的生物学物种; 但是, 不同倍性的物种之间仍可能存在少量的杂交后代和基因流(Grant, 1981 )。因此, “生物学物种”概念应修订为“种间生殖隔离程度远大于种内居群间生殖隔离程度”才更加符合实际情况。
此外, 生物学物种概念在异域分布的地理隔离的近缘物种中应用起来十分不方便。地理隔离物种之间可能存在由于遗传漂变等机制导致多位点分化产生的合子后生殖隔离以及其他机制产生的交配前生殖隔离(Coyne & Orr, 2004 ); 但是, 这些隔离机制也可能在一些最近才异域隔离分布的姊妹形态学种中都不存在。
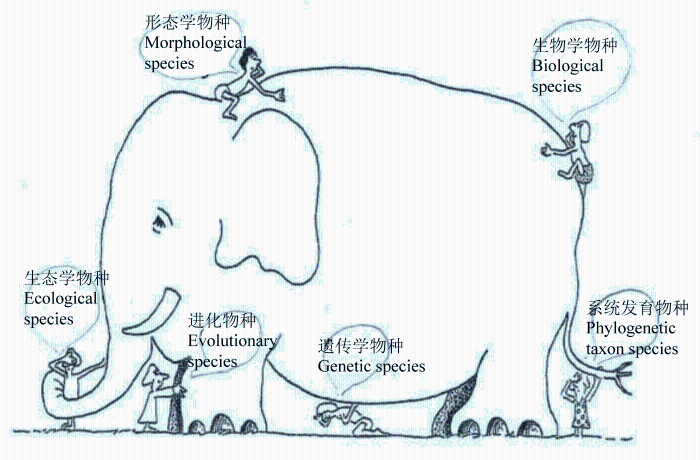
不同的物种概念都是用来对自然界存在的某类生物进行物种划分, 只是各自强调的重点不一样。如果换一个角度来比较, 这些概念强调的是分化完全、“成熟”物种的某个方面。对于来自同一个最近共同祖先的两个分化较好的以有性生殖为主的“好种”, 既有形态上的统计间断, 也有程度比较高的生殖隔离, 还有明显的生态位分化, 各为单系, 具有显著不一样的基因型(遗传聚合群, genetic clusters)、各属于遗传分化比较大的进化单元……等等。每个物种概念的定义, 十分类似于盲人摸象(图1 ), 摸到的分别只是大象的某个方面; 当强调这一方面时, 则或多或少忽略了其他方面。因此, 科学而完整的物种概念是“睁开眼睛看大象”, 是涵盖以前定义所有物种概念条件的“整合物种概念”。但是, 自然界中的“物种”能涵盖所有这些方面吗?非也。正是这种不能覆盖各方面的大量“物种” (没有发展到成熟物种阶段, 见下面论述)的存在, 影响了大家对物种的认识, 从而提出了若干“物种概念”定义, 引起了若干争论。此外, 尽管多数以有性生殖为主的动植物类群符合上述情况; 但也有很多生物类群本身也存在例外, 如一些微生物类群不具有肉眼可见的形态性状, 一些植物类群以无融合生殖为主等; 这些类群就如残疾大象, 缺了鼻子或眼睛等; 即使是分化后期的成熟物种, 也满足不了所有的物种概念。
2 “分化路上的物种”
现在看到的物种实际是其“进化”长河中的一个切面, 物种始终是在“分化路上”, 或处于分化的早期, 或处于中期, 或处于后期。产生的物种或者存留至今, 或者永久灭绝, 但更多情况下则是灭绝后又开始新一轮的物种分化, 周而复始。当处于分化的早中期时, 物种显示出来的特征可能只满足其中一个或两个物种概念条件, 而不能满足多个条件; 当处于分化后期时, 则能满足更多甚至所有物种概念的条件。但是, 自然界中处于分化后期的物种相对较少。古老孑遗单种属中的物种如银杏(Ginkgo biloba )、大熊猫(Ailuropoda melanoleuca )等, 以及少种属中分化时间相对较长的一些物种应属于这种情况, 这些物种就是分类上所谓的“好种”。但是, 有些古老孑遗属的少种属尽管属起源古老, 但所包括的物种却可能是新近历史上才开始分化的年轻物种, 如领春木(Cercidiphyllum magnificum )和日本领春木(C. japonicum ) (Mao & Liu, 2012 ; Qi et al, 2012 ), 它们之间的分化不一定符合所有物种概念的条件。这也说明一些所谓的“孑遗物种”实际上还是处在分化路上的“年轻”物种。
图1 以前定义的每个物种概念就像盲人摸象一样, 只看到了一个成熟物种的某一方面
Fig. 1 One of the diverse species concepts defined before may only perceive one aspect of the mature species like “the blind men feel the elephant”
尽管很难科学检验和统计, 但自然界中可能更多的物种应处于物种分化的早中期, 不符合所有物种概念定义的条件。极端的例子是多倍化新产生的四倍体群体, 在形态特征上与祖先二倍体没有统计上的显著差异; 但多倍化导致其生殖隔离程度较高(尽管不是100%隔离, 因为可能还存在通过三倍体桥的基因流动, Grant, 1981 ), 符合修订后的生物学物种概念, 是生物学物种, 但不符合其他物种概念条件。另一类例子是形态隐存种(morphologically cryptic species), 在最近的研究中出现十分频繁(如Yassin et al, 2008 ), 甚至包括知名物种长颈鹿(Giraffa camelopardalis , Fennessy et al, 2016如Fennessy et al, 2016 ); 但是形态性状差异统计不显著, 不符合形态学物种概念。
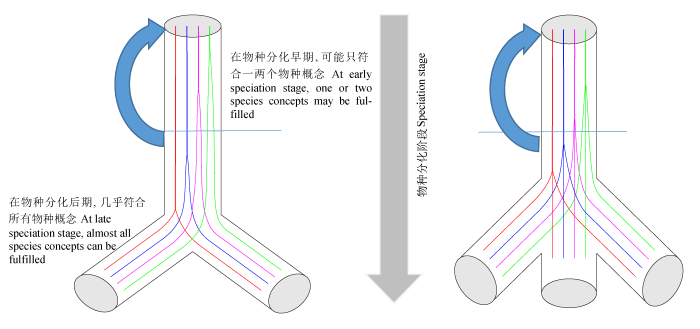
现代遗传学已经证实群体或物种间的分化都有其遗传学基础, 即本质上是DNA或基因的分化(Feder et al, 2013 )。每个物种概念成立所需的条件实际上是相应基因的分化。“一因多效” (一个基因多个效果)确实存在, 如控制形态性状分化的基因同时也导致生殖隔离以及生态位分化(Coyne & Orr, 2004 )。这种情况下, 一个基因的分化可能会满足形态学、生物学、遗传学和生态学4个甚至更多物种概念的条件。但是, 这样的基因和例子相对少见。多数情况下, 每个物种概念所需条件涉及的基因可能是不同的, 如系统发育分析常用的基因或DNA片段是近中性进化的, 而生态物种概念所依托的基因可能具有选择适应性; 生殖隔离和形态差异基因既可能是随机中性进化的产物, 也可能是自然选择导致的差异性表现。各个物种概念所需条件与两个物种群体水平上多个基因的分化式样一致: 有些基因快(选择或其他因素), 有些基因慢。在物种分化后期, 所有的基因都分离, 相当于所有物种概念的条件都得到满足, 成为“好种”。因此, 各物种概念所需条件在“分化路上”的分离与物种分化中的多基因分化式样是基本类似和相同的(图2 )。
因为“整合物种概念”只有在物种分化后期才能完整体现, 所以在分化早中期尝试定义一个针对所有生物的物种概念似乎不可能做到: 因为对于不同的物种对来说, 它们的基因组结构、进化历史以及所受到的选择压力等都是不同的。在物种分化过程中, 有些可能是生态位分化早, 有些可能是形态分化较早, 而另一些则可能是生殖隔离分化较早, 情况极为复杂; 各物种分化条件在不同物种对中应是随机的。还须认识到, 自然界的物种分化不是进行到后期, 即两个或多个后代物种之间(如图2 )已经完整分化(符合所有物种概念)时才开始新一轮的物种分化; 往往是还在分化中(如早中期)就已开始新一轮的物种分化(图2 箭头所示), 这可能导致自然界的很多物种始终处于物种分化的早中期。理论上, 分化时间越长, 遗传差异越大、生殖隔离程度越高; 处于分化早中期的物种, 种间不可避免地产生一定的基因流。种间杂交以及基因流一方面会引起一些基因或DNA片段在种间遗传界限上弱化以及种间关系网状化; 另一方面也可能促进被渗入物种的环境适应性提高和分布区扩展, 甚至这些杂交后代还会形成一个新的同倍性杂交或异源多倍体物种(Coyne & Orr, 2004 ; Abbott et al, 2010 )。
图2 在1个祖先种分化为2至3个(辐射多样化)物种过程中, 满足不同物种概念(不同颜色)条件分化的非一致性。未达到物种分化的最后期, 如箭头所示又开始一轮新的物种分化。由于不完全的生殖隔离, 这些物种之间的基因流与遗传交换应十分频繁。
Fig. 2 Different species concepts (in different colors) may be gradually fulfilled along the divergences of one ancestral species into two or three (radiative diversification) species. Before the late speciation stage, the incomplete diverged daughter species undergo further cycles of speciation (arrows). Gene flow and genetic changes may occur frequently between these assumed “species” due to the incomplete reproductive isolations.
3 物种界定
目前依据形态特征建立的很多物种采用的还是模式物种概念, 而没有采用统计学方法来验证它们是否是真实的形态聚合群。今后采用“整合物种概念”进行物种界定、分类处理和发表新种时, 要尽可能考虑物种分化的各个方面, 也就是目前各物种概念所强调的各项条件, 即“整合”的思想和“整合”的方法。具体来说:
第一步是要对记载的物种或拟发表的新种及其近缘种开展形态性状的统计聚类分析。分析方法需要采用统计学上的聚类分析(如差异显著性比较分析、主成分分析等), 而不是简单的间断性与连续性分析, 因为分化上统计显著的两个聚合群(即物种)之间也可有部分个体处于中间状态或偏离聚合群。这是自然界中多数物种存在的常态, 种间杂交导致的部分基因流以及居群内部分个体的部分基因产生的新生突变等都可能导致此类情况发生。对化石物种也建议开展类似的工作。由于每个化石物种的化石标本不多, 多数古生物学家感觉难以全面采用形态性状差异统计显著的“形态学物种概念”, 但是对那些采集标本较多的类群应该进行此类物种界定的研究工作。古生物学家在物种界定和分类时还存在另一个比较重要的问题, 即把同一谱系不同时间段的化石命名为不同的物种。这些化石物种在系统发育的分子钟标记中很容易被误用, 被错误地置于一个属的系统发育树的冠(crown)支或干(stem)支, 从而缩短或增加拟计算节点的分化时间。
其次, 应采用常用的分子标记, 如DNA条形码序列变异或其他简单的分子标记对所研究物种对或物种群的多个居群进行遗传学分析。根据分析结果来确认拟划分的物种是否可划分为不同的遗传聚合群, 是否是单系, 是否符合遗传学以及系统发育物种概念。这种分析对于没有可直接观察的形态性状以及无融合生殖等类群尤其重要。但是, 这种遗传学研究仍然要注意“群体”和“统计”概念, 只要能将多数个体划分为不同的遗传聚合群, 就说明是不同的进化单元, 应承认种间基因流导致的“中间” (如自然界存在的大量种间杂交个体)或“异常”个体的存在。
第三, 通过生态位模拟以及实地观察来验证是否符合生态学物种概念, 检查物种之间是否具有明显的生态位分化(如花期、行为、资源利用以及时空分布等)。生态模型模拟表明, 两个独立进化的物种不可能具有完全重叠的生态位(Coyne & Orr, 2004 )。
第四, 通过同质园种植(养殖)、同域分布杂交可行性的各种实验以及染色体分析等研究来确认是否符合生物学物种概念。这里采用的生物学物种概念应该是修订过的生物学概念(modified biological species concept): 即种间存在较高程度的生殖隔离, 种间生殖隔离程度远大于种内居群间生殖隔离。
当然, 除上述这些物种概念的标准之外, 也可依据其他物种概念所需条件进行检查。检查的条件越多, 所依据的证据越多, 最后的分类处理应越客观和科学。对一些特定类群(如微生物、以无融合生殖为主的植物类群等)进行研究时, 要注意采用适合类群特点的物种概念。
对于所检查类群中的“物种”, 可能存在只符合一个物种概念到符合所有已检查物种概念的情况, 后者是毫无疑问的“好种”。关键是那些只符合一个至两个物种概念条件的两个类群, 是独立为两个种还是合并为一个物种?两种处理并没有本质上的区别(正如种和变种没有本质的区别一样, Darwin, 1859 ), 因为处于分化前期的物种总是这种情况; 但应尽量减少分类名称上的混乱, 遵循已有的分类处理。对于符合形态学物种概念的类群, 应优先考虑形态学标准, 因为这更容易让其他科学工作者认识和使用这样界定和发表的物种。发表新物种时, 至少应该符合两个物种概念条件。对于那些一个物种概念条件都不符合的物种, 应予以合并或命名为自然杂种(hybrid), 如: 依据居群内少数基因突变产生的变异个体以及种间杂交个体命名的物种, 这些个体尚未形成独立的进化单元。经过这样的物种界定和分类处理, 就能知道所处理的物种处于物种分化的什么时期, 也知道它们之间的分化主要存在于哪个方面, 从而为研究它们的物种形成机制及其保护提供更为可靠的分类学背景。
致谢: 感谢马克平主编邀请撰写这篇短文。感谢郑红蕾等同学的帮助和讨论。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Abbott RJ Hegarty MJ Hiscock SJ Brennan AC 2010 ) Homoploid hybrid speciation in action.
Taxon , 59 , 1375 -1386 .
[本文引用: 1]
[2]
Coyne JA Orr A 2004 ) Speciation. Sinauer Associates, Sunderland
, MA.
[本文引用: 6]
[3]
Darwin C 1859 ) On the Origin of Species by Means of Natural Selection or the Presentation of Favored Races in the Struggle for Life. John Murray, London.
[本文引用: 2]
[4]
de Queiroz K Donoghue MJ 1988 ) Phylogenetic systematics and the species problem.
Cladistics , 4 , 317 -338 .
[本文引用: 1]
[5]
Feder JL Flaxman SM Egan SP Comeault AA Nosil P 2013 ) Geographic mode of speciation and genomic divergence. Annual Review of Ecology,
Evolution and Systematics , 44 , 73 -97 .
[本文引用: 1]
[6]
Fennessy J Bidon T Reuss F Vamberger M Fritz W Jank A 2016 ) Multi-locus analyses reveal four giraffe species.
Current Biology , 26 , 1 -7 .
[本文引用: 2]
[7]
Grant V 1981 ) Plant Speciation . Columbia University Press , New York .
[本文引用: 2]
[8]
Mallet J 1995 ) A species definition for the modern synthesis.
Trends in Ecology & Evolution , 10 , 294 -299 .
[本文引用: 3]
[9]
Mao KS Liu JQ 2012 ) Current ‘relicts’ more dynamic in history than previously thought.
New Phytologist , 196 , 329 -331 .
[本文引用: 1]
[10]
Mayr E 1963 ) Animal Species and Evolution . Harvard University Press , Cambridge , Massachusetts.
[本文引用: 1]
[11]
Qi XS Chen C Comes HP Sakaguchi S Liu YH Tanaka N Sakio H Qiu YX 2012 ) Molecular data and ecological niche modelling reveal a highly dynamic evolutionary history of the East Asian Tertiary relict Cercidiphyllum (Cercidiphyllaceae).
New Phytologist , 196 , 617 -630 .
[本文引用: 1]
[12]
van Valen L 1976 ) Ecological species, multispecies, and oaks.
Taxon , 25 , 233 -239 .
[13]
Wilkins JS 2006 ) A list of 26 species “concepts” source.
. (accessed on 2016-08-01
URL
[本文引用: 2]
[14]
Wilkins JS 2009 ) Species : A History of the Idea. University of California Press, Berkeley .
[本文引用: 2]
[15]
Yassin A Capy P Madi-Ravazzi L Ogereau D David JR 2008 ) DNA barcode discovers two cryptic species and two geographical radiations in the invasive drosophilid Zaprionus indianus.
Molecular Ecology Resources , 8 , 491 -501 .
[本文引用: 1]
[16]
Zhou CF Yang G 2011 ) Existence and Definition of Species . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[周长发 , 杨光 (2011 ) 物种的存在与定义. 科学出版社 , 北京.]
[本文引用: 1]
Homoploid hybrid speciation in action.
1
2010
... 因为“整合物种概念”只有在物种分化后期才能完整体现, 所以在分化早中期尝试定义一个针对所有生物的物种概念似乎不可能做到: 因为对于不同的物种对来说, 它们的基因组结构、进化历史以及所受到的选择压力等都是不同的.在物种分化过程中, 有些可能是生态位分化早, 有些可能是形态分化较早, 而另一些则可能是生殖隔离分化较早, 情况极为复杂; 各物种分化条件在不同物种对中应是随机的.还须认识到, 自然界的物种分化不是进行到后期, 即两个或多个后代物种之间(如图2 )已经完整分化(符合所有物种概念)时才开始新一轮的物种分化; 往往是还在分化中(如早中期)就已开始新一轮的物种分化(图2 箭头所示), 这可能导致自然界的很多物种始终处于物种分化的早中期.理论上, 分化时间越长, 遗传差异越大、生殖隔离程度越高; 处于分化早中期的物种, 种间不可避免地产生一定的基因流.种间杂交以及基因流一方面会引起一些基因或DNA片段在种间遗传界限上弱化以及种间关系网状化; 另一方面也可能促进被渗入物种的环境适应性提高和分布区扩展, 甚至这些杂交后代还会形成一个新的同倍性杂交或异源多倍体物种(Coyne & Orr, 2004 ; Abbott et al, 2010 ). ...
Speciation. Sinauer Associates, Sunderland
6
2004
... 物种是生物学研究的基石, 任何生物学研究都离不开物种, 即以一个或多个物种为研究对象开展各种不同的研究(Darwin, 1859 ; Mallet, 1995 ; Coyne & Orr, 2004 ).但是, 物种又是生物学中争论最大的定义或概念(Mallet, 1995 ).如何定义物种?物种概念如何指导物种界定和分类修订以及其他研究?下面是我对这些问题的思考. ...
... 生物学物种是所有教科书介绍“物种”时都要介绍的经典概念.生殖隔离是生物学物种概念的核心(Mayr, 1963 ); 但在定义和阐述生物学物种概念时, 人们往往忽略了在群体水平上两个物种的生殖隔离存在一定的“度”, 从100% (所有的个体都存在生殖隔离)到0% (所有的个体之间都不存在生殖隔离).合子后的生殖隔离是生物学物种中最常见的支持证据; 但是两个近缘物种杂交后代雄性不育、雌性可育(Haldane’s rule)在很多动物类群中都十分常见(Coyne & Orr, 2004 ), 从而说明这些所谓的“生物学物种”也不是100%的生殖隔离, 因为雌性可育也可能导致种间界限打破, 使得基因在种间转移(Mallet, 1995 ).植物以及部分动物属中还广泛存在多倍化现象, 多倍化形成的物种一般认为是比较理想的生物学物种; 但是, 不同倍性的物种之间仍可能存在少量的杂交后代和基因流(Grant, 1981 ).因此, “生物学物种”概念应修订为“种间生殖隔离程度远大于种内居群间生殖隔离程度”才更加符合实际情况. ...
... 此外, 生物学物种概念在异域分布的地理隔离的近缘物种中应用起来十分不方便.地理隔离物种之间可能存在由于遗传漂变等机制导致多位点分化产生的合子后生殖隔离以及其他机制产生的交配前生殖隔离(Coyne & Orr, 2004 ); 但是, 这些隔离机制也可能在一些最近才异域隔离分布的姊妹形态学种中都不存在. ...
... 现代遗传学已经证实群体或物种间的分化都有其遗传学基础, 即本质上是DNA或基因的分化(Feder et al, 2013 ).每个物种概念成立所需的条件实际上是相应基因的分化.“一因多效” (一个基因多个效果)确实存在, 如控制形态性状分化的基因同时也导致生殖隔离以及生态位分化(Coyne & Orr, 2004 ).这种情况下, 一个基因的分化可能会满足形态学、生物学、遗传学和生态学4个甚至更多物种概念的条件.但是, 这样的基因和例子相对少见.多数情况下, 每个物种概念所需条件涉及的基因可能是不同的, 如系统发育分析常用的基因或DNA片段是近中性进化的, 而生态物种概念所依托的基因可能具有选择适应性; 生殖隔离和形态差异基因既可能是随机中性进化的产物, 也可能是自然选择导致的差异性表现.各个物种概念所需条件与两个物种群体水平上多个基因的分化式样一致: 有些基因快(选择或其他因素), 有些基因慢.在物种分化后期, 所有的基因都分离, 相当于所有物种概念的条件都得到满足, 成为“好种”.因此, 各物种概念所需条件在“分化路上”的分离与物种分化中的多基因分化式样是基本类似和相同的(图2 ). ...
... 因为“整合物种概念”只有在物种分化后期才能完整体现, 所以在分化早中期尝试定义一个针对所有生物的物种概念似乎不可能做到: 因为对于不同的物种对来说, 它们的基因组结构、进化历史以及所受到的选择压力等都是不同的.在物种分化过程中, 有些可能是生态位分化早, 有些可能是形态分化较早, 而另一些则可能是生殖隔离分化较早, 情况极为复杂; 各物种分化条件在不同物种对中应是随机的.还须认识到, 自然界的物种分化不是进行到后期, 即两个或多个后代物种之间(如图2 )已经完整分化(符合所有物种概念)时才开始新一轮的物种分化; 往往是还在分化中(如早中期)就已开始新一轮的物种分化(图2 箭头所示), 这可能导致自然界的很多物种始终处于物种分化的早中期.理论上, 分化时间越长, 遗传差异越大、生殖隔离程度越高; 处于分化早中期的物种, 种间不可避免地产生一定的基因流.种间杂交以及基因流一方面会引起一些基因或DNA片段在种间遗传界限上弱化以及种间关系网状化; 另一方面也可能促进被渗入物种的环境适应性提高和分布区扩展, 甚至这些杂交后代还会形成一个新的同倍性杂交或异源多倍体物种(Coyne & Orr, 2004 ; Abbott et al, 2010 ). ...
... 第三, 通过生态位模拟以及实地观察来验证是否符合生态学物种概念, 检查物种之间是否具有明显的生态位分化(如花期、行为、资源利用以及时空分布等).生态模型模拟表明, 两个独立进化的物种不可能具有完全重叠的生态位(Coyne & Orr, 2004 ). ...
On the Origin of Species by Means of Natural Selection or the Presentation of Favored Races in the Struggle for Life. John Murray, London.
2
1859
... 物种是生物学研究的基石, 任何生物学研究都离不开物种, 即以一个或多个物种为研究对象开展各种不同的研究(Darwin, 1859 ; Mallet, 1995 ; Coyne & Orr, 2004 ).但是, 物种又是生物学中争论最大的定义或概念(Mallet, 1995 ).如何定义物种?物种概念如何指导物种界定和分类修订以及其他研究?下面是我对这些问题的思考. ...
... 对于所检查类群中的“物种”, 可能存在只符合一个物种概念到符合所有已检查物种概念的情况, 后者是毫无疑问的“好种”.关键是那些只符合一个至两个物种概念条件的两个类群, 是独立为两个种还是合并为一个物种?两种处理并没有本质上的区别(正如种和变种没有本质的区别一样, Darwin, 1859 ), 因为处于分化前期的物种总是这种情况; 但应尽量减少分类名称上的混乱, 遵循已有的分类处理.对于符合形态学物种概念的类群, 应优先考虑形态学标准, 因为这更容易让其他科学工作者认识和使用这样界定和发表的物种.发表新物种时, 至少应该符合两个物种概念条件.对于那些一个物种概念条件都不符合的物种, 应予以合并或命名为自然杂种(hybrid), 如: 依据居群内少数基因突变产生的变异个体以及种间杂交个体命名的物种, 这些个体尚未形成独立的进化单元.经过这样的物种界定和分类处理, 就能知道所处理的物种处于物种分化的什么时期, 也知道它们之间的分化主要存在于哪个方面, 从而为研究它们的物种形成机制及其保护提供更为可靠的分类学背景. ...
Phylogenetic systematics and the species problem.
1
1988
... 到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
Geographic mode of speciation and genomic divergence. Annual Review of Ecology,
1
2013
... 现代遗传学已经证实群体或物种间的分化都有其遗传学基础, 即本质上是DNA或基因的分化(Feder et al, 2013 ).每个物种概念成立所需的条件实际上是相应基因的分化.“一因多效” (一个基因多个效果)确实存在, 如控制形态性状分化的基因同时也导致生殖隔离以及生态位分化(Coyne & Orr, 2004 ).这种情况下, 一个基因的分化可能会满足形态学、生物学、遗传学和生态学4个甚至更多物种概念的条件.但是, 这样的基因和例子相对少见.多数情况下, 每个物种概念所需条件涉及的基因可能是不同的, 如系统发育分析常用的基因或DNA片段是近中性进化的, 而生态物种概念所依托的基因可能具有选择适应性; 生殖隔离和形态差异基因既可能是随机中性进化的产物, 也可能是自然选择导致的差异性表现.各个物种概念所需条件与两个物种群体水平上多个基因的分化式样一致: 有些基因快(选择或其他因素), 有些基因慢.在物种分化后期, 所有的基因都分离, 相当于所有物种概念的条件都得到满足, 成为“好种”.因此, 各物种概念所需条件在“分化路上”的分离与物种分化中的多基因分化式样是基本类似和相同的(图2 ). ...
Multi-locus analyses reveal four giraffe species.
2
2016
... 尽管很难科学检验和统计, 但自然界中可能更多的物种应处于物种分化的早中期, 不符合所有物种概念定义的条件.极端的例子是多倍化新产生的四倍体群体, 在形态特征上与祖先二倍体没有统计上的显著差异; 但多倍化导致其生殖隔离程度较高(尽管不是100%隔离, 因为可能还存在通过三倍体桥的基因流动, Grant, 1981 ), 符合修订后的生物学物种概念, 是生物学物种, 但不符合其他物种概念条件.另一类例子是形态隐存种(morphologically cryptic species), 在最近的研究中出现十分频繁(如Yassin et al, 2008 ), 甚至包括知名物种长颈鹿(Giraffa camelopardalis , Fennessy et al, 2016如Fennessy et al, 2016 ); 但是形态性状差异统计不显著, 不符合形态学物种概念. ...
... ); 不同的隐存种之间存在遗传差异, 为遗传聚合群, 且各自为单系群, 因而符合遗传以及系统发育物种概念, 甚至也可能有地理分布差异而符合生态学物种概念(如Fennessy et al, 2016 ); 但是形态性状差异统计不显著, 不符合形态学物种概念. ...
2
1981
... 生物学物种是所有教科书介绍“物种”时都要介绍的经典概念.生殖隔离是生物学物种概念的核心(Mayr, 1963 ); 但在定义和阐述生物学物种概念时, 人们往往忽略了在群体水平上两个物种的生殖隔离存在一定的“度”, 从100% (所有的个体都存在生殖隔离)到0% (所有的个体之间都不存在生殖隔离).合子后的生殖隔离是生物学物种中最常见的支持证据; 但是两个近缘物种杂交后代雄性不育、雌性可育(Haldane’s rule)在很多动物类群中都十分常见(Coyne & Orr, 2004 ), 从而说明这些所谓的“生物学物种”也不是100%的生殖隔离, 因为雌性可育也可能导致种间界限打破, 使得基因在种间转移(Mallet, 1995 ).植物以及部分动物属中还广泛存在多倍化现象, 多倍化形成的物种一般认为是比较理想的生物学物种; 但是, 不同倍性的物种之间仍可能存在少量的杂交后代和基因流(Grant, 1981 ).因此, “生物学物种”概念应修订为“种间生殖隔离程度远大于种内居群间生殖隔离程度”才更加符合实际情况. ...
... 尽管很难科学检验和统计, 但自然界中可能更多的物种应处于物种分化的早中期, 不符合所有物种概念定义的条件.极端的例子是多倍化新产生的四倍体群体, 在形态特征上与祖先二倍体没有统计上的显著差异; 但多倍化导致其生殖隔离程度较高(尽管不是100%隔离, 因为可能还存在通过三倍体桥的基因流动, Grant, 1981 ), 符合修订后的生物学物种概念, 是生物学物种, 但不符合其他物种概念条件.另一类例子是形态隐存种(morphologically cryptic species), 在最近的研究中出现十分频繁(如Yassin et al, 2008 ), 甚至包括知名物种长颈鹿(Giraffa camelopardalis , Fennessy et al, 2016如Fennessy et al, 2016 ); 但是形态性状差异统计不显著, 不符合形态学物种概念. ...
A species definition for the modern synthesis.
3
1995
... 物种是生物学研究的基石, 任何生物学研究都离不开物种, 即以一个或多个物种为研究对象开展各种不同的研究(Darwin, 1859 ; Mallet, 1995 ; Coyne & Orr, 2004 ).但是, 物种又是生物学中争论最大的定义或概念(Mallet, 1995 ).如何定义物种?物种概念如何指导物种界定和分类修订以及其他研究?下面是我对这些问题的思考. ...
... ).但是, 物种又是生物学中争论最大的定义或概念(Mallet, 1995 ).如何定义物种?物种概念如何指导物种界定和分类修订以及其他研究?下面是我对这些问题的思考. ...
... 生物学物种是所有教科书介绍“物种”时都要介绍的经典概念.生殖隔离是生物学物种概念的核心(Mayr, 1963 ); 但在定义和阐述生物学物种概念时, 人们往往忽略了在群体水平上两个物种的生殖隔离存在一定的“度”, 从100% (所有的个体都存在生殖隔离)到0% (所有的个体之间都不存在生殖隔离).合子后的生殖隔离是生物学物种中最常见的支持证据; 但是两个近缘物种杂交后代雄性不育、雌性可育(Haldane’s rule)在很多动物类群中都十分常见(Coyne & Orr, 2004 ), 从而说明这些所谓的“生物学物种”也不是100%的生殖隔离, 因为雌性可育也可能导致种间界限打破, 使得基因在种间转移(Mallet, 1995 ).植物以及部分动物属中还广泛存在多倍化现象, 多倍化形成的物种一般认为是比较理想的生物学物种; 但是, 不同倍性的物种之间仍可能存在少量的杂交后代和基因流(Grant, 1981 ).因此, “生物学物种”概念应修订为“种间生殖隔离程度远大于种内居群间生殖隔离程度”才更加符合实际情况. ...
Current ‘relicts’ more dynamic in history than previously thought.
1
2012
... 现在看到的物种实际是其“进化”长河中的一个切面, 物种始终是在“分化路上”, 或处于分化的早期, 或处于中期, 或处于后期.产生的物种或者存留至今, 或者永久灭绝, 但更多情况下则是灭绝后又开始新一轮的物种分化, 周而复始.当处于分化的早中期时, 物种显示出来的特征可能只满足其中一个或两个物种概念条件, 而不能满足多个条件; 当处于分化后期时, 则能满足更多甚至所有物种概念的条件.但是, 自然界中处于分化后期的物种相对较少.古老孑遗单种属中的物种如银杏(Ginkgo biloba )、大熊猫(Ailuropoda melanoleuca )等, 以及少种属中分化时间相对较长的一些物种应属于这种情况, 这些物种就是分类上所谓的“好种”.但是, 有些古老孑遗属的少种属尽管属起源古老, 但所包括的物种却可能是新近历史上才开始分化的年轻物种, 如领春木(Cercidiphyllum magnificum )和日本领春木(C. japonicum ) (Mao & Liu, 2012 ; Qi et al, 2012 ), 它们之间的分化不一定符合所有物种概念的条件.这也说明一些所谓的“孑遗物种”实际上还是处在分化路上的“年轻”物种. ...
1
1963
... 生物学物种是所有教科书介绍“物种”时都要介绍的经典概念.生殖隔离是生物学物种概念的核心(Mayr, 1963 ); 但在定义和阐述生物学物种概念时, 人们往往忽略了在群体水平上两个物种的生殖隔离存在一定的“度”, 从100% (所有的个体都存在生殖隔离)到0% (所有的个体之间都不存在生殖隔离).合子后的生殖隔离是生物学物种中最常见的支持证据; 但是两个近缘物种杂交后代雄性不育、雌性可育(Haldane’s rule)在很多动物类群中都十分常见(Coyne & Orr, 2004 ), 从而说明这些所谓的“生物学物种”也不是100%的生殖隔离, 因为雌性可育也可能导致种间界限打破, 使得基因在种间转移(Mallet, 1995 ).植物以及部分动物属中还广泛存在多倍化现象, 多倍化形成的物种一般认为是比较理想的生物学物种; 但是, 不同倍性的物种之间仍可能存在少量的杂交后代和基因流(Grant, 1981 ).因此, “生物学物种”概念应修订为“种间生殖隔离程度远大于种内居群间生殖隔离程度”才更加符合实际情况. ...
Molecular data and ecological niche modelling reveal a highly dynamic evolutionary history of the East Asian Tertiary relict Cercidiphyllum (Cercidiphyllaceae).
1
2012
... 现在看到的物种实际是其“进化”长河中的一个切面, 物种始终是在“分化路上”, 或处于分化的早期, 或处于中期, 或处于后期.产生的物种或者存留至今, 或者永久灭绝, 但更多情况下则是灭绝后又开始新一轮的物种分化, 周而复始.当处于分化的早中期时, 物种显示出来的特征可能只满足其中一个或两个物种概念条件, 而不能满足多个条件; 当处于分化后期时, 则能满足更多甚至所有物种概念的条件.但是, 自然界中处于分化后期的物种相对较少.古老孑遗单种属中的物种如银杏(Ginkgo biloba )、大熊猫(Ailuropoda melanoleuca )等, 以及少种属中分化时间相对较长的一些物种应属于这种情况, 这些物种就是分类上所谓的“好种”.但是, 有些古老孑遗属的少种属尽管属起源古老, 但所包括的物种却可能是新近历史上才开始分化的年轻物种, 如领春木(Cercidiphyllum magnificum )和日本领春木(C. japonicum ) (Mao & Liu, 2012 ; Qi et al, 2012 ), 它们之间的分化不一定符合所有物种概念的条件.这也说明一些所谓的“孑遗物种”实际上还是处在分化路上的“年轻”物种. ...
Ecological species, multispecies, and oaks.
1976
A list of 26 species “concepts” source.
2
2006
... 到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
... )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
2
2009
... 到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
... , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
DNA barcode discovers two cryptic species and two geographical radiations in the invasive drosophilid Zaprionus indianus.
1
2008
... 尽管很难科学检验和统计, 但自然界中可能更多的物种应处于物种分化的早中期, 不符合所有物种概念定义的条件.极端的例子是多倍化新产生的四倍体群体, 在形态特征上与祖先二倍体没有统计上的显著差异; 但多倍化导致其生殖隔离程度较高(尽管不是100%隔离, 因为可能还存在通过三倍体桥的基因流动, Grant, 1981 ), 符合修订后的生物学物种概念, 是生物学物种, 但不符合其他物种概念条件.另一类例子是形态隐存种(morphologically cryptic species), 在最近的研究中出现十分频繁(如Yassin et al, 2008 ), 甚至包括知名物种长颈鹿(Giraffa camelopardalis , Fennessy et al, 2016如Fennessy et al, 2016 ); 但是形态性状差异统计不显著, 不符合形态学物种概念. ...
1
2011
... 到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...
1
2011
... 到目前为止, 有接近30个(Wilkins, 2006 , 2009 )甚至更多(周长发和杨光, 2001 )有关物种的定义.这些物种定义最常用的有5-7个(Wilkins, 2006 , 2009 ), 如形态学物种概念(morphological species concept)、生物学物种概念(biological species concept)、生态学物种概念(ecological species concept)、遗传学物种概念(genetic species concept)、进化物种概念(evolutionary species concept)和系统发育物种概念(phylogenetic taxon species concept)等, 其他定义都是从中衍生或综合其中2-3个物种概念而形成的(de Queiroz & Dono- ghue, 1988 ).其中, 形态学物种概念是生物多样性研究中定义肉眼可见的动植物物种时用得最多的物种概念, 是从模式物种概念(typological species concept)衍生而来.但是, 目前各种志书类书籍中记载的多数物种, 与其近缘物种的划分所依据的形态学性状大都没有进行过居群水平的统计检验, 尤其是数量性状.在形态学物种的现代处理过程中, “居群”和“统计显著差异”两个概念十分重要.一旦领悟了这两个概念, 就不难想象近缘物种之间可能存在部分中间个体; 但它们的存在不影响“统计显著差异”和是否存在不同的“形态聚合群” (morphological clusters)这一用于区分不同形态学物种的标准. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}