生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 )。生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能。例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 )。由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种。
物种丰富度的空间格局一直是生态学和生物地理学的研究热点。物种丰富度变化和周转速率在纬度和海拔梯度上往往较为显著, 因此备受关注(Lomolino, 2001 )。乡土物种丰富度一般随纬度和海拔单调递减或者呈单峰格局, 尤其是当低海拔人类干扰严重时(Stevens, 1992 ; Rahbek, 1995 ); 而入侵物种丰富度一般随海拔升高呈现单调递减的格局, 其纬度格局则与研究区域的位置和尺度有关。尽管对物种分布空间格局的研究已有很长的历史, 但对外来种随纬度和海拔梯度的入侵格局的研究仍有待深入。开展这方面研究显然有助于增加对物种丰富度决定机制的理解(Lembrechts et al, 2014 )。
植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素。生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 )。种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵。然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 )。
滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 )。包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 )。目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献。本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小。希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据。
1 材料与方法
1.1 研究区概况
怒江是中国西南地区的大河之一, 发源于青藏高原的唐古拉山南麓, 在云南省内流经怒江州的贡山、福贡、泸水和保山地区的保山、施甸、龙陵、昌宁等7县市, 出国境后称为萨尔温江。云南省内的怒江段位于98º07'-100º02' E, 24º07'-28º23' N, 长度650 km, 流域面积3.35万km2 , 占云南省面积的8.7%。云南怒江流域受西南季风影响强烈, 年均气温5.2-16.6℃, 年均降水量961.7-1,639.6 mm。研究区段河谷海拔从南到北递增, 气温相应降低, 而降水量呈递增趋势, 具有亚热带半干旱至半湿润气候; 河谷低地植被稀疏, 以云南松(Pinus yunnanensis )为优势种的针叶林或混交林和灌草丛间断分布(李晖等, 2009 )。
研究区内的公路主要是沿江呈南北分布的省道S228, 沿线有县、乡(镇)和村级居民点分布。公路两旁植被长期受到不同程度的人为干扰, 生物入侵现象十分普遍。
1.2 野外植物群落与生境调查
为了探究怒江地区外来植物的入侵现状及影响因素, 我们在公路沿线进行了系统的植物群落和生境调查及土壤采样。采取分层系统设计, 沿怒江河谷延伸的省道S228两侧布设调查样点, 参考各生境因子调整采样强度, 包括: (1)公路等级: 包括省道(138个样方)、县道(112个样方)和乡村道(24个样方); (2)道路上、下坡: 沿样线在公路两侧等距离布设样方各1-3个; (3)与道路距离: 样方到公路边距离依次为10 m、30 m、60 m, 以反映公路相关的生境干扰衰减的梯度; (4)土地覆盖类型: 避开建筑用地和生产用地。考虑到怒江河谷植被以灌草丛为主, 样方统一为10 m × 10 m, 以保证足够的取样面积。
群落调查线路上至滇藏交界的云南怒江州贡山县丙中洛, 下至保山市潞江县境内, 全长500余km, 共调查273个样方(图1 )。用GPS测定每个样方的地理坐标和海拔。记录样方内所有维管束植物(包括外来物种); 测量所有胸径≥ 2 cm的乔木高度、胸径和个体数。利用罗盘仪测定样方的坡向、坡度, 记录坡位。生境干扰因子由3个变量表示, 即地表裸露比例、公路等级、样方到公路的距离。
图1 怒江地区公路沿线植物调查样方分布图
Fig. 1 Distribution map of plant investigation plots alongside road in Nujiang River region
1.3 样方气候数据
由于缺乏每个样方所在地气候状况的实际观测数据, 本研究采用Worldclim全球气候数据集(http://www.worldclim.org/)提供的月均气温和降水数据。该数据是全球气象台站50年(1950-2000年) 观测数据平均值经空间插值得到的栅格数据, 空间分辨率为1 km。利用样方地点的地理坐标, 从美国航空航天局(NASA)发布的全球数字高程模型(Global Digital Elevation Model, GDEM, 100 m分辨率)中提取研究区内的数字地形数据, 并采用刘晔(2015) 提供的方法, 结合三江并流地区气象台站过去54年观测记录数据的平均值, 采用Co-Kriging插值修正, 得到每一样方点的12个月平均气温和降水数据。
进一步参考相关文献计算多个生物气候指标(Fang et al, 2002 ; Hawkins et al, 2003 ), 分为能量、水分和气候变异性3组。能量因子指标包括年均温(MAT)、最冷季均温(MTCQ)、最暖季均温(MTWQ)和年潜在蒸散量(PET) (Thornthwaite & Hare, 1955 )。其中, 年潜在蒸散量表示在理想状态下, 一年内地表(植被、土壤和水体表面)向大气蒸腾水分的最大可能值, 反映了一个地区的环境能量, 其计算公式如下:
其中k i 为第i个月的标准月换算因子, 可以通过换算因子表查得。e i 为第i个月的标准月份最大蒸散量, e i = 16 × (10MMT i /I)a /(S/360), MMTi 为月均温, I为12个月总和的热量指标(o C), 计算公式为I = ∑(MMTi /5)1.514 , a = 6.75 × 10-7 I3 - 7.71 × 10-5 I2 + 1.79 × 10-2 I + 0.4923, S是月实际日照时数。上述公式仅适用于月均温在0-26.5ºC之间。当MMT i ≤ 0℃时, PET设定为0, 当MMTi ≥ 26.5℃时, 由月均温直接查得e并计算PET。
水分因子包括年降水量(MAP)、最干季降水量(PDQ)、最湿季降水量(PWQ)、年实际蒸散量(AET) (Brutsaert, 1982 )和水分亏缺(water deficit, WD) (Stephenson, 1990 )。其中, 最干(或湿)季降水量是一年内降水最少(或最多)的3个月内的降水总量, 即一个地区冬季(或夏季)的降水量; 年实际蒸散量是一年内地表向大气蒸腾水分的实际值, 受水分和能量双重影响; 水分亏缺反映一个地方的干燥程度。其计算公式如下:
AET = MAP/[0.9 + (MAP/L)2 ]1/2 (2)
WD = AET - PET (3)
其中, L为平均蒸发量, L = 300 + 25MAT + 0.05MAT3 。
气候变异性因子包括温度季节性(temperature seasonality, TSN)和降水季节性(precipitation seasonality, PSN) (Fang & Lechowicz, 2006 )。温度和降水季节性反映一个地区温度和降水的季节变化程度, 其计算公式如下:
其中, MMP i
1.4 数据分析
在分析乡土物种或入侵物种丰富度与纬度、海拔的关系时, 在模型中加入纬度和海拔的一次和二次项, 如果二次项结果显著(P < 0.05), 则认为二者的关系非线性。
由于物种丰富度通常符合Poisson分布, 采用广义线性模型(generalized linear model, GLM)拟合入侵物种与乡土物种丰富度与环境因子的关系。当出现残差过度分散, 即残差/剩余自由度> 1.2时, 采用quasi-Poisson分布。由于部分气候因子之间存在严重的共线性, 因此先计算自变量之间的相关系数, 在气温、降水、气候变异性3组变量中各选1个组内相关性高而与组间变量相关性低的变量, 加入物种丰富度的多元广义线性模型, 根据赤池信息准则(Akaike Information Criterion, AIC)选出最优模型。基于GLM最优模型, 进一步采用等级方差分离(hierarchical variation partitioning, HVP)算法来定量评价不同自变量对丰富度变异性的独立贡献(Nally, 2002 )。该方法通过穷尽比较包含1个到考虑的全部自变量的所有变量组合构建的模型, 分离出每个自变量的独立效应和联合效应, 并用Z-scores值检验自变量的独立效应是否统计显著。
为了获取环境因子约束下入侵物种和乡土物种丰富度的关系, 采用结构方程模型(structural equation model, SEM)进行分析, SEM能够同时分析影响入侵和乡土物种丰富度的因素之间的相互作用。数据采用Z-scores进行标准化处理, 使得不同变量间的过程系数可以比较。首先利用主成分分析(PCA)对所有地形、气候环境和人类干扰等因子做降维处理, 将这些存在高度相关性的变量转化为少数线性不相关的综合变量。我们根据PCA的特征值提取了贡献较大的前4个主成分。然后将获取的主成分与入侵物种和乡土物种丰富度一起纳入结构方程模型进行分析。
样方气候指标推算通过地理信息系统软件Arcgis 9.3完成。统计分析全部在统计软件R 3.2.0 (http://www.r-project.org/)中完成。
2 结果
2.1 入侵植物和乡土植物的组成和分布
本次调查的273个样方中共出现26种外来植物, 属于13科21属(附录1), 其中菊科植物最多, 占外来植物种的38.5%, 其次是茄科和大戟科。不同种的频度差异很大, 如番石榴(Psidium guajava )、曼陀罗(Datura stramonium )、婆婆纳(Veronica didyma )等均仅见于1个样方, 而紫茎泽兰出现在106个样方中。样方入侵物种丰富度介于0-7, 密度为2.03 ± 1.45种/100 m2 。入侵植物原产地主要为美洲, 占76.9%, 其次来自欧洲、非洲热带和亚洲热带。26种外来入侵植物中草本17种, 占65.4%。入侵程度最严重的是鬼针草(Bidens spilosa ), 出现的样方数和平均重要值分别为143和5.94, 其次是小白酒草(Conyza canadensis )、紫茎泽兰、苏门白酒草(C. sumatrensis )和藿香蓟(Ageratum conyzoides )。紫茎泽兰的平均重要值最高(21.44), 构成比较普遍的优势群落。
样方中共记录1,145种乡土植物, 分属于158科628属, 其中禾本科、豆科和菊科植物最为丰富。样方中乡土植物丰富度介于6-91种, 平均密度为35.8 ± 16.3种/100 m2 。
在怒江流域, 入侵植物分布的纬度范围为25.068°-28.069° N, 最北出现的种类包括鬼针草、小白酒草、苏门白酒草和蓖麻(Risinus communi )。入侵种丰富度呈现随纬度增加而减少的趋势(P = 0.009)。入侵植物的海拔分布范围为726-3,135 m, 分布海拔最高的物种为苦苣菜(Sonchus oleraceus )。入侵物种丰富度随海拔升高而线性减少的趋势非常显著(海拔, P < 0.001; 海拔的二次项, P = 0.189)。乡土植物丰富度则表现出明显不同的格局, 丰富度随纬度增加显著上升(P < 0.001), 随海拔上升表现出先增加后减少的单峰型格局(海拔, P < 0.001; 海拔的二次项, P < 0.001), 在中等海拔(2,000 m)处丰富度达到最高值(图2 )。
图2 怒江河谷入侵(A)与乡土植物(B)丰富度随纬度和海拔的分布
Fig. 2 Distribution of invasive (A) and native (B) species richness along latitude and altitude in Nujiang River valley
2.2 入侵植物和乡土植物丰富度与环境因子的 关系
关于入侵物种丰富度的GLM最优模型包括4个因子: 公路等级、年均温、最湿季降水量和最干季降水量, 共解释了入侵物种丰富度变异的9.8%。其中公路等级、最干季降水量和年降水量分别贡献了全部解释量的54.1%、22.0%和16.71% (图3 )。
乡土物种丰富度的GLM最优模型包括5个因子: 有无乔木(0/1)、坡向、坡度、裸地比例和最干季降水量, 共解释乡土物种丰富度的32.1%。其中, 裸地比例贡献最大(58.0%), 坡向(21.5%)和有无乔木(17.3%)是另外两个影响显著的因子。坡度和气候因子影响微弱(图3 )。
图3 基于等级方差分离的怒江河谷入侵(a)和乡土植物(b)物种丰富度格局的环境因子独立和联合解释能力。*表示在0.05水平显著。
Fig. 3 Independent and joint contributions of environmental variables accounting for the spatial pattern of invasive and native species richness in Nujiang River valley, based on hierarchical variation partitioning model. * indicates that influence is significant at the 0.05 level. RG, Road grade; MAP, Mean annual precipitation; PDQ, Precipitation of driest quarter; PWQ, Precipitation of wettest quarter.
2.3 入侵与乡土植物物种丰富度的关系
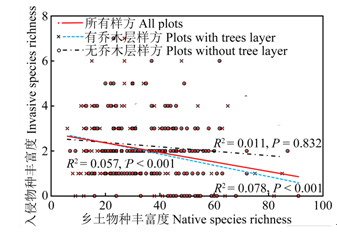
本研究表明, 在群落尺度上外来植物与乡土植物的多样性之间存在显著负相关关系(P < 0.001) (图4 ), 即乡土物种的丰富度越高, 入侵物种的丰富度越低, 反映了群落复杂性对入侵植物的抵抗性。将样方分为有、无乔木层两类后, 在有乔木层的样方中, 入侵物种与乡土物种丰富度之间的负相关关系更为显著(P < 0.001), 而无乔木层的样方中二者无明显关系(P = 0.832), 表明在不同生境中驱动乡土物种和外来物种丰富度格局的机制可能不同。有乔木层存在的情况下, 入侵物种丰富度显著低于只有灌草的样方(F = 4.581, P = 0.033), 而乡土物种丰富度显著高于只有灌草的样方(F = 20.736, P = 0.000)。二者在干扰程度不同的生境中复杂的相互作用可能是不同的生物和非生物因子共同作用的结果。无乔木层的样方中没有相关性, 说明由于优势乡土竞争种的缺乏, 导致可利用资源较多, 在这些生境中可能没有强烈的种间相互作用。
图4 入侵与乡土植物物种丰富度的关系
Fig. 4 Relationship between richness of invasive and native species
根据各主成分的特征值, 我们选取了前4个主成分代表人类干扰、地形和水热因子变量(表1 )。前4个主成分累计代表了80.5%的因子信息量, 第一主成分贡献率为48.6%, 且大部分因子在这一主成分上的载荷为负值。第二主成分贡献率为15.6%, 与成分载荷较大的因子呈正相关关系。其中, 第一主成分主要表征热量因子, 第二主成分则表征了水分因子, 第三主成分主要代表降水季节性的信息, 而第四主成分则代表了人为干扰和地形因子。
结构方程模型结果显示(图5 ), 外来入侵物种丰富度只与第一主成分表现出极显著的负相关关系, 与第二主成分负相关但并不显著, 与第三和第四主成分正相关但并不显著。乡土物种丰富度与第一主成分为极显著正相关关系, 与第二主成分之间为显著的正相关关系, 与第三主成分正相关但并不显著, 而与第四主成分为显著负相关关系。在控制了丰富度与环境因子的关系后, 入侵种丰富度和乡土种丰富度之间没有表现出显著的相关性。
图5 环境因子主成分(PCA1, PCA2, PCA3, PCA4)和乡土种丰富度(nat_ric)对入侵种丰富度(inv_ric)影响的结构方程模型。箭头方向表示因果关系, 数字表示路径相关系数, 方框旁的数字表示模型对变量变异的解释系数, 星号表示相关的显著性水平: *P < 0.05, **P < 0.01, ***P < 0.001。
Fig. 5 A structural equation model showing the multivariate effects on invasive plant richness (inv_ric) by native plant richness (nat_ric) and four principal components (PCA1, PCA2, PCA3, PCA4) of environmental factors. The direction of arrows shows the causal relationships, numbers are standardized path coefficients, accompanied by positive (+) or negative (-) illustrations. The proportion of variation explained by the model is indicated by the number near each textbox. Asterisks imply the level of significance: *P < 0.05, **P < 0.01, ***P < 0.001.
3 讨论
3.1 入侵和乡土物种丰富度的空间格局
研究区内入侵物种与乡土物种丰富度沿海拔梯度存在显著不同的空间分布格局, 这个差异可能反映了入侵植物不同于乡土植物的生活史性状与环境需求(van Kleunen et al, 2010 )。外来入侵物种丰富度随海拔上升而下降的格局与其他研究相似, 但这种格局的成因目前还有很大争议, 可能是由于逐渐严酷的非生物环境因子限制了外来种进一步扩散, 或是人为影响减少导致繁殖体扩散能力降低, 也可能仅仅是由于入侵时间较短, 还没有随海拔上升完全占据可利用的生境(Daehler, 2005 ; Giorgis et al, 2011 ; Marini et al, 2012 )。
3.2 环境因素对入侵和乡土物种丰富度格局的 影响
探讨入侵和乡土植物的分布格局及形成机制是保护生物学研究的重要组成部分。关于环境因子对外来物种分布及其多样性格局的影响存在不同假说。Elton (1958)认为乡土物种丰富度高的地方对入侵的抵抗性强, 但支持这一观点的证据模棱两可(Kennedy et al, 2002 ; Fargione et al, 2003 )。干扰被认为是入侵的先导, 但很多未被干扰的群落也存在外来物种入侵(Lozon & MacIsaac, 1997 ; Davis et al, 2000 )。乡土物种在长期历史过程中已经适应的环境压力可能限制了外来物种的存活和定殖, 但资源可利用性低的栖息地依然受到入侵物种的影响(Harrison, 1999 )。
怒江河谷地区以干热气候为主。能量对入侵和乡土物种丰富度的影响均不显著; 而水分条件, 尤其是最干季降水量的影响非常显著, 显示了干旱对植物分布的限制作用, 且入侵物种对干旱的耐性比乡土物种更低。Baruch和Fernandez (1993)发现一种乡土C4 草本植物对于干旱的耐受性高于另一种入侵C4 草本植物; Hamilton等(1999) 也发现, 美国加州南部草地的干旱环境有利于多年生乡土草本物种的萌芽而不利于一年生入侵草本。怒江河谷的乡土植物在长期适应干旱环境的进化过程中变得更加耐旱, 而入侵种由于进化时间较短, 对干旱生境的适应性较差。对于乡土物种, 裸地比例即路侧生境干扰具有最强的胁迫; 而样方到公路的距离与入侵物种丰富度显著负相关, 这充分显示, 在怒江河谷, 公路沿线的生境干扰对乡土植物群落的退化和外来物种的迁移入侵具有关键作用。
3.3 群落物种多样性与可入侵性
物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 )。支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 )。且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 )。相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵。也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关。
本研究表明, 在群落尺度上外来入侵植物与乡土植物的多样性之间存在显著负相关关系(P < 0.001), 特别是乔木层存在的情况下, 入侵物种丰富度显著低于只有灌草的样方。这表明局域尺度群落结构完整性可能对生物入侵具有抵抗作用。乔木层是否存在对入侵的影响可能与光资源有关。乔木层下的生境光照强度低, 可能抑制了外来植物的竞争力(Watkins et al, 2003 , Hansen & Clevenger, 2005 ); 而光照较强的生境如草地、开放林地以及森林边缘等, 常伴随高温、裸露土壤等条件, 容易被外来植物入侵, 并促使外来植物定居后向更远的距离扩散(Parendes & Jones, 2000 ; Hansen & Clevenger, 2005)。
但是, 当我们进一步使用结构方程模型分析入侵和乡土植物对环境因子的响应发现, 入侵和乡土植物的丰富度格局对温度、降水、人类干扰和地形等环境因子的响应完全相反, 并且在控制了外部环境因子的影响后, 二者的关系并不显著。这表明二者丰富度的负相关关系更多是各自对环境约束的响应差异, 而非直接竞争排斥的结果。
本文从群落水平对怒江河谷乡土和入侵植物多样性进行研究, 发现二者的空间格局显著不同, 对环境因子的响应也基本相反。结果支持物种入侵的资源可利用性限制假说, 同时强调了人为干扰对生物多样性的负面影响; 乡土植物或已较好地适应了干旱河谷气候, 但并没有显示出对外来物种入侵的抵抗作用。
参考文献
文献选项
[1]
Baker HG 1974 ) The evolution of weeds
. Annual Review of Ecology and Systematics , 5 , 1 -24 .
[本文引用: 1]
[2]
Baruch Z Fernandez DS 1993 ) Water relations of native and introduced C4 grasses in a neotropical savanna
. Oecologia , 96 , 179 -185 .
[3]
Brooks ML D’Antonio CM Richardson DM Grace JB Keeley JE DiTomaso JM Hobbs RJ Pellant M Pyke D 2004 ) Effects of invasive alien plants on fire regimes
. BioScience , 54 , 677 -688 .
[本文引用: 1]
[4]
Brutsaert WH 1982 ) Evaporation in the atmosphere : theory, history and application. D. Reidel Publishing Company, Dordrecht .
[本文引用: 1]
[5]
Bulleri F Bruno JF Benedetti-Cecchi L 2008 ) Beyond competition: incorporating positive interactions between species to predict ecosystem invasibility
. PLoS Biology , 6 , 1136 -1140 .
[本文引用: 1]
[6]
Christian CE 2001 ) Consequences of a biological invasion reveal the importance of mutualism for plant communities
. Nature , 413 , 635 -639 .
[本文引用: 1]
[7]
Daehler CC 2005 ) Upper-montane plant invasions in the Hawaiian Islands: patterns and opportunities. Perspectives in Plant Ecology,
Evolution and Systematics , 7 , 203 -216 .
[本文引用: 1]
[8]
Davis MA Grime JP Thompson K 2000 ) Fluctuating resources in plant communities: a general theory of invasibility
. Journal of Ecology , 88 , 528 -534 .
[本文引用: 1]
[9]
Deutschewitz K Lausch A Kühn I Klotz S 2003 ) Native and alien plant species in relation to spatial heterogeneity on a regional scale in Germany
. Global Ecology and Biogeography , 12 , 299 -311 .
[本文引用: 1]
[10]
Ehrenfeld JG 2003 ) Effects of exotic plant invasions on soil nutrient cycling processes
. Ecosystems , 6 , 503 -523 .
[本文引用: 1]
[11]
Ehrenfeld JG 2010 ) Ecosystem consequences of biological invasions. Annual Review of Ecology, Evolution,
and Systematics , 41 , 59 -80 .
[本文引用: 1]
[12]
Elton CS 1958 ) The Ecology of Invasions by Animals and Plants . University of Chicago Press, Chicago .
[本文引用: 1]
[13]
Fang JY Lechowicz MJ 2006 ) Climatic limits for the present distribution of beech (Fagus L.) species in the world
. Journal of Biogeography , 33 , 1804 -1819 .
[本文引用: 1]
[14]
Fang JY Song YC Liu HY Piao SL 2002 ) Vegetation-climate relationship and its application in the division of vegetation zone in China
. Acta Botanica Sinica , 44 , 1105 -1122 .
[本文引用: 1]
[15]
Fargione J Brown CS Tilman D 2003 ) Community assembly and invasion: an experimental test of neutral versus niche processes. Proceedings of the National Academy of Sciences,
USA , 100 , 8916 -8920 .
[本文引用: 1]
[16]
Funk J Zachary V 2010 ) Physiological responses to short-term water and light stress in native and invasive plant species in southern California
. Biological Invasions , 12 , 1685 -1694 .
[本文引用: 1]
[17]
Gerhardt F Collinge SK 2007 ) Abiotic constraints eclipse biotic resistance in determining invasibility along experimental vernal pool gradients
. Ecological Applications , 17 , 922 -933 .
[本文引用: 1]
[18]
Giorgis MA Tecco PA Cingolani AM Renison D Marcora P Paiaro V 2011 ) Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina
. Biological Invasions , 13 , 1423 -1434 .
[本文引用: 1]
[19]
Grotkopp E Rejmánek M Rost TL 2002 ) Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species
. The American Naturalist , 159 , 396 -419 .
[本文引用: 1]
[20]
Halpern SL Underwood N 2006 ) Approaches for testing herbivore effects on plant population dynamics
. Journal of Applied Ecology , 43 , 922 -929 .
[本文引用: 1]
[21]
Hamilton JG Holzapfel C Mahall BE 1999 ) Coexistence and interference between a native perennial grass and non-native annual grasses in California
. Oecologia , 121 , 518 -526 .
[本文引用: 1]
[22]
Hansen MJ Clevenger AP 2005 ) The influence of disturbance and habitat on the presence of non-native plant species along transport corridors
. Biological Conservation , 125 , 249 -259 .
[本文引用: 1]
[23]
Harrison S 1999 ) Native and alien species diversity at the local and regional scales in a grazed California grassland
. Oecologia , 121 , 99 -106 .
[本文引用: 1]
[24]
Hawkins BA Field R Cornell HV Currie DJ Guégan JF Kaufman DM Kerr MJ Mittelbach GG Oberdorff T O’Brien EM Porter RE Turner JRG 2003 ) Energy, water, and broad-scale geographic patterns of species richness
. Ecology , 84 , 3105 -3117 .
[本文引用: 1]
[25]
Hu FG Duan CF Liu GH 2007 ) Exotic invasive weeds in farmland of dry-hot valley in Yunnan Nujiang region
. Weed Sciences , (4 ), 20 -23 .(in Chinese)
[本文引用: 1]
[胡发广 , 段春芳 , 刘光华 (2007 ) 云南怒江干热河谷农田外来入侵杂草的调查
. 杂草科学 , (4 ), 20 -23 .]
[本文引用: 1]
[26]
Hulme PE Pyšek P Jarošík V Pergl J Schaffner U Vila M 2013 ) Bias and error in understanding plant invasion impacts
. Trends in Ecology & Evolution , 28 , 212 -218 .
[本文引用: 1]
[27]
Kennedy TA Naeem S Howe KM Knops JMH Tilman D Reich P 2002 ) Biodiversity as a barrier to ecological invasion
. Nature , 417 , 636 -638 .
[本文引用: 1]
[28]
Kissling WD Lord JM Schnittler M 2006 ) Agamospermous seed production of the invasive tussock grass Nardus stricta L. (Poaceae) in New Zealand: evidence from pollination experiments
. Flora , 201 , 144 -151 .
[本文引用: 1]
[29]
Kueffer C P Pyšek Richardson DM 2013 ) Integrative invasion science: model systems, multi-site studies, focused meta-analysis and invasion syndromes
. New Phytologist , 200 , 615 -633 .
[本文引用: 1]
[30]
Lembrechts JJ Milbau A Nijs I 2014 ) Alien roadside species more easily invade alpine than lowland plant communities in a subarctic mountain ecosystem
. PloS ONE , 9 , e89664 .
[本文引用: 1]
[31]
Levine JM 2000 ) Species diversity and biological invasions: relating local process to community pattern
. Science , 288 , 852 -854 .
[本文引用: 1]
[32]
Levine JM D’Antonio CM 1999 ) Elton revisited: a review of evidence linking diversity and invasibility
. Oikos , 1999 , 15 -26 .
[本文引用: 1]
[33]
Li H Bai Y Yang SH Zhu X Zhao K 2009 ) Prediction of vegetations dynamic changes in central Nujiang watershed based on Markov process mode
. Chinese Journal of Ecology , 28 , 371 -376 .(in Chinese with English abstract)
[本文引用: 1]
[李晖 , 白杨 , 杨树华 , 朱雪 , 赵凯 (2009 ) 基于马尔柯夫模型的怒江流域中段植被动态变化预测
. 生态学杂志 , 28 , 371 -376 .]
[本文引用: 1]
[34]
Liu J Dong M Miao SL Li ZY Song MH Wang RQ 2006 ) Invasive alien plants in China: role of clonality and geographical origin
. Biological Invasions , 8 , 1461 -1470 .
[本文引用: 1]
[35]
Liu Y 2015 ) Plant diversity and biogeography of the arid valleys in Southwest China. PhD dissertation, Peking University, Beijing
.(in Chinese with English abstract)
[刘晔 (2015 ) 中国西南干旱河谷的植物多样性地理格局
. 博士学位论文, 北京大学, 北京.]
[36]
Liu Y Shen LF Wang T Fang R Jiang R Shen ZH 2013 ) Invasive pattern and influential factors of Eupatorium adenophorum along the highway in three parallel rivers region in northwestern Yunnan
. Highway Traffic Science and Technology , 5 , 284 -288 .(in Chinese with English abstract)
[本文引用: 1]
[刘晔 , 沈利峰 , 王韬 , 房锐 , 姜睿 , 沈泽昊 (2013 ) 滇西北三江并流地区公路沿线紫茎泽兰的入侵格局及影响因素
. 公路交通科技 , 5 , 284 -288 .]
[本文引用: 1]
[37]
Lomolino MV 2001 ) Elevation gradients of species-density: historical and prospective views
. Global Ecological Biogeogrophy , 10 , 3 -13 .
[本文引用: 1]
[38]
Lozon JD MacIssac HJ 1997 ) Biological invasions: are they dependent on disturbance?
Environmental Reviews , 5 , 131 -144 .
[39]
Lu P Sang WG Ma KP 2005 ) Progress and prospects in research of exotic invasive species, Eupatorium adenophorum
. Acta Phytoecologica Sinica , 29 , 1029 -1037 .(in Chinese with English abstract)
[本文引用: 1]
[鲁萍 , 桑卫国 , 马克平 (2005 ) 外来入侵种紫茎泽兰研究进展与展望
. 植物生态学报 , 29 , 1029 -1037 .]
[本文引用: 1]
[40]
Lu ZJ Ma KP 2004 ) The influence of topographical factors on the invasion of the alien species, Eupatorium adenophorum
. Acta Phytoecologica Sinica , 28 , 761 -767 .(in Chinese with English abstract)
[本文引用: 1]
[卢志军 , 马克平 (2004 ) 地形因素对外来入侵种紫茎泽兰的影响
. 植物生态学报 , 28 , 761 -767 .]
[本文引用: 1]
[41]
Lu ZJ Ma KP 2005 ) Scale dependent relationships between native plant diversity and the invasion of croftonweed (Eupatorium adenophorum) in Southwest China
. Weed Science , 53 , 600 -604 .
[本文引用: 1]
[42]
Nally RM 2002 ) Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables
. Biodiversity and Conservation , 11 , 1397 -1401 .
[本文引用: 1]
[43]
Marini L Battisti A Bona E Federici G Martini F Pautasso M Hulme PE 2012 ) Alien and native plant life-forms respond differently to human and climate pressures
. Global Ecology and Biogeography , 21 , 534 -544 .
[本文引用: 1]
[44]
Matzek V 2011 ) Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment
. Biological Invasions , 13 , 3005 -3014 .
[本文引用: 1]
[45]
Mooney HA Hobbs RJ 2000 ) Invasive Species in a Changing World . Island Press, Washington, DC .
[本文引用: 1]
[46]
Palmer MW Maurer TA 1997 ) Does diversity beget diversity? A case study of crops and weeds
. Journal of Vegetation Science , 8 , 235 -240 .
[本文引用: 1]
[47]
Parendes LA Jones JA 2000 ) Role of light availability and dispersal in exotic plant invasion along roads and streams in the H. J. Andrews experimental forest, Oregon
. Conservation Biology , 14 , 64 -75 .
[本文引用: 1]
[48]
Pimentel D Lach L Zuniga R Morrison D 2000 ) Environmental and economic costs of nonindigenous species in the United States
. BioScience , 50 , 53 -65 .
[本文引用: 1]
[49]
Pokorny ML Sheley RL Zabinski CA Engel RE Svejcar TJ Borkowski JJ 2005 ) Plant functional group diversity as a mechanism for invasion resistance
. Restoration Ecology , 13 , 448 -459 .
[本文引用: 1]
[50]
Rahbek C 1995 ) The elevational gradient of species richness: a uniform pattern?
Ecography , 18 , 200 -205 .
[本文引用: 1]
[51]
Rudgers JA Mattingly WB Koslow JM 2005 ) Mutualistic fungus promotes plant invasion into diverse communities
. Oecologia , 144 , 463 -471 .
[本文引用: 1]
[52]
Souza L Bunn WA Simberloff D Lawton RM Sanders NJ 2011 ) Biotic and abiotic influences on native and exotic richness relationship across spatial scales: favourable environments for native species are highly invasible
. Functional Ecology , 25 , 1106 -1112 .
[本文引用: 1]
[53]
Spence LA Dickie IA Coomes DA 2011 ) Arbuscular mycorrhizal inoculum potential: a mechanism promoting positive diversity-invasibility relationships in mountain beech forests in New Zealand?
Mycorrhiza , 21 , 309 -314 .
[本文引用: 1]
[54]
Stephenson NJ 1990 ) Climatic control of vegetation distribution: the role of the water balance
. The American Naturalist , 135 , 649 -670 .
[本文引用: 1]
[55]
Stevens GC 1992 ) The elevational gradient in altitudinal range: an extension of Rapoport’s latitudinal rule to altitude
. The American Naturalist , 140 , 893 -911 .
[本文引用: 1]
[56]
Thornthwaite CW Hare FK 1955 ) Climatic classification in forestry
. Unasylva , 9 , 51 -59 .
[本文引用: 1]
[57]
Toledo M Poorter L Pena-Claros M Alarcon A Balcazar J Leano C Licona JC Llanque O Vroomans V Zuidema P Bongers F 2011 ) Climate is a stronger driver of tree and forest growth rates than soil and disturbance
. Journal of Ecology , 99 , 254 -264 .
[本文引用: 1]
[58]
van Kleunen MV Weber E Fischer M 2010 ) A meta-analysis of trait differences between invasive and non-invasive plant species
. Ecology Letters , 13 , 235 -245 .
[本文引用: 2]
[59]
Vilà M Espinar JL Hejda M Hulme PE Jarošík V Maron JL Pergl J Schaffner U Sun Y Pyšek P 2011 ) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems
. Ecology Letters , 14 , 702 -708 .
[本文引用: 1]
[60]
Wang DN 2012 ) Prevention and control situation and the countermeasures of exotic invasive species Mikania micrantha. Yunnan Agricultural Science and Technology, Suppl
.1 , 235 -236 .(in Chinese)
[本文引用: 1]
[王大能 (2012 ) 西盟县外来入侵有害生物薇甘菊防控现状及对策
. , 云南农业科技 , 235 -236 .]
[本文引用: 1]
[61]
Watkins RZ Chen JQ Pickens J Brosofske KD 2003 ) Effects of forest roads on understory plants in a managed hardwood landscape
. Conservation Biology , 17 , 411 -419 .
[本文引用: 1]
[62]
Yunnan Province Three Parallel Rivers Administration Bureau (2010 ) An overview of the world natural heritage “Three Parallel Rivers” and the protection progress
. Chinese Landscape Architecture , 26 (5 ), 52 -55 .(in Chinese with English abstract)
[本文引用: 1]
[云南省三江并流管理局 (2010 ) 世界自然遗产地——“三江并流”的概况及其保护工作的进展
. 中国园林 ,26 (5 ), 52 -55 .]
[本文引用: 1]
[63]
Zeiter M Stampfli A 2012 ) Positive diversity-invasibility relationship in species-rich semi-natural grassland at the neighbourhood scale
. Annals of Botany , 110 , 1385 -1393 .
[本文引用: 1]
[64]
Zheng L Feng YL 2005 ) The effects of ecophysiological traits on carbon gain in invasive plants
. Acta Ecologica Sinica , 25 , 2782 -2787 .(in Chinese with English abstract)
[本文引用: 1]
[郑丽 , 冯玉龙 (2005 ) 入侵植物的生理生态特性对碳积累的影响
. 生态学报 , 25 , 2782 -2787 .]
[本文引用: 1]
The evolution of weeds
1
1974
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Water relations of native and introduced C4 grasses in a neotropical savanna
1993
Effects of invasive alien plants on fire regimes
1
2004
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
1
1982
... 水分因子包括年降水量(MAP)、最干季降水量(PDQ)、最湿季降水量(PWQ)、年实际蒸散量(AET) (Brutsaert, 1982 )和水分亏缺(water deficit, WD) (Stephenson, 1990 ).其中, 最干(或湿)季降水量是一年内降水最少(或最多)的3个月内的降水总量, 即一个地区冬季(或夏季)的降水量; 年实际蒸散量是一年内地表向大气蒸腾水分的实际值, 受水分和能量双重影响; 水分亏缺反映一个地方的干燥程度.其计算公式如下: ...
Beyond competition: incorporating positive interactions between species to predict ecosystem invasibility
1
2008
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Consequences of a biological invasion reveal the importance of mutualism for plant communities
1
2001
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Upper-montane plant invasions in the Hawaiian Islands: patterns and opportunities. Perspectives in Plant Ecology,
1
2005
... 研究区内入侵物种与乡土物种丰富度沿海拔梯度存在显著不同的空间分布格局, 这个差异可能反映了入侵植物不同于乡土植物的生活史性状与环境需求(van Kleunen et al, 2010 ).外来入侵物种丰富度随海拔上升而下降的格局与其他研究相似, 但这种格局的成因目前还有很大争议, 可能是由于逐渐严酷的非生物环境因子限制了外来种进一步扩散, 或是人为影响减少导致繁殖体扩散能力降低, 也可能仅仅是由于入侵时间较短, 还没有随海拔上升完全占据可利用的生境(Daehler, 2005 ; Giorgis et al, 2011 ; Marini et al, 2012 ). ...
Fluctuating resources in plant communities: a general theory of invasibility
1
2000
... 探讨入侵和乡土植物的分布格局及形成机制是保护生物学研究的重要组成部分.关于环境因子对外来物种分布及其多样性格局的影响存在不同假说.Elton (1958)认为乡土物种丰富度高的地方对入侵的抵抗性强, 但支持这一观点的证据模棱两可(Kennedy et al, 2002 ; Fargione et al, 2003 ).干扰被认为是入侵的先导, 但很多未被干扰的群落也存在外来物种入侵(Lozon & MacIsaac, 1997 ; Davis et al, 2000 ).乡土物种在长期历史过程中已经适应的环境压力可能限制了外来物种的存活和定殖, 但资源可利用性低的栖息地依然受到入侵物种的影响(Harrison, 1999 ). ...
Native and alien plant species in relation to spatial heterogeneity on a regional scale in Germany
1
2003
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Effects of exotic plant invasions on soil nutrient cycling processes
1
2003
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Ecosystem consequences of biological invasions. Annual Review of Ecology, Evolution,
1
2010
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
1
1958
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Climatic limits for the present distribution of beech (Fagus L.) species in the world
1
2006
... 气候变异性因子包括温度季节性(temperature seasonality, TSN)和降水季节性(precipitation seasonality, PSN) (Fang & Lechowicz, 2006 ).温度和降水季节性反映一个地区温度和降水的季节变化程度, 其计算公式如下: ...
Vegetation-climate relationship and its application in the division of vegetation zone in China
1
2002
... 进一步参考相关文献计算多个生物气候指标(Fang et al, 2002 ; Hawkins et al, 2003 ), 分为能量、水分和气候变异性3组.能量因子指标包括年均温(MAT)、最冷季均温(MTCQ)、最暖季均温(MTWQ)和年潜在蒸散量(PET) (Thornthwaite & Hare, 1955 ).其中, 年潜在蒸散量表示在理想状态下, 一年内地表(植被、土壤和水体表面)向大气蒸腾水分的最大可能值, 反映了一个地区的环境能量, 其计算公式如下: ...
Community assembly and invasion: an experimental test of neutral versus niche processes. Proceedings of the National Academy of Sciences,
1
2003
... 探讨入侵和乡土植物的分布格局及形成机制是保护生物学研究的重要组成部分.关于环境因子对外来物种分布及其多样性格局的影响存在不同假说.Elton (1958)认为乡土物种丰富度高的地方对入侵的抵抗性强, 但支持这一观点的证据模棱两可(Kennedy et al, 2002 ; Fargione et al, 2003 ).干扰被认为是入侵的先导, 但很多未被干扰的群落也存在外来物种入侵(Lozon & MacIsaac, 1997 ; Davis et al, 2000 ).乡土物种在长期历史过程中已经适应的环境压力可能限制了外来物种的存活和定殖, 但资源可利用性低的栖息地依然受到入侵物种的影响(Harrison, 1999 ). ...
Physiological responses to short-term water and light stress in native and invasive plant species in southern California
1
2010
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
Abiotic constraints eclipse biotic resistance in determining invasibility along experimental vernal pool gradients
1
2007
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina
1
2011
... 研究区内入侵物种与乡土物种丰富度沿海拔梯度存在显著不同的空间分布格局, 这个差异可能反映了入侵植物不同于乡土植物的生活史性状与环境需求(van Kleunen et al, 2010 ).外来入侵物种丰富度随海拔上升而下降的格局与其他研究相似, 但这种格局的成因目前还有很大争议, 可能是由于逐渐严酷的非生物环境因子限制了外来种进一步扩散, 或是人为影响减少导致繁殖体扩散能力降低, 也可能仅仅是由于入侵时间较短, 还没有随海拔上升完全占据可利用的生境(Daehler, 2005 ; Giorgis et al, 2011 ; Marini et al, 2012 ). ...
Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species
1
2002
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Approaches for testing herbivore effects on plant population dynamics
1
2006
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
Coexistence and interference between a native perennial grass and non-native annual grasses in California
1
1999
... 怒江河谷地区以干热气候为主.能量对入侵和乡土物种丰富度的影响均不显著; 而水分条件, 尤其是最干季降水量的影响非常显著, 显示了干旱对植物分布的限制作用, 且入侵物种对干旱的耐性比乡土物种更低.Baruch和Fernandez (1993)发现一种乡土C4 草本植物对于干旱的耐受性高于另一种入侵C4 草本植物; Hamilton等(1999) 也发现, 美国加州南部草地的干旱环境有利于多年生乡土草本物种的萌芽而不利于一年生入侵草本.怒江河谷的乡土植物在长期适应干旱环境的进化过程中变得更加耐旱, 而入侵种由于进化时间较短, 对干旱生境的适应性较差.对于乡土物种, 裸地比例即路侧生境干扰具有最强的胁迫; 而样方到公路的距离与入侵物种丰富度显著负相关, 这充分显示, 在怒江河谷, 公路沿线的生境干扰对乡土植物群落的退化和外来物种的迁移入侵具有关键作用. ...
The influence of disturbance and habitat on the presence of non-native plant species along transport corridors
1
2005
... 本研究表明, 在群落尺度上外来入侵植物与乡土植物的多样性之间存在显著负相关关系(P < 0.001), 特别是乔木层存在的情况下, 入侵物种丰富度显著低于只有灌草的样方.这表明局域尺度群落结构完整性可能对生物入侵具有抵抗作用.乔木层是否存在对入侵的影响可能与光资源有关.乔木层下的生境光照强度低, 可能抑制了外来植物的竞争力(Watkins et al, 2003 , Hansen & Clevenger, 2005 ); 而光照较强的生境如草地、开放林地以及森林边缘等, 常伴随高温、裸露土壤等条件, 容易被外来植物入侵, 并促使外来植物定居后向更远的距离扩散(Parendes & Jones, 2000 ; Hansen & Clevenger, 2005). ...
Native and alien species diversity at the local and regional scales in a grazed California grassland
1
1999
... 探讨入侵和乡土植物的分布格局及形成机制是保护生物学研究的重要组成部分.关于环境因子对外来物种分布及其多样性格局的影响存在不同假说.Elton (1958)认为乡土物种丰富度高的地方对入侵的抵抗性强, 但支持这一观点的证据模棱两可(Kennedy et al, 2002 ; Fargione et al, 2003 ).干扰被认为是入侵的先导, 但很多未被干扰的群落也存在外来物种入侵(Lozon & MacIsaac, 1997 ; Davis et al, 2000 ).乡土物种在长期历史过程中已经适应的环境压力可能限制了外来物种的存活和定殖, 但资源可利用性低的栖息地依然受到入侵物种的影响(Harrison, 1999 ). ...
Energy, water, and broad-scale geographic patterns of species richness
1
2003
... 进一步参考相关文献计算多个生物气候指标(Fang et al, 2002 ; Hawkins et al, 2003 ), 分为能量、水分和气候变异性3组.能量因子指标包括年均温(MAT)、最冷季均温(MTCQ)、最暖季均温(MTWQ)和年潜在蒸散量(PET) (Thornthwaite & Hare, 1955 ).其中, 年潜在蒸散量表示在理想状态下, 一年内地表(植被、土壤和水体表面)向大气蒸腾水分的最大可能值, 反映了一个地区的环境能量, 其计算公式如下: ...
云南怒江干热河谷农田外来入侵杂草的调查
1
2007
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
云南怒江干热河谷农田外来入侵杂草的调查
1
2007
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Bias and error in understanding plant invasion impacts
1
2013
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Biodiversity as a barrier to ecological invasion
1
2002
... 探讨入侵和乡土植物的分布格局及形成机制是保护生物学研究的重要组成部分.关于环境因子对外来物种分布及其多样性格局的影响存在不同假说.Elton (1958)认为乡土物种丰富度高的地方对入侵的抵抗性强, 但支持这一观点的证据模棱两可(Kennedy et al, 2002 ; Fargione et al, 2003 ).干扰被认为是入侵的先导, 但很多未被干扰的群落也存在外来物种入侵(Lozon & MacIsaac, 1997 ; Davis et al, 2000 ).乡土物种在长期历史过程中已经适应的环境压力可能限制了外来物种的存活和定殖, 但资源可利用性低的栖息地依然受到入侵物种的影响(Harrison, 1999 ). ...
Agamospermous seed production of the invasive tussock grass Nardus stricta L. (Poaceae) in New Zealand: evidence from pollination experiments
1
2006
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Integrative invasion science: model systems, multi-site studies, focused meta-analysis and invasion syndromes
1
2013
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
Alien roadside species more easily invade alpine than lowland plant communities in a subarctic mountain ecosystem
1
2014
... 物种丰富度的空间格局一直是生态学和生物地理学的研究热点.物种丰富度变化和周转速率在纬度和海拔梯度上往往较为显著, 因此备受关注(Lomolino, 2001 ).乡土物种丰富度一般随纬度和海拔单调递减或者呈单峰格局, 尤其是当低海拔人类干扰严重时(Stevens, 1992 ; Rahbek, 1995 ); 而入侵物种丰富度一般随海拔升高呈现单调递减的格局, 其纬度格局则与研究区域的位置和尺度有关.尽管对物种分布空间格局的研究已有很长的历史, 但对外来种随纬度和海拔梯度的入侵格局的研究仍有待深入.开展这方面研究显然有助于增加对物种丰富度决定机制的理解(Lembrechts et al, 2014 ). ...
Species diversity and biological invasions: relating local process to community pattern
1
2000
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Elton revisited: a review of evidence linking diversity and invasibility
1
1999
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
基于马尔柯夫模型的怒江流域中段植被动态变化预测
1
2009
... 怒江是中国西南地区的大河之一, 发源于青藏高原的唐古拉山南麓, 在云南省内流经怒江州的贡山、福贡、泸水和保山地区的保山、施甸、龙陵、昌宁等7县市, 出国境后称为萨尔温江.云南省内的怒江段位于98º07'-100º02' E, 24º07'-28º23' N, 长度650 km, 流域面积3.35万km2 , 占云南省面积的8.7%.云南怒江流域受西南季风影响强烈, 年均气温5.2-16.6℃, 年均降水量961.7-1,639.6 mm.研究区段河谷海拔从南到北递增, 气温相应降低, 而降水量呈递增趋势, 具有亚热带半干旱至半湿润气候; 河谷低地植被稀疏, 以云南松(Pinus yunnanensis )为优势种的针叶林或混交林和灌草丛间断分布(李晖等, 2009 ). ...
基于马尔柯夫模型的怒江流域中段植被动态变化预测
1
2009
... 怒江是中国西南地区的大河之一, 发源于青藏高原的唐古拉山南麓, 在云南省内流经怒江州的贡山、福贡、泸水和保山地区的保山、施甸、龙陵、昌宁等7县市, 出国境后称为萨尔温江.云南省内的怒江段位于98º07'-100º02' E, 24º07'-28º23' N, 长度650 km, 流域面积3.35万km2 , 占云南省面积的8.7%.云南怒江流域受西南季风影响强烈, 年均气温5.2-16.6℃, 年均降水量961.7-1,639.6 mm.研究区段河谷海拔从南到北递增, 气温相应降低, 而降水量呈递增趋势, 具有亚热带半干旱至半湿润气候; 河谷低地植被稀疏, 以云南松(Pinus yunnanensis )为优势种的针叶林或混交林和灌草丛间断分布(李晖等, 2009 ). ...
Invasive alien plants in China: role of clonality and geographical origin
1
2006
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
滇西北三江并流地区公路沿线紫茎泽兰的入侵格局及影响因素
1
2013
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
滇西北三江并流地区公路沿线紫茎泽兰的入侵格局及影响因素
1
2013
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Elevation gradients of species-density: historical and prospective views
1
2001
... 物种丰富度的空间格局一直是生态学和生物地理学的研究热点.物种丰富度变化和周转速率在纬度和海拔梯度上往往较为显著, 因此备受关注(Lomolino, 2001 ).乡土物种丰富度一般随纬度和海拔单调递减或者呈单峰格局, 尤其是当低海拔人类干扰严重时(Stevens, 1992 ; Rahbek, 1995 ); 而入侵物种丰富度一般随海拔升高呈现单调递减的格局, 其纬度格局则与研究区域的位置和尺度有关.尽管对物种分布空间格局的研究已有很长的历史, 但对外来种随纬度和海拔梯度的入侵格局的研究仍有待深入.开展这方面研究显然有助于增加对物种丰富度决定机制的理解(Lembrechts et al, 2014 ). ...
Biological invasions: are they dependent on disturbance?
1997
外来入侵种紫茎泽兰研究进展与展望
1
2005
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
外来入侵种紫茎泽兰研究进展与展望
1
2005
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
地形因素对外来入侵种紫茎泽兰的影响
1
2004
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
地形因素对外来入侵种紫茎泽兰的影响
1
2004
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Scale dependent relationships between native plant diversity and the invasion of croftonweed (Eupatorium adenophorum) in Southwest China
1
2005
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables
1
2002
... 由于物种丰富度通常符合Poisson分布, 采用广义线性模型(generalized linear model, GLM)拟合入侵物种与乡土物种丰富度与环境因子的关系.当出现残差过度分散, 即残差/剩余自由度> 1.2时, 采用quasi-Poisson分布.由于部分气候因子之间存在严重的共线性, 因此先计算自变量之间的相关系数, 在气温、降水、气候变异性3组变量中各选1个组内相关性高而与组间变量相关性低的变量, 加入物种丰富度的多元广义线性模型, 根据赤池信息准则(Akaike Information Criterion, AIC)选出最优模型.基于GLM最优模型, 进一步采用等级方差分离(hierarchical variation partitioning, HVP)算法来定量评价不同自变量对丰富度变异性的独立贡献(Nally, 2002 ).该方法通过穷尽比较包含1个到考虑的全部自变量的所有变量组合构建的模型, 分离出每个自变量的独立效应和联合效应, 并用Z-scores值检验自变量的独立效应是否统计显著. ...
Alien and native plant life-forms respond differently to human and climate pressures
1
2012
... 研究区内入侵物种与乡土物种丰富度沿海拔梯度存在显著不同的空间分布格局, 这个差异可能反映了入侵植物不同于乡土植物的生活史性状与环境需求(van Kleunen et al, 2010 ).外来入侵物种丰富度随海拔上升而下降的格局与其他研究相似, 但这种格局的成因目前还有很大争议, 可能是由于逐渐严酷的非生物环境因子限制了外来种进一步扩散, 或是人为影响减少导致繁殖体扩散能力降低, 也可能仅仅是由于入侵时间较短, 还没有随海拔上升完全占据可利用的生境(Daehler, 2005 ; Giorgis et al, 2011 ; Marini et al, 2012 ). ...
Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment
1
2011
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
1
2000
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Does diversity beget diversity? A case study of crops and weeds
1
1997
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Role of light availability and dispersal in exotic plant invasion along roads and streams in the H. J. Andrews experimental forest, Oregon
1
2000
... 本研究表明, 在群落尺度上外来入侵植物与乡土植物的多样性之间存在显著负相关关系(P < 0.001), 特别是乔木层存在的情况下, 入侵物种丰富度显著低于只有灌草的样方.这表明局域尺度群落结构完整性可能对生物入侵具有抵抗作用.乔木层是否存在对入侵的影响可能与光资源有关.乔木层下的生境光照强度低, 可能抑制了外来植物的竞争力(Watkins et al, 2003 , Hansen & Clevenger, 2005 ); 而光照较强的生境如草地、开放林地以及森林边缘等, 常伴随高温、裸露土壤等条件, 容易被外来植物入侵, 并促使外来植物定居后向更远的距离扩散(Parendes & Jones, 2000 ; Hansen & Clevenger, 2005). ...
Environmental and economic costs of nonindigenous species in the United States
1
2000
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
Plant functional group diversity as a mechanism for invasion resistance
1
2005
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
The elevational gradient of species richness: a uniform pattern?
1
1995
... 物种丰富度的空间格局一直是生态学和生物地理学的研究热点.物种丰富度变化和周转速率在纬度和海拔梯度上往往较为显著, 因此备受关注(Lomolino, 2001 ).乡土物种丰富度一般随纬度和海拔单调递减或者呈单峰格局, 尤其是当低海拔人类干扰严重时(Stevens, 1992 ; Rahbek, 1995 ); 而入侵物种丰富度一般随海拔升高呈现单调递减的格局, 其纬度格局则与研究区域的位置和尺度有关.尽管对物种分布空间格局的研究已有很长的历史, 但对外来种随纬度和海拔梯度的入侵格局的研究仍有待深入.开展这方面研究显然有助于增加对物种丰富度决定机制的理解(Lembrechts et al, 2014 ). ...
Mutualistic fungus promotes plant invasion into diverse communities
1
2005
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Biotic and abiotic influences on native and exotic richness relationship across spatial scales: favourable environments for native species are highly invasible
1
2011
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Arbuscular mycorrhizal inoculum potential: a mechanism promoting positive diversity-invasibility relationships in mountain beech forests in New Zealand?
1
2011
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
Climatic control of vegetation distribution: the role of the water balance
1
1990
... 水分因子包括年降水量(MAP)、最干季降水量(PDQ)、最湿季降水量(PWQ)、年实际蒸散量(AET) (Brutsaert, 1982 )和水分亏缺(water deficit, WD) (Stephenson, 1990 ).其中, 最干(或湿)季降水量是一年内降水最少(或最多)的3个月内的降水总量, 即一个地区冬季(或夏季)的降水量; 年实际蒸散量是一年内地表向大气蒸腾水分的实际值, 受水分和能量双重影响; 水分亏缺反映一个地方的干燥程度.其计算公式如下: ...
The elevational gradient in altitudinal range: an extension of Rapoport’s latitudinal rule to altitude
1
1992
... 物种丰富度的空间格局一直是生态学和生物地理学的研究热点.物种丰富度变化和周转速率在纬度和海拔梯度上往往较为显著, 因此备受关注(Lomolino, 2001 ).乡土物种丰富度一般随纬度和海拔单调递减或者呈单峰格局, 尤其是当低海拔人类干扰严重时(Stevens, 1992 ; Rahbek, 1995 ); 而入侵物种丰富度一般随海拔升高呈现单调递减的格局, 其纬度格局则与研究区域的位置和尺度有关.尽管对物种分布空间格局的研究已有很长的历史, 但对外来种随纬度和海拔梯度的入侵格局的研究仍有待深入.开展这方面研究显然有助于增加对物种丰富度决定机制的理解(Lembrechts et al, 2014 ). ...
Climatic classification in forestry
1
1955
... 进一步参考相关文献计算多个生物气候指标(Fang et al, 2002 ; Hawkins et al, 2003 ), 分为能量、水分和气候变异性3组.能量因子指标包括年均温(MAT)、最冷季均温(MTCQ)、最暖季均温(MTWQ)和年潜在蒸散量(PET) (Thornthwaite & Hare, 1955 ).其中, 年潜在蒸散量表示在理想状态下, 一年内地表(植被、土壤和水体表面)向大气蒸腾水分的最大可能值, 反映了一个地区的环境能量, 其计算公式如下: ...
Climate is a stronger driver of tree and forest growth rates than soil and disturbance
1
2011
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
A meta-analysis of trait differences between invasive and non-invasive plant species
2
2010
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
... 研究区内入侵物种与乡土物种丰富度沿海拔梯度存在显著不同的空间分布格局, 这个差异可能反映了入侵植物不同于乡土植物的生活史性状与环境需求(van Kleunen et al, 2010 ).外来入侵物种丰富度随海拔上升而下降的格局与其他研究相似, 但这种格局的成因目前还有很大争议, 可能是由于逐渐严酷的非生物环境因子限制了外来种进一步扩散, 或是人为影响减少导致繁殖体扩散能力降低, 也可能仅仅是由于入侵时间较短, 还没有随海拔上升完全占据可利用的生境(Daehler, 2005 ; Giorgis et al, 2011 ; Marini et al, 2012 ). ...
Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems
1
2011
... 植物群落中生物入侵程度的影响因子一般可分为生物与非生物因素.生物因素主要包括扩散限制、物种间的竞争或互利关系、天敌是否存在等(Halpern & Underwood, 2006 ; Vilà et al, 2011 ); 非生物因素包括气温、降水、养分及光资源的可利用性等, 以及干扰带来的生态位空缺(Funk & Zachary, 2010 ; Matzek, 2011 ; Toledo et al, 2011 ).种间竞争较弱、环境压力较小而可利用资源丰富, 或干扰体系发生改变的群落容易被入侵.然而, 生物及非生物因子之间存在复杂的相互作用, 不同机制对生物入侵影响的相对重要性尚无定论(Kueffer et al, 2013 ). ...
西盟县外来入侵有害生物薇甘菊防控现状及对策
1
2012
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
西盟县外来入侵有害生物薇甘菊防控现状及对策
1
2012
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Effects of forest roads on understory plants in a managed hardwood landscape
1
2003
... 本研究表明, 在群落尺度上外来入侵植物与乡土植物的多样性之间存在显著负相关关系(P < 0.001), 特别是乔木层存在的情况下, 入侵物种丰富度显著低于只有灌草的样方.这表明局域尺度群落结构完整性可能对生物入侵具有抵抗作用.乔木层是否存在对入侵的影响可能与光资源有关.乔木层下的生境光照强度低, 可能抑制了外来植物的竞争力(Watkins et al, 2003 , Hansen & Clevenger, 2005 ); 而光照较强的生境如草地、开放林地以及森林边缘等, 常伴随高温、裸露土壤等条件, 容易被外来植物入侵, 并促使外来植物定居后向更远的距离扩散(Parendes & Jones, 2000 ; Hansen & Clevenger, 2005). ...
世界自然遗产地——“三江并流”的概况及其保护工作的进展
1
2010
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
世界自然遗产地——“三江并流”的概况及其保护工作的进展
1
2010
... 滇西北三江并流地区是具有全球重要性的生物多样性热点地区(云南省三江并流管理局, 2010 ).包括紫茎泽兰(Eupatorium adenophorum )在内的外来植物已经入侵这一区域, 在一定程度上对当地生态环境和国民经济造成了严重影响(卢志军和马克平, 2004 ; Lu & Ma, 2005 ; 胡发广等, 2007 ).目前, 关于这一区域生物入侵的研究或是基于单个物种(刘晔等, 2013 ), 或是以人工生态系统为本底进行调查(王大能, 2012 ), 缺少入侵物种和乡土物种丰富度的对比, 并且不能确定自然和人类干扰因素对生物入侵影响的贡献.本文基于对怒江河谷公路沿线植物入侵的详尽群落学调查, 试图研究以下3方面的问题: (1)怒江河谷地区入侵植物和乡土植物丰富度的空间格局; (2)入侵和乡土物种多样性之间的关系; (3)人为干扰、怒江河谷的自然环境以及植物群落结构等因子对入侵物种和乡土物种丰富度影响的相对大小.希望通过此研究, 能够定量评价在生物多样性丰富的区域, 决定入侵植物多样性的不同机制及其相互之间的关系, 为外来入侵植物的预防、控制和管理提供科学依据. ...
Positive diversity-invasibility relationship in species-rich semi-natural grassland at the neighbourhood scale
1
2012
... 物种多样性和群落可入侵性之间的关系对被入侵植物群落的恢复与管理十分重要, 而关于外来入侵物种与乡土物种丰富度之间的关系仍存在争议(Gerhardt & Collinge, 2007 ; Souza et al, 2011 ).支持乡土物种与入侵物种多样性负相关的观点认为, 群落生态位空间有限, 竞争在群落构建中起主导作用, 因此物种丰富的群落利用了大部分资源, 为新物种留下的资源较少, 从而减少了外来物种入侵的可能性(Palmer & Maurer, 1997 ; Zeiter & Stampfli, 2012 ).且物种丰富的群落含有与外来种性状相似的乡土物种的可能性更高, 因而可入侵性更低(Levine & D’Antonio, 1999 ; Pokorny et al, 2005 ).相反的观点认为, 群落中存在种间促进(facilitate)效应, 如遮阴、菌根供养(Rudgers et al, 2005 ; Bulleri et al, 2008 ; Spence et al, 2011 ), 导致物种越丰富的群落能够共存的物种也越多, 因而促进外来种入侵.也有观点认为乡土和入侵物种丰富度的关系取决于空间尺度: 即当外在干扰程度较轻或中等时, 局域尺度的乡土物种丰富度通过种间相互作用, 增加了群落对物种入侵的抵抗(Elton, 1958 ); 而在更大的空间尺度上则呈现正相关关系(Levine, 2000 ; Deutschewitz et al, 2003 ), 这可能与二者在不同尺度上对环境因子的响应有差异相关. ...
入侵植物的生理生态特性对碳积累的影响
1
2005
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...
入侵植物的生理生态特性对碳积累的影响
1
2005
... 生物入侵是全球变化的重要组成部分(Mooney & Hobbs, 2000 ), 已在从局域到全球各个尺度上造成严重的生态后果(Christian, 2001 ; Ehrenfeld, 2003 )和经济损失(Pimentel et al, 2000 ).生物入侵威胁入侵地的生物多样性, 削弱生态系统结构和功能.例如, 生物入侵通过改变可燃物组成和性状, 影响到生态系统的火烧频率和强度(Brooks et al, 2004 ), 改变生态系统的养分循环、水分动态以及土壤生化环境, 进而改变乡土物种丰富度和种群多度(Ehrenfeld, 2010 ; Hulme, 2013 ).由于入侵种一般生长速度快(Grotkopp et al, 2002 ; van Kleunen et al, 2010 ), 光合速率(郑丽和冯玉龙, 2005 )和种子产量高(鲁萍等, 2005 ; Kissling et al, 2006 ), 许多具有克隆繁殖能力(Baker, 1974 ; Liu et al, 2006 ), 因此入侵后易于成为群落优势种. ...










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}