生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 )。随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重。鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 )。但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 )。针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 )。此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 )。
我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 )。Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 )。与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束。
图1 Web of Science 中关于入侵鱼类的文献数量(检索关键词为‘fish’和‘invasive’, 数据截止于2015年10月15日)
Fig. 1 Number of global studies on freshwater fish invasion in the world based on Web of Science
本文综述了全球淡水鱼类入侵的现状和研究进展, 包括鱼类入侵的定义及分布、入侵途径和机制、产生的生态和社会经济影响以及预防措施等诸方面的研究进展, 并展望了未来我国鱼类入侵的研究方向。
图2 2010-2015年世界各国淡水鱼类入侵文献量及其研究领域(数据来源于Web of Science, 截止于2015年10月15日)
Fig. 2 Fields and number of global studies in freshwater invasion during 2010-2015 (data acquired from Web of Science, by 2015.10.15)
1 鱼类入侵的定义及分布
1.1 鱼类入侵的定义
鱼类入侵是指鱼类由于人类干预到达历史上没有分布的水域系统, 并且能够自然繁殖, 维持种群稳定的过程。由于行政地理和自然地理的共同作用, 鱼类本地种和入侵种的划分还存在困难(Davis & Thompson, 2000 ; Copp et al, 2005a ; Gozlan et al, 2010 )。根据行政地理的定义, 鱼类跨越国界却在同一水系的移动被认为是入侵过程, 而从生态学的定义出发, 并非如此, 鱼类在同一个国家不同水系间的人为移动和建群, 同样能造成捕食、种内种间竞争、杂交、疾病传播等负面生态影响, 也是鱼类入侵(Gozlan et al, 2010 )。因此, 世界自然保护联盟(International Union for Conservation of Nature, IUCN)和《生物多样性公约》(Convention on Biological Diversity, CBD)规定入侵物种应基于自然地理而不是行政地理来定义。
1.2 全球外来鱼类的分布
全球外来鱼类共624种, 以丽鱼科和鲤科种类为主(Gozlan et al, 2008 )。已在引入地建群的外来种多为大量水产养殖的品种, 如多种鲟鱼和罗非鱼。部分岛国海水外来鱼类较多, 淡水外来鱼类较少。按地域分, 亚洲、南美洲、非洲的外来鱼类数量较多: 亚洲外来鱼类较多的国家有印度(228种)、中国(199种)、越南(149种)等; 南美洲外来鱼类较多的国家有巴西(235种)、玻利维亚(177种)、巴哈马(149种)等; 非洲外来鱼类较多的国家有安哥拉(198种)、埃及(189种)、博茨瓦纳(180种)等(fishbase.org)。除此之外, 全球外来鱼类较多的国家还有澳大利亚(254种)及美国(224种) (fishbase.org)。欧洲各国外来鱼类的数量都相对较少。由于对外来种定义和调查力度的差异, 以上统计数据可能并不能完全反映各国外来物种的现状, 如亚洲的鱼类总数大约是欧洲的10倍, 然而外来种的统计总数(957种)却与欧洲(705种)相当(Gozlan et al, 2008 ) (Fishbase.org)。因此, 各国统计数量还有待验证。
2 鱼类入侵途径和机制
2.1 鱼类入侵的路线
全球淡水鱼类入侵路线可分为以下三种: (1)大陆间的鱼类入侵; (2)大陆内的鱼类入侵; (3)同一国家不同水系之间的入侵。
2.1.1 大陆间的鱼类入侵
随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害。因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早。(1)欧洲到北美的入侵。鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河。虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 )。19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降。欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 )。(2)北美到欧洲的入侵。美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲。例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 )。(3)南美到欧洲的入侵。食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 )。
2.1.2 大陆内的鱼类入侵
人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵。(1)欧亚大陆内的入侵。20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 )。麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 )。除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 )。(2)欧洲各国间的入侵。来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 )。其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 )。(3)亚洲各国间的入侵。中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 )。
2.1.3 同一国家不同水系间的入侵
一些国家幅员辽阔, 水体类型多样, 仅本国境内就包括多条河流, 由于垂钓、饮食、种群保护等原因, 鱼种被引进到同一国家的不同水系, 形成入侵。如原分布于英格兰东南部的密歇根梅花鲈(Gymnocephalus cernua )和拟鲤(Rutilus rutilus )现已扩散到岛内各水系(Davies et al, 2004 ); 多瑙哲罗鱼(Hucho hucho )原产于波兰的2条河流, 后政府为保护种群将其投放到本国其他淡水水域, 造成入侵(Copp et al, 2005a )。原分布于美国落基山脉西侧的鲑鱼因垂钓需求被引进到加拿大, 现已广泛入侵加拿大各地区(Crossman, 1991 )。中国因水产养殖需求进行了大量不同区域之间的引种, 如从长江流域引进, 现广布于新疆博斯腾湖的河鲈(Perca fluviatilis ); 从广东、广西、湖南、湖北等地引入到云南的鳙、太湖新银鱼(Neosalanx taihuensis )、虾虎鱼、麦穗鱼等34种外来鱼类(楼允东, 2000 )。
2.2 鱼类入侵途径
与植物和鸟类入侵的途径不同, 鱼类入侵大多是由有意引种或者随有意引种的无意带入造成的。已有研究结果表明, 鱼类主要通过水产养殖(51%)、观赏渔业(21%)、休闲垂钓(12%)、渔业捕捞运输(7%)等多种途径成功入侵(Gozlan et al, 2010 ), 并对本土鱼类种群造成严重影响, 甚至导致了部分地方本土种群的灭绝(Gurevitch & Padilla, 2004 ; Gozlan et al, 2008 )。
(1)水产养殖。外来种由于其低投入、高产出和对环境高抗压性的优势, 被引入到各地作为水产养殖品种, 养殖过程中的逃逸和弃养造成了入侵, 如罗非鱼和非洲鲶鱼都属于这种入侵方式。统计表明, 水产养殖量与外来鱼类数量直接相关, 亚洲淡水水产养殖量排名前五的国家为中国(60.5%)、印度(9.9%)、越南(5.4%)、印度尼西亚(5.4%)、孟加拉国(4.0%), 而这些国家的外来鱼类数量也位于世界前列, 分别为199、228、149、107、132种, 以上数据间接验证了水产养殖为外来鱼类入侵的主要途径之一(附录1)。
(2)观赏渔业。欧洲和北美很多国家大量从东南亚、非洲、南美引进颜色鲜艳的观赏型鱼类用于展览和商品养殖, 在养殖过程中部分种群形成野外逃逸入侵(Keller & Lodge, 2007 ), 如鲫(Carassius auratus auratus )、非洲丽鱼、鲤鱼等。
(3)休闲垂钓。Cambray (2003)指出, 垂钓需求也是导致鱼类入侵及水生生物多样性下降的主要原因之一, 每年欧洲各国的垂钓业花费已经超过250亿欧元, 鲑鱼、鲈鱼等外来鱼类因当地垂钓和食用的需求被引入欧洲, 并且已广泛入侵到欧洲各国(Gozlan et al, 2008 )。鱼饵的使用也会造成鱼类入侵, 例如爱尔兰鲤科鱼种常用于白斑狗鱼(Esox lucius )垂钓的鱼饵(Caffrey et al, 2008 )。
(4)渔业捕捞运输。渔业捕捞运输会造成外来鱼类无意识带入, 其中包括随船舶压舱水带入、因水利工程的修建带入、或者混入引进养殖种类带入等(Bax et al, 2001 )。欧洲有大约25%的外来淡水水生生物通过船舶压舱水进入当地水域并成为入侵种(Gollasch, 2007 )。美国五大湖地区是世界上入侵物种最多的地区之一, 其中海七鳃鳗和虾虎鱼就是通过船舶压舱水被带入到五大湖地区并迅速形成大范围入侵(Wonham et al, 2000 )。
(5)其他。部分物种被引进以控制本地生态系统中的其他物种, 例如食蚊鱼被世界各国引进控制蚊子和疟疾; 亚洲鲤鱼被北美引进控制水生植物和藻类(Ciruna et al, 2004 )。我国鱼类入侵还与民间放生及政府增殖放流密切相关。民间放生为我国放流的主要来源之一, 大多由民间自发组织, 民众从水产市场或观赏鱼市场购买活鱼, 在就近水域进行放生。这些放生活动缺乏对放生种类和方法的调研, 其中很多放生种并非土著鱼种, 其放生对土著鱼类种质资源和当地水生生态环境造成破坏, 因此我国亟需对民间放生活动进行科学指导和规范管理。另外, 从20世纪50年代以来, 我国政府加大了内陆及近海的增殖放流, 按目的可分为“生态放流”和“生产性放流”。其中, “生态放流”的目标为提高水域的生物多样性(周军等, 2012 ); “生产性放流”的目标为增加江河湖泊及近海的渔业产量, 放流的淡水鱼类主要有青、草、鲢、鲤、鲂、鲑、鲴、鳗、鲟鱼等养殖常见种, 成为造成鱼类入侵的主要原因(尹增强和章守宇, 2008 )。
综上所述, 鱼类入侵大多由人类活动直接或间接造成。
2.3 鱼类成功入侵的机制
2.3.1 入侵鱼类的生物学特性
成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 )。(1)环境适应力。引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境。如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 )。(2)表型适应性。入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 )。(3)繁殖力。入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 )。(4)竞争力。ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点。入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 )。
2.3.2 入侵地的生态特征
鱼类的成功入侵与入侵地的生物多样性和生态位空缺有密切关系。鱼类的成功入侵往往发生在生物多样性低、生态结构不稳固的地区, 如欧洲波罗的海地区和北美五大湖地区相对年轻且物种本地化程度低, 入侵鱼类易成功入侵(Reid & Orlova, 2002 )。成功入侵的鱼类往往填补了入侵地空缺的生态位(Pettitt-Wade & Heath, 2015 )。入侵种和本地种虽可能在生态位的多个维度(dimensions of niche)发生重合, 但Hill等(2015)通过同位素的分析得知入侵物种至少填补了入侵地某一维度的空缺生态位。
2.4 我国鱼类入侵途径与现状
我国作为世界上淡水鱼类引种最多的国家, 现有外来鱼类共439种, 隶属于22目67科256属, 该数量远远超过多次统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xu HG et al, 2006 , 2012 ; 万方浩, 2009 )。其中外来种最多的四目为鲈形目(169种)、脂鲤目(65种)、鲇形目(61种)、鲤形目(49种), 已经确定入侵的外来鱼类共有53种(Xiong et al, 2015 )。
2000年之前, 水产养殖为我国鱼类入侵的主要原因(71种, 78.02%), 外来经济鱼类广泛分布在水产养殖量大的地区, 如广东、海南、广西、福建、云南等地, 数量最多的种类为尼罗罗非鱼(Tilapia nilotica ), 斑点叉尾鮰和四种鲟鱼(俄罗斯鲟Acipenser gueldenstaedti , 西伯利亚鲟A. baerii , 黑海鳇Huso huso , 匙吻鲟Polyodon spathula )。2000年以后, 观赏渔业因市场需求与引进政策的缺失逐渐成为我国鱼类入侵的第一大原因(347种, 79.04%) (Xiong et al, 2015 )。因其带来的经济效益, 政府鼓励发展观赏渔业并且提供技术支持, 除了西藏、新疆、宁夏、内蒙古等少数地区, 观赏鱼市场已遍布全国大部分地区, 尤其是北京、上海、广州等大城市, 部分物种由于逃逸或人为放生形成入侵。
我国从国外引进的经济鱼类主要来源于南美洲(156种, 35.54%)、亚洲(101种, 23.01%)、非洲(94种, 21.41%)和北美洲(52种, 11.85%)(Xiong et al, 2015 )。其中, 从非洲、越南、泰国等地区引进了多种罗非鱼, 造成了其在中国南方水系的入侵, 如齐式罗非鱼(T. zillii )、尼罗罗非鱼、加利亚罗非鱼(Sarotherodon galilaeus )等; 虹鳟于1959年由朝鲜引进到中国北方, 现已在中国北方低温淡水水域形成自然种群; 美国红鱼(Sciaenops ocellata )原分布于美国东南海岸和墨西哥湾沿岸, 目前已被引入到福建等多个地区养殖, 形成入侵(阮少江, 2002 )。除了从国外引进经济鱼类, 国内不同水系之间的引种也不断得到推广, 如现已遍布我国各地的四大家鱼青鱼(Mylopharyngodon piceus )、草鱼、鲢、鳙, 20世纪90年代在全国推广的银鱼科鱼类、银鲫(Carassius gibelio )、中华倒刺鲃(Spinibarbus sinesis )等(楼允东, 2000 )。
国内外来鱼类较多的水系主要有云南和青藏高原的淡水湖泊、新疆的塔里木河、博斯腾湖、额尔齐斯河等。自20世纪60年代以来, 云南先后从广东、广西、湖南、湖北等地引入外来种34种, 如鳙、太湖新银鱼、虾虎鱼、麦穗鱼等, 其中虾虎鱼随四大家鱼被引入到云南和青藏高原淡水湖泊后, 因食物和空间竞争, 造成了本地种数量的急剧减少, 如云南鲴(Xenocypris yunnanensis )、大头鲤(Cyprinus pellegrini )、裂腹鱼(Schizothorax spp.)等(李振宇和解焱, 2002 ; 潘勇等, 2007 )。从额尔齐斯河引入到博斯腾湖的河鲈直接导致了湖中扁吻鱼(Aspiorhynchus laticeps )的灭绝(李振宇和解焱, 2002 )。
3 鱼类入侵的影响
3.1 生态影响
入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等。
(1)捕食。入侵肉食鱼类因其强攻击性, 在本地大量捕食小型鱼类及幼鱼, 使其种群遭受威胁。如丰满叶唇鱼(Ptychocheilus grandis )造成美国加州本地幼鱼种群数的下降(Brown & Moyle, 1991 ); 尼罗河尖吻鲈(Lates niloticus )在非洲Victoria湖的入侵导致本地丽鱼种群的大量减少(Kitchell et al, 1997 )。一部分本地鱼为躲避入侵捕食者, 改变了原有的生活习性, 而被迫选择了其他生境。
(2)种间竞争。除了对食物网结构和功能的改变以外, 入侵活动还会导致生态位接近的物种间(interspecific)的竞争。比如北美五大湖地区两种入侵虾虎鱼(云斑原吻虾虎、黑口新虾虎)对北美小型淡水鲈类(darter)和鳅类(loach)等底栖鱼类种群造成了严重的威胁, 而黑口新虾虎又因其极强的可塑性(niche plasticity)在入侵范围和程度上超过云斑原吻虾虎(Pettitt-Wade & Heath, 2015 )。Caiola和Sostoa (2005)的研究表明霍氏食蚊鱼入侵伊比利亚半岛后与本地两种鳉形目小型鱼类(伊比利亚祕鳉Aphanius iberus , 西班牙鳉Valencia hispanica )竞争栖息地和食物, 造成了本地鱼种群数量下降; 食蚊鱼由于其极强的攻击性, 在入侵地大量食用本地鱼类的卵及幼鱼, 造成本地种种群数量严重下降(Goodsell & Kats, 1999 )。相近物种在同一入侵过程中可能对本地物种产生不同的影响。如Blanchet等(2007)通过实验室和野外实验研究了美洲红点鲑和虹鳟对本地欧洲褐鳟的生态影响, 结果证明虹鳟对欧洲褐鳟造成了危害, 而美洲红点鲑并没有对欧洲褐鳟造成生存或者生长的负面影响。
(3)入侵种与本地种的杂交。入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 )。比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 )。也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 )。
(4)栖息地破坏。入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 )。鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 )。再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 )。栖息地环境的改变也促进了其他适生物种的入侵。
(5)疾病传播。因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 )。外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 )。如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 )。有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究。例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题。再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害。
3.2 社会经济影响
在研究入侵物种带来的生态影响的同时, 社会经济影响也不可忽视。对同一入侵过程的评价往往因评价的角度不同得到完全相反的结果。
(1)负面影响。Pimentel等(2005)的研究结果表明, 美国每年由入侵鱼类造成的直接经济损失为54亿美元, 其中还不包括入侵物种的治理费用及本地种的保护费用。圣劳伦斯运河开通后, 入侵鱼类对本地经济鱼类造成严重危害, 每年用于治理和投苗的花费大约为2,000万美元左右(Lovell & Stone, 2006 )。五大湖渔业委员会(the Great Lakes Fishery Commission, GLFC)因七鳃鳗种群数得到控制一度减弱了对其治理的力度, 但是其种群数在短期内明显增加, 导致巨大的渔业经济损失(Adair & Sullivan, 2014 )。这也表明入侵物种一旦形成自然繁殖的种群之后很难根除, 并且治理花费大。
(2)正面影响。很多研究仅仅关注入侵活动产生的生态危害和负面社会经济影响, 却忽视了其可能带来的正面经济效应, 如入侵物种提供的休闲娱乐、食品供给和生态污染治理等功能(Gozlan et al, 2008 )。例如三文鱼在引进地智利的产量占世界产量的30%, 且给数十万人提供了就业机会(Iizuka & Katz, 2015 )。美国每年由入侵鱼类带来的垂钓收益为690亿美元。圣劳伦斯运河开通后, 北美五大湖地区年经济收益为40亿美元, 且提供了75,000个工作机会(FAO)。
以上案例表明因社会经济需求产生的鱼类引种不能避免, 我们必须从正反两方面论证和评价入侵活动带来的影响, 寻找有效的防范管理鱼类入侵的方法(Gozlan et al, 2008 )。
4 鱼类入侵的管理和预防
4.1 国际鱼类入侵的管理和预防
4.1.1 入侵管理机构和法律法规
世界各国为严格监控和管理包括鱼类入侵在内的生物入侵, 成立了很多政府和非政府机构, 并制定了相关法案。如世界自然保护联盟的外来入侵种专家组(the Invasive Species Specialist Group, ISSG)专门负责关注生物入侵问题; 有178个国家共同签署的《生物多样性公约》呼吁各国慎重引入外来种, 并严格控制入侵种的传播; 欧洲基于2007年通过的水产养殖过程中外来物种的引进议案, 建立了欧洲水产外来物种风险分析预测制度(European Non-native Species in Aquaculture Risk Analysis Scheme, ENSARS)来预测入侵风险; 美国于1990年通过的外来有害物种预防控制法案(the Nonindigenous Aquatic Nuisance Prevention and Control Act)严格监控引进种对生态环境的影响, 一旦发现某物种达到一定量的生态和经济危害, 即宣布其为有害物种, 并禁止引入(https://www.fws.gov/laws/law- sdigest/NONINDI.HTML)。
4.1.2 预测预警研究
由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 )。鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 )。近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009)。具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b )。由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 )。
4.2 我国鱼类入侵的管理和预防
近年来, 我国已充分意识到生物入侵的危害, 并在农业部、国家林业局、环境保护部、国家质检总局等多个单位设立了管理外来种的部门, 制定了与外来种相关的法律法规, 如《中华人民共和国进出境动植物检疫法》、《中华人民共和国渔业法》等。环境保护部在2003年和2010年先后发布了入侵种清单; 国家质检总局在海关口岸加强了对外来种的排查; 农业部和国家林业局制定了入侵种的应急预案。然而我国重大入侵种的管理与控制研究主要针对与农林业相关的陆生物种, 如互花米草(Spartina alterniflora )、椰心叶甲(Brontispa longissima )、美国白蛾(Hyphantria cunea )等(鞠瑞亭等, 2012 ), 入侵鱼类由于其隐蔽性, 研究案例较少。力度比较大的一次是2003年对入侵鱼类食人鲳的管控, 政府组织了全国范围食人鲳的捕杀, 但因我国还并未有专门的生物入侵管理部门, 对入侵物种的管控力度仍然不够, 因此食人鲳于近些年又偶见于花鸟鱼虫市场。除了物理捕杀外, 化学毒杀也是国际常用控制入侵鱼类的方法。尼罗罗非鱼为我国南方水域常见入侵种, 中国科学院珠江水产研究所研制了“灭非灵”对其进行毒杀, 取得了良好的实验效果, 目前未见对其他同水域常见鱼类造成危害(Ma et al, 2014 )。然而并未见类似化学药品的使用规范, 其对环境的影响评估也有待完善。
入侵风险评价系统能科学管理和预防生物入侵。在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系。以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性。鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系。
由此可见, 我国对外来鱼类的管理和预防还远远不够, 仍需在科研与管理层面上充分重视鱼类入侵问题。胡隐昌等(2012)指出, 我国对外来种入侵的防控存在“自然与经济错位”、“生态系统与行政区域错位”的问题, 即防控重视经济效益、忽视生态保护, 以行政区域而非生态系统为单位制定相关条例管理外来物种。这也造成了鱼类入侵研究投入经费不足且不集中, 各地的管理和监测不规范、投入力度差异大的现象。
5 展望
随着社会经济的发展, 生物入侵将持续影响人类环境和物种进化(Davis, 2010 )。联合国粮食与农业组织(FAO)在2014年的报告中指出: 全球渔业产量在过去的50年翻了5倍, 鱼类作为食品的年增长量为3.2%, 并且报告开篇就提出中国淡水鱼类养殖量占全球养殖量的60%, 是对全球渔业影响最大的国家(FAO, 2014 )。因为水产养殖是淡水鱼类入侵的最主要途径(51%), 因此我国面临严重的淡水鱼类入侵风险。然而我国在淡水鱼类入侵方面的研究文献数仅占全球的3.7%, 并且与全球相比, 我国尚无有效的外来鱼类入侵的管理和预防制度, 因此我国亟需加强鱼类入侵的研究, 制定相关制度和法案, 科学防范鱼类入侵。我们在全球的研究基础上, 结合中国鱼类入侵的特点和现状, 对未来鱼类入侵的研究提出以下几点建议:
(1)开展全国范围的本底调查并建立数据库, 实现数据共享, 明确鱼类入侵的历史与分布现状。由于大多数鱼类入侵由人为引种造成, 且鱼类入侵通常以不同自然水系为基本单位, 我们建议: (a)成立外来鱼类调查组对环保、农业、水产等各部门现有数据进行整合, 进行跨水域、跨地区的全国性调查; (b)突破已有的以省、直辖市为基本调查单位的现状, 以自然水域为单位开展调查, 调查范围应不仅局限于江河及入海口, 而应更深入到各水系上游; (c)在调查方案中明确外来种和入侵种的定义, 细化调查范围及步骤, 并在调查过程中严格取证, 撰写详细可验证的调查报告; (d)因水产养殖及观赏渔业为外来鱼类入侵的主要途径, 应对人工养殖场的饲养条件及废弃鱼塘的处理方法进行调查, 对易造成鱼类入侵的隐患加以规范。调查结果汇总之后, 应对其进行全面深入地分析, 聚焦外来鱼类入侵的重点研究区域和物种, 为进一步防控和管理提供依据。
(2)深入研究鱼类入侵的机制和影响。只有充分了解鱼类入侵的机制和影响, 才能有效合理地控制和防范鱼类入侵。我们建议从以下几个方面开展研究: (a)结合本底调查的结果, 重点选择入侵范围广、程度深的物种和研究地, 借鉴已知的生物入侵机制, 结合鱼类入侵的特性, 进行野外和室内实验, 由个别案例总结出入侵的普遍规律和机制; (b)建立入侵鱼类基因组数据库, 研究决定外来鱼类成功入侵的基因型, 基于基因组水平解释鱼类入侵机制; (c)突破以单一物种为研究对象的研究方式, 以水生生态系统为单位, 综合分析甲壳类、软体动物、水生植物、藻类等多种水生生物门类在鱼类入侵活动中的作用和变化, 评价鱼类入侵对整个水生生态系统结构、功能及能量流动的影响; (d)联合多部门进行长期观测, 基于现有数据客观评价鱼类入侵的生态及社会经济影响, 避免过早定性鱼类入侵活动及其产生的影响。
(3)加强鱼类入侵管理及预警的研究。鱼类入侵一旦发生就无法根除, 且持续治理花费大。因此应加大预警系统的研究力度, 并在此基础上促进相关法律法规的建立, 加强科普宣传, 建立合理的管理制度, 提高公众防范外来种的意识。我们建议: (a)承担水产管理职责的农业部门不应同时负责外来鱼类的入侵管理, 而应由环境保护部门成立专门的鱼类入侵防治部门, 该部门应对鱼类入侵严重的水域进行长期监测, 对目前鱼类入侵不严重但面临入侵风险的水域进行定期调查, 以自然水域为单位撰写调查报告, 并量化每年的治理方案; (b)整合现有的数据资源, 以自然水域为单位构建外来鱼类入侵风险评价系统, 该风险评价系统应对研究地外来鱼类是否入侵进行量化甄别, 聚焦重点研究物种, 为入侵种的长期跟踪分析提供依据; (c)严格规范水产养殖和观赏渔业的外来物种引进, 将预防和管理入侵鱼类的具体要求列入到相关部门和机构的工作计划和预算中; (d)建立公开的、可视的、专业的、合作的外来鱼类信息系统, 实现政府、科研单位、商业机构等多个利益方的数据共享, 从而更好地分析鱼类入侵的途径和机制, 对入侵活动进行实时的监督和报告; (e)加强科普宣传, 建立科学的管理制度, 对固定的放生活动, 尤其是大规模的宗教放生进行科学指导和管理, 在现场进行科普宣传, 并在常见放生区域组织科普活动, 树立明显的放生规范及咨询方式标识, 将鱼类入侵防范在源头。
(4)规范增殖放流。针对我国目前“重放流, 轻管理”的现象, 我们建议: (a)加强放流技术的研究, 制定科学的种质资源筛选标准, 对放流方法和放流环境进行充分前期调研, 避免放流种群与本地种群形成远缘杂交、遗传同化等负面生态影响; (b)设立增殖放流的管理机构, 科学有效地管理放流活动, 对放流个体进入渔业阶段后的状况进行长期监测和评估。
致谢: 特别感谢中国科学院水生生物研究所张鹗研究员、何德奎副研究员及华中农业大学杨瑞斌副教授对本文工作的支持; 感谢上海科技馆王晓娟教授对本文初稿提出的宝贵建议。
附录 Supplementary Material
Appendix 1 Farmed food fish production by top 15 producers in 2012 (data acquired from FAO)
http://www.biodiversity-science.net/fileup/PDF/2015374-1.pdf
参考文献
文献选项
[1]
Adair R Sullivan P 2014 ) Sea lamprey control in the Great Lakes. Annual report to the great lakes fishery commission. GLFC annual meeting grand rapids, Michigan.
(accessed on 2016-06-24
URL
[本文引用: 1]
[2]
Allendorf FW Leary RF Hitt NP Knudsen KL Lundquist LL Spruell AP 2004 ) Intercrosses and the U. S. Endangered Species Act: should hybridized populations be included as westslope cutthroat trout?
Conservation Biology , 18 , 1203 -1213 .
[本文引用: 1]
[3]
Almeida D Ribeiro F Leunda PM Vilizzi L Copp GH 2013 ) Effectiveness of FISK, an invasiveness screening tool for non-native freshwater fishes, to perform risk identification assessments in the Iberian Peninsula.
Risk Analysis , 33 , 1404 -1413 .
[本文引用: 2]
[4]
ArbaČIauskas K Lesutien ĖJ GasiŪNait ZR 2013 ) Feeding strategies and elemental composition in Ponto-Caspian peracaridans from contrasting environments: can stoichiometric plasticity promote invasion success?
Freshwater Biology , 58 , 1052 -1068 .
[本文引用: 1]
[5]
Ayala JR Rader RB Belk MC Schaalje GB 2007 ) Ground-truthing the impact of invasive species: spatio-temporal overlap between native least chub and introduced western mosquitofish.
Biological Invasions , 9 , 857 -869 .
[本文引用: 1]
[6]
Banarescu P 1964 ) Fauna. Republicii Populare Romine. Pisces-Osteichthyes, Vol. XIII. Academiei Republicii populare Romine, Bucuresti.
(in Romanian)
[本文引用: 1]
[7]
Bax N Carlton T Mathews-Amos A Headrich RL Howarth FG Purcell JE Rieser A Gray A 2001 ) The control of biological invasions in the world’s oceans.
Conservation Biology , 15 , 1234 -1246 .
[本文引用: 1]
[8]
Bergstrom MA Mensinger AF 2009 ) Interspecific resource competition between the invasive round goby and three native species: logperch, slimy sculpin, and spoonhead sculpin.
Transactions of the American Fisheries Society , 138 , 1009 -1017 .
[本文引用: 1]
[9]
Bianco PG 1988 ) Occurrence of the asiatic gobionid Pseudorasbora parva (Temminck and Schlegel) in south-eastern Europe.
Journal of Fish Biology , 32 , 973 -974 .
[本文引用: 1]
[10]
Bianco PG 1998 ) Freshwater fish transfers in Italy: history, local modification of fish composition, and a prediction on the future of native populations. In: Stocking and Introductions of Fishes (ed. Cowx IG) , pp. 165 -197. Blackwell , Oxford .
[本文引用: 1]
[11]
Blanc G 1997 ) Introduced pathogens in European aquatic ecosystems: theoretical aspects and realities.
Bulletin Francais de la Peche et de la Pisciculture , 0 , 489 -513 .
[本文引用: 1]
[12]
Blanchet S Loot G Grenouillet G Brosse S 2007 ) Competitive interactions between native and exotic salmonids: a combined field and laboratory demonstration.
Ecology of Freshwater Fish , 16 , 133 -143 .
[13]
Bogutskaya NG Naseka AM 2002 ) An overview of nonindigenous fishes in inland waters of Russia.
Proceedings of the Zoological Institute , 296 , 21 -30 .
[本文引用: 1]
[14]
Brown LR Moyle PB 1991 ) Changes in habitat and microhabitat partitioning within an assemblage of stream fishes in response to predation by Sacramento squawfish (Ptychocheilus grandis).
Canadian Journal of Fisheries and Aquatic Sciences , 48 , 849 -856 .
[本文引用: 4]
[15]
Brown LR Moyle PB 1997 ) Invading species in the Eel River, California: successes, failures, and relationships with resident species.
Environmental Biology of Fishes , 49 , 271 -291 .
[本文引用: 1]
[16]
Caffrey J Acevedo S Gallagher K Britton R 2008 ) Chub (Leuciscus cephalus): a new potentially invasive fish species in Ireland.
Aquatic Invasions , 3 , 201 -209 .
[本文引用: 1]
[17]
Caiola N Sostoa AD 2005 ) Possible reasons for the decline of two native toothcarps in the Iberian Peninsula: evidence of competition with the introduced eastern mosquitofish.
Journal of Applied Ichthyology , 21 , 358 -363 .
[本文引用: 1]
[18]
Cambray JA 2003 ) Impact on indigenous species biodiversity caused by the globalisation of alien recreational freshwater fisheries.
Hydrobiologia , 500 , 217 -230 .
[19]
Ciruna KA Meyerson LA Gutierrez A 2004 ) The ecological and socio-economic impacts of invasive alien species in inland water ecosystems. Report to the Conservation on Biological Diversity on behalf of the Global Invasive Species Programme, Washington, D.C.
pp. 34 .
[本文引用: 1]
[20]
Copp GH Fox MG Kovac V 2002 ) Growth, morphology and life history traits of a cool-water European population of pumpkinseed Lepomis gibbosus.
Archiv für Hydrobiologie , 155 , 585 -614 .
[本文引用: 1]
[21]
Copp GH 2013 ) The Fish Invasiveness Screening Kit (FISK) for non-native freshwater fishes: a summary of current applications.
Risk Analysis , 33 , 1394 -1396 .
[本文引用: 2]
[22]
Copp GH Bianco PG Bogutskaya NG Erős T Falka I Ferreira MT Fox MG Freyhof J Gozlan RE Grabowska J Kovac V Moreno-Amich R Naseka AM Penaz M Povz M Orzybylski M Robillard M Russell IC Stakenas S Sumer S Vila-Gispert A Wiesner C 2005 a) To be, or not to be, a non-native freshwater fish?
Journal of Applied Ichthyology , 21 , 242 -262 .
[本文引用: 4]
[23]
Copp GH Garthwaite R Gozlan RE 2005 b) Risk identification and assessment of non-native freshwater fishes: a summary of concepts and perspectives on protocols for the UK.
Journal of Applied Ichthyology , 21 , 371 .
[本文引用: 5]
[24]
Copp GH Vilizzi L Mumford J Fenwick GV Godard MJ Gozlan RE 2009 ) Calibration of FISK, an invasiveness screening tool for nonnative freshwater fishes.
Risk Analysis , 29 , 457 -467 .
[本文引用: 2]
[25]
Copp GH Wesley KJ Kovac V Ives MJ Carter MG 2003 ) Introduction and establishment of the pikeperch Stizostedion lucioperca (L.) in Stanborough Lake (Hertfordshire) and its dispersal in the Thames catchment.
The London Naturalist , 82 , 139 -153 .
[本文引用: 1]
[26]
Costedoat C Pech N Salducci MD Chappaz R Gilles A 2005 ) Evolution of mosaic hybrid zone between invasive and endemic species of Cyprinidae through space and time.
Biological Journal of the Linnean Society , 85 , 135 -155 .
[本文引用: 2]
[27]
Crossman EJ 1991 ) Introduced freshwater fishes: a review of the North American perspective with emphasis on Canada.
Canadian Journal of Fisheries and Aquatic Sciences , 48 , 46 -57 .
[本文引用: 1]
[28]
D’Amato ME Esterhuyse MM van der Waal BC Brink D Volckaert FA 2007 ) Hybridization and phylogeography of the Mozambique tilapia Oreochromis mossambicus in southern Africa evidenced by mitochondrial and microsatellite DNA genotyping.
Conservation Genetics , 8 , 475 -488 .
[本文引用: 2]
[29]
Davies C Shelly J Harding P McLean I Gardiner R Peirson G 2004 ) Freshwater Fishes in Britain: the Species and Their Distribution. Harley Books, Colchester.
[本文引用: 1]
[30]
Davis MA 2010 ) Biological invasions and society’s response.
BioScience , 60 , 651 -652 .
[本文引用: 1]
[31]
Davis MA Thompson K 2000 ) Eight ways to be a colonizer: two ways to be an invader: a proposed nomenclature scheme for invasion ecology.
Bulletin of the Ecological Society of America , 81 , 226 -230 .
[本文引用: 1]
[32]
Dextrase AJ Mandrak NE 2006 ) Impacts of alien invasive species on freshwater fauna at risk in Canada.
Biological Invasions , 8 , 13 -24 .
[本文引用: 1]
[33]
Didham RK Tylianakis JM Gemmell NJ Rand TA Ewers RM 2007 ) Interactive effects of habitat modification and species invasion on native species decline.
Trends in Ecology and Evolution , 22 , 489 -496 .
[本文引用: 1]
[34]
Didham RK Tylianakis JM Hutchison MA Ewers RM Gemmell NJ 2005 ) Are invasive species the drivers of ecological change?
Trends in Ecology and Evolution , 20 , 470 -474 .
[本文引用: 1]
[35]
Ding H Shi BQ Xu HG 2006 ) Index system and methodology for assessment of alien species.
Journal of Ecology and Rural Environment , 22 (2 ), 92 -96 . (in Chinese with English abstract)
[丁晖 , 石碧清 , 徐海根 (2006 ) 外来物种风险评估指标体系和评估方法
. 生态与农村环境学报 , 22 (2 ), 92 -96 .]
[36]
Ding H Xu HG Qiang S Meng L Han ZM Miao JL Hu BS Sun HY Huang C Lei JC Le ZF 2011 ) Status quo and trends of biological invasion into China.
Journal of Ecology and Rural Environment , 27 (3 ), 35 -41 . (in Chinese with English abstract)
[本文引用: 1]
[丁晖 , 徐海根 , 强胜 , 孟玲 , 韩正敏 , 缪锦来 , 胡白石 , 孙红英 , 黄成 , 雷军成 , 乐志芳 (2011 ) 中国生物入侵的现状与趋势
. 生态与农村环境学报 , 27 (3 ), 35 -41 .]
[本文引用: 1]
[37]
Dou Y Wu J Huang C 2011 ) Risk assessment system and method for invasion of alien fishes.
Journal of Ecology and Rural Environment , 27 (1 ), 12 -16 . (in Chinese with English abstract)
[本文引用: 1]
[窦寅 , 吴军 , 黄成 (2011 ) 外来鱼类入侵风险评估体系及方法
. 生态与农村环境学报 , 27 (1 ), 12 -16 .]
[本文引用: 1]
[38]
Food and Agriculture Organization of the United Nations (FAO ) (2014 ) The State of World Fisheries and Aquaculture.
(accessed on 2016- 06-18
URL
[本文引用: 2]
[39]
Gaughan DJ 2001 ) Disease-translocation across geographic boundaries must be recognized as a risk even in the absence of disease identification: the case with Australian Sardinops.
Reviews in Fish Biology and Fisheries , 11 , 113 -123 .
[40]
Giery ST Layman CA Langerhans RB 2015 ) Anthropogenic ecosystem fragmentation drives shared and unique patterns of sexual signal divergence among three species of Bahamian mosquitofish.
Evolutionary Applications , 8 , 679 -691 .
[41]
Gollasch S 2007 ) Marine vs. freshwater invaders: is shipping the key vector for species introductions to Europe? In: Biological Invaders in Inland Waters: Profiles, Distribution, and Threats. Francesca Gherardi, Springer, Houten.
[42]
Goodsell JA Kats LB 1999 ) Effect of introduced mosquitofish on Pacific treefrogs and the role of alternative prey.
Conservation Biology , 13 , 921 -924 .
[本文引用: 2]
[43]
Gozlan RE Britton JR Cowx I Copp GH 2010 ) Current knowledge on non-native freshwater fish introductions.
Journal of Fish Biology , 76 , 751 -786 .
[本文引用: 2]
[44]
Gozlan RE Newton AC Hulme PE Nentwig W Pysek P Vila M 2008 ) Biological invasions: benefits versus risks.
Science , 324 , 1015 .
[本文引用: 1]
[45]
Gozlan RE Peeler EJ Longshaw M St-Hilaire S Feist SW 2006 ) Effect of microbial pathogens on the diversity of aquatic populations, notably in Europe.
Microbes Infect , 8 , 1358 -1364 .
[本文引用: 1]
[46]
Gozlan RE St-Hilaire S Feist SW Martin P Kent ML 2005 ) Biodiversity: disease threat to European fish.
Nature , 435 , 1046 .
[本文引用: 5]
[47]
Grabowska J 2005 ) Reproductive biology of racer goby Neogobius gymnotrachelus in the Włocławski Reservoir (Vistula River, Poland).
Journal of Applied Ichthyology , 21 , 296 -299 .
[本文引用: 10]
[48]
Gurevitch J Marrow LL Wallace A Walsh JS 1992 ) A meta-analysis of competition in field experiments.
The American Naturalist , 140 , 539 -572 .
[本文引用: 2]
[49]
Gurevitch J Padilla DK 2004 ) Are invasive species a major cause of extinctions?
Trends in Ecology and Evolution , 19 , 470 -474 .
[本文引用: 3]
[50]
Hill JM Jones RW Hill MP Weyl OL 2015 ) Comparisons of isotopic niche widths of some invasive and indigenous fauna in a South African river.
Freshwater Biology , 60 , 893 -902 .
[本文引用: 3]
[51]
Hu YC Li Y Luo JR Tan XC 2006 ) Risk assessment system for alien aquatic animals.
Journal of Huazhong University of Science and Technology , 34 (10 ), 113 -115 . (in Chinese with English abstract)
[本文引用: 1]
[胡隐昌 , 李勇 , 罗建仁 , 谭细畅 (2006 ) 水生动物外来物种入侵风险评估系统的建立
. 华中科技大学学报 , 34 (10 ), 113 -115 .]
[本文引用: 1]
[52]
Hu YC Song HM Mou XD Luo JR 2012 ) Invasion of exotic species and their control measures taken in China.
Journal of Biosafety , 21 , 256 -261 . (in Chinese with English abstract)
[本文引用: 2]
[胡隐昌 , 宋红梅 , 牟希东 , 罗建仁 (2012 ) 浅议我国外来物种入侵问题及其防治对策
. 生物安全学报 , 21 , 256 -261 .]
[本文引用: 2]
[53]
Iizuka M Katz J 2015 ) Globalisation, sustainability and the role of institutions: the case of the Chilean salmon industry.
Tijdschrift Voor Economische En Sociale Geografie , 106 , 140 -153 .
[54]
Jeschke JM Strayer DL 2008 ) Are threat status and invasion success two sides of the same coin?
Ecography , 31 , 124 -130 .
[55]
Ju RT Li H Shi ZR Li B 2012 ) Progress of biological invasions research in China over the last decade.
Biodiversity Science , 20 , 581 -611 . (in Chinese with English abstract)
[本文引用: 1]
[鞠瑞亭 , 李慧 , 石正人 , 李博 (2012 ) 近十年中国生物入侵研究进展
. 生物多样性 , 20 , 581 -611 .]
[本文引用: 1]
[56]
Keller RP Lodge DM 2007 ) Species invasions from commerce in live aquatic organisms: problems and possible solutions.
BioScience , 57 , 428 -436 .
[57]
Khan TA Wilson ME Khan MT 2003 ) Evidence for invasive carp mediated trophic cascade in shallow lakes of western Victoria, Australia.
Hydrobiologia , 506 , 465 -472 .
[58]
Kitchell JF Schindler DE Ogutu-Ohwayo R Reinthal PN 1997 ) The Nile perch in Lake Victoria: interactions between predation and fisheries.
Ecological Applications , 7 , 653 -664 .
[本文引用: 1]
[59]
Koehn JD 2004 ) Carp (Cyprinus carpio) as a powerful invader in Australian waterways.
Freshwater Biology , 49 , 882 -894 .
[本文引用: 1]
[60]
Kolar CS Lodge DM 2002 ) Ecological predictions and risk assessment for alien fishes in North America.
Science , 298 , 1233 -1236 .
[61]
Kováč V Siryova S 2005 ) Ontogenetic variability in external morphology of bighead goby Neogobius kessleri from the Middle Danube, Slovakia.
Journal of Applied Ichthyology , 21 , 312 -315 .
[本文引用: 1]
[62]
Krupka I Meszaros J Rab P Šlechtová V 1989 ) Toward the preservation of endangered fish gene pools: example of wild danube carp (Cyprinus carpio carpio).
Práce VURHVodnany , 18 , 27 -33 .
[本文引用: 1]
[63]
Lawson LL Jr Hill JE Vilizzi L Hardin S Copp GH 2013 ) Revisions of the Fish Invasiveness Screening Kit (FISK) for its application in warmer climatic zones, with particular reference to Peninsular Florida.
Risk Analysis , 33 , 1414 -1431 .
[本文引用: 1]
[64]
Li ZY Xie Y 2002 ) Invasive Alien Species in China . China Forestry Publishing House , Beijing . (in Chinese)
[本文引用: 3]
[李振宇 , 解焱 (2002 ) 中国外来入侵种 . 中国林业出版社 , 北京 .]
[本文引用: 3]
[65]
Lou YD 2000 ) Present situation and countermeasures of the fish introduction in China.
Journal of Fisheries of China , 24 , 185 -192 . (in Chinese)
[本文引用: 1]
[楼允东 (2000 ) 我国鱼类引种研究的现状与对策
. 水产学报 , 24 , 185 -192 .]
[本文引用: 1]
[66]
Lovell SJ Stone SF 2006 ) The economic impacts of aquatic invasive species: a review of the literature.
Agricultural and Resource Economics Review , 35 , 195 .
[67]
Ma GM Gu DE Mou XD Luo JR Hu YC 2014 ) Poison effect of “Mie fei ling” on the alien Nile tilapia (Oreochromis niloticus).
Chinese Journal of Ecology , 33 , 2442 -2447 . (in Chinese with English abstract)
[本文引用: 1]
[马光明 , 顾党恩 , 牟希东 , 罗建仁 , 胡隐昌 (2014 ) “灭非灵”对外来鱼类尼罗罗非鱼的毒杀效果
. 生态学杂志 , 33 , 2442 -2447 .]
[本文引用: 1]
[68]
Ma XF Xiong BX Wang YD Wang MX 2003 ) Intentionally introduced and transferred fishes in China’s inland waters.
Asian Fisheries Science , 16 , 279 -290 .
[本文引用: 1]
[69]
Ma Y Xiong HJ Lin YH Tang SM 2009 ) Establishment of risk assessment system of marine invasive species.
Journal of Fisheries of China , 33 , 617 -623 . (in Chinese)
[本文引用: 1]
[马英 , 熊何健 , 林源洪 , 唐森铭 (2009 ) 外来海洋物种入侵风险评估体系的构建
. 水产学报 , 33 , 617 -623 .]
[本文引用: 1]
[70]
MacNeil C Prenter J 2000 ) Differential microdistributions and interspecific interactions in coexisting native and introduced Gammarus spp. (Crustacea: Amphipoda).
Journal of Zoology , 251 , 377 -384 .
[71]
Maitland PS 2004 ) Keys to the freshwater fish of Great Britain and Ireland with notes on their distribution and ecology.
Aquaculture Research , 36 , 828 .
[本文引用: 4]
[72]
McDowall RM 2006 ) Crying wolf, crying foul, or crying shame: alien salmonids and a biodiversity crisis in the southern cool-temperate galaxioid fishes?
Reviews in Fish Biology and Fisheries , 16 , 233 -422 .
[73]
McIntosh AR Townsend CR 1995 ) Contrasting predation risks presented by introduced brown trout and native common river galaxias in New Zealand streams.
Canadian Journal of Fisheries and Aquatic Sciences , 52 , 1821 -1833 .
[本文引用: 3]
[74]
Mikschi E 2002 ) Fische (Pisces). In: Neobiota in Osterreich (eds Essl F, Rabitsch W) , pp. 197 -204. Umweltbundesamt , Wien .
[本文引用: 1]
[75]
Miller SA Crowl TA 2006 ) Effects of common carp (Cyprinus carpio) on macrophytes and invertebrate communities in a shallow lake.
Freshwater Biology , 51 , 85 -94 .
[本文引用: 1]
[76]
Mills EL Leach JH Carlton JT Secor CL 1993 ) Exotic species in the Great Lakes: a history of biotic crises and anthropogenic introductions.
Journal of Great Lakes Research , 19 , 1 -54 .
[77]
Naseka AM Boldyrev VS Bogutskaya NG Delitsyn VV 2005 ) New data on the historical and expanded range of Proterorhinus marmoratus (Pallas, 1814) (Teleostei: Gobiidae) in eastern Europe.
Journal of Applied Ichthyology , 21 , 300 -305 .
[本文引用: 2]
[78]
Navodaru I Buijse AD Staras M 2002 ) Effects of hydrology and water quality on the fish community in Danube delta lakes.
International Review of Hydrobiology , 87 , 329 -348 .
[79]
Ondračková M Mávidová M Pečínková M Blažek R Gelnar M Valová Z Černý J Jurajda P 2005 ) Metazoan parasites of Neogobius fishes in the Slovak section of the River Danube.
Journal of Applied Ichthyology , 21 , 345 -349 .
[本文引用: 1]
[80]
Onikura N Nakajima J Inui R Mizutani H Kobayakawa M Fukuda S Mukai T 2011 ) Evaluating the potential for invasion by alien freshwater fishes in northern Kyushu Island, Japan, using the Fish Invasiveness Screening Kit.
Ichthyological Research , 58 , 382 -387 .
[本文引用: 1]
[81]
Ou J Lu CY 2007 ) Research of alien plant invasion risk assessment in Xiamen.
Journal of Xiamen University (Natural Science) , 45 , 883 -888 . (in Chinese)
[本文引用: 2]
[欧健 , 卢昌义 (2007 ) 厦门市外来植物入侵风险评价指标体系的研究
. 厦门大学学报(自然科学版) , 45 , 883 -888 .]
[本文引用: 2]
[82]
Pan Y Cao WX Xu LP Yin SR Bai L 2007 ) Process, mechanism, and research method of fish invasion.
Chinese Journal of Applied Ecology , 18 , 687 -692 . (in Chinese with English abstract)
[本文引用: 2]
[潘勇 , 曹文宣 , 徐立蒲 , 殷守仁 , 白璐 (2007 ) 鱼类入侵的过程、机制及研究方法
. 应用生态学报 , 18 , 687 -692 .]
[本文引用: 2]
[83]
Pettitt-Wade H Heath DD 2015 ) Niche plasticity in invasive fishes in the Great Lakes.
Biological Invasions , 17 , 2565 -2580 .
[本文引用: 1]
[84]
Pimentel D Zuniga R Morrison D 2005 ) Update on the environmental and economic costs associated with alien-invasive species in the United States.
Ecological Economics , 52 , 273 -288 .
[本文引用: 1]
[85]
Povž M Šumer S 2005 ) A brief review of non-native freshwater fishes in Slovenia.
Journal of Applied Ichthyology , 21 , 316 -318 .
[本文引用: 1]
[86]
Power ME 1992 ) Habitat heterogeneity and the functional signifiance of fish in river food webs.
Ecology , 73 , 1675 -1688 .
[本文引用: 1]
[87]
Puntila R Vilizzi L Lehtiniemi M Copp GH 2013 ) First application of FISK, the freshwater fish invasiveness screening kit, in northern Europe: example of southern Finland.
Risk Analysis , 33 , 1397 -1403 .
[本文引用: 1]
[88]
Rahel FJ Olden JD 2008 ) Assessing the effects of climate change on aquatic invasive species.
Conservation Biology , 22 , 521 -533 .
[本文引用: 1]
[89]
Rees DM 1945 ) Supplemental notes on mosquitofish in Utah, Gambusia affinis (Baird and Girard).
Copeia , 4 , 236 .
[本文引用: 1]
[90]
Reid D Orlova MI 2002 ) Geological and evolutionary underpinnings for the success of Ponto-Caspian species invasions in the Baltic Sea and North American Great Lakes.
Canadian Journal of Fisheries and Aquatic Sciences , 59 , 1144 -1158 .
[本文引用: 1]
[91]
Ruan SJ 2002 ) Reseach of alien species invasion in Mindong.
Journal of Ningde Teachers College (Natural Science) , 14 , 196 -198 . (in Chinese with English abstract)
[阮少江 (2002 ) 闽东外来生物入侵的初步研究
. 宁德师专学报(自然科学版) , 14 , 196 -198 .
[92]
Sagoff M 2005 ) Do non-native species threaten the natural environment?
Journal of Agricultural and Environmental Ethics , 18 , 215 -236 .
[93]
Sala OE Chapin FS Armesto JJ Berlow E Bloomfield J Dirzo R Huber-Sanwald E Huenneke LF Jackson RB Kinziq A Leemans R Lodge DM Mooney HA Oesterheld M Poff NL Sykes MT Walker BH Walker M Wall DH 2000 ) Global biodiversity scenarios for the year 2100.
Science , 287 , 1770 -1774 .
[94]
Scott B 1977 ) On the question of the introduction of the grass carp (Ctenopharyngodon idella Val.) into the United Kingdom.
Aquaculture Research , 8 , 63 -71 .
[本文引用: 1]
[95]
Scribner KT Page KS Bartron ML 2000 ) Hybridization in freshwater fishes: a review of case studies and cytonuclear methods of biological inference.
Reviews in Fish Biology and Fisheries , 10 , 293 -323 .
[本文引用: 2]
[96]
Simon KS Townsend CR Biggs BJF Bowden WB Frew RD 2004 ) Habitat-specific nitrogen dynamics in New Zealand streams containing native or invasive fish.
Ecosystems , 7 , 777 -792 .
[97]
Smartt J 2007 ) A possible genetic basis for species replacement: preliminary results of interspecific hybridisation between native crucian carp Carassius carassius (L.) and introduced goldfish Carassius auratus (L.).
Aquatic Invasions , 2 , 59 -62 .
[本文引用: 1]
[98]
Staub BP Hopkins WA Novak J Congdon JD 2004 ) Respiratory and reproductive characteristics of eastern mosquitofish (Gambusia holbrooki) inhabiting a coal ash settling basin.
Archives of Environmental Contamination and Toxicology , 46 , 96 -101 .
[本文引用: 1]
[99]
Stohlgren TJ Barnett D Flather C Fuller P Peterjohn B Kartesz J Master LL 2006 ) Species richness and patterns of invasion in plants, birds, and fishes in the United States.
Biological Invasions , 8 , 427 -447 .
[本文引用: 1]
[100]
Tarkan AS Ekmekci FG Vilizzi L Copp GH 2014 ) Risk screening of non-native freshwater fishes at the frontier between Asia and Europe: first application in Turkey of the fish invasiveness screening kit.
Journal of Applied Ichthyology , 30 , 392 -398 .
[本文引用: 2]
[101]
Vilizzi L Copp GH 2013 ) Application of FISK, an invasiveness screening tool for non-native freshwater fishes, in the Murray-Darling Basin (southeastern Australia).
Risk Analysis , 33 , 1432 -1440 .
[本文引用: 1]
[102]
Wan FH 2009 ) Research of Biological Invasions in China . Science Press , Beijing . (in Chinese)
[本文引用: 2]
[万方浩 (2009 ) 中国生物入侵研究 . 科学出版社 , 北京 .]
[本文引用: 2]
[103]
Wonham MJ Carlton JT Ruiz GM Smith LD 2000 ) Fish and ships: relating dispersal frequency to success in biological invasions.
Marine Biology , 136 , 1111 -1121 .
[104]
Xiong W Sui XY Liang SH Chen YF 2015 ) Non-native freshwater fish species in China.
Reviews in Fish Biology and Fisheries , 25 , 651 -687 .
[本文引用: 1]
[105]
Xu H Chen K Ouyang ZY Pan XB Zhu SF 2012 ) Threats of invasive species for China caused by expanding international trade.
Environmental Science and Technology , 46 , 7063 -7064 .
[本文引用: 1]
[106]
Xu HG Qiang S Genovesi P Ding H Wu J Meng L Han ZM Miao JL Hu BS Guo JG Sun HY Huang C Lei JC Le ZF Zhang XP He SP Wu Y Zheng Z Chen L Jarošík V Pyšek P 2012 ) An inventory of invasive alien species in China.
Neobiota , 15 , 1 -26 .
[本文引用: 2]
[107]
Xu HG Qiang S Han ZM Guo JY Huang ZG 2006 ) The status and causes of alien species invasion in China.
Biodiversity and Conservation , 15 , 2893 -2904 .
[本文引用: 2]
[108]
Yin ZQ Zhang SY 2008 ) Research of freshwater fish breeding and releasing station in China.
China Fisheries , (3 ), 9 -11 . (in Chinese)
[本文引用: 1]
[尹增强 , 章守宇 (2008 ) 对我国渔业资源增值放流问题的思考
. 中国水产 , (3 ), 9 -11 .]
[本文引用: 1]
[109]
Yonekura R Kohmatsu Y Yuma M 2007 ) Difference in the predation impact enhanced by morphological divergence between introduced fish populations.
Biological Journal of the Linnean Society , 91 , 601 -610 .
[本文引用: 1]
[110]
Zhou J Zhao ZL Xu YF Zhang HP Li YQ Fu Z Gao WB 2012 ) Current status, issue and suggestions of oceanic stock enhancement in Hebei Province, China.
Chinese Fisheries Economics , 6 (30 ), 111 -117 . (in Chinese)
[本文引用: 1]
[周军 , 赵振良 , 许玉甫 , 张海鹏 , 李怡群 , 付仲 , 高文斌 (2012 ) 河北省海洋渔业增值放流现状、问题及建议
. 中国渔业经济 , 6 (30 ), 111 -117 .]
[本文引用: 1]
[111]
Zimmerman JKH Vondracek B 2006 ) Interactions of slimy sculpin (Cottus cognatus) with native and nonnative trout: consequences for growth.
Canadian Journal of Fisheries and Aquatic Sciences , 63 , 1526 -1535 .
[本文引用: 1]
Sea lamprey control in the Great Lakes. Annual report to the great lakes fishery commission. GLFC annual meeting grand rapids, Michigan.
1
2014
... (1)负面影响.Pimentel等(2005)的研究结果表明, 美国每年由入侵鱼类造成的直接经济损失为54亿美元, 其中还不包括入侵物种的治理费用及本地种的保护费用.圣劳伦斯运河开通后, 入侵鱼类对本地经济鱼类造成严重危害, 每年用于治理和投苗的花费大约为2,000万美元左右(Lovell & Stone, 2006 ).五大湖渔业委员会(the Great Lakes Fishery Commission, GLFC)因七鳃鳗种群数得到控制一度减弱了对其治理的力度, 但是其种群数在短期内明显增加, 导致巨大的渔业经济损失(Adair & Sullivan, 2014 ).这也表明入侵物种一旦形成自然繁殖的种群之后很难根除, 并且治理花费大. ...
Intercrosses and the U. S. Endangered Species Act: should hybridized populations be included as westslope cutthroat trout?
1
2004
... (3)入侵种与本地种的杂交.入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 ).比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 ).也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 ). ...
Effectiveness of FISK, an invasiveness screening tool for non-native freshwater fishes, to perform risk identification assessments in the Iberian Peninsula.
2
2013
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Feeding strategies and elemental composition in Ponto-Caspian peracaridans from contrasting environments: can stoichiometric plasticity promote invasion success?
1
2013
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
Ground-truthing the impact of invasive species: spatio-temporal overlap between native least chub and introduced western mosquitofish.
1
2007
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
Fauna. Republicii Populare Romine. Pisces-Osteichthyes, Vol. XIII. Academiei Republicii populare Romine, Bucuresti.
1
1964
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
The control of biological invasions in the world’s oceans.
1
2001
... (4)渔业捕捞运输.渔业捕捞运输会造成外来鱼类无意识带入, 其中包括随船舶压舱水带入、因水利工程的修建带入、或者混入引进养殖种类带入等(Bax et al, 2001 ).欧洲有大约25%的外来淡水水生生物通过船舶压舱水进入当地水域并成为入侵种(Gollasch, 2007 ).美国五大湖地区是世界上入侵物种最多的地区之一, 其中海七鳃鳗和虾虎鱼就是通过船舶压舱水被带入到五大湖地区并迅速形成大范围入侵(Wonham et al, 2000 ). ...
Interspecific resource competition between the invasive round goby and three native species: logperch, slimy sculpin, and spoonhead sculpin.
1
2009
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
Occurrence of the asiatic gobionid Pseudorasbora parva (Temminck and Schlegel) in south-eastern Europe.
1
1988
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
1
1998
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Introduced pathogens in European aquatic ecosystems: theoretical aspects and realities.
1
1997
... (5)疾病传播.因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 ).外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 ).如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
Competitive interactions between native and exotic salmonids: a combined field and laboratory demonstration.
2007
An overview of nonindigenous fishes in inland waters of Russia.
1
2002
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Changes in habitat and microhabitat partitioning within an assemblage of stream fishes in response to predation by Sacramento squawfish (Ptychocheilus grandis).
4
1991
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (1)捕食.入侵肉食鱼类因其强攻击性, 在本地大量捕食小型鱼类及幼鱼, 使其种群遭受威胁.如丰满叶唇鱼(Ptychocheilus grandis )造成美国加州本地幼鱼种群数的下降(Brown & Moyle, 1991 ); 尼罗河尖吻鲈(Lates niloticus )在非洲Victoria湖的入侵导致本地丽鱼种群的大量减少(Kitchell et al, 1997 ).一部分本地鱼为躲避入侵捕食者, 改变了原有的生活习性, 而被迫选择了其他生境. ...
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
Invading species in the Eel River, California: successes, failures, and relationships with resident species.
1
1997
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
Chub (Leuciscus cephalus): a new potentially invasive fish species in Ireland.
1
2008
... (3)休闲垂钓.Cambray (2003)指出, 垂钓需求也是导致鱼类入侵及水生生物多样性下降的主要原因之一, 每年欧洲各国的垂钓业花费已经超过250亿欧元, 鲑鱼、鲈鱼等外来鱼类因当地垂钓和食用的需求被引入欧洲, 并且已广泛入侵到欧洲各国(Gozlan et al, 2008 ).鱼饵的使用也会造成鱼类入侵, 例如爱尔兰鲤科鱼种常用于白斑狗鱼(Esox lucius )垂钓的鱼饵(Caffrey et al, 2008 ). ...
Possible reasons for the decline of two native toothcarps in the Iberian Peninsula: evidence of competition with the introduced eastern mosquitofish.
1
2005
... 随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害.因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早.(1)欧洲到北美的入侵.鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河.虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 ).19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降.欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 ).(2)北美到欧洲的入侵.美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲.例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 ).(3)南美到欧洲的入侵.食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 ). ...
Impact on indigenous species biodiversity caused by the globalisation of alien recreational freshwater fisheries.
2003
The ecological and socio-economic impacts of invasive alien species in inland water ecosystems. Report to the Conservation on Biological Diversity on behalf of the Global Invasive Species Programme, Washington, D.C.
1
2004
... (5)其他.部分物种被引进以控制本地生态系统中的其他物种, 例如食蚊鱼被世界各国引进控制蚊子和疟疾; 亚洲鲤鱼被北美引进控制水生植物和藻类(Ciruna et al, 2004 ).我国鱼类入侵还与民间放生及政府增殖放流密切相关.民间放生为我国放流的主要来源之一, 大多由民间自发组织, 民众从水产市场或观赏鱼市场购买活鱼, 在就近水域进行放生.这些放生活动缺乏对放生种类和方法的调研, 其中很多放生种并非土著鱼种, 其放生对土著鱼类种质资源和当地水生生态环境造成破坏, 因此我国亟需对民间放生活动进行科学指导和规范管理.另外, 从20世纪50年代以来, 我国政府加大了内陆及近海的增殖放流, 按目的可分为“生态放流”和“生产性放流”.其中, “生态放流”的目标为提高水域的生物多样性(周军等, 2012 ); “生产性放流”的目标为增加江河湖泊及近海的渔业产量, 放流的淡水鱼类主要有青、草、鲢、鲤、鲂、鲑、鲴、鳗、鲟鱼等养殖常见种, 成为造成鱼类入侵的主要原因(尹增强和章守宇, 2008 ). ...
Growth, morphology and life history traits of a cool-water European population of pumpkinseed Lepomis gibbosus.
1
2002
... 随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害.因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早.(1)欧洲到北美的入侵.鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河.虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 ).19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降.欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 ).(2)北美到欧洲的入侵.美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲.例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 ).(3)南美到欧洲的入侵.食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 ). ...
The Fish Invasiveness Screening Kit (FISK) for non-native freshwater fishes: a summary of current applications.
2
2013
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
a) To be, or not to be, a non-native freshwater fish?
4
2005
... 鱼类入侵是指鱼类由于人类干预到达历史上没有分布的水域系统, 并且能够自然繁殖, 维持种群稳定的过程.由于行政地理和自然地理的共同作用, 鱼类本地种和入侵种的划分还存在困难(Davis & Thompson, 2000 ; Copp et al, 2005a ; Gozlan et al, 2010 ).根据行政地理的定义, 鱼类跨越国界却在同一水系的移动被认为是入侵过程, 而从生态学的定义出发, 并非如此, 鱼类在同一个国家不同水系间的人为移动和建群, 同样能造成捕食、种内种间竞争、杂交、疾病传播等负面生态影响, 也是鱼类入侵(Gozlan et al, 2010 ).因此, 世界自然保护联盟(International Union for Conservation of Nature, IUCN)和《生物多样性公约》(Convention on Biological Diversity, CBD)规定入侵物种应基于自然地理而不是行政地理来定义. ...
... 一些国家幅员辽阔, 水体类型多样, 仅本国境内就包括多条河流, 由于垂钓、饮食、种群保护等原因, 鱼种被引进到同一国家的不同水系, 形成入侵.如原分布于英格兰东南部的密歇根梅花鲈(Gymnocephalus cernua )和拟鲤(Rutilus rutilus )现已扩散到岛内各水系(Davies et al, 2004 ); 多瑙哲罗鱼(Hucho hucho )原产于波兰的2条河流, 后政府为保护种群将其投放到本国其他淡水水域, 造成入侵(Copp et al, 2005a ).原分布于美国落基山脉西侧的鲑鱼因垂钓需求被引进到加拿大, 现已广泛入侵加拿大各地区(Crossman, 1991 ).中国因水产养殖需求进行了大量不同区域之间的引种, 如从长江流域引进, 现广布于新疆博斯腾湖的河鲈(Perca fluviatilis ); 从广东、广西、湖南、湖北等地引入到云南的鳙、太湖新银鱼(Neosalanx taihuensis )、虾虎鱼、麦穗鱼等34种外来鱼类(楼允东, 2000 ). ...
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
b) Risk identification and assessment of non-native freshwater fishes: a summary of concepts and perspectives on protocols for the UK.
5
2005
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
... ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Calibration of FISK, an invasiveness screening tool for nonnative freshwater fishes.
2
2009
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Introduction and establishment of the pikeperch Stizostedion lucioperca (L.) in Stanborough Lake (Hertfordshire) and its dispersal in the Thames catchment.
1
2003
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Evolution of mosaic hybrid zone between invasive and endemic species of Cyprinidae through space and time.
2
2005
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (3)入侵种与本地种的杂交.入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 ).比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 ).也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 ). ...
Introduced freshwater fishes: a review of the North American perspective with emphasis on Canada.
1
1991
... 一些国家幅员辽阔, 水体类型多样, 仅本国境内就包括多条河流, 由于垂钓、饮食、种群保护等原因, 鱼种被引进到同一国家的不同水系, 形成入侵.如原分布于英格兰东南部的密歇根梅花鲈(Gymnocephalus cernua )和拟鲤(Rutilus rutilus )现已扩散到岛内各水系(Davies et al, 2004 ); 多瑙哲罗鱼(Hucho hucho )原产于波兰的2条河流, 后政府为保护种群将其投放到本国其他淡水水域, 造成入侵(Copp et al, 2005a ).原分布于美国落基山脉西侧的鲑鱼因垂钓需求被引进到加拿大, 现已广泛入侵加拿大各地区(Crossman, 1991 ).中国因水产养殖需求进行了大量不同区域之间的引种, 如从长江流域引进, 现广布于新疆博斯腾湖的河鲈(Perca fluviatilis ); 从广东、广西、湖南、湖北等地引入到云南的鳙、太湖新银鱼(Neosalanx taihuensis )、虾虎鱼、麦穗鱼等34种外来鱼类(楼允东, 2000 ). ...
Hybridization and phylogeography of the Mozambique tilapia Oreochromis mossambicus in southern Africa evidenced by mitochondrial and microsatellite DNA genotyping.
2
2007
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (3)入侵种与本地种的杂交.入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 ).比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 ).也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 ). ...
Freshwater Fishes in Britain: the Species and Their Distribution. Harley Books, Colchester.
1
2004
... 一些国家幅员辽阔, 水体类型多样, 仅本国境内就包括多条河流, 由于垂钓、饮食、种群保护等原因, 鱼种被引进到同一国家的不同水系, 形成入侵.如原分布于英格兰东南部的密歇根梅花鲈(Gymnocephalus cernua )和拟鲤(Rutilus rutilus )现已扩散到岛内各水系(Davies et al, 2004 ); 多瑙哲罗鱼(Hucho hucho )原产于波兰的2条河流, 后政府为保护种群将其投放到本国其他淡水水域, 造成入侵(Copp et al, 2005a ).原分布于美国落基山脉西侧的鲑鱼因垂钓需求被引进到加拿大, 现已广泛入侵加拿大各地区(Crossman, 1991 ).中国因水产养殖需求进行了大量不同区域之间的引种, 如从长江流域引进, 现广布于新疆博斯腾湖的河鲈(Perca fluviatilis ); 从广东、广西、湖南、湖北等地引入到云南的鳙、太湖新银鱼(Neosalanx taihuensis )、虾虎鱼、麦穗鱼等34种外来鱼类(楼允东, 2000 ). ...
Biological invasions and society’s response.
1
2010
... 随着社会经济的发展, 生物入侵将持续影响人类环境和物种进化(Davis, 2010 ).联合国粮食与农业组织(FAO)在2014年的报告中指出: 全球渔业产量在过去的50年翻了5倍, 鱼类作为食品的年增长量为3.2%, 并且报告开篇就提出中国淡水鱼类养殖量占全球养殖量的60%, 是对全球渔业影响最大的国家(FAO, 2014 ).因为水产养殖是淡水鱼类入侵的最主要途径(51%), 因此我国面临严重的淡水鱼类入侵风险.然而我国在淡水鱼类入侵方面的研究文献数仅占全球的3.7%, 并且与全球相比, 我国尚无有效的外来鱼类入侵的管理和预防制度, 因此我国亟需加强鱼类入侵的研究, 制定相关制度和法案, 科学防范鱼类入侵.我们在全球的研究基础上, 结合中国鱼类入侵的特点和现状, 对未来鱼类入侵的研究提出以下几点建议: ...
Eight ways to be a colonizer: two ways to be an invader: a proposed nomenclature scheme for invasion ecology.
1
2000
... 鱼类入侵是指鱼类由于人类干预到达历史上没有分布的水域系统, 并且能够自然繁殖, 维持种群稳定的过程.由于行政地理和自然地理的共同作用, 鱼类本地种和入侵种的划分还存在困难(Davis & Thompson, 2000 ; Copp et al, 2005a ; Gozlan et al, 2010 ).根据行政地理的定义, 鱼类跨越国界却在同一水系的移动被认为是入侵过程, 而从生态学的定义出发, 并非如此, 鱼类在同一个国家不同水系间的人为移动和建群, 同样能造成捕食、种内种间竞争、杂交、疾病传播等负面生态影响, 也是鱼类入侵(Gozlan et al, 2010 ).因此, 世界自然保护联盟(International Union for Conservation of Nature, IUCN)和《生物多样性公约》(Convention on Biological Diversity, CBD)规定入侵物种应基于自然地理而不是行政地理来定义. ...
Impacts of alien invasive species on freshwater fauna at risk in Canada.
1
2006
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
Interactive effects of habitat modification and species invasion on native species decline.
1
2007
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
Are invasive species the drivers of ecological change?
1
2005
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
中国生物入侵的现状与趋势
1
2011
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
中国生物入侵的现状与趋势
1
2011
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
外来鱼类入侵风险评估体系及方法
1
2011
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
外来鱼类入侵风险评估体系及方法
1
2011
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
The State of World Fisheries and Aquaculture.
2
2014
... 我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 ).Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 ).与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束. ...
... 随着社会经济的发展, 生物入侵将持续影响人类环境和物种进化(Davis, 2010 ).联合国粮食与农业组织(FAO)在2014年的报告中指出: 全球渔业产量在过去的50年翻了5倍, 鱼类作为食品的年增长量为3.2%, 并且报告开篇就提出中国淡水鱼类养殖量占全球养殖量的60%, 是对全球渔业影响最大的国家(FAO, 2014 ).因为水产养殖是淡水鱼类入侵的最主要途径(51%), 因此我国面临严重的淡水鱼类入侵风险.然而我国在淡水鱼类入侵方面的研究文献数仅占全球的3.7%, 并且与全球相比, 我国尚无有效的外来鱼类入侵的管理和预防制度, 因此我国亟需加强鱼类入侵的研究, 制定相关制度和法案, 科学防范鱼类入侵.我们在全球的研究基础上, 结合中国鱼类入侵的特点和现状, 对未来鱼类入侵的研究提出以下几点建议: ...
Disease-translocation across geographic boundaries must be recognized as a risk even in the absence of disease identification: the case with Australian Sardinops.
2001
Anthropogenic ecosystem fragmentation drives shared and unique patterns of sexual signal divergence among three species of Bahamian mosquitofish.
2015
Effect of introduced mosquitofish on Pacific treefrogs and the role of alternative prey.
2
1999
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (5)疾病传播.因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 ).外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 ).如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
Current knowledge on non-native freshwater fish introductions.
2
2010
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
... ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
Biological invasions: benefits versus risks.
1
2008
... (4)渔业捕捞运输.渔业捕捞运输会造成外来鱼类无意识带入, 其中包括随船舶压舱水带入、因水利工程的修建带入、或者混入引进养殖种类带入等(Bax et al, 2001 ).欧洲有大约25%的外来淡水水生生物通过船舶压舱水进入当地水域并成为入侵种(Gollasch, 2007 ).美国五大湖地区是世界上入侵物种最多的地区之一, 其中海七鳃鳗和虾虎鱼就是通过船舶压舱水被带入到五大湖地区并迅速形成大范围入侵(Wonham et al, 2000 ). ...
Effect of microbial pathogens on the diversity of aquatic populations, notably in Europe.
1
2006
... (2)种间竞争.除了对食物网结构和功能的改变以外, 入侵活动还会导致生态位接近的物种间(interspecific)的竞争.比如北美五大湖地区两种入侵虾虎鱼(云斑原吻虾虎、黑口新虾虎)对北美小型淡水鲈类(darter)和鳅类(loach)等底栖鱼类种群造成了严重的威胁, 而黑口新虾虎又因其极强的可塑性(niche plasticity)在入侵范围和程度上超过云斑原吻虾虎(Pettitt-Wade & Heath, 2015 ).Caiola和Sostoa (2005)的研究表明霍氏食蚊鱼入侵伊比利亚半岛后与本地两种鳉形目小型鱼类(伊比利亚祕鳉Aphanius iberus , 西班牙鳉Valencia hispanica )竞争栖息地和食物, 造成了本地鱼种群数量下降; 食蚊鱼由于其极强的攻击性, 在入侵地大量食用本地鱼类的卵及幼鱼, 造成本地种种群数量严重下降(Goodsell & Kats, 1999 ).相近物种在同一入侵过程中可能对本地物种产生不同的影响.如Blanchet等(2007)通过实验室和野外实验研究了美洲红点鲑和虹鳟对本地欧洲褐鳟的生态影响, 结果证明虹鳟对欧洲褐鳟造成了危害, 而美洲红点鲑并没有对欧洲褐鳟造成生存或者生长的负面影响. ...
Biodiversity: disease threat to European fish.
5
2005
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 鱼类入侵是指鱼类由于人类干预到达历史上没有分布的水域系统, 并且能够自然繁殖, 维持种群稳定的过程.由于行政地理和自然地理的共同作用, 鱼类本地种和入侵种的划分还存在困难(Davis & Thompson, 2000 ; Copp et al, 2005a ; Gozlan et al, 2010 ).根据行政地理的定义, 鱼类跨越国界却在同一水系的移动被认为是入侵过程, 而从生态学的定义出发, 并非如此, 鱼类在同一个国家不同水系间的人为移动和建群, 同样能造成捕食、种内种间竞争、杂交、疾病传播等负面生态影响, 也是鱼类入侵(Gozlan et al, 2010 ).因此, 世界自然保护联盟(International Union for Conservation of Nature, IUCN)和《生物多样性公约》(Convention on Biological Diversity, CBD)规定入侵物种应基于自然地理而不是行政地理来定义. ...
... ).根据行政地理的定义, 鱼类跨越国界却在同一水系的移动被认为是入侵过程, 而从生态学的定义出发, 并非如此, 鱼类在同一个国家不同水系间的人为移动和建群, 同样能造成捕食、种内种间竞争、杂交、疾病传播等负面生态影响, 也是鱼类入侵(Gozlan et al, 2010 ).因此, 世界自然保护联盟(International Union for Conservation of Nature, IUCN)和《生物多样性公约》(Convention on Biological Diversity, CBD)规定入侵物种应基于自然地理而不是行政地理来定义. ...
... 与植物和鸟类入侵的途径不同, 鱼类入侵大多是由有意引种或者随有意引种的无意带入造成的.已有研究结果表明, 鱼类主要通过水产养殖(51%)、观赏渔业(21%)、休闲垂钓(12%)、渔业捕捞运输(7%)等多种途径成功入侵(Gozlan et al, 2010 ), 并对本土鱼类种群造成严重影响, 甚至导致了部分地方本土种群的灭绝(Gurevitch & Padilla, 2004 ; Gozlan et al, 2008 ). ...
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Reproductive biology of racer goby Neogobius gymnotrachelus in the Włocławski Reservoir (Vistula River, Poland).
10
2005
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 全球外来鱼类共624种, 以丽鱼科和鲤科种类为主(Gozlan et al, 2008 ).已在引入地建群的外来种多为大量水产养殖的品种, 如多种鲟鱼和罗非鱼.部分岛国海水外来鱼类较多, 淡水外来鱼类较少.按地域分, 亚洲、南美洲、非洲的外来鱼类数量较多: 亚洲外来鱼类较多的国家有印度(228种)、中国(199种)、越南(149种)等; 南美洲外来鱼类较多的国家有巴西(235种)、玻利维亚(177种)、巴哈马(149种)等; 非洲外来鱼类较多的国家有安哥拉(198种)、埃及(189种)、博茨瓦纳(180种)等(fishbase.org).除此之外, 全球外来鱼类较多的国家还有澳大利亚(254种)及美国(224种) (fishbase.org).欧洲各国外来鱼类的数量都相对较少.由于对外来种定义和调查力度的差异, 以上统计数据可能并不能完全反映各国外来物种的现状, 如亚洲的鱼类总数大约是欧洲的10倍, 然而外来种的统计总数(957种)却与欧洲(705种)相当(Gozlan et al, 2008 ) (Fishbase.org).因此, 各国统计数量还有待验证. ...
... ).已在引入地建群的外来种多为大量水产养殖的品种, 如多种鲟鱼和罗非鱼.部分岛国海水外来鱼类较多, 淡水外来鱼类较少.按地域分, 亚洲、南美洲、非洲的外来鱼类数量较多: 亚洲外来鱼类较多的国家有印度(228种)、中国(199种)、越南(149种)等; 南美洲外来鱼类较多的国家有巴西(235种)、玻利维亚(177种)、巴哈马(149种)等; 非洲外来鱼类较多的国家有安哥拉(198种)、埃及(189种)、博茨瓦纳(180种)等(fishbase.org).除此之外, 全球外来鱼类较多的国家还有澳大利亚(254种)及美国(224种) (fishbase.org).欧洲各国外来鱼类的数量都相对较少.由于对外来种定义和调查力度的差异, 以上统计数据可能并不能完全反映各国外来物种的现状, 如亚洲的鱼类总数大约是欧洲的10倍, 然而外来种的统计总数(957种)却与欧洲(705种)相当(Gozlan et al, 2008 ) (Fishbase.org).因此, 各国统计数量还有待验证. ...
... 与植物和鸟类入侵的途径不同, 鱼类入侵大多是由有意引种或者随有意引种的无意带入造成的.已有研究结果表明, 鱼类主要通过水产养殖(51%)、观赏渔业(21%)、休闲垂钓(12%)、渔业捕捞运输(7%)等多种途径成功入侵(Gozlan et al, 2010 ), 并对本土鱼类种群造成严重影响, 甚至导致了部分地方本土种群的灭绝(Gurevitch & Padilla, 2004 ; Gozlan et al, 2008 ). ...
... (3)休闲垂钓.Cambray (2003)指出, 垂钓需求也是导致鱼类入侵及水生生物多样性下降的主要原因之一, 每年欧洲各国的垂钓业花费已经超过250亿欧元, 鲑鱼、鲈鱼等外来鱼类因当地垂钓和食用的需求被引入欧洲, 并且已广泛入侵到欧洲各国(Gozlan et al, 2008 ).鱼饵的使用也会造成鱼类入侵, 例如爱尔兰鲤科鱼种常用于白斑狗鱼(Esox lucius )垂钓的鱼饵(Caffrey et al, 2008 ). ...
... (5)疾病传播.因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 ).外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 ).如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
... , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
... (2)正面影响.很多研究仅仅关注入侵活动产生的生态危害和负面社会经济影响, 却忽视了其可能带来的正面经济效应, 如入侵物种提供的休闲娱乐、食品供给和生态污染治理等功能(Gozlan et al, 2008 ).例如三文鱼在引进地智利的产量占世界产量的30%, 且给数十万人提供了就业机会(Iizuka & Katz, 2015 ).美国每年由入侵鱼类带来的垂钓收益为690亿美元.圣劳伦斯运河开通后, 北美五大湖地区年经济收益为40亿美元, 且提供了75,000个工作机会(FAO). ...
... 以上案例表明因社会经济需求产生的鱼类引种不能避免, 我们必须从正反两方面论证和评价入侵活动带来的影响, 寻找有效的防范管理鱼类入侵的方法(Gozlan et al, 2008 ). ...
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
A meta-analysis of competition in field experiments.
2
1992
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (5)疾病传播.因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 ).外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 ).如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
Are invasive species a major cause of extinctions?
3
2004
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (5)疾病传播.因水产养殖需求引进的外来种还会导致疾病传播(Blanc, 1997 ; Gozlan et al, 2006 , 2008 ).外来种作为病原体的载体, 将疾病传给本地种, 而本地物种往往不具备该种疾病的抗体, 大面积的感染导致本地种种群数下降(Gaughan, 2001 ; Gozlan et al, 2005 ).如麦穗鱼于20世纪60年代被引入罗马尼亚并扩散到整个欧洲地区, 其携带的病原体Sphaerothecum destruens 对本地物种小赤梢鱼(Leucaspius delineates )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
... )造成危害, 导致了该种在全欧洲范围内濒危(Gozlan et al, 2005 , 2008 ).有些疾病对本地种和本地生态环境长期缓慢的影响不容易被发现, 因而有很多由外来种造成的疾病案例还有待研究.例如, 亚洲鲤鱼并没有在所有引入地产生良好的控制水生植物和藻类的效果, 反而将其寄生生物带入引入地成为当地生态环境新的问题.再如, 寄生于草鱼的鳋属(Ergasilus spp.)随鱼类引进被带入英国, 对本地鱼种造成危害. ...
Comparisons of isotopic niche widths of some invasive and indigenous fauna in a South African river.
3
2015
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
... ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
水生动物外来物种入侵风险评估系统的建立
1
2006
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
水生动物外来物种入侵风险评估系统的建立
1
2006
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
浅议我国外来物种入侵问题及其防治对策
2
2012
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 与植物和鸟类入侵的途径不同, 鱼类入侵大多是由有意引种或者随有意引种的无意带入造成的.已有研究结果表明, 鱼类主要通过水产养殖(51%)、观赏渔业(21%)、休闲垂钓(12%)、渔业捕捞运输(7%)等多种途径成功入侵(Gozlan et al, 2010 ), 并对本土鱼类种群造成严重影响, 甚至导致了部分地方本土种群的灭绝(Gurevitch & Padilla, 2004 ; Gozlan et al, 2008 ). ...
浅议我国外来物种入侵问题及其防治对策
2
2012
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 与植物和鸟类入侵的途径不同, 鱼类入侵大多是由有意引种或者随有意引种的无意带入造成的.已有研究结果表明, 鱼类主要通过水产养殖(51%)、观赏渔业(21%)、休闲垂钓(12%)、渔业捕捞运输(7%)等多种途径成功入侵(Gozlan et al, 2010 ), 并对本土鱼类种群造成严重影响, 甚至导致了部分地方本土种群的灭绝(Gurevitch & Padilla, 2004 ; Gozlan et al, 2008 ). ...
Globalisation, sustainability and the role of institutions: the case of the Chilean salmon industry.
2015
Are threat status and invasion success two sides of the same coin?
2008
近十年中国生物入侵研究进展
1
2012
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
近十年中国生物入侵研究进展
1
2012
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
Species invasions from commerce in live aquatic organisms: problems and possible solutions.
2007
Evidence for invasive carp mediated trophic cascade in shallow lakes of western Victoria, Australia.
2003
The Nile perch in Lake Victoria: interactions between predation and fisheries.
1
1997
... (2)正面影响.很多研究仅仅关注入侵活动产生的生态危害和负面社会经济影响, 却忽视了其可能带来的正面经济效应, 如入侵物种提供的休闲娱乐、食品供给和生态污染治理等功能(Gozlan et al, 2008 ).例如三文鱼在引进地智利的产量占世界产量的30%, 且给数十万人提供了就业机会(Iizuka & Katz, 2015 ).美国每年由入侵鱼类带来的垂钓收益为690亿美元.圣劳伦斯运河开通后, 北美五大湖地区年经济收益为40亿美元, 且提供了75,000个工作机会(FAO). ...
Carp (Cyprinus carpio) as a powerful invader in Australian waterways.
1
2004
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
Ecological predictions and risk assessment for alien fishes in North America.
2002
Ontogenetic variability in external morphology of bighead goby Neogobius kessleri from the Middle Danube, Slovakia.
1
2005
... 近年来, 我国已充分意识到生物入侵的危害, 并在农业部、国家林业局、环境保护部、国家质检总局等多个单位设立了管理外来种的部门, 制定了与外来种相关的法律法规, 如《中华人民共和国进出境动植物检疫法》、《中华人民共和国渔业法》等.环境保护部在2003年和2010年先后发布了入侵种清单; 国家质检总局在海关口岸加强了对外来种的排查; 农业部和国家林业局制定了入侵种的应急预案.然而我国重大入侵种的管理与控制研究主要针对与农林业相关的陆生物种, 如互花米草(Spartina alterniflora )、椰心叶甲(Brontispa longissima )、美国白蛾(Hyphantria cunea )等(鞠瑞亭等, 2012 ), 入侵鱼类由于其隐蔽性, 研究案例较少.力度比较大的一次是2003年对入侵鱼类食人鲳的管控, 政府组织了全国范围食人鲳的捕杀, 但因我国还并未有专门的生物入侵管理部门, 对入侵物种的管控力度仍然不够, 因此食人鲳于近些年又偶见于花鸟鱼虫市场.除了物理捕杀外, 化学毒杀也是国际常用控制入侵鱼类的方法.尼罗罗非鱼为我国南方水域常见入侵种, 中国科学院珠江水产研究所研制了“灭非灵”对其进行毒杀, 取得了良好的实验效果, 目前未见对其他同水域常见鱼类造成危害(Ma et al, 2014 ).然而并未见类似化学药品的使用规范, 其对环境的影响评估也有待完善. ...
Toward the preservation of endangered fish gene pools: example of wild danube carp (Cyprinus carpio carpio).
1
1989
... (2)观赏渔业.欧洲和北美很多国家大量从东南亚、非洲、南美引进颜色鲜艳的观赏型鱼类用于展览和商品养殖, 在养殖过程中部分种群形成野外逃逸入侵(Keller & Lodge, 2007 ), 如鲫(Carassius auratus auratus )、非洲丽鱼、鲤鱼等. ...
Revisions of the Fish Invasiveness Screening Kit (FISK) for its application in warmer climatic zones, with particular reference to Peninsular Florida.
1
2013
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
3
2002
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (1)捕食.入侵肉食鱼类因其强攻击性, 在本地大量捕食小型鱼类及幼鱼, 使其种群遭受威胁.如丰满叶唇鱼(Ptychocheilus grandis )造成美国加州本地幼鱼种群数的下降(Brown & Moyle, 1991 ); 尼罗河尖吻鲈(Lates niloticus )在非洲Victoria湖的入侵导致本地丽鱼种群的大量减少(Kitchell et al, 1997 ).一部分本地鱼为躲避入侵捕食者, 改变了原有的生活习性, 而被迫选择了其他生境. ...
3
2002
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (1)捕食.入侵肉食鱼类因其强攻击性, 在本地大量捕食小型鱼类及幼鱼, 使其种群遭受威胁.如丰满叶唇鱼(Ptychocheilus grandis )造成美国加州本地幼鱼种群数的下降(Brown & Moyle, 1991 ); 尼罗河尖吻鲈(Lates niloticus )在非洲Victoria湖的入侵导致本地丽鱼种群的大量减少(Kitchell et al, 1997 ).一部分本地鱼为躲避入侵捕食者, 改变了原有的生活习性, 而被迫选择了其他生境. ...
我国鱼类引种研究的现状与对策
1
2000
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
我国鱼类引种研究的现状与对策
1
2000
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
The economic impacts of aquatic invasive species: a review of the literature.
2006
“灭非灵”对外来鱼类尼罗罗非鱼的毒杀效果
1
2014
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
“灭非灵”对外来鱼类尼罗罗非鱼的毒杀效果
1
2014
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Intentionally introduced and transferred fishes in China’s inland waters.
1
2003
... 随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害.因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早.(1)欧洲到北美的入侵.鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河.虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 ).19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降.欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 ).(2)北美到欧洲的入侵.美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲.例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 ).(3)南美到欧洲的入侵.食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 ). ...
外来海洋物种入侵风险评估体系的构建
1
2009
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
外来海洋物种入侵风险评估体系的构建
1
2009
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Differential microdistributions and interspecific interactions in coexisting native and introduced Gammarus spp. (Crustacea: Amphipoda).
2000
Keys to the freshwater fish of Great Britain and Ireland with notes on their distribution and ecology.
4
2004
... 我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 ).Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 ).与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束. ...
... 我国作为世界上淡水鱼类引种最多的国家, 现有外来鱼类共439种, 隶属于22目67科256属, 该数量远远超过多次统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xu HG et al, 2006 , 2012 ; 万方浩, 2009 ).其中外来种最多的四目为鲈形目(169种)、脂鲤目(65种)、鲇形目(61种)、鲤形目(49种), 已经确定入侵的外来鱼类共有53种(Xiong et al, 2015 ). ...
... 国内外来鱼类较多的水系主要有云南和青藏高原的淡水湖泊、新疆的塔里木河、博斯腾湖、额尔齐斯河等.自20世纪60年代以来, 云南先后从广东、广西、湖南、湖北等地引入外来种34种, 如鳙、太湖新银鱼、虾虎鱼、麦穗鱼等, 其中虾虎鱼随四大家鱼被引入到云南和青藏高原淡水湖泊后, 因食物和空间竞争, 造成了本地种数量的急剧减少, 如云南鲴(Xenocypris yunnanensis )、大头鲤(Cyprinus pellegrini )、裂腹鱼(Schizothorax spp.)等(李振宇和解焱, 2002 ; 潘勇等, 2007 ).从额尔齐斯河引入到博斯腾湖的河鲈直接导致了湖中扁吻鱼(Aspiorhynchus laticeps )的灭绝(李振宇和解焱, 2002 ). ...
... )的灭绝(李振宇和解焱, 2002 ). ...
Crying wolf, crying foul, or crying shame: alien salmonids and a biodiversity crisis in the southern cool-temperate galaxioid fishes?
2006
Contrasting predation risks presented by introduced brown trout and native common river galaxias in New Zealand streams.
3
1995
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
... 一些国家幅员辽阔, 水体类型多样, 仅本国境内就包括多条河流, 由于垂钓、饮食、种群保护等原因, 鱼种被引进到同一国家的不同水系, 形成入侵.如原分布于英格兰东南部的密歇根梅花鲈(Gymnocephalus cernua )和拟鲤(Rutilus rutilus )现已扩散到岛内各水系(Davies et al, 2004 ); 多瑙哲罗鱼(Hucho hucho )原产于波兰的2条河流, 后政府为保护种群将其投放到本国其他淡水水域, 造成入侵(Copp et al, 2005a ).原分布于美国落基山脉西侧的鲑鱼因垂钓需求被引进到加拿大, 现已广泛入侵加拿大各地区(Crossman, 1991 ).中国因水产养殖需求进行了大量不同区域之间的引种, 如从长江流域引进, 现广布于新疆博斯腾湖的河鲈(Perca fluviatilis ); 从广东、广西、湖南、湖北等地引入到云南的鳙、太湖新银鱼(Neosalanx taihuensis )、虾虎鱼、麦穗鱼等34种外来鱼类(楼允东, 2000 ). ...
... 我国从国外引进的经济鱼类主要来源于南美洲(156种, 35.54%)、亚洲(101种, 23.01%)、非洲(94种, 21.41%)和北美洲(52种, 11.85%)(Xiong et al, 2015 ).其中, 从非洲、越南、泰国等地区引进了多种罗非鱼, 造成了其在中国南方水系的入侵, 如齐式罗非鱼(T. zillii )、尼罗罗非鱼、加利亚罗非鱼(Sarotherodon galilaeus )等; 虹鳟于1959年由朝鲜引进到中国北方, 现已在中国北方低温淡水水域形成自然种群; 美国红鱼(Sciaenops ocellata )原分布于美国东南海岸和墨西哥湾沿岸, 目前已被引入到福建等多个地区养殖, 形成入侵(阮少江, 2002 ).除了从国外引进经济鱼类, 国内不同水系之间的引种也不断得到推广, 如现已遍布我国各地的四大家鱼青鱼(Mylopharyngodon piceus )、草鱼、鲢、鳙, 20世纪90年代在全国推广的银鱼科鱼类、银鲫(Carassius gibelio )、中华倒刺鲃(Spinibarbus sinesis )等(楼允东, 2000 ). ...
1
2002
... (1)负面影响.Pimentel等(2005)的研究结果表明, 美国每年由入侵鱼类造成的直接经济损失为54亿美元, 其中还不包括入侵物种的治理费用及本地种的保护费用.圣劳伦斯运河开通后, 入侵鱼类对本地经济鱼类造成严重危害, 每年用于治理和投苗的花费大约为2,000万美元左右(Lovell & Stone, 2006 ).五大湖渔业委员会(the Great Lakes Fishery Commission, GLFC)因七鳃鳗种群数得到控制一度减弱了对其治理的力度, 但是其种群数在短期内明显增加, 导致巨大的渔业经济损失(Adair & Sullivan, 2014 ).这也表明入侵物种一旦形成自然繁殖的种群之后很难根除, 并且治理花费大. ...
Effects of common carp (Cyprinus carpio) on macrophytes and invertebrate communities in a shallow lake.
1
2006
... 近年来, 我国已充分意识到生物入侵的危害, 并在农业部、国家林业局、环境保护部、国家质检总局等多个单位设立了管理外来种的部门, 制定了与外来种相关的法律法规, 如《中华人民共和国进出境动植物检疫法》、《中华人民共和国渔业法》等.环境保护部在2003年和2010年先后发布了入侵种清单; 国家质检总局在海关口岸加强了对外来种的排查; 农业部和国家林业局制定了入侵种的应急预案.然而我国重大入侵种的管理与控制研究主要针对与农林业相关的陆生物种, 如互花米草(Spartina alterniflora )、椰心叶甲(Brontispa longissima )、美国白蛾(Hyphantria cunea )等(鞠瑞亭等, 2012 ), 入侵鱼类由于其隐蔽性, 研究案例较少.力度比较大的一次是2003年对入侵鱼类食人鲳的管控, 政府组织了全国范围食人鲳的捕杀, 但因我国还并未有专门的生物入侵管理部门, 对入侵物种的管控力度仍然不够, 因此食人鲳于近些年又偶见于花鸟鱼虫市场.除了物理捕杀外, 化学毒杀也是国际常用控制入侵鱼类的方法.尼罗罗非鱼为我国南方水域常见入侵种, 中国科学院珠江水产研究所研制了“灭非灵”对其进行毒杀, 取得了良好的实验效果, 目前未见对其他同水域常见鱼类造成危害(Ma et al, 2014 ).然而并未见类似化学药品的使用规范, 其对环境的影响评估也有待完善. ...
Exotic species in the Great Lakes: a history of biotic crises and anthropogenic introductions.
1993
New data on the historical and expanded range of Proterorhinus marmoratus (Pallas, 1814) (Teleostei: Gobiidae) in eastern Europe.
2
2005
... 我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 ).Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 ).与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束. ...
... 我国作为世界上淡水鱼类引种最多的国家, 现有外来鱼类共439种, 隶属于22目67科256属, 该数量远远超过多次统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xu HG et al, 2006 , 2012 ; 万方浩, 2009 ).其中外来种最多的四目为鲈形目(169种)、脂鲤目(65种)、鲇形目(61种)、鲤形目(49种), 已经确定入侵的外来鱼类共有53种(Xiong et al, 2015 ). ...
Effects of hydrology and water quality on the fish community in Danube delta lakes.
2002
Metazoan parasites of Neogobius fishes in the Slovak section of the River Danube.
1
2005
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
Evaluating the potential for invasion by alien freshwater fishes in northern Kyushu Island, Japan, using the Fish Invasiveness Screening Kit.
1
2011
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
厦门市外来植物入侵风险评价指标体系的研究
2
2007
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
厦门市外来植物入侵风险评价指标体系的研究
2
2007
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
... 入侵风险评价系统能科学管理和预防生物入侵.在这方面, 我国已有相关研究: 如欧健和卢昌义(2007) 以互花米草入侵风险评价为例建立了厦门地区外来植物入侵风险评价体系及方法; 丁晖等(2006) 结合外来物种的种类、引进途径、生物生态学特征以及危害特点的差异, 提出了基于3个层次、17个指标层构建的外来物种风险评估体系; 胡隐昌等(2006) 建立了水生动物外来物种信息数据库, 包括名录检索、引用文献检索和专家名录检索等相关内容, 初步构建了水生动物外来物种入侵风险评估的基础; 马英等(2009) 建立了外来海洋物种入侵风险评估体系.以上研究多以个别物种在较小的研究区域内的应用和研究为主, 缺乏研究深度及普适性.鱼类入侵评价体系的研究很少, 仅有窦寅等(2011) 初步构建了一套包含5个一级指标、12个二级指标、44个三级指标的外来鱼类入侵风险量化评估指标体系. ...
鱼类入侵的过程、机制及研究方法
2
2007
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
鱼类入侵的过程、机制及研究方法
2
2007
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
Niche plasticity in invasive fishes in the Great Lakes.
1
2015
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
Update on the environmental and economic costs associated with alien-invasive species in the United States.
1
2005
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
A brief review of non-native freshwater fishes in Slovenia.
1
2005
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
Habitat heterogeneity and the functional signifiance of fish in river food webs.
1
1992
... 随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害.因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早.(1)欧洲到北美的入侵.鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河.虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 ).19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降.欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 ).(2)北美到欧洲的入侵.美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲.例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 ).(3)南美到欧洲的入侵.食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 ). ...
First application of FISK, the freshwater fish invasiveness screening kit, in northern Europe: example of southern Finland.
1
2013
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Assessing the effects of climate change on aquatic invasive species.
1
2008
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Supplemental notes on mosquitofish in Utah, Gambusia affinis (Baird and Girard).
1
1945
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
Geological and evolutionary underpinnings for the success of Ponto-Caspian species invasions in the Baltic Sea and North American Great Lakes.
1
2002
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Do non-native species threaten the natural environment?
2005
Global biodiversity scenarios for the year 2100.
2000
On the question of the introduction of the grass carp (Ctenopharyngodon idella Val.) into the United Kingdom.
1
1977
... 国内外来鱼类较多的水系主要有云南和青藏高原的淡水湖泊、新疆的塔里木河、博斯腾湖、额尔齐斯河等.自20世纪60年代以来, 云南先后从广东、广西、湖南、湖北等地引入外来种34种, 如鳙、太湖新银鱼、虾虎鱼、麦穗鱼等, 其中虾虎鱼随四大家鱼被引入到云南和青藏高原淡水湖泊后, 因食物和空间竞争, 造成了本地种数量的急剧减少, 如云南鲴(Xenocypris yunnanensis )、大头鲤(Cyprinus pellegrini )、裂腹鱼(Schizothorax spp.)等(李振宇和解焱, 2002 ; 潘勇等, 2007 ).从额尔齐斯河引入到博斯腾湖的河鲈直接导致了湖中扁吻鱼(Aspiorhynchus laticeps )的灭绝(李振宇和解焱, 2002 ). ...
Hybridization in freshwater fishes: a review of case studies and cytonuclear methods of biological inference.
2
2000
... 鱼类的成功入侵与入侵地的生物多样性和生态位空缺有密切关系.鱼类的成功入侵往往发生在生物多样性低、生态结构不稳固的地区, 如欧洲波罗的海地区和北美五大湖地区相对年轻且物种本地化程度低, 入侵鱼类易成功入侵(Reid & Orlova, 2002 ).成功入侵的鱼类往往填补了入侵地空缺的生态位(Pettitt-Wade & Heath, 2015 ).入侵种和本地种虽可能在生态位的多个维度(dimensions of niche)发生重合, 但Hill等(2015)通过同位素的分析得知入侵物种至少填补了入侵地某一维度的空缺生态位. ...
... (2)种间竞争.除了对食物网结构和功能的改变以外, 入侵活动还会导致生态位接近的物种间(interspecific)的竞争.比如北美五大湖地区两种入侵虾虎鱼(云斑原吻虾虎、黑口新虾虎)对北美小型淡水鲈类(darter)和鳅类(loach)等底栖鱼类种群造成了严重的威胁, 而黑口新虾虎又因其极强的可塑性(niche plasticity)在入侵范围和程度上超过云斑原吻虾虎(Pettitt-Wade & Heath, 2015 ).Caiola和Sostoa (2005)的研究表明霍氏食蚊鱼入侵伊比利亚半岛后与本地两种鳉形目小型鱼类(伊比利亚祕鳉Aphanius iberus , 西班牙鳉Valencia hispanica )竞争栖息地和食物, 造成了本地鱼种群数量下降; 食蚊鱼由于其极强的攻击性, 在入侵地大量食用本地鱼类的卵及幼鱼, 造成本地种种群数量严重下降(Goodsell & Kats, 1999 ).相近物种在同一入侵过程中可能对本地物种产生不同的影响.如Blanchet等(2007)通过实验室和野外实验研究了美洲红点鲑和虹鳟对本地欧洲褐鳟的生态影响, 结果证明虹鳟对欧洲褐鳟造成了危害, 而美洲红点鲑并没有对欧洲褐鳟造成生存或者生长的负面影响. ...
Habitat-specific nitrogen dynamics in New Zealand streams containing native or invasive fish.
2004
A possible genetic basis for species replacement: preliminary results of interspecific hybridisation between native crucian carp Carassius carassius (L.) and introduced goldfish Carassius auratus (L.).
1
2007
... 随着国际间贸易交流的日益深化和航运的发展, 大陆间的鱼类入侵愈发普遍, 且因在引入地的绝对优势, 对本地种和本地生态系统造成严重危害.因欧洲与美洲间贸易交流非常频繁, 导致了鱼类入侵现象严重, 相关研究开始较早.(1)欧洲到北美的入侵.鲤鱼作为最早从亚洲经欧洲引入到北美洲的物种, 于19世纪50年代被有意引入到安大略湖和哈得孙河.虽然鲤鱼在欧洲的入侵早于北美, 但是鲤鱼在北美本土迅速形成了自然繁殖的种群, 并且扩散速度和范围远超过欧洲(Krupka et al, 1989 ).19世纪后期欧洲褐鳟(Salmo trutta )被引入北美, 造成了美国其他几种本土鳟鱼种群数的下降.欧洲本土鱼种, 如海七鳃鳗(Petromyzon marinus )、云斑原吻虾虎(Proterorhinus marmoratus )和黑口新虾虎(Neogobius melanostomus )经圣劳伦斯河, 由船舶压舱水被带入北美五大湖地区, 迅速形成自然繁殖种群, 严重威胁了本地底栖小型鱼类的生存(Mills et al, 1993 ).(2)北美到欧洲的入侵.美国淡水鱼类从19世纪中后期开始因饮食和垂钓的需求被大规模引进到欧洲.例如德国从北美引进的美洲红点鲑(Salvelinus fontinalis )、 虹鳟(Oncorhynchus mykiss )、云斑鮰(Ameiurus nebulosus )、小口黑鲈(Micropterus dolomieu )、大口黑鲈(M. salmoides ); 立陶宛从北美引进大口黑鲈、小口黑鲈、小体鲟(Acipenser ruthenus )、鲱形白鲑(Coregonus clupeaformis ); 意大利除引进与德国相似品种, 还引进了黑鮰(Ameiurus melas )、斑点叉尾鮰(Ictalurus punctatus )和突吻红点鲑(Salvelinus namaycush ); 法国、英国、西班牙从美国引进了驼背太阳鱼(Lepomis gibbosus ) (Copp et al, 2002 ; Povž & Šumer, 2005 ).(3)南美到欧洲的入侵.食蚊鱼因其对疟疾的控制作用首先从南美被引进到西班牙, 后到德国、意大利等国家, 因其极强的环境适应性, 现已遍布欧洲温水水域, 造成了对本地种的危害(Caiola & Sostoa, 2005 ). ...
Respiratory and reproductive characteristics of eastern mosquitofish (Gambusia holbrooki) inhabiting a coal ash settling basin.
1
2004
... (4)栖息地破坏.入侵物种能够通过改变生态系统的物理条件, 例如水生植物或者藻类的种群数量, 对生态系统产生长期复杂的影响(Brown & Moyle, 1991 ; Power, 1992 ; McDowall, 2006 ).鲤科鱼类的入侵范围现已遍布全球, 入侵初期可导致生态环境的物理变化, 如水生植物、浮游生物和小型脊椎动物的种群数量的明显减少, 继而水温、溶氧量、浑浊度等多种物理化学参数产生变化, 使底砾重新沉淀(Khan et al, 2003 ; Koehn, 2004 ; Miller & Crowl, 2006 ).再如食蚊鱼在美国犹他州的入侵造成了本地生态系统破坏, 同时被认为直接导致了产卵栖息地环境类似的本地种阴河鱼(Iotichthys phlegethontis )种群数量的下降(Ayala et al, 2007 ).栖息地环境的改变也促进了其他适生物种的入侵. ...
Species richness and patterns of invasion in plants, birds, and fishes in the United States.
1
2006
... 由于经济和社会需求, 淡水鱼类引种不可避免, 但一旦形成入侵, 评价过程复杂且治理花费大; 因此, 制定全球化的管理和预防机制非常重要(Gozlan et al, 2008 , 2010 ).鱼类入侵的评价是复杂且主观的, 过去往往依赖评价人的主观评价, 很多评价体系缺少量化分析(Stohlgren et al, 2006 ).近年来随着多种量化分析工具的开发, 入侵预警和评价系统帮助生态管理者对外来种和入侵种进行了客观有效的评价和监管(Copp et al, 2005a , b , 2009).具体的研究方法如Kolar和Lodge(2002)提出的分类回归树(classification and regression tree, CART), 该评价体系综合考虑了外来种的生物生态特征, 对五大湖的鱼类按建群与否、扩散速度和是否已成为有害种3个阶段进行分析, 通过外来种不同阶段不同特征的判别式系统预测外来鱼类可能形成的危害; 另一种鱼类入侵评价工具FISK (The Fish Invasiveness Screening Kit)和FISK v2也已被世界各国和地区广泛应用(Copp, 2013 ; Copp et al, 2005b , 2009 ; Onikura et al, 2011 ; Almeida et al, 2013 ; Lawson et al, 2013 ; Vilizzi & Copp, 2013 ), 该工具能较为客观地评价和预测鱼类入侵活动, 结合气候环境变化, 通过对物种分布、本土化过程、捕食、繁殖及扩散能力分析得出鱼类入侵的临界值, 该值可作为判别鱼类入侵与否的重要标准(Copp et al, 2005a , b ).由于不同国家和地区的气候、环境不同, FISK需要在各地区经校准后应用, 因而所得临界值不同(Copp et al, 2005b ; Almeida et al, 2013 ; Puntila et al, 2013 ; Tarkan et al, 2014 ). ...
Risk screening of non-native freshwater fishes at the frontier between Asia and Europe: first application in Turkey of the fish invasiveness screening kit.
2
2014
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
Application of FISK, an invasiveness screening tool for non-native freshwater fishes, in the Murray-Darling Basin (southeastern Australia).
1
2013
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
2
2009
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
... 鱼类的成功入侵与入侵地的生物多样性和生态位空缺有密切关系.鱼类的成功入侵往往发生在生物多样性低、生态结构不稳固的地区, 如欧洲波罗的海地区和北美五大湖地区相对年轻且物种本地化程度低, 入侵鱼类易成功入侵(Reid & Orlova, 2002 ).成功入侵的鱼类往往填补了入侵地空缺的生态位(Pettitt-Wade & Heath, 2015 ).入侵种和本地种虽可能在生态位的多个维度(dimensions of niche)发生重合, 但Hill等(2015)通过同位素的分析得知入侵物种至少填补了入侵地某一维度的空缺生态位. ...
2
2009
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...
... 鱼类的成功入侵与入侵地的生物多样性和生态位空缺有密切关系.鱼类的成功入侵往往发生在生物多样性低、生态结构不稳固的地区, 如欧洲波罗的海地区和北美五大湖地区相对年轻且物种本地化程度低, 入侵鱼类易成功入侵(Reid & Orlova, 2002 ).成功入侵的鱼类往往填补了入侵地空缺的生态位(Pettitt-Wade & Heath, 2015 ).入侵种和本地种虽可能在生态位的多个维度(dimensions of niche)发生重合, 但Hill等(2015)通过同位素的分析得知入侵物种至少填补了入侵地某一维度的空缺生态位. ...
Fish and ships: relating dispersal frequency to success in biological invasions.
2000
Non-native freshwater fish species in China.
1
2015
... 我国从国外引进的经济鱼类主要来源于南美洲(156种, 35.54%)、亚洲(101种, 23.01%)、非洲(94种, 21.41%)和北美洲(52种, 11.85%)(Xiong et al, 2015 ).其中, 从非洲、越南、泰国等地区引进了多种罗非鱼, 造成了其在中国南方水系的入侵, 如齐式罗非鱼(T. zillii )、尼罗罗非鱼、加利亚罗非鱼(Sarotherodon galilaeus )等; 虹鳟于1959年由朝鲜引进到中国北方, 现已在中国北方低温淡水水域形成自然种群; 美国红鱼(Sciaenops ocellata )原分布于美国东南海岸和墨西哥湾沿岸, 目前已被引入到福建等多个地区养殖, 形成入侵(阮少江, 2002 ).除了从国外引进经济鱼类, 国内不同水系之间的引种也不断得到推广, 如现已遍布我国各地的四大家鱼青鱼(Mylopharyngodon piceus )、草鱼、鲢、鳙, 20世纪90年代在全国推广的银鱼科鱼类、银鲫(Carassius gibelio )、中华倒刺鲃(Spinibarbus sinesis )等(楼允东, 2000 ). ...
Threats of invasive species for China caused by expanding international trade.
1
2012
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
An inventory of invasive alien species in China.
2
2012
... 生物入侵作为影响全球生态系统结构和功能的重要因素, 与气候变化和生物栖息地丧失并列为全球环境面临的三大问题(Sala et al, 2000 ; Rahel & Olden, 2008 ; Gozlan et al, 2010 ).随着全球经济一体化进程的深化, 水产养殖、观赏渔业、生态治理等原因导致的跨国家或者跨水系的引种愈加频繁, 其中部分外来种逃逸至野外并形成自然种群, 导致全球范围内的淡水鱼类入侵问题日益严重.鱼类入侵会破坏入侵地的生态环境, 并造成入侵水域生态类型单一及本地物种濒危等负面影响(Gurevitch & Padilla, 2004 ; Dextrase & Mandrak, 2006 ; Didham et al, 2007 ; Rahel & Olden, 2008 ).但也有研究表明鱼类入侵带来的生态危害被过分放大, 入侵种与本地种的濒危并没有直接联系, 且鱼类引种能带来正面的社会经济影响, 相关产业提供了淡水渔产品及工作机会, 因此应该客观对入侵物种的风险进行评估(Copp et al, 2005b , 2009 ; Didham et al, 2005 ; Sagoff, 2005 ; Gozlan et al, 2008 ; Jeschke & Strayer, 2008 ).针锋相对的学术争论还会持续, 但不争的事实是: 近20年鱼类入侵问题已被全球广泛关注, 相关研究论文数量翻了8倍(图1 ).此外, 早期鱼类入侵研究主要集中在入侵鱼类生物学特性及其带来的生态影响上, 近年来针对鱼类入侵风险评估的研究显著增加, 可见全球已经加强了对外来鱼类入侵的防范和管理(图2 ). ...
... 我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 ).Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 ).与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束. ...
The status and causes of alien species invasion in China.
2
2006
... 我国环境保护部门分别于2001-2003年, 2008-2010年两次对入侵物种进行全国范围的普查, 发现全国入侵物种共529种, 其中鱼类31种(万方浩, 2009 ; 丁晖等, 2011 ).Xiong等(2015)通过文献统计我国共有淡水外来鱼类439种, 该统计数据远高于全国普查数据及其他统计数据(李振宇和解焱, 2002 ; Ma et al, 2003 ; Xu H et al, 2012 ; Xue HG et al, 2006 , 2012 ).与其他国家相比, 我国对外来鱼类的研究力度仍然不够, 相关论文数量仅占全球总数的3.7%, 主要研究方向集中在少数已产生大规模危害的入侵种的地理分布(41.2%)及生物学特性(44.1%)上(该数据远高于全球均值24.2%和27.9%), 而缺乏对入侵种的生态影响(8.8%)及风险评估(0)的研究(该数据远低于全球均值26.1%和9.5%) (图2 ), 且未见具体的法律法规对外来鱼类的引进和管理进行约束. ...
... 人类活动不仅造成了大陆间的鱼类入侵, 也造成了大陆内不同水系间的鱼类入侵.(1)欧亚大陆内的入侵.20世纪中叶以后, 中国的草鱼(Ctenoph- aryngodon idella )、鲢(Hypophthalmichthys molitrix )、鳙(H. nobilis )等鲤科鱼类被引进到欧洲罗马尼亚、奥地利等国家, 造成了欧洲鲫鱼(Carassius auratus )种群数的下降(Scott, 1977 ; Mikschi, 2002 ; Navodaru et al, 2002 ).麦穗鱼(Pseudorasbora parva )于20世纪60年代随草鱼、鲢等鲤科鱼类的引进被携带至罗马尼亚(Banarescu, 1964 ), 随后迅速扩散到欧洲各国, 如奥地利、意大利等国家(Bianco, 1988 ).除了鲤科鱼类之外, 葛氏鲈塘鳢(Perccottus glenii )、虾虎鱼等其他科鱼类也在近30年从亚洲国家入侵到了欧洲各国(Bogutskaya & Naseka, 2002 ).(2)欧洲各国间的入侵.来源于东欧或中欧的欧洲鲶鱼(Silurus glanis )、云斑原吻虾虎、黑口新虾虎、河棲新虾虎(Neogobius fluviatilis )、戈拉高加索虾虎(Ponticola gorlap )、裸喉虾虎(Babka gymnotrachelus )和白梭吻鲈(Sander lucioperca ), 现已入侵到法国、意大利, 甚至英格兰、威尔士(Bianco, 1998 ; Copp et al, 2003 ; Maitland, 2004 ).其中虾虎鱼由于极强的表型适应性(Kováč & Siryova, 2005 )、繁殖的优越性(Grabowska, 2005 )以及对寄生生物的高抗性(Ondračková et al, 2005 ), 广泛入侵到欧洲各国, 人类对河道的改造也促使其成为欧洲入侵最为猖獗的鱼种之一(Naseka et al, 2005 ).(3)亚洲各国间的入侵.中国从亚洲其他国家引进水产养殖品种, 如多种罗非鱼等, 也造成了全国范围内(尤其是南方水系)外来鱼类的入侵(楼允东, 2000 ). ...
对我国渔业资源增值放流问题的思考
1
2008
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
对我国渔业资源增值放流问题的思考
1
2008
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
Difference in the predation impact enhanced by morphological divergence between introduced fish populations.
1
2007
... 入侵鱼类一般对本地生态系统和本地种产生以下生态影响: 捕食(Brown & Moyle, 1991 ; McIntosh & Townsend, 1995 ; Kitchell et al, 1997 ; Yonekura et al, 2007 )、种间竞争(Gurevitch et al, 1992 ; Simon et al, 2004 ; Zimmerman & Vondracek, 2006 )、杂交(Scribner et al, 2000 ; Costedoat et al, 2005 ; D’Amato et al, 2007 )、栖息地破坏(Brown & Moyle, 1991 , 1997 ; Kitchell et al, 1997 ; McDowall, 2006 )和疾病传播(Gaughan, 2001 ; Gozlan et al, 2005 , 2006 )等. ...
河北省海洋渔业增值放流现状、问题及建议
1
2012
... (3)入侵种与本地种的杂交.入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 ).比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 ).也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 ). ...
河北省海洋渔业增值放流现状、问题及建议
1
2012
... (3)入侵种与本地种的杂交.入侵种与本地种的杂交作为环境压力的一种, 导致了鱼类种群和遗传多样性的下降(Allendorf et al, 2004 ; D’Amato et al, 2007 ).比如欧洲鲫鱼与鲫和鲤(Carpio carpio )杂交导致了欧洲鲫鱼在本地的种群数量下降(Smartt, 2007 ).也有研究表明杂交并不一定总是降低物种多样性, 比如入侵法国的一种大鼻软口鱼(Chondrostoma nasus )和法国本地弓口副软口鱼(C. toxostoma )经过几十年的杂交已经形成了稳定的种群, 同时也在同一水系保持了原有种群数量(Costedoat et al, 2005 ). ...
Interactions of slimy sculpin (Cottus cognatus) with native and nonnative trout: consequences for growth.
1
2006
... 成功入侵的鱼类有很强的环境适应力(ArbaČIauskas et al, 2013 )、表型适应性(Reid & Orlova, 2002 ; Giery et al, 2015 )、繁殖力(Grabowska, 2005 )和竞争力(MacNeil & Prenter, 2000 ).(1)环境适应力.引入地野外环境不一定是入侵种的最优生境, 但成功入侵的鱼种往往能迅速找到适合生存的生境.如食蚊鱼表现出了很宽的盐幅和温幅, 研究表明霍氏食蚊鱼(Gambusia holbrooki )可以在40℃的生境生存, 也可以在0℃的生境中越冬, 还可以在严重污染的水体及低溶氧的环境中生存(Rees, 1945 ; Staub et al, 2004 ).(2)表型适应性.入侵物种和本地物种在应对相同的环境压力时也可表现出不同的表型适应性, 如入侵雄性食蚊鱼可在同一环境压力下表现出不同颜色和花纹的变化, 从而在交配中对异性发出更有效的信号(Giery et al, 2015 ).(3)繁殖力.入侵鱼类有很强的繁殖力, 表现为产卵量高、孵化成功率高、性成熟早、繁殖周期短等方面(Grabowska, 2005 ).(4)竞争力.ArbaČIauskas等(2013)通过同位素分析发现化学适应力与鱼类捕食习性相关, 而化学适应性强的鱼类具备更强的竞争力, 入侵鱼类往往具备这种特点.入侵物种在入侵地的竞争中占优势, 本地种往往被迫改变习性和栖息地, 从而导致本地种数量急剧下降(Bergstrom & Mensinger, 2009 ). ...



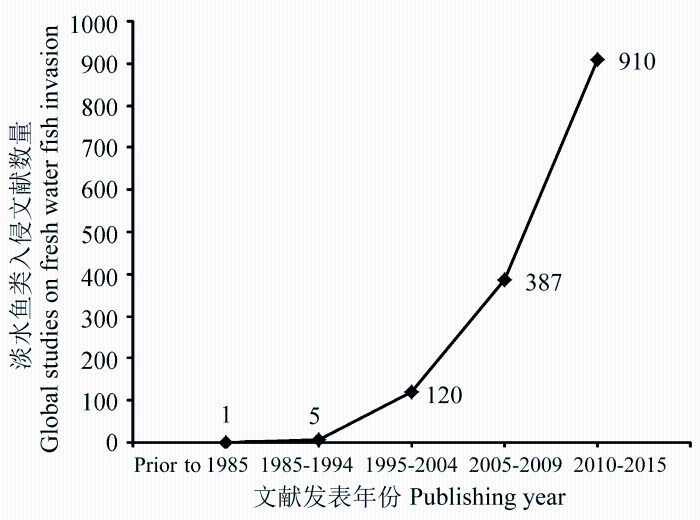
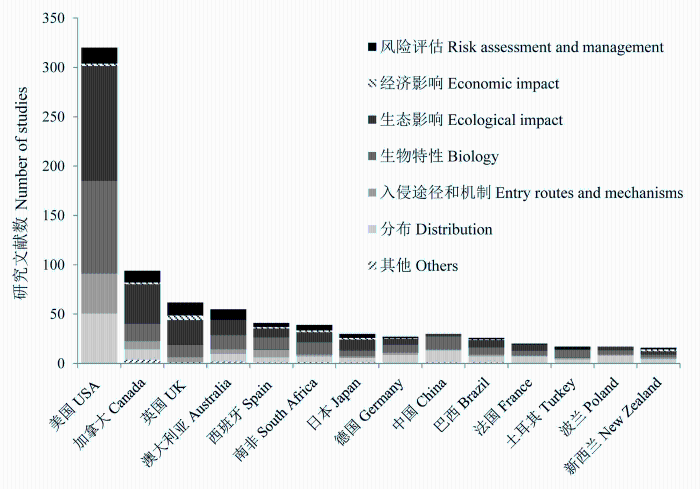
{kind=link}
{kind=link}
{kind=link}
{kind=link}