传统的生物多样性与生态系统功能(biodiver- sity and ecosystem functioning, BEF)研究多关注植物物种丰富度和生态系统净初级生产力(NPP)之间的关系。显然, 单个生态系统功能指标(如NPP)并不能完全反映多样性对生态系统功能的影响(Hector & Bagchi, 2007 ; Gamfeldt et al, 2008 ; Zavaleta et al, 2010 ), 因为一个生态系统具有同时提供多种功能和服务的能力, 这被称为生态系统多功能性(ecosystem multifunctionality, EMF) (Hector & Bagchi, 2007 )。
EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识。与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 )。单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 )。不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平。
目前, 生物多样性与生态系统多功能性之间的关系(biodiversity and ecosystem multifunctionality, BEMF)已成为生态学研究的热点之一; 但国际上对研究这一问题的关键技术——EMF的量化方法存在不一致性, 使得不同的研究之间难以进行比较(Byrnes et al, 2014a ), 限制了BEMF研究的进一步发展。Hector和Bagchi (2007)首次用功能-物种替代法(turnover approach)来量化EMF, 有着里程碑式的意义。这一研究让BEMF走入人们的视野, 很多研究者由此意识到EMF的量化对研究BEMF是不可或 缺的。
其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF。比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法。之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF。Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平。然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多。除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 )。
本文根据目前已发表的文献, 对其中报道的EMF测度方法进行分类, 同时还对各方法的使用情况进行了统计(表1 )。
根据表1 的结果可知, 本文介绍的7种EMF测度方法几乎涵盖了所有已发表文献中所用的方法。单功能法可参考传统的BEF研究, 而直系同源基因法因自身的特点(将在下文详细介绍)使用得较少。在不考虑期刊影响因子差异的情况下, 平均值法在文献中的使用频率显著高于其他方法, 这说明研究者偏向于使用较为简单、直观的EMF量化方法; 若是考虑到期刊影响因子的差异, 单从重要的综合性期刊(如Nature 、Science 、Nature Communications 等)上发表的文章来看, 研究者倾向于多种方法(即单功能法、功能-物种替代法、平均值法、单阈值法和多阈值法这5种方法)并用; 除了单功能法, 其他几种方法的使用频率相差较小。因多阈值法出现的时间较晚(2014年), 所以当前的使用频率较低, 但未来可能会有更多的应用。此外, 在多阈值法出现后, 因其克服了单阈值法的缺点, 已基本取代了单阈值法, 在2015年已发表的文献中, 没有研究者再采用单阈值法来量化EMF。多元模型法在2015年才被提出, 它克服了上述方法的一些缺陷, 分析过程中丢失的信息最少, 但其实用性仍需要在未来的研究中进行检验。
上述提及的EMF的测度方法各有其优缺点, 每种方法的侧重方面也不尽相同。研究者常常根据自身研究的需要, 使用相对合适的方法。有的研究者(如Bradford et al, 2014a ; Jing et al, 2015 ; Lefcheck et al, 2015 )则选择了1种以上的测度方法, 这样不同方法可以互相补充。这虽然有助于更好地研究生物多样性对EMF的潜在影响, 但是也突显了不同方法之间存在的缺陷, 不利于研究间的对比。因此, 未来研究的一个重要任务是开发一种具有普遍适用性的EMF量化方法(Bradford et al, 2014b ; Byrnes et al, 2014b ), 从而实现不同研究之间的整合分析, 促进BEMF研究的发展。
22年前, 《生物多样性》对生物群落多样性测度方法的介绍(马克平, 1994 ),在一定程度上促进了国内对生物多样性的研究。在此启发下, 本文简述了7种EMF量化方法的基本原理及其特点, 以帮助研究者选择合适的测度方法。对于其中较难理解的多阈值法, 将利用我们所收集的青藏高原样带数据(Jing et al, 2015 )说明其在BEMF研究中的应用。
1 生态系统多功能性测度方法
1.1 单功能法
检验生物多样性对EMF的影响最简单的方法是测定生态系统的各种功能, 进而与生物多样性建立关系, 从而判定EMF与生物多样性的关系。研究者常用一般线性模型(general linear model)进行分析, 各个生态系统功能独立地和生物多样性建立回归关系, 根据回归结果定性地判断EMF是否随着多样性的增加而提高。
单功能法源自经典的BEF实验, 最早由Duffy等(2003) 提出, 用于检验以海草为食的海生无脊椎动物物种丰富度对生态系统功能的影响。结果发现,当无脊椎动物的物种丰富度达到最高水平(6个物种)时, 生态系统的多个功能能够同时达到最大值。作者由此认为维持多个较高水平的生态系统功能需要更多的物种来支持。
该方法的结果易受到抽样效应(sampling effect)和互补效应(complementarity)的共同影响(Tilman et al, 1997a ; Loreau & Hector, 2001 ), 其中互补效应起主要作用(详见Loreau & Hector, 2001 ; Cardinale et al, 2007 ; Fargione et al, 2007 )。所谓抽样效应是一个统计学的概念, 指在同一生境中, 竞争力强的物种对资源的利用率更高, 故而有更高的生产力, 随着物种丰富度的增加, 从物种库中抽到高产物种的概率也将增大(Tilman et al, 1997b ); 所以在随机抽样的情况下, 高多样性的群落包含高产物种的概率更大。将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 从物种库中得到对某几个功能或所有功能都有很强促进作用的物种的概率会增大, 从而得出物种丰富度增加会提高多功能性的结论。互补效应则是指在一个群落中, 物种间存在生态位差异, 故而群落中物种数越多, 所占据的“功能空间” (functional space)范围越广(Tilman et al, 1997b )。因此, 生态系统的物种丰富度越高, 对资源的利用率就越高, 相应的生产力也会越高。将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 物种间的生态位差异增大, 不同物种支持不同的功能, 同样会得到因物种丰富度增加而多功能性提高的结论。
1.2 功能-物种替代法
单功能法的缺陷之一是某些功能可能受到一个或几个物种的影响较大。为了解决这个问题, 研究者提出了功能-物种替代法(简称物种替代法)。所谓物种替代, 即不同的功能是由不同的物种所驱动, 即功能-物种替代。如果完全相同的物种驱动不同的功能, 则不存在物种替代; 反之, 不同的功能由不同的物种来驱动, 则存在完全物种替代。物种替代率的高低可根据物种重叠率(overlap)的高低来判断, 该方法最早由Hector和Bagchi (2007) 提出。
该方法通过量化维持EMF的物种数和每个物种对EMF贡献的冗余度来评估功能的替代, 其计算分为两步:
第一步, 通过模型模拟每个物种的作用, 以此来量化物种对EMF的影响。依据最小AIC (Akaike Information Criterion)标准来选择模型, 用反向-剔除多元回归分析(backward-elimination multiple regression analysis)将物种依次从模型中剔除, 计算相应的AIC值。通过比较各物种剔除前后的AIC值, 将导致AIC值减小最多的物种彻底剔除, 不断重复此过程, 直至剔除任意一个物种都会使AIC值增加, 这时便得到了最优模型。未被剔除的物种集合就是影响该生态系统功能的最简物种组合(Hector & Bagchi, 2007 )。该模型对数据的要求较高, 通常要求不同多样性梯度下都有很好的可重复性(Byrnes et al, 2014a )。
利用上述分析得到的物种集合, 通过模型拟合以检测对某一功能组合有影响的物种, 从而判断支持不同功能的物种重叠度, 以此获取功能数与影响一个或一个以上功能的最小物种数之间的关系(Hector & Bagchi, 2007 )。因不同功能间物种的替代率与重叠度成反比, 故物种的替代率可由重叠度来计算。影响每一对功能的物种, 其重叠度可用Sørenson指数o 表示, 计算公式为:
式中, |Ei |表示对功能i 有贡献的物种数目, |Ej |表示对对功能j 有贡献的物种数目, |Ei ∩Ej |表示对功能i 和j 都有贡献的物种数目。
第二步, 进行生态系统多功能性分析。一般利用泊松广义线性模型(Poisson generalized linear model)计算影响单个生态系统功能的平均物种数目 E )与维持这些功能所需的平均物种数(SE )之间的关系即可根据以下公式计算:
根据上式可以定量预测一个生态系统中, 随着所考虑的生态系统功能数的增加, 维持这些功能所需的平均物种数。当对所考虑的功能与维持这些功能所需的物种数进行回归时, 有2种极端的情况: (1)如果一个物种只影响一个功能( ) 之间存在正向线性关系, 其斜率为1 (Hector & Bagchi, 2007 ); (2)如果生态系统所有的功能都受同一物种集合的影响( Hector & Bagchi, 2007 )。在实际研究中, 结果往往介于两者之间, 表明物种和功能间存在一定的替代(Hector & Bagchi, 2007 ; Isbell et al, 2011 )。
然而, 因为物种库中的物种对不同的功能有不同的作用, 而多样性对多功能性的影响取决于正负效应之间的权衡。若正效应大于负效应, 则随着多样性的增加, 多功能性将增加; 反之则降低。Hector和Bagchi (2007)的方法目前仅被用于各物种对生态系统功能具有正效应的情况(He et al, 2009 ; Isbell et al, 2011 ), 而在很多情况下并非如此。当将负效应考虑进来时, 就可以通过检验物种正效应是否强于、等于或者弱于负效应以及是否有物种主导整体功能的正负效应, 来更好地揭示BEMF的潜在关系。为此, Byrnes等(2014a)提出了2个额外的分析步骤来完善该方法:
(1)分别测定各物种对不同生态系统功能的作用, 及受这些物种显著影响的功能间的相关性。通过检验可以评估功能之间潜在的权衡或者是否有物种对生态系统功能有显著的正/负效应; (2)利用上述数据, 通过检验各物种对其影响的所有功能的平均正/负效应(均需标准化)间的关系, 进而量化功能间的权衡关系。通过这两步就可以知道该物种集合对生态系统整体功能正/负效应的强弱关系(Byrnes et al, 2014a )。当物种集合对不同的生态系统功能有显著的正效应, 且不同功能的平均正效应大于平均负效应, 那么物种替代很可能会对生物多样性和生态系统多功能性的关系有正向作用。
1.3 平均值法
顾名思义, 该方法就是将不同功能的测定值进行转化、平均, 最后得到一个可以代表所测功能平均水平的指数, 即多功能性指数(multifunctionality index)。该方法最早由Hooper和Vitousek (1998)提出, 当时被称为相对资源利用指数(index of relative resource use), 可用于量化不同植物对养分的消耗。这是研究者第一次将多个生态系统过程用一个指数来进行表征, 实际上就是对多功能性的一种量化, 因此该方法可以说是最早出现的多功能性量化方法。其实, 用主成分分析的方法也可以得到类似的多功能性指数: 对所有的生态系统功能作主成分分析, 得到轴一、轴二的得分, 用其表示生态系统的多功能性。目前, 这种量化方法还未有研究者使用, 在未来的研究中可以作进一步的探索。
用平均值法计算多功能性较为简单、直观, 先将样方中的生态系统功能值进行转化, 然后将数据标准化, 最后计算各个功能值的均值作为多功能性指数。当样方面积较大(如30 m × 30 m)、植被覆盖率较低时, 需先根据裸地面积和有植被覆盖区域面积的比例对所测功能值进行加权平均, 然后进行转化和标准化(Maestre et al, 2012b )。计算多功能性指数MFa 的公式为:
(3)
式中, F 表示测定的功能数, fi 表示功能i 的测定值, ri 是将fi 转换成正值的数学函数, g 表示将所有的测定值标准化。由于有些功能值不能直接反映生态系统的功能, 比如土壤N含量表征的是植物N吸收的能力, 一般情况下, N含量越低, 植物N吸收能力越高。因此, ri (fi ) 可以采用max(fi ) - fi 的形式表示, max(fi )为前5%的观测值的平均值, 所得结果越大, 表明植物N吸收能力越强。在求平均值之前, 所有的功能值要先标准化, 以保证不同功能是在同一尺度上进行相互比较。虽然功能值标准化的方法有很多, 但最终的结果都较为相近(Maestre et al, 2012b )。这里推荐最常用的两种标准化方法。一是Z得分(Z Scores) (Mouillot et al, 2011 ; Maestre et al, 2012b ; Bradford et al, 2014a ; Wagg et al, 2014 ): 即用平均值和标准偏差来进行标准化(测定值减去平均值后除以标准偏差); 二是用最大值转化(Hooper & Vitousek, 1998 ; Maestre et al, 2012a ): 每个功能排名前5%的测定值取平均值后作为该功能的最大值, 然后计算每个测定值与最大值的比值。
当然, 这两种标准化方法均存在一定的缺陷: Z得分法使得功能值更适合用线性模型分析(Maestre et al, 2012b ; Bradford et al, 2014a ), 而用其他模型计算时可能会产生较大的误差; 最大值转化则易受异常值的影响。一般情况下, 在计算MFa 时, 各个功能值的权重是相同的, 研究者亦可根据研究需要对权重进行调整; 例如Maestre等(2012b)曾根据样方内植被覆盖面积和裸地面积的比值对所测功能赋以不同的权重。
1.4 单阈值法
单阈值法的出现将多功能性的测度转向了另一个方向, 即评估随着多样性的增加达到某一阈值水平的功能数的变化。该方法通过计算每个生态系统中达到某一阈值的功能数来求得一个指数, 该指数表示在该阈值条件下这个生态系统整体功能的水平(Zavaleta et al, 2010 )。
此方法的一个关键步骤是阈值的确定。阈值通常是指每个功能所观测到的最大值的比例(如25%、50%、75%等) (Zavaleta et al, 2010 ), 通常用最大值法来确定某一功能在所研究生态系统中的最高水平。实验中观测到的最大值因实验误差或其他原因可能是异常值, 为此, 研究者常取某一个功能前n + 1个观测值的平均值作为最大值(n 表示单个物种处理时的最小样本量) (Byrnes et al, 2014a ; Perkins et al, 2015 ), 或取观测值前5%的均值作为最大值(与平均值法中介绍的最大值转换类似) (Zavaleta et al, 2010 )。得到最大值之后, 下一步就是确定阈值ti , ti 由最大值乘以一定的百分比得到。研究者可根据研究目的自行选择某一百分比。但必须注意的是, 对于所选的百分比, 必须有合适的理由进行解释, 否则不同的阈值可能导致不同的结果, 所得的结果难以让人信服。
Gamfeldt等(2008) 最早使用该方法来评价生态系统多功能性, 他们选择所测功能最大值的50%作为阈值来评价每个功能在生态系统中的表现, 理由有两点:
(1)因为缺少一个普遍的标准去判断一个生态系统功能是否处于危险状态(即该功能处于消失的边缘), 他们借鉴生态毒理学中的半数致死效应浓度(concentration giving 50% of maximum effect, EC50 ), 选择了一个较低的阈值。如果一个生态系统中包含能提供某个功能最大值50%的物种, 他们就认为该生态系统能维持此功能。
(2)根据Cardinale等(2006)的整合分析(meta-analysis)结果: 对于任意一个功能(如养分吸收), 维持其过程的最大速率的60%, 一般需要1个物种。因此, 当使用最大值的50%作为阈值时, 维持一个功能将只需要1个物种, 这就能使各功能达到阈值的概率最大化。
在确定了每个功能的阈值后, 根据多功能性指数计算公式, 即可求得对应阈值下的多功能性指数。此方法与平均值法计算多功能性指数不同, 因各个功能分别与对应的阈值进行比较, 这样各个功能值不需要经过尺度上的转化。多功能性指数计算公式如下:
(4)
式中, F 表示总的功能数, fi 表示某一生态系统内功能i 的值(用数学函数ri 进行转换), ti 表示阈值。由公式(4)可知, 这是一个累加的方程, 可以根据需要对不同的功能选择不同的阈值。在计算时只需改变对应的阈值ti 即可。
1.5 多阈值法
阈值的选择与最终得到的多功能性指数有直接的关系, 但在实际应用中, 往往很难准确地选择恰当的阈值, 于是便出现了多阈值法。多阈值法由Byrnes等(2014a)提出, 计算多功能性指数MFt 的方法与单阈值法相同, 即计算达到阈值的功能数, 每个阈值的确定也与单阈值法相同。两者的差异是多阈值法有多个阈值, 包含了0-100%的所有阈值, 对每个阈值都计算相应的多功能性指数。
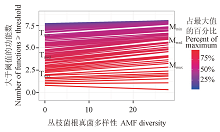
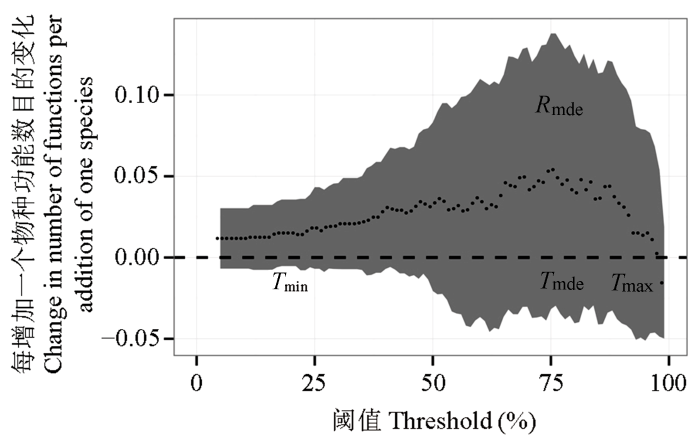
为了便于读者理解, 本文根据青藏高原样带数据(Jing et al, 2015 )进行说明。通过检验不同阈值下丛枝菌根真菌多样性与多功能性指数MFt 拟合曲线的斜率变化(图1 )来检测多样性对多功能性的影响。将阈值与对应的拟合曲线的斜率建立关系(图2 ), 可以得到多样性如何影响多功能性的大量信息, 而不仅仅局限于何时多样性的作用最大(即斜率最大)。
图1 丛枝菌根真菌(AMF)多样性和达到所测功能最大值的某一阈值的功能数之间的关系。 不同颜色表示不同的阈值, 冷色调表示低阈值, 暖色调表示高阈值。T min 表示斜率不等于0的最小阈值, T mde 表示斜率最大时的阈值, T max 是斜率与0有显著差异时的最大阈值。带M 的指标表示相应阈值下, 达到该阈值的功能数(数据来自Jing et al, 2015 )。
Fig. 1 The relationship between diversity of arbuscular mycorrhizal fungi (AMF) and the number of functions beyond a threshold of the maximum observed value. Different colors indicate different thresholds. Cooler colors imply lower thresholds; and warmer colors higher thresholds. T min is the minimum threshold whose slope is significantly different from 0. T mde is the threshold with the steepest slope. T max is the maximum threshold where the slope is not significantly different from 0 again. All indices preceded by M indicate the number of functions at or beyond the threshold (Data from Jing et al, 2015 ).
图2 在不同的阈值下, 丛枝菌根真菌(AMF)多样性和达到所测功能最大值的某一阈值的功能数之间关系的斜率。各个点是拟合值, 阴影表示±1的置信区间。T min 表示斜率不等于0的最小阈值, T mde 表示斜率最大时的阈值, T max 是斜率再次不等于0时的最大阈值, R mde 表示在T mde 处估计的最大斜率(数据来自Jing et al, 2015 )。
Fig. 2 The slope of the relationship between diversity of arbuscular mycorrhizal fungi (AMF) and the number of functions beyond a threshold of the maximum observed value at different thresholds. Points are the fitted values and shading indicated ±1 CI. T min is the lowest threshold whose slope is significantly different from 0. T mde is the threshold with the steepest slope. T max is the maximum threshold where the slope is not significantly different from 0 again. R mde indicates the maximum slope estimated at T mde (Data are from Jing et al, 2015 ).
结合图1 和图2 的曲线, 可以获得一些关键的度量指标来帮助理解多样性与多功能性之间的关系。通过4个主要指标可以获得多样性如何影响多功能性的信息; 而借助4个辅助指标可以对不同的系统和实验进行对比(Byrnes et al, 2014a ; Perkins et al, 2015 )。4个主要指标为:
(1)最低阈值(minimum threshold, T min ): 表示多样性对多功能性有影响时(斜率显著大于或小于0)最小的阈值。
(2)最高阈值(maximum threshold, T max ): 表示从最低阈值开始, 随着阈值的增大, 多样性对多功能性的影响变得不显著(斜率与0没有显著差异)时的阈值, 即斜率不为0时的最大阈值。
(3)多样性效应最大时的阈值(threshold of maximum diversity, T mde ): 表示多样性具有最大的正效应或负效应(斜率最大或最小)时的阈值。
(4)多样性的最大效应(realized maximum effect of diversity, R mde ): 表示多样性对多功能性影响最大时的斜率, 表征了生物多样性对达到阈值的功能数的最大影响, 可在T mde 处计算。
根据上述4个主要指标就可以判断多样性与多功能性的关系: 当T min 很小, T max 很大, 且T mde 、R mde 都较大时, 说明多样性可以有效地驱动多功能性(Byrnes et al, 2014a )。为进行不同系统和实验之间的对比, 需要结合以下4个辅助指标:
(1)最低阈值处的多功能性(minimum diversity-independent multifunctionality, M min ): 表示达到最低阈值(T min )的功能数;
(2)最高阈值处的多功能性(maximum diversity-independent multifunctionality, M max ): 表示达到最高阈值(T max )的功能数;
(3)多样性作用最大时的多功能性(diversity- maximized multifunctionality, M mde ): 多样性对多功能性的作用最大(斜率最大或最小)时, 达到相应阈值的功能数;
(4)可能的最大多样性作用实现的百分比(percentage of maximum possible diversity effect, P mde ): 即R mde 除以实验中可能出现的最大斜率(总功能数除以总物种数), 表示多样性对多功能性的相对重要性在实验中实现的百分比。
在获得以上的辅助指标后, 结合T min 和M min 可以判定一个生态系统中多样性对多功能性影响的最低阈值, 及在该阈值水平下多样性对多功能性有多大的影响; 结合T max 和M max 可以获得在该生态系统中能同时维持的高水平功能数; 结合T mde 和M mde 可以获得多样性作用最大时达到相应阈值的功能数; 结合R mde 和P mde 可以获得多样性对多功能性的相对重要性在该生态系统中实现的程度(Byrnes et al, 2014a )。通过以上分析就可以对比不同实验或生态系统的多功能性。
1.6 直系同源基因法
直系同源基因是指不同物种中来自共同祖先的基因, 通常认为其功能保守, 因此具有相似的功能(Fang et al, 2010 )。该方法由Miki等(2014)提出, 用于测度微生物群落的多功能性, 直系同源基因丰富度越高则群落的多功能性越高。Miki等(2014)先在理论上进行探讨, 认为不同物种所拥有的同类直系同源基因具有相似的功能, 不同类别的直系同源基因具有不同的功能, 因此通过评估微生物群落中直系同源基因的丰富度就能知道群落功能的大致范围。之后, 他们通过微宇宙实验证明, 利用群落中微生物的直系同源基因序列丰富度可以预测实际的多功能性(Miki et al, 2014 )。用该方法测度多功能性时, 最好选择系统发生相近的微生物类群。
该方法的第一步是根据研究需要从微生物基因组数据库(microbial genome database, MBGD)中 (http://mbgd.genome.ad.jp/ )下载相关数据作为默认序列表(default orthologue table) (Uchiyama et al, 2010 ), 数据以碱基序列表征。因为一个物种可能会有很多具有完全相同序列的基因组, 因此需要将每个物种的序列簇表(orthologue cluster table)转换成二进制的矩阵, 以1或0表示某一序列存在与否, 1表示存在, 0表示不存在。转换后的表格就是简单的物种×序列矩阵, 可以方便地对测定的每种微生物所拥有的基因序列进行统计。
根据预防原则(precautionary principle), 为避免低估微生物群落的多功能性, 将群落中已知功能的序列组和没有特定功能的序列组(通过假设蛋白或没有特定功能的蛋白来预测直系同源基因)的总数定义为多功能性指数MF , 以此来判断一个群落多功能性的高低。
之后, 可以根据一个群落中微生物物种丰富度(species richness, SR )和多功能性指数MF 建立幂指数关系来估计群落的多功能冗余, 如下式所示:
(5)
式中, c 表示群落中只有一个物种时的平均多功能性, 即群落中每个物种所拥有的平均直系同源基因数; a 代表多功能冗余指数, a 越小表示多功能冗余越大, 根据该指数可以估计微生物群落的多功能冗余情况。
此方法目前应用不多, 仅有一篇文章(Miki etal, 2014 )采用了该方法。尽管如此, 从开发新的多功能性量化方法的角度来看, 这种方法从基因的角 度出发, 对量化微生物多功能性进行了有益的探 索, 对未来建立具有普遍适用性的量化方法有启发作用。
1.7 多元模型法
以上介绍的6种方法, 在分析过程中或多或少都会丢失一些重要的信息, 导致对EMF的分析不够全面。为了尽可能全面地了解BEMF的关系, Dooley等(2015)提出了以BEF研究中使用的多样性交互模型(Kirwan et al, 2009 )为基础扩展而成的多元模型法。该方法克服了上述方法仅将物种丰富度作为多样性的缺点, 在分析过程中考虑了不同方面的多样性(如物种相对多度、均匀度、物种组成等)对EMF的影响。同时, 该方法还可以分析功能之间的权衡和相关性, 并能检验整体功能在何时达到最大值。
多元模型法的基础是多样性交互模型(详见Kirwan et al, 2009 ), 该模型可以评估物种属性和多样性对生态系统功能的影响, 如下式所示:
(6)
式中, y 表示一个生态系统功能, Pi 和Pj 分别表示第i 和第j 个物种的相对多度(i , j = 1, 2, …, s)。A 包含了群落多度或不同的处理, α 则是一个包含了若干系数的矢量。βi 表示第i 个物种在单作时该功能预期的表现。δij 表示物种i 和j 之间的相互作用。 ε 为残差, 且ε ~ N(0, σ2 )。
公式(6)是针对一个生态系统功能的模型, 将之推广到多个功能后, 任意一个功能可以表示为:
(7)
式中, yk 表示功能k , βik 表示物种i 在单作时, 功能k 的预期表现, δijk 表示物种i 和j 对功能k 的交互作用(Dooley et al, 2015 )。为了减少模型系数, 使整个模型更为简洁, 通过一些有生物学意义的假设, 可以将表示物种间交互作用的系数δijk 进行简化(Kirwan et al, 2009 )。例如,假设功能群决定物种间的交互作用, 以2个功能群为例, 当物种i , j 都来自功能群1 (功能群2)时, δijk = δwfg1k (δwfg2k ) ; 当i , j 来自不同功能群时, δijk = δbfgk ; 此处wfg 表示属于同一个功能群, bfg 表示2个不同的功能群(Dooley et al, 2015 )。
在用该方法分析前, 需先对所测的功能值进行转化, 将不同功能转化到同一尺度, 转化方法可参照前文平均值法。将原始值转化后代入上述模型, 用最大似然法或限制性最大似然法对每个功能进行拟合, 根据模型拟合的系数和预测能力, 对拟合的模型进行检验和比较, 以期找出最优模型(最简模型), 不同模型间的比较可用似然比值。
在确定了最优模型后, 先用t 检验对模型系数进行比较, 但在比较过程中要注意多次成对比较所带来的问题, 如比较3个功能e 、f 、k 所得模型的系数βie 、βif 和βik , 在比较时, 有3次成对t 检验, 这时可以用Bonferroni校验法来避免多次比较造成的误差(Dooley et al, 2015 )。之后, 根据不同物种相对多度、物种组成、均匀度及所设置的处理, 利用上述模型, 对各生态系统功能的表现进行预测, 并预先设置一个阈值(尽量取值大一些, 如70%)来与各功能值进行比较。根据对模型系数的分析和模型预测的各功能的表现, 可以从不同角度探讨BEMF的关系。讨论的内容主要包括两个方面:
(1)单作时的多功能性。通过比较不同模型的系数β , 可以得到不同物种在单作时各功能表现的差异, 从而得到各物种对不同功能的相对重要性、不同功能表现的变异性、相关性以及功能之间的权衡(如单作时, 当某个功能值很高时, 其他的功能处于相对较低水平, 说明这个功能与其他功能之间存在权衡) (Dooley et al, 2015 )。
(2)混种时的多功能性。通过比较不同模型的系数δ , 可以得到物种间的交互作用对各功能的影响, 从而判断种间交互作用能否驱动某个功能。根据功能值是否超过提前设定的阈值, 从而判断该功能是否达到预期的水平, 因此, 可以在特定的物种均匀度下, 根据不同的物种组成, 将预测的功能值与阈值进行对比, 从而可以判断哪个物种为优势种时, 各功能的表现最好。在不同的物种均匀度下, 可以检验随着物种均匀度的增加, 各个功能的表现是否也随之增加; 同时, 考虑到在一定的物种组成条件下, 不同功能间可能存在较大的差异, 因此, 可以检验随着物种均匀度的增加, 功能间的变异性是否发生变化。此外, 在相同的物种组成和均匀度下, 则可以比较物种相对多度对各个功能的影响(Dooley et al, 2015 )。
总的来讲, 通过以上两个方面的探讨, 研究者可以同时掌握单个功能的信息和多个功能的整体情况, 相当于是单功能法和平均值法的结合, 同时又有物种替代法的优点, 可以获得不同物种对各个功能相对重要性的信息, 还可以根据研究目的设置阈值, 将功能值与阈值进行对比, 从而可以实现单阈值法的功能。
2 不同量化方法的特点
2.1 单功能法
该方法是阈值法的前身, 对生态系统的每个功能和多样性之间的关系都进行了分析, 有助于阐明多功能性是由哪一个功能所驱动的。但该方法有多个方面的不足: (1)难以评估生态系统的整体功能; 当一个实验中, 多样性对一个生态系统的不同功能同时出现正、负效应时, 难以解释实验结果。(2)不能完全体现生物多样性效应;对于一个具有高多样性的群落而言, 有些物种对于一些功能来说可能是冗余的, 而对于其他功能来说, 这些物种又起着一定的作用。(3)只能作定性描述而不能进行定量分析。(4)实验结果易受个别物种的影响。(5)未考虑功能之间的相关性(Dooley et al, 2015 )。(6)不同功能对生态系统的重要性不同, 该方法未考虑不同功能的权重。
该方法单独使用时所得结果不易让人信服, 但是可以作为辅助方法, 与其他测度方法(如平均值法、单阈值法等)一起使用(Jing et al, 2015 )。
2.2 功能-物种替代法
物种替代法主要关注不同的功能是否由不同物种驱动以及维持一定数量的功能需要多少个物种, 这和其他方法探讨在不同多样性水平下生态系统能维持的功能数或整体功能水平是不同的。其最大的优点是,能揭示不同的物种是否驱动不同的生态系统功能, 并能检测每个物种对不同功能的重要性。同时, 该方法还能对某一生态系统中维持特定数量的功能所需的平均物种数进行定量预测(Hector & Bagchi, 2007 )。但是, 该方法同样存在缺点: (1)未直接测定多功能性, 故不能用多功能性指数来定量估计多样性改变对多功能性的影响。(2)需要大量数据支持, 且对数据有较高的要求; 分析每个物种对各个功能的作用, 需要一系列的多样性梯度, 且每个梯度都要有很好的可重复性。(3)数据分析步骤较繁琐。(4)未考虑不同功能的权重。
2.3 平均值法
该方法简单直观且所得结果容易解释, 可以较明确地评估多样性的改变对生态系统多个功能的平均影响,以及多个功能的平均水平是否随多样性的增加而增加, 还能衡量群落同时维持多个生态系统功能的能力(Maestre et al, 2012b )。但其缺点也很明显: (1)该方法不能区分一个物种对不同功能重要性的差异(Maestre et al, 2012b ), 且无法分辨多样性对单个功能的影响, 这将不能真实地反映生态系统的基本情况。(2)虽然考虑了不同功能的权重, 但功能间的权重不易衡量。多数情况下, 研究者将不同功能视为相同的权重, 而事实上某个功能可能会对最终的结果产生很大的影响。因不同生态系统中的功能权重不尽相同, 即使确定了某一系统各功能的权重, 也难以将之推广。(3)难以区分由相似水平的几个功能所得的多功能性指数与部分较高水平和部分较低水平的几个功能平均所得的多功能性指数(Dooley et al, 2015 )。(4)平均值法在计算时默认一种功能的降低可以由另一种功能的上升来弥补, 而在一个生态系统中, 不同的功能并非都可以互相替代, 即某一功能的减弱并不一定能由另一功能的增加来补偿(Gamfeldt et al, 2008 ; Maestre et al, 2012b )。
鉴于上述缺陷, 在用平均值法量化多功能性时, 可以结合单功能法建立各个功能与多样性之间的关系。若条件允许, 再结合物种替代法分析每个物种对不同功能的作用, 可以弥补该方法的一些 不足。
2.4 单阈值法
该方法可以评估达到某一阈值的功能数是否随多样性的增加而增加。其优点是较灵活, 因为测定的是超过某一水平的功能数量, 故而多样性与多功能性的关系不管是线性还是非线性都不影响测定的结果, 即使功能间存在权衡、交互作用等问题, 也能很好地获取达到阈值的功能数, 并且适用范围广(Byrnes et al, 2014a )。但是单阈值法也有缺点: (1)阈值的选择较随意, 研究者往往会选择有利于实验结果表述的阈值, 而不同的阈值可能得到截然不同的结果。因此, 没有恰当的理由时, 单个阈值的结果往往让人难以信服。(2)对每个功能的测度存在不足, 不能反映超过阈值或低于阈值的程度, 只能体现生态系统的整体功能达到了哪个水平(Gamfeldt et al, 2008 )。(3)因缺少统一的标准, 在实际应用中很难准确地判断哪个阈值最能反映多样性与多功能性的关系(Gamfeldt et al, 2008 ), 只检验单个阈值可能会丢失一些重要的阈值, 如多样性作用最强时的阈值(Byrnes et al, 2014a )。(4)不能测定多样性对单个生态系统功能的影响。(5)未考虑功能之间的 相关性(Dooley et al, 2015 )。(6)未考虑不同功能的权重。
在应用单阈值法时, 为了尽量避免遗漏某些关键的阈值信息, Zavaleta等(2010) 在0-100%的范围内选择了9个阈值: 10%、20%、30%、40%、50%、60%、70%、80%、90%。这种阈值选择方式虽然没有多阈值法所选择的阈值全面, 但是也覆盖了从低到高的各个水平, 可以获得较多的信息, 增强对单个功能超过或低于某阈值的程度的度量。但是, 对于一些较小的阈值(如低于最大值30%), 几乎任何多样性水平的群落都能实现所测定的功能, 因此, 过小的阈值显得有些赘余; 相反, 可以在多样性与多功能性的关系发生变化的拐点处多设置几个阈值, 以确定最接近生态系统发生状态转变的阈值水平。同时, 在用该方法测定多功能性时, 可以结合单功能法分别建立各个功能与多样性之间的关系, 从不同的角度进行分析, 从而找出多样性和多功能性之间的潜在关系。
2.5 多阈值法
多阈值法在本质上与单阈值法是同一种方法, 但它将所有的阈值都进行了计算, 因此比单阈值法提供的信息更多, 不仅拥有单阈值法的优点, 并且比单阈值法更灵活、全面(Byrnes et al, 2014a ), 用该方法测得的多功能性水平可以在不同实验间进行直接比较。因为设置的阈值为0-100%, 因此不必考虑阈值的选择, 可以在各个阈值水平检验多样性对多功能性的影响。但其存在4个明显的缺点: (1)提供了一系列的测度指标, 相对而言比较繁琐(Byrnes et al, 2014a );(2)不能测定多样性对单个生态系统功能的影响;(3)未考虑功能之间的相关性(Dooley et al, 2015 );(4)未考虑不同功能的权重。
同样, 在采用该方法测定多功能性时可以结合单功能法, 这样有助于找出多样性和多功能性之间的潜在关系。
2.6 直系同源基因法
直系同源基因法本质上衡量的是群落中物种所拥有的功能基因的多样性, 而不是测度该群落真正能够行使的功能。该方法是分辨不同微生物功能的有效方法, 且不会低估微生物的功能, 可以定量估计微生物群落多样性的改变对生态系统功能的影响。同时, 直系同源基因法还能反映进化多样性, 从进化的角度与生物多样性进行了结合(Faith, 1992 ; Hendry et al, 2010 )。此外, 由于能从许多研究地点获取直系同源基因丰富度的数据, 因此可以在较大尺度上研究BEMF关系。
直系同源基因法的主要缺点如下: (1)由于将所有的直系同源基因计算在内, 没有考虑这些功能是否在微生物基因组数据库(MBGD)里被预测过, 因此对多功能性的预测能力较低, 若要提高预测能力, 需要更详细的分析(Miki et al, 2014 )。(2)微生物的功能除受物种基因影响外, 还可能受到种间相互作用和环境因素的影响, 单纯用直系同源基因丰富度来估计, 可能会高估多功能性(可以采用基于物种特征的功能多样性进行分析来弥补此缺点)。(3)通过直系同源基因不能判断对应功能的强弱。(4)对于微生物丰富度和直系同源基因变异较大的大尺度实验, 还需阐明群落直系同源基因作为生态系统多功能性指数的有效性(Miki et al, 2014 )。(5)适用范围小, 只能用于微生物群落多功能性的分析。
2.7 多元模型法
多元模型法是最新提出的一种检测BEMF关系的方法, 也是目前最能充分利用数据信息的方法, 它充分考虑了不同方面的多样性、功能间的权衡和相关性对多功能性的影响, 可以从多个角度获得较为全面的信息; 同时, 综合了单功能法、平均值法、物种替代法和单阈值法的优点, 实现了以往的方法没能达成的目标, 即在分析单个功能的同时量化生态系统多功能性。此外, 该方法的主要优点有(Dooley et al, 2015 ): (1)可以检测生态系统整体功能达到最大值时的物种组成和相对多度。(2)可以检测功能之间的权衡和相关性, 更好地解释多样性与多功能性的关系。(3)可以检测各物种及种间交互作用对各个生态系统功能的作用及相对重要性。(4)具有很强的预测能力, 如当物种丰富度相同时, 其他方法预测的多功能性是相同的, 但该方法根据不同的物种组成和相对多度可以识别出不同的多功能性。(5)该模型还可引入时间和空间变量来分析多功 能性。
尽管多元模型法有很多优点, 但其应用也有较大局限性, 主要为: (1)该方法需要一系列多样性梯度, 仅有单作和包含所有物种的混种这两个处理不适用该方法。(2)当物种数量较多时, 会产生很多的种间交互系数δ , 故而需要用一定的方法进行降维处理, 如根据物种所属功能群来划分δ (其他的方法可参考Kirwan et al, 2009 )。(3)目前只适合功能数较少(如3个功能)的研究, 在功能数较多时, 因分析时会产生大量的系数, 使得模型变得很复杂, 因而不适用这种方法(Dooley et al, 2015 )。此外, 该方法也同样没有考虑不同功能的权重问题。
2.8 不同方法的应用
根据对各方法原理和特点的分析, 研究者应根据研究问题的需要以及实验所得的数据选择合适的多功能性测度方法。事实上, 由于目前的每种方法都存在相应的缺点, 大多数分析方法会损失关键信息, 多元模型法虽然损失的信息较少, 但不适合功能数量较多时的情况, 而常规的多功能性研究往往功能数较多(Hector & Bagchi, 2007 ; Isbell et al, 2011 ; Jing et al, 2015 )。所以,仅靠一种方法很难对BEMF进行全面分析。
因此, 在多功能性量化方法有所突破之前, 在条件允许的情况下, 功能数较少时, 可采用多元模型法; 当功能数较多时, 研究者可根据不同方法的特点, 结合多种分析方法, 取长补短。例如, 在用平均值法量化多功能性时结合单功能法(Jing et al, 2015 ), 则既能得到多样性改变对生态系统多个功能的平均影响, 又可根据单功能法建立的每个功能和多样性之间的关系得到每个功能如何随着多样性的变化而变化, 从而有助于研究者发现BEMF的潜在关系。
本文介绍的前5种方法的多功能性数据的统计分析可以使用R中的“Multifunc”程序包。“Multifunc”是由Byrnes等(2014a)开发, 免费开源的程序包, 用于处理生物多样性与生态系统多功能性数据(http://github.com/jebyrnes/multifunc )。多元模型法使用的R代码和SAS代码可参考Dooley等(2015)的附加代码。
目前, BEMF研究正处于一个快速发展阶段, 越来越多的研究者投入到BEMF的研究中。然而, 与BEF相比, BEMF的研究数据仍相对缺乏。此外, 因不同研究者采用不同的方法来测度多功能性, 使得已有研究数据之间很难进行对比, 更加剧了这种情况。为此, 希望在大力开展BEMF实验的同时, 能开发一种简洁且具有兼顾单个功能和多功能性整体水平的测度方法, 以实现不同研究之间的整合分析, 从而促进BEMF研究的发展。
(责任编委: 蒋 林 责任编辑: 黄祥忠)
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[21]
Hendry AP Lohmann LG Conti E Cracraft J Crandall KA Faith DP Hauser C Joly CA Kogure K Larigauderie A Magallon S Moritz C Tillier S Zardoya R Prieur-Richard AH Walther BA Yahara T Donoghue MJ 2010 ) Evolutionary biology in biodiversity science, conservation, and policy: a call to action.
Evolution , 64 , 1517 -1528 .
[本文引用: 1]
[22]
Hooper DU Vitousek PM 1998 ) Effects of plant composition and diversity on nutrient cycling.
Ecological Monographs , 68 , 121 -149 .
[本文引用: 1]
[23]
Isbell F Calcagno V Hector A Connolly J Harpole WS Reich PB Scherer-Lorenzen M Schmid B Tilman D van Ruijven J Weigelt A Wilsey BJ Zavaleta ES Loreau M 2011 ) High plant diversity is needed to maintain ecosystem services.
Nature , 477 , 199 -202 .
[本文引用: 3]
[24]
Jing X Sanders NJ Shi Y Chu HY Classen AT Zhao K Chen LT Shi Y Jiang YX He JS 2015 ) The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate.
Nature Communications , 6 , doi:10.1038/ncomms9159 .
[本文引用: 7]
[25]
Kirwan L Connolly J Finn JA Brophy C Luscher A Nyfeler D Sebastia MT 2009 ) Diversity-interaction modeling: estimating contributions of species identities and interactions to ecosystem function.
Ecology , 90 , 2032 -2038 .
[本文引用: 3]
[26]
Lefcheck JS Byrnes JEK Isbell F Gamfeldt L Griffin JN Eisenhauer N Hensel MJS Hector A Cardinale BJ Duffy JE 2015 ) Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.
Nature Communications , 6 , 6936 .
[本文引用: 3]
[27]
Loreau M Hector A 2001 ) Partitioning selection and complementarity in biodiversity experiments.
Nature , 412 , 72 -76 .
[本文引用: 2]
[28]
Lundholm JT 2015 ) Green roof plant species diversity improves ecosystem multifunctionality.
Journal of Applied Ecology , 52 , 726 -734 .
[29]
Maestre FT Castillo-Monroy AP Bowker MA Ochoa-Hueso R 2012 a) Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.
Journal of Ecology , 100 , 317 -330 .
[30]
Maestre FT Quero JL Gotelli NJ Escudero A Ochoa V Delgado-Baquerizo M Garcia-Gomez M Bowker MA Soliveres S Escolar C Garcia-Palacios P Berdugo M Valencia E Gozalo B Gallardo A Aguilera L Arredondo T Blones J Boeken B Bran D Conceicao AA Cabrera O Chaieb M Derak M Eldridge DJ Espinosa CI Florentino A Gaitan J Gatica MG Ghiloufi W Gomez-Gonzalez S Gutierrez JR Hernandez RM Huang X Huber-Sannwald E Jankju M Miriti M Monerris J Mau RL Morici E Naseri K Ospina A Polo V Prina A Pucheta E Ramirez- Collantes DA Romao R Tighe M Torres-Diaz C Val J Veiga JP Wang D Zaady E 2012 b) Plant species richness and ecosystem multifunctionality in global drylands.
Science , 335 , 214 -218 .
[31]
Ma KP 1994 ) Measurement of biotic community diversity. I. α diversity (part 1).
Chinese Biodiversity , 2 , 162 -168 . (in Chinese)
[本文引用: 1]
[马克平 (1994 ) 生物群落多样性的测度方法 I. α多样性的测度方法(上)
. 生物多样性 , 2 , 162 -168 .]
[本文引用: 1]
[32]
Miki T Yokokawa T Matsui K 2014 ) Biodiversity and multifunctionality in a microbial community: a novel theoretical approach to quantify functional redundancy.
Proceedings of the Royal Society B: Biological Sciences , 281 , 1 -9 .
[本文引用: 5]
[33]
Mooney HA Cropper A Reid W 2004 ) The Millennium Ecosystem Assessment: what is it all about?
Trends in Ecology and Evolution , 19 , 221 -224 .
[34]
Mouillot D Villéger S Scherer-Lorenzen M Mason NWH 2011 ) Functional structure of biological cmmunities predicts ecosystem multifunctionality.
PLoS ONE , 6 , e17476 .
[本文引用: 1]
[35]
Pasari JR Levi T Zavaleta ES Tilman D 2013 ) Several scales of biodiversity affect ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
USA , 110 , 10219 -10222 .
[36]
Pendleton RM Hoeinghaus DJ Gomes LC Agostinho AA 2014 ) Loss of rare fish species from tropical floodplain food webs affects community structure and ecosystem multifunctionality in a mesocosm experiment.
PLoS ONE , 9 , e84568 .
[37]
Perkins DM Bailey RA Dossena M Gamfeldt L Reiss J Trimmer M Woodward G 2015 ) Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes.
Global Change Biology , 21 , 396 -406 .
[本文引用: 2]
[38]
Peter H Ylla I Gudasz C Romaní AM Sabater S Tranvik LJ 2011 ) Multifunctionality and diversity in bacterial biofilms.
PLoS ONE , 6 , e23225 .
[39]
Quero JL Maestre FT Ochoa V García-Gómez M Delgado- Baquerizo M 2013 ) On the importance of shrub encroachment by sprouters, climate, species richness and anthropic factors for ecosystem multifunctionality in semi-arid mediterranean ecosystems.
Ecosystems , 16 , 1248 -1261 .
[40]
Soliveres S Maestre FT Eldridge DJ Delgado-Baquerizo M Quero JL Bowker MA Gallardo A 2014 ) Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands.
Global Ecology and Biogeography , 23 , 1408 -1416 .
[41]
Spehn EM Hector A Joshi J Scherer-Lorenzen M Schmid B Bazeley-White E Beierkuhnlein C Caldeira MC Diemer M Dimitrakopoulos PG Finn JA Freitas H Giller PS Good J Harris R Högberg P Huss-Danell K Jumpponen A Koricheva J Leadley PW Loreau M Minns A Mulder CPH O’Donovan G Otway SJ Palmborg C Pereira JS Pfisterer AB Prinz A Read DJ Schulze ED Siamantziouras ASD Terry AC Troumbis AY Woodward FI Yachi S Lawton JH 2005 ) Ecosystem effects of biodiversity manipulations in European grasslands.
Ecological Monographs , 75 , 37 -63 .
[本文引用: 1]
[42]
Tilman D Knops J Wedin D Reich P Ritchie M Siemann E 1997 a) The influence of functional diversity and composition on ecosystem processes.
Science , 277 , 1300 -1302 .
[43]
Tilman D Lehman CL Thomson KT 1997 b) Plant diversity and ecosystem productivity: theoretical considerations. Proceedings of the National Academy of Sciences,
USA , 94 , 1857 .
[44]
Uchiyama I Higuchi T Kawai M 2010 ) MBGD update 2010: toward a comprehensive resource for exploring microbial genome diversity.
Nucleic Acids Research , 38 , 361 -365 .
[本文引用: 1]
[45]
Valencia E Maestre FT Le Bagousse-Pinguet Y Quero JL Tamme R Börger L García-Gómez M Gross N 2015 ) Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands.
New Phytologist , 206 , 660 -671 .
[46]
van der Heijden MGA Martin FM Selosse M Sanders IR 2015 ) Mycorrhizal ecology and evolution: the past, the present, and the future.
New Phytologist , 205 , 1406 -1423 .
[47]
Wagg C Bender SF Widmer F van der Heijden MGA 2014 ) Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
USA , 111 , 5266 -5270 .
[本文引用: 1]
[48]
Xu W Ma ZY Jing X He JS 2016 ) Biodiversity and ecosystem multifunctionality: status and perspectives.
Biodiversity Science , 24 , 55 -71 . (in Chinese with English abstract)
[本文引用: 1]
[徐炜 , 马志远 , 井新 , 贺金生 (2016 ) 生物多样性与生态系统多功能性: 进展与展望
. 生物多样性 , 24 , 55 -71 .]
[本文引用: 1]
[49]
Zavaleta ES Pasari JR Hulvey KB Tilman GD 2010 ) Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences,
USA , 107 , 1443 -1446 .
[本文引用: 7]
[1]
Bowker MA Maestre FT Mau RL 2013 ) Diversity and patch-size distributions of biological soil crusts regulate dryland ecosystem multifunctionality.
Ecosystems , 16 , 923 -933 .
[2]
Bradford MA Wood SA Bardgett RD Black HIJ Bonkowski M Eggers T Grayston SJ Kandeler E Manning P Setälä H Jones TH 2014 a) Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proceedings of the National Academy of Sciences,
USA , 111 , 14478 -14483 .
[3]
Bradford MA Wood SA Bardgett RD Black HIJ Bonkowski M Eggers T Grayston SJ Kandeler E Manning P Setälä H Jones TH 2014 b) Reply to Byrnes et al.: Aggregation can obscure understanding of ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
USA , 111 , E5491 .
[4]
Byrnes JEK Gamfeldt L Isbell F Lefcheck JS Griffin JN Hector A Cardinale BJ Hooper DU Dee LE Emmett Duffy J 2014 a) Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions.
Methods in Ecology and Evolution , 5 , 111 -124 .
[5]
Byrnes JEK Lefcheck JS Gamfeldt L Griffin JN Isbell F Hector A 2014 b) Multifunctionality does not imply that all functions are positively correlated. Proceedings of the National Academy of Sciences,
USA , 111 , E5490 .
[6]
Cardinale BJ Wright JP Cadotte MW Carroll IT Hector A Srivastava DS Loreau M Weis JJ 2007 ) Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences,
USA , 104 , 18123 -18128 .
[本文引用: 1]
[7]
Cardinale BJ 2011 ) Biodiversity improves water quality through niche partitioning.
Nature , 472 , 86 -89 .
[8]
Cardinale BJ Srivastava DS Emmett Duffy J Wright JP Downing AL Sankaran M Jouseau C 2006 ) Effects of biodiversity on the functioning of trophic groups and ecosystems.
Nature , 443 , 989 -992 .
[9]
Constán-Nava S Soliveres S Torices R Serra L Bonet A 2015 ) Direct and indirect effects of invasion by the alien tree Ailanthus altissima on riparian plant communities and ecosystem multifunctionality.
Biological Invasions , 17 , 1095 -1108 .
[10]
Dooley A Isbell F Kirwan L Connolly J Finn JA Brophy C 2015 ) Testing the effects of diversity on ecosystem multifunctionality using a multivariate model.
Ecology Letters , 18 , 1242 -1251 .
[本文引用: 12]
[11]
Downing AL 2005 ) Relative effects of species composition and richness on ecosystem properties in ponds.
Ecology , 86 , 701 -715 .
[本文引用: 1]
[12]
Duffy JE Macdonald KS Rhode JM Parker JD 2001 ) Grazer diversity, functional redundancy, and productivity in seagrass beds: an experimental test.
Ecology , 82 , 2417 -2434 .
[本文引用: 1]
[13]
Duffy JE Richardson JP Canuel EA 2003 ) Grazer diversity effects on ecosystem functioning in seagrass beds.
Ecology Letters , 6 , 637 -645 .
[本文引用: 1]
[14]
Faith DP 1992 ) Conservation evaluation and phylogenetic diversity.
Biological Conservation , 61 , 1 -10 .
[本文引用: 1]
[15]
Fang G Bhardwaj N Robilotto R Gerstein MB 2010 ) Getting started in gene orthology and functional analysis.
PLoS Computational Biology , 6 , e1000703 .
[本文引用: 1]
[16]
Fargione J Tilman D Dybzinski R Lambers JHR Clark C Harpole WS Knops JMH Reich PB Loreau M 2007 ) From selection to complementarity: shifts in the causes of biodiversity-productivity relationships in a long-term biodiversity experiment.
Proceedings of the Royal Society B: Biological Sciences , 274 , 871 -876 .
[本文引用: 1]
[17]
Gamfeldt L Hillebrand H Jonsson PR 2008 ) Multiple functions increase the importance of biodiversity for overall ecosystem functioning.
Ecology , 89 , 1223 .
[本文引用: 6]
[18]
Gamfeldt L Snäll T Bagchi R Jonsson M Gustafsson L Kjellander P Ruiz-Jaen MC Fröberg M Stendahl J Philipson CD Mikusiński G Andersson E Westerlund B Andrén H Moberg F Moen J Bengtsson J 2013 ) Higher levels of multiple ecosystem services are found in forests with more tree species.
Nature Communications , 4 , doi: 10.1038/ncomms2328 .
[本文引用: 1]
[19]
He J Ge Y Xu Z Chen C 2009 ) Linking soil bacterial diversity to ecosystem multifunctionality using backward- elimination boosted trees analysis.
Journal of Soils and Sediments , 9 , 547 -554 .
[本文引用: 1]
[20]
Hector A Bagchi R 2007 ) Biodiversity and ecosystem multifunctionality.
Nature , 448 , 188 -190 .
[本文引用: 10]
Evolutionary biology in biodiversity science, conservation, and policy: a call to action.
1
2010
... 直系同源基因法本质上衡量的是群落中物种所拥有的功能基因的多样性, 而不是测度该群落真正能够行使的功能.该方法是分辨不同微生物功能的有效方法, 且不会低估微生物的功能, 可以定量估计微生物群落多样性的改变对生态系统功能的影响.同时, 直系同源基因法还能反映进化多样性, 从进化的角度与生物多样性进行了结合(Faith, 1992 ; Hendry et al, 2010 ).此外, 由于能从许多研究地点获取直系同源基因丰富度的数据, 因此可以在较大尺度上研究BEMF关系. ...
Effects of plant composition and diversity on nutrient cycling.
1
1998
... 式中, F 表示测定的功能数, fi 表示功能i 的测定值, ri 是将fi 转换成正值的数学函数, g 表示将所有的测定值标准化.由于有些功能值不能直接反映生态系统的功能, 比如土壤N含量表征的是植物N吸收的能力, 一般情况下, N含量越低, 植物N吸收能力越高.因此, ri (fi ) 可以采用max(fi ) - fi 的形式表示, max(fi )为前5%的观测值的平均值, 所得结果越大, 表明植物N吸收能力越强.在求平均值之前, 所有的功能值要先标准化, 以保证不同功能是在同一尺度上进行相互比较.虽然功能值标准化的方法有很多, 但最终的结果都较为相近(Maestre et al, 2012b ).这里推荐最常用的两种标准化方法.一是Z得分(Z Scores) (Mouillot et al, 2011 ; Maestre et al, 2012b ; Bradford et al, 2014a ; Wagg et al, 2014 ): 即用平均值和标准偏差来进行标准化(测定值减去平均值后除以标准偏差); 二是用最大值转化(Hooper & Vitousek, 1998 ; Maestre et al, 2012a ): 每个功能排名前5%的测定值取平均值后作为该功能的最大值, 然后计算每个测定值与最大值的比值. ...
High plant diversity is needed to maintain ecosystem services.
3
2011
... 根据上式可以定量预测一个生态系统中, 随着所考虑的生态系统功能数的增加, 维持这些功能所需的平均物种数.当对所考虑的功能与维持这些功能所需的物种数进行回归时, 有2种极端的情况: (1)如果一个物种只影响一个功能( ) 之间存在正向线性关系, 其斜率为1 (Hector & Bagchi, 2007 ); (2)如果生态系统所有的功能都受同一物种集合的影响( Hector & Bagchi, 2007 ).在实际研究中, 结果往往介于两者之间, 表明物种和功能间存在一定的替代(Hector & Bagchi, 2007 ; Isbell et al, 2011 ). ...
... 然而, 因为物种库中的物种对不同的功能有不同的作用, 而多样性对多功能性的影响取决于正负效应之间的权衡.若正效应大于负效应, 则随着多样性的增加, 多功能性将增加; 反之则降低.Hector和Bagchi (2007)的方法目前仅被用于各物种对生态系统功能具有正效应的情况(He et al, 2009 ; Isbell et al, 2011 ), 而在很多情况下并非如此.当将负效应考虑进来时, 就可以通过检验物种正效应是否强于、等于或者弱于负效应以及是否有物种主导整体功能的正负效应, 来更好地揭示BEMF的潜在关系.为此, Byrnes等(2014a)提出了2个额外的分析步骤来完善该方法: ...
... 根据对各方法原理和特点的分析, 研究者应根据研究问题的需要以及实验所得的数据选择合适的多功能性测度方法.事实上, 由于目前的每种方法都存在相应的缺点, 大多数分析方法会损失关键信息, 多元模型法虽然损失的信息较少, 但不适合功能数量较多时的情况, 而常规的多功能性研究往往功能数较多(Hector & Bagchi, 2007 ; Isbell et al, 2011 ; Jing et al, 2015 ).所以,仅靠一种方法很难对BEMF进行全面分析. ...
The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate.
7
2015
... (1) The journals used in this survey include the top journals such as Nature , Science and Nature Communications ; (2) Bradford et al (2014a) , Lefcheck et al (2015) and Jing et al (2015) all used three metrics to qualify EMF; (3) Although Byrnes et al (2014a) used five metrics, the authors’ purpose was to propose the multiple-threshold approach. Thus, this paper is only listed in the multiple-threshold approach; (4) Review articles are here excluded. ...
... 上述提及的EMF的测度方法各有其优缺点, 每种方法的侧重方面也不尽相同.研究者常常根据自身研究的需要, 使用相对合适的方法.有的研究者(如Bradford et al, 2014a ; Jing et al, 2015 ; Lefcheck et al, 2015 )则选择了1种以上的测度方法, 这样不同方法可以互相补充.这虽然有助于更好地研究生物多样性对EMF的潜在影响, 但是也突显了不同方法之间存在的缺陷, 不利于研究间的对比.因此, 未来研究的一个重要任务是开发一种具有普遍适用性的EMF量化方法(Bradford et al, 2014b ; Byrnes et al, 2014b ), 从而实现不同研究之间的整合分析, 促进BEMF研究的发展. ...
... 22年前, 《生物多样性》对生物群落多样性测度方法的介绍(马克平, 1994 ),在一定程度上促进了国内对生物多样性的研究.在此启发下, 本文简述了7种EMF量化方法的基本原理及其特点, 以帮助研究者选择合适的测度方法.对于其中较难理解的多阈值法, 将利用我们所收集的青藏高原样带数据(Jing et al, 2015 )说明其在BEMF研究中的应用. ...
... 为了便于读者理解, 本文根据青藏高原样带数据(Jing et al, 2015 )进行说明.通过检验不同阈值下丛枝菌根真菌多样性与多功能性指数MFt 拟合曲线的斜率变化(图1 )来检测多样性对多功能性的影响.将阈值与对应的拟合曲线的斜率建立关系(图2 ), 可以得到多样性如何影响多功能性的大量信息, 而不仅仅局限于何时多样性的作用最大(即斜率最大). ...
... 该方法单独使用时所得结果不易让人信服, 但是可以作为辅助方法, 与其他测度方法(如平均值法、单阈值法等)一起使用(Jing et al, 2015 ). ...
... 根据对各方法原理和特点的分析, 研究者应根据研究问题的需要以及实验所得的数据选择合适的多功能性测度方法.事实上, 由于目前的每种方法都存在相应的缺点, 大多数分析方法会损失关键信息, 多元模型法虽然损失的信息较少, 但不适合功能数量较多时的情况, 而常规的多功能性研究往往功能数较多(Hector & Bagchi, 2007 ; Isbell et al, 2011 ; Jing et al, 2015 ).所以,仅靠一种方法很难对BEMF进行全面分析. ...
... 因此, 在多功能性量化方法有所突破之前, 在条件允许的情况下, 功能数较少时, 可采用多元模型法; 当功能数较多时, 研究者可根据不同方法的特点, 结合多种分析方法, 取长补短.例如, 在用平均值法量化多功能性时结合单功能法(Jing et al, 2015 ), 则既能得到多样性改变对生态系统多个功能的平均影响, 又可根据单功能法建立的每个功能和多样性之间的关系得到每个功能如何随着多样性的变化而变化, 从而有助于研究者发现BEMF的潜在关系. ...
Diversity-interaction modeling: estimating contributions of species identities and interactions to ecosystem function.
3
2009
... 以上介绍的6种方法, 在分析过程中或多或少都会丢失一些重要的信息, 导致对EMF的分析不够全面.为了尽可能全面地了解BEMF的关系, Dooley等(2015)提出了以BEF研究中使用的多样性交互模型(Kirwan et al, 2009 )为基础扩展而成的多元模型法.该方法克服了上述方法仅将物种丰富度作为多样性的缺点, 在分析过程中考虑了不同方面的多样性(如物种相对多度、均匀度、物种组成等)对EMF的影响.同时, 该方法还可以分析功能之间的权衡和相关性, 并能检验整体功能在何时达到最大值. ...
... 式中, yk 表示功能k , βik 表示物种i 在单作时, 功能k 的预期表现, δijk 表示物种i 和j 对功能k 的交互作用(Dooley et al, 2015 ).为了减少模型系数, 使整个模型更为简洁, 通过一些有生物学意义的假设, 可以将表示物种间交互作用的系数δijk 进行简化(Kirwan et al, 2009 ).例如,假设功能群决定物种间的交互作用, 以2个功能群为例, 当物种i , j 都来自功能群1 (功能群2)时, δijk = δwfg1k (δwfg2k ) ; 当i , j 来自不同功能群时, δijk = δbfgk ; 此处wfg 表示属于同一个功能群, bfg 表示2个不同的功能群(Dooley et al, 2015 ). ...
... 尽管多元模型法有很多优点, 但其应用也有较大局限性, 主要为: (1)该方法需要一系列多样性梯度, 仅有单作和包含所有物种的混种这两个处理不适用该方法.(2)当物种数量较多时, 会产生很多的种间交互系数δ , 故而需要用一定的方法进行降维处理, 如根据物种所属功能群来划分δ (其他的方法可参考Kirwan et al, 2009 ).(3)目前只适合功能数较少(如3个功能)的研究, 在功能数较多时, 因分析时会产生大量的系数, 使得模型变得很复杂, 因而不适用这种方法(Dooley et al, 2015 ).此外, 该方法也同样没有考虑不同功能的权重问题. ...
Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.
3
2015
... (1) The journals used in this survey include the top journals such as Nature , Science and Nature Communications ; (2) Bradford et al (2014a) , Lefcheck et al (2015) and Jing et al (2015) all used three metrics to qualify EMF; (3) Although Byrnes et al (2014a) used five metrics, the authors’ purpose was to propose the multiple-threshold approach. Thus, this paper is only listed in the multiple-threshold approach; (4) Review articles are here excluded. ...
... (1)重要刊物指Nature 、Science 、Nature Communications 等综合性的国际重要期刊; (2) Bradford等(2014a) , Lefcheck等(2015) 以及Jing等(2015)这3篇文章每篇都使用了3种EMF量化方法; (3)尽管Byrnes等(2014a)用了5种方法, 但其目的是为了引出多阈值法, 故只将其列入多阈值法的文献中; (4)表中所统计的文献不包括评论文章. ...
... 上述提及的EMF的测度方法各有其优缺点, 每种方法的侧重方面也不尽相同.研究者常常根据自身研究的需要, 使用相对合适的方法.有的研究者(如Bradford et al, 2014a ; Jing et al, 2015 ; Lefcheck et al, 2015 )则选择了1种以上的测度方法, 这样不同方法可以互相补充.这虽然有助于更好地研究生物多样性对EMF的潜在影响, 但是也突显了不同方法之间存在的缺陷, 不利于研究间的对比.因此, 未来研究的一个重要任务是开发一种具有普遍适用性的EMF量化方法(Bradford et al, 2014b ; Byrnes et al, 2014b ), 从而实现不同研究之间的整合分析, 促进BEMF研究的发展. ...
Partitioning selection and complementarity in biodiversity experiments.
2
2001
... 该方法的结果易受到抽样效应(sampling effect)和互补效应(complementarity)的共同影响(Tilman et al, 1997a ; Loreau & Hector, 2001 ), 其中互补效应起主要作用(详见Loreau & Hector, 2001 ; Cardinale et al, 2007 ; Fargione et al, 2007 ).所谓抽样效应是一个统计学的概念, 指在同一生境中, 竞争力强的物种对资源的利用率更高, 故而有更高的生产力, 随着物种丰富度的增加, 从物种库中抽到高产物种的概率也将增大(Tilman et al, 1997b ); 所以在随机抽样的情况下, 高多样性的群落包含高产物种的概率更大.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 从物种库中得到对某几个功能或所有功能都有很强促进作用的物种的概率会增大, 从而得出物种丰富度增加会提高多功能性的结论.互补效应则是指在一个群落中, 物种间存在生态位差异, 故而群落中物种数越多, 所占据的“功能空间” (functional space)范围越广(Tilman et al, 1997b ).因此, 生态系统的物种丰富度越高, 对资源的利用率就越高, 相应的生产力也会越高.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 物种间的生态位差异增大, 不同物种支持不同的功能, 同样会得到因物种丰富度增加而多功能性提高的结论. ...
... ), 其中互补效应起主要作用(详见Loreau & Hector, 2001 ; Cardinale et al, 2007 ; Fargione et al, 2007 ).所谓抽样效应是一个统计学的概念, 指在同一生境中, 竞争力强的物种对资源的利用率更高, 故而有更高的生产力, 随着物种丰富度的增加, 从物种库中抽到高产物种的概率也将增大(Tilman et al, 1997b ); 所以在随机抽样的情况下, 高多样性的群落包含高产物种的概率更大.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 从物种库中得到对某几个功能或所有功能都有很强促进作用的物种的概率会增大, 从而得出物种丰富度增加会提高多功能性的结论.互补效应则是指在一个群落中, 物种间存在生态位差异, 故而群落中物种数越多, 所占据的“功能空间” (functional space)范围越广(Tilman et al, 1997b ).因此, 生态系统的物种丰富度越高, 对资源的利用率就越高, 相应的生产力也会越高.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 物种间的生态位差异增大, 不同物种支持不同的功能, 同样会得到因物种丰富度增加而多功能性提高的结论. ...
Green roof plant species diversity improves ecosystem multifunctionality.
2015
a) Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.
2012
b) Plant species richness and ecosystem multifunctionality in global drylands.
2012
生物群落多样性的测度方法 I. α多样性的测度方法(上)
1
1994
... 22年前, 《生物多样性》对生物群落多样性测度方法的介绍(马克平, 1994 ),在一定程度上促进了国内对生物多样性的研究.在此启发下, 本文简述了7种EMF量化方法的基本原理及其特点, 以帮助研究者选择合适的测度方法.对于其中较难理解的多阈值法, 将利用我们所收集的青藏高原样带数据(Jing et al, 2015 )说明其在BEMF研究中的应用. ...
生物群落多样性的测度方法 I. α多样性的测度方法(上)
1
1994
... 22年前, 《生物多样性》对生物群落多样性测度方法的介绍(马克平, 1994 ),在一定程度上促进了国内对生物多样性的研究.在此启发下, 本文简述了7种EMF量化方法的基本原理及其特点, 以帮助研究者选择合适的测度方法.对于其中较难理解的多阈值法, 将利用我们所收集的青藏高原样带数据(Jing et al, 2015 )说明其在BEMF研究中的应用. ...
Biodiversity and multifunctionality in a microbial community: a novel theoretical approach to quantify functional redundancy.
5
2014
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
... 直系同源基因是指不同物种中来自共同祖先的基因, 通常认为其功能保守, 因此具有相似的功能(Fang et al, 2010 ).该方法由Miki等(2014)提出, 用于测度微生物群落的多功能性, 直系同源基因丰富度越高则群落的多功能性越高.Miki等(2014)先在理论上进行探讨, 认为不同物种所拥有的同类直系同源基因具有相似的功能, 不同类别的直系同源基因具有不同的功能, 因此通过评估微生物群落中直系同源基因的丰富度就能知道群落功能的大致范围.之后, 他们通过微宇宙实验证明, 利用群落中微生物的直系同源基因序列丰富度可以预测实际的多功能性(Miki et al, 2014 ).用该方法测度多功能性时, 最好选择系统发生相近的微生物类群. ...
... 此方法目前应用不多, 仅有一篇文章(Miki etal, 2014 )采用了该方法.尽管如此, 从开发新的多功能性量化方法的角度来看, 这种方法从基因的角 度出发, 对量化微生物多功能性进行了有益的探 索, 对未来建立具有普遍适用性的量化方法有启发作用. ...
... 直系同源基因法的主要缺点如下: (1)由于将所有的直系同源基因计算在内, 没有考虑这些功能是否在微生物基因组数据库(MBGD)里被预测过, 因此对多功能性的预测能力较低, 若要提高预测能力, 需要更详细的分析(Miki et al, 2014 ).(2)微生物的功能除受物种基因影响外, 还可能受到种间相互作用和环境因素的影响, 单纯用直系同源基因丰富度来估计, 可能会高估多功能性(可以采用基于物种特征的功能多样性进行分析来弥补此缺点).(3)通过直系同源基因不能判断对应功能的强弱.(4)对于微生物丰富度和直系同源基因变异较大的大尺度实验, 还需阐明群落直系同源基因作为生态系统多功能性指数的有效性(Miki et al, 2014 ).(5)适用范围小, 只能用于微生物群落多功能性的分析. ...
... ).(2)微生物的功能除受物种基因影响外, 还可能受到种间相互作用和环境因素的影响, 单纯用直系同源基因丰富度来估计, 可能会高估多功能性(可以采用基于物种特征的功能多样性进行分析来弥补此缺点).(3)通过直系同源基因不能判断对应功能的强弱.(4)对于微生物丰富度和直系同源基因变异较大的大尺度实验, 还需阐明群落直系同源基因作为生态系统多功能性指数的有效性(Miki et al, 2014 ).(5)适用范围小, 只能用于微生物群落多功能性的分析. ...
The Millennium Ecosystem Assessment: what is it all about?
2004
Functional structure of biological cmmunities predicts ecosystem multifunctionality.
1
2011
... 式中, F 表示测定的功能数, fi 表示功能i 的测定值, ri 是将fi 转换成正值的数学函数, g 表示将所有的测定值标准化.由于有些功能值不能直接反映生态系统的功能, 比如土壤N含量表征的是植物N吸收的能力, 一般情况下, N含量越低, 植物N吸收能力越高.因此, ri (fi ) 可以采用max(fi ) - fi 的形式表示, max(fi )为前5%的观测值的平均值, 所得结果越大, 表明植物N吸收能力越强.在求平均值之前, 所有的功能值要先标准化, 以保证不同功能是在同一尺度上进行相互比较.虽然功能值标准化的方法有很多, 但最终的结果都较为相近(Maestre et al, 2012b ).这里推荐最常用的两种标准化方法.一是Z得分(Z Scores) (Mouillot et al, 2011 ; Maestre et al, 2012b ; Bradford et al, 2014a ; Wagg et al, 2014 ): 即用平均值和标准偏差来进行标准化(测定值减去平均值后除以标准偏差); 二是用最大值转化(Hooper & Vitousek, 1998 ; Maestre et al, 2012a ): 每个功能排名前5%的测定值取平均值后作为该功能的最大值, 然后计算每个测定值与最大值的比值. ...
Several scales of biodiversity affect ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
2013
Loss of rare fish species from tropical floodplain food webs affects community structure and ecosystem multifunctionality in a mesocosm experiment.
2014
Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes.
2
2015
... 此方法的一个关键步骤是阈值的确定.阈值通常是指每个功能所观测到的最大值的比例(如25%、50%、75%等) (Zavaleta et al, 2010 ), 通常用最大值法来确定某一功能在所研究生态系统中的最高水平.实验中观测到的最大值因实验误差或其他原因可能是异常值, 为此, 研究者常取某一个功能前n + 1个观测值的平均值作为最大值(n 表示单个物种处理时的最小样本量) (Byrnes et al, 2014a ; Perkins et al, 2015 ), 或取观测值前5%的均值作为最大值(与平均值法中介绍的最大值转换类似) (Zavaleta et al, 2010 ).得到最大值之后, 下一步就是确定阈值ti , ti 由最大值乘以一定的百分比得到.研究者可根据研究目的自行选择某一百分比.但必须注意的是, 对于所选的百分比, 必须有合适的理由进行解释, 否则不同的阈值可能导致不同的结果, 所得的结果难以让人信服. ...
... 结合图1 和图2 的曲线, 可以获得一些关键的度量指标来帮助理解多样性与多功能性之间的关系.通过4个主要指标可以获得多样性如何影响多功能性的信息; 而借助4个辅助指标可以对不同的系统和实验进行对比(Byrnes et al, 2014a ; Perkins et al, 2015 ).4个主要指标为: ...
Multifunctionality and diversity in bacterial biofilms.
2011
On the importance of shrub encroachment by sprouters, climate, species richness and anthropic factors for ecosystem multifunctionality in semi-arid mediterranean ecosystems.
2013
Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands.
2014
Ecosystem effects of biodiversity manipulations in European grasslands.
1
2005
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
a) The influence of functional diversity and composition on ecosystem processes.
1997
b) Plant diversity and ecosystem productivity: theoretical considerations. Proceedings of the National Academy of Sciences,
1997
MBGD update 2010: toward a comprehensive resource for exploring microbial genome diversity.
1
2010
... 该方法的第一步是根据研究需要从微生物基因组数据库(microbial genome database, MBGD)中 (http://mbgd.genome.ad.jp/ )下载相关数据作为默认序列表(default orthologue table) (Uchiyama et al, 2010 ), 数据以碱基序列表征.因为一个物种可能会有很多具有完全相同序列的基因组, 因此需要将每个物种的序列簇表(orthologue cluster table)转换成二进制的矩阵, 以1或0表示某一序列存在与否, 1表示存在, 0表示不存在.转换后的表格就是简单的物种×序列矩阵, 可以方便地对测定的每种微生物所拥有的基因序列进行统计. ...
Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands.
2015
Mycorrhizal ecology and evolution: the past, the present, and the future.
2015
Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
1
2014
... 式中, F 表示测定的功能数, fi 表示功能i 的测定值, ri 是将fi 转换成正值的数学函数, g 表示将所有的测定值标准化.由于有些功能值不能直接反映生态系统的功能, 比如土壤N含量表征的是植物N吸收的能力, 一般情况下, N含量越低, 植物N吸收能力越高.因此, ri (fi ) 可以采用max(fi ) - fi 的形式表示, max(fi )为前5%的观测值的平均值, 所得结果越大, 表明植物N吸收能力越强.在求平均值之前, 所有的功能值要先标准化, 以保证不同功能是在同一尺度上进行相互比较.虽然功能值标准化的方法有很多, 但最终的结果都较为相近(Maestre et al, 2012b ).这里推荐最常用的两种标准化方法.一是Z得分(Z Scores) (Mouillot et al, 2011 ; Maestre et al, 2012b ; Bradford et al, 2014a ; Wagg et al, 2014 ): 即用平均值和标准偏差来进行标准化(测定值减去平均值后除以标准偏差); 二是用最大值转化(Hooper & Vitousek, 1998 ; Maestre et al, 2012a ): 每个功能排名前5%的测定值取平均值后作为该功能的最大值, 然后计算每个测定值与最大值的比值. ...
生物多样性与生态系统多功能性: 进展与展望
1
2016
... EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识.与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 ).单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
生物多样性与生态系统多功能性: 进展与展望
1
2016
... EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识.与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 ).单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences,
7
2010
... 传统的生物多样性与生态系统功能(biodiver- sity and ecosystem functioning, BEF)研究多关注植物物种丰富度和生态系统净初级生产力(NPP)之间的关系.显然, 单个生态系统功能指标(如NPP)并不能完全反映多样性对生态系统功能的影响(Hector & Bagchi, 2007 ; Gamfeldt et al, 2008 ; Zavaleta et al, 2010 ), 因为一个生态系统具有同时提供多种功能和服务的能力, 这被称为生态系统多功能性(ecosystem multifunctionality, EMF) (Hector & Bagchi, 2007 ). ...
... EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识.与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 ).单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
... ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
... 单阈值法的出现将多功能性的测度转向了另一个方向, 即评估随着多样性的增加达到某一阈值水平的功能数的变化.该方法通过计算每个生态系统中达到某一阈值的功能数来求得一个指数, 该指数表示在该阈值条件下这个生态系统整体功能的水平(Zavaleta et al, 2010 ). ...
... 此方法的一个关键步骤是阈值的确定.阈值通常是指每个功能所观测到的最大值的比例(如25%、50%、75%等) (Zavaleta et al, 2010 ), 通常用最大值法来确定某一功能在所研究生态系统中的最高水平.实验中观测到的最大值因实验误差或其他原因可能是异常值, 为此, 研究者常取某一个功能前n + 1个观测值的平均值作为最大值(n 表示单个物种处理时的最小样本量) (Byrnes et al, 2014a ; Perkins et al, 2015 ), 或取观测值前5%的均值作为最大值(与平均值法中介绍的最大值转换类似) (Zavaleta et al, 2010 ).得到最大值之后, 下一步就是确定阈值ti , ti 由最大值乘以一定的百分比得到.研究者可根据研究目的自行选择某一百分比.但必须注意的是, 对于所选的百分比, 必须有合适的理由进行解释, 否则不同的阈值可能导致不同的结果, 所得的结果难以让人信服. ...
... ), 或取观测值前5%的均值作为最大值(与平均值法中介绍的最大值转换类似) (Zavaleta et al, 2010 ).得到最大值之后, 下一步就是确定阈值ti , ti 由最大值乘以一定的百分比得到.研究者可根据研究目的自行选择某一百分比.但必须注意的是, 对于所选的百分比, 必须有合适的理由进行解释, 否则不同的阈值可能导致不同的结果, 所得的结果难以让人信服. ...
... 在应用单阈值法时, 为了尽量避免遗漏某些关键的阈值信息, Zavaleta等(2010) 在0-100%的范围内选择了9个阈值: 10%、20%、30%、40%、50%、60%、70%、80%、90%.这种阈值选择方式虽然没有多阈值法所选择的阈值全面, 但是也覆盖了从低到高的各个水平, 可以获得较多的信息, 增强对单个功能超过或低于某阈值的程度的度量.但是, 对于一些较小的阈值(如低于最大值30%), 几乎任何多样性水平的群落都能实现所测定的功能, 因此, 过小的阈值显得有些赘余; 相反, 可以在多样性与多功能性的关系发生变化的拐点处多设置几个阈值, 以确定最接近生态系统发生状态转变的阈值水平.同时, 在用该方法测定多功能性时, 可以结合单功能法分别建立各个功能与多样性之间的关系, 从不同的角度进行分析, 从而找出多样性和多功能性之间的潜在关系. ...
Diversity and patch-size distributions of biological soil crusts regulate dryland ecosystem multifunctionality.
2013
a) Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition. Proceedings of the National Academy of Sciences,
2014
b) Reply to Byrnes et al.: Aggregation can obscure understanding of ecosystem multifunctionality. Proceedings of the National Academy of Sciences,
2014
a) Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions.
2014
b) Multifunctionality does not imply that all functions are positively correlated. Proceedings of the National Academy of Sciences,
2014
Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences,
1
2007
... 该方法的结果易受到抽样效应(sampling effect)和互补效应(complementarity)的共同影响(Tilman et al, 1997a ; Loreau & Hector, 2001 ), 其中互补效应起主要作用(详见Loreau & Hector, 2001 ; Cardinale et al, 2007 ; Fargione et al, 2007 ).所谓抽样效应是一个统计学的概念, 指在同一生境中, 竞争力强的物种对资源的利用率更高, 故而有更高的生产力, 随着物种丰富度的增加, 从物种库中抽到高产物种的概率也将增大(Tilman et al, 1997b ); 所以在随机抽样的情况下, 高多样性的群落包含高产物种的概率更大.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 从物种库中得到对某几个功能或所有功能都有很强促进作用的物种的概率会增大, 从而得出物种丰富度增加会提高多功能性的结论.互补效应则是指在一个群落中, 物种间存在生态位差异, 故而群落中物种数越多, 所占据的“功能空间” (functional space)范围越广(Tilman et al, 1997b ).因此, 生态系统的物种丰富度越高, 对资源的利用率就越高, 相应的生产力也会越高.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 物种间的生态位差异增大, 不同物种支持不同的功能, 同样会得到因物种丰富度增加而多功能性提高的结论. ...
Biodiversity improves water quality through niche partitioning.
2011
Effects of biodiversity on the functioning of trophic groups and ecosystems.
2006
Direct and indirect effects of invasion by the alien tree Ailanthus altissima on riparian plant communities and ecosystem multifunctionality.
2015
Testing the effects of diversity on ecosystem multifunctionality using a multivariate model.
12
2015
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
... 式中, yk 表示功能k , βik 表示物种i 在单作时, 功能k 的预期表现, δijk 表示物种i 和j 对功能k 的交互作用(Dooley et al, 2015 ).为了减少模型系数, 使整个模型更为简洁, 通过一些有生物学意义的假设, 可以将表示物种间交互作用的系数δijk 进行简化(Kirwan et al, 2009 ).例如,假设功能群决定物种间的交互作用, 以2个功能群为例, 当物种i , j 都来自功能群1 (功能群2)时, δijk = δwfg1k (δwfg2k ) ; 当i , j 来自不同功能群时, δijk = δbfgk ; 此处wfg 表示属于同一个功能群, bfg 表示2个不同的功能群(Dooley et al, 2015 ). ...
... 表示2个不同的功能群(Dooley et al, 2015 ). ...
... 在确定了最优模型后, 先用t 检验对模型系数进行比较, 但在比较过程中要注意多次成对比较所带来的问题, 如比较3个功能e 、f 、k 所得模型的系数βie 、βif 和βik , 在比较时, 有3次成对t 检验, 这时可以用Bonferroni校验法来避免多次比较造成的误差(Dooley et al, 2015 ).之后, 根据不同物种相对多度、物种组成、均匀度及所设置的处理, 利用上述模型, 对各生态系统功能的表现进行预测, 并预先设置一个阈值(尽量取值大一些, 如70%)来与各功能值进行比较.根据对模型系数的分析和模型预测的各功能的表现, 可以从不同角度探讨BEMF的关系.讨论的内容主要包括两个方面: ...
... (1)单作时的多功能性.通过比较不同模型的系数β , 可以得到不同物种在单作时各功能表现的差异, 从而得到各物种对不同功能的相对重要性、不同功能表现的变异性、相关性以及功能之间的权衡(如单作时, 当某个功能值很高时, 其他的功能处于相对较低水平, 说明这个功能与其他功能之间存在权衡) (Dooley et al, 2015 ). ...
... (2)混种时的多功能性.通过比较不同模型的系数δ , 可以得到物种间的交互作用对各功能的影响, 从而判断种间交互作用能否驱动某个功能.根据功能值是否超过提前设定的阈值, 从而判断该功能是否达到预期的水平, 因此, 可以在特定的物种均匀度下, 根据不同的物种组成, 将预测的功能值与阈值进行对比, 从而可以判断哪个物种为优势种时, 各功能的表现最好.在不同的物种均匀度下, 可以检验随着物种均匀度的增加, 各个功能的表现是否也随之增加; 同时, 考虑到在一定的物种组成条件下, 不同功能间可能存在较大的差异, 因此, 可以检验随着物种均匀度的增加, 功能间的变异性是否发生变化.此外, 在相同的物种组成和均匀度下, 则可以比较物种相对多度对各个功能的影响(Dooley et al, 2015 ). ...
... 该方法是阈值法的前身, 对生态系统的每个功能和多样性之间的关系都进行了分析, 有助于阐明多功能性是由哪一个功能所驱动的.但该方法有多个方面的不足: (1)难以评估生态系统的整体功能; 当一个实验中, 多样性对一个生态系统的不同功能同时出现正、负效应时, 难以解释实验结果.(2)不能完全体现生物多样性效应;对于一个具有高多样性的群落而言, 有些物种对于一些功能来说可能是冗余的, 而对于其他功能来说, 这些物种又起着一定的作用.(3)只能作定性描述而不能进行定量分析.(4)实验结果易受个别物种的影响.(5)未考虑功能之间的相关性(Dooley et al, 2015 ).(6)不同功能对生态系统的重要性不同, 该方法未考虑不同功能的权重. ...
... 该方法简单直观且所得结果容易解释, 可以较明确地评估多样性的改变对生态系统多个功能的平均影响,以及多个功能的平均水平是否随多样性的增加而增加, 还能衡量群落同时维持多个生态系统功能的能力(Maestre et al, 2012b ).但其缺点也很明显: (1)该方法不能区分一个物种对不同功能重要性的差异(Maestre et al, 2012b ), 且无法分辨多样性对单个功能的影响, 这将不能真实地反映生态系统的基本情况.(2)虽然考虑了不同功能的权重, 但功能间的权重不易衡量.多数情况下, 研究者将不同功能视为相同的权重, 而事实上某个功能可能会对最终的结果产生很大的影响.因不同生态系统中的功能权重不尽相同, 即使确定了某一系统各功能的权重, 也难以将之推广.(3)难以区分由相似水平的几个功能所得的多功能性指数与部分较高水平和部分较低水平的几个功能平均所得的多功能性指数(Dooley et al, 2015 ).(4)平均值法在计算时默认一种功能的降低可以由另一种功能的上升来弥补, 而在一个生态系统中, 不同的功能并非都可以互相替代, 即某一功能的减弱并不一定能由另一功能的增加来补偿(Gamfeldt et al, 2008 ; Maestre et al, 2012b ). ...
... 该方法可以评估达到某一阈值的功能数是否随多样性的增加而增加.其优点是较灵活, 因为测定的是超过某一水平的功能数量, 故而多样性与多功能性的关系不管是线性还是非线性都不影响测定的结果, 即使功能间存在权衡、交互作用等问题, 也能很好地获取达到阈值的功能数, 并且适用范围广(Byrnes et al, 2014a ).但是单阈值法也有缺点: (1)阈值的选择较随意, 研究者往往会选择有利于实验结果表述的阈值, 而不同的阈值可能得到截然不同的结果.因此, 没有恰当的理由时, 单个阈值的结果往往让人难以信服.(2)对每个功能的测度存在不足, 不能反映超过阈值或低于阈值的程度, 只能体现生态系统的整体功能达到了哪个水平(Gamfeldt et al, 2008 ).(3)因缺少统一的标准, 在实际应用中很难准确地判断哪个阈值最能反映多样性与多功能性的关系(Gamfeldt et al, 2008 ), 只检验单个阈值可能会丢失一些重要的阈值, 如多样性作用最强时的阈值(Byrnes et al, 2014a ).(4)不能测定多样性对单个生态系统功能的影响.(5)未考虑功能之间的 相关性(Dooley et al, 2015 ).(6)未考虑不同功能的权重. ...
... 多阈值法在本质上与单阈值法是同一种方法, 但它将所有的阈值都进行了计算, 因此比单阈值法提供的信息更多, 不仅拥有单阈值法的优点, 并且比单阈值法更灵活、全面(Byrnes et al, 2014a ), 用该方法测得的多功能性水平可以在不同实验间进行直接比较.因为设置的阈值为0-100%, 因此不必考虑阈值的选择, 可以在各个阈值水平检验多样性对多功能性的影响.但其存在4个明显的缺点: (1)提供了一系列的测度指标, 相对而言比较繁琐(Byrnes et al, 2014a );(2)不能测定多样性对单个生态系统功能的影响;(3)未考虑功能之间的相关性(Dooley et al, 2015 );(4)未考虑不同功能的权重. ...
... 多元模型法是最新提出的一种检测BEMF关系的方法, 也是目前最能充分利用数据信息的方法, 它充分考虑了不同方面的多样性、功能间的权衡和相关性对多功能性的影响, 可以从多个角度获得较为全面的信息; 同时, 综合了单功能法、平均值法、物种替代法和单阈值法的优点, 实现了以往的方法没能达成的目标, 即在分析单个功能的同时量化生态系统多功能性.此外, 该方法的主要优点有(Dooley et al, 2015 ): (1)可以检测生态系统整体功能达到最大值时的物种组成和相对多度.(2)可以检测功能之间的权衡和相关性, 更好地解释多样性与多功能性的关系.(3)可以检测各物种及种间交互作用对各个生态系统功能的作用及相对重要性.(4)具有很强的预测能力, 如当物种丰富度相同时, 其他方法预测的多功能性是相同的, 但该方法根据不同的物种组成和相对多度可以识别出不同的多功能性.(5)该模型还可引入时间和空间变量来分析多功 能性. ...
... 尽管多元模型法有很多优点, 但其应用也有较大局限性, 主要为: (1)该方法需要一系列多样性梯度, 仅有单作和包含所有物种的混种这两个处理不适用该方法.(2)当物种数量较多时, 会产生很多的种间交互系数δ , 故而需要用一定的方法进行降维处理, 如根据物种所属功能群来划分δ (其他的方法可参考Kirwan et al, 2009 ).(3)目前只适合功能数较少(如3个功能)的研究, 在功能数较多时, 因分析时会产生大量的系数, 使得模型变得很复杂, 因而不适用这种方法(Dooley et al, 2015 ).此外, 该方法也同样没有考虑不同功能的权重问题. ...
Relative effects of species composition and richness on ecosystem properties in ponds.
1
2005
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
Grazer diversity, functional redundancy, and productivity in seagrass beds: an experimental test.
1
2001
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
Grazer diversity effects on ecosystem functioning in seagrass beds.
1
2003
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
Conservation evaluation and phylogenetic diversity.
1
1992
... 直系同源基因法本质上衡量的是群落中物种所拥有的功能基因的多样性, 而不是测度该群落真正能够行使的功能.该方法是分辨不同微生物功能的有效方法, 且不会低估微生物的功能, 可以定量估计微生物群落多样性的改变对生态系统功能的影响.同时, 直系同源基因法还能反映进化多样性, 从进化的角度与生物多样性进行了结合(Faith, 1992 ; Hendry et al, 2010 ).此外, 由于能从许多研究地点获取直系同源基因丰富度的数据, 因此可以在较大尺度上研究BEMF关系. ...
Getting started in gene orthology and functional analysis.
1
2010
... 直系同源基因是指不同物种中来自共同祖先的基因, 通常认为其功能保守, 因此具有相似的功能(Fang et al, 2010 ).该方法由Miki等(2014)提出, 用于测度微生物群落的多功能性, 直系同源基因丰富度越高则群落的多功能性越高.Miki等(2014)先在理论上进行探讨, 认为不同物种所拥有的同类直系同源基因具有相似的功能, 不同类别的直系同源基因具有不同的功能, 因此通过评估微生物群落中直系同源基因的丰富度就能知道群落功能的大致范围.之后, 他们通过微宇宙实验证明, 利用群落中微生物的直系同源基因序列丰富度可以预测实际的多功能性(Miki et al, 2014 ).用该方法测度多功能性时, 最好选择系统发生相近的微生物类群. ...
From selection to complementarity: shifts in the causes of biodiversity-productivity relationships in a long-term biodiversity experiment.
1
2007
... 该方法的结果易受到抽样效应(sampling effect)和互补效应(complementarity)的共同影响(Tilman et al, 1997a ; Loreau & Hector, 2001 ), 其中互补效应起主要作用(详见Loreau & Hector, 2001 ; Cardinale et al, 2007 ; Fargione et al, 2007 ).所谓抽样效应是一个统计学的概念, 指在同一生境中, 竞争力强的物种对资源的利用率更高, 故而有更高的生产力, 随着物种丰富度的增加, 从物种库中抽到高产物种的概率也将增大(Tilman et al, 1997b ); 所以在随机抽样的情况下, 高多样性的群落包含高产物种的概率更大.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 从物种库中得到对某几个功能或所有功能都有很强促进作用的物种的概率会增大, 从而得出物种丰富度增加会提高多功能性的结论.互补效应则是指在一个群落中, 物种间存在生态位差异, 故而群落中物种数越多, 所占据的“功能空间” (functional space)范围越广(Tilman et al, 1997b ).因此, 生态系统的物种丰富度越高, 对资源的利用率就越高, 相应的生产力也会越高.将之推广到多功能性, 则可以理解为: 随着物种丰富度的增加, 物种间的生态位差异增大, 不同物种支持不同的功能, 同样会得到因物种丰富度增加而多功能性提高的结论. ...
Multiple functions increase the importance of biodiversity for overall ecosystem functioning.
6
2008
... 传统的生物多样性与生态系统功能(biodiver- sity and ecosystem functioning, BEF)研究多关注植物物种丰富度和生态系统净初级生产力(NPP)之间的关系.显然, 单个生态系统功能指标(如NPP)并不能完全反映多样性对生态系统功能的影响(Hector & Bagchi, 2007 ; Gamfeldt et al, 2008 ; Zavaleta et al, 2010 ), 因为一个生态系统具有同时提供多种功能和服务的能力, 这被称为生态系统多功能性(ecosystem multifunctionality, EMF) (Hector & Bagchi, 2007 ). ...
... 其实在Hector和Bagchi之前, 已经有研究者尝试测度EMF.比如, Hooper和Vitousek (1998)最早利用平均值法(averaging approach)量化EMF, 当时还没有生态系统多功能性的概念, 他们将多个生态系统功能的表现转换成一个能反映整体功能的指标, 由此形成了现在使用最普遍的EMF测度方法——平均值法.之后, 又出现了单功能法(single function approach) (Duffy et al, 2003 ), 这种方法源自经典的BEF研究(Tilman et al, 1997a ; Duffy et al, 2001 ; Downing, 2005 ; Spehn et al, 2005 ), 只是传统研究关注的是多样性对单个功能的影响, 即便同时考虑了多个功能, 也未涉及EMF.Duffy等(2003)最先用单功能法描述了多个功能在同一时间内的表现水平.然而, 该方法并没有对EMF进行定量分析, 而是采用了定性描述, 因此, 这种方法在EMF研究中使用不多.除上述方法外, 近期又出现了几种新的方法, 包括单阈值法(single threshold approach) (Gamfeldt et al, 2008 )、多阈值法(multiple-threshold approach) (Byrnes et al, 2014a )、直系同源基因法(orthologous approach) (Miki et al, 2014 )和多元模型法(multivariate model approach) (Dooley et al, 2015 ). ...
... Gamfeldt等(2008) 最早使用该方法来评价生态系统多功能性, 他们选择所测功能最大值的50%作为阈值来评价每个功能在生态系统中的表现, 理由有两点: ...
... 该方法简单直观且所得结果容易解释, 可以较明确地评估多样性的改变对生态系统多个功能的平均影响,以及多个功能的平均水平是否随多样性的增加而增加, 还能衡量群落同时维持多个生态系统功能的能力(Maestre et al, 2012b ).但其缺点也很明显: (1)该方法不能区分一个物种对不同功能重要性的差异(Maestre et al, 2012b ), 且无法分辨多样性对单个功能的影响, 这将不能真实地反映生态系统的基本情况.(2)虽然考虑了不同功能的权重, 但功能间的权重不易衡量.多数情况下, 研究者将不同功能视为相同的权重, 而事实上某个功能可能会对最终的结果产生很大的影响.因不同生态系统中的功能权重不尽相同, 即使确定了某一系统各功能的权重, 也难以将之推广.(3)难以区分由相似水平的几个功能所得的多功能性指数与部分较高水平和部分较低水平的几个功能平均所得的多功能性指数(Dooley et al, 2015 ).(4)平均值法在计算时默认一种功能的降低可以由另一种功能的上升来弥补, 而在一个生态系统中, 不同的功能并非都可以互相替代, 即某一功能的减弱并不一定能由另一功能的增加来补偿(Gamfeldt et al, 2008 ; Maestre et al, 2012b ). ...
... 该方法可以评估达到某一阈值的功能数是否随多样性的增加而增加.其优点是较灵活, 因为测定的是超过某一水平的功能数量, 故而多样性与多功能性的关系不管是线性还是非线性都不影响测定的结果, 即使功能间存在权衡、交互作用等问题, 也能很好地获取达到阈值的功能数, 并且适用范围广(Byrnes et al, 2014a ).但是单阈值法也有缺点: (1)阈值的选择较随意, 研究者往往会选择有利于实验结果表述的阈值, 而不同的阈值可能得到截然不同的结果.因此, 没有恰当的理由时, 单个阈值的结果往往让人难以信服.(2)对每个功能的测度存在不足, 不能反映超过阈值或低于阈值的程度, 只能体现生态系统的整体功能达到了哪个水平(Gamfeldt et al, 2008 ).(3)因缺少统一的标准, 在实际应用中很难准确地判断哪个阈值最能反映多样性与多功能性的关系(Gamfeldt et al, 2008 ), 只检验单个阈值可能会丢失一些重要的阈值, 如多样性作用最强时的阈值(Byrnes et al, 2014a ).(4)不能测定多样性对单个生态系统功能的影响.(5)未考虑功能之间的 相关性(Dooley et al, 2015 ).(6)未考虑不同功能的权重. ...
... ).(3)因缺少统一的标准, 在实际应用中很难准确地判断哪个阈值最能反映多样性与多功能性的关系(Gamfeldt et al, 2008 ), 只检验单个阈值可能会丢失一些重要的阈值, 如多样性作用最强时的阈值(Byrnes et al, 2014a ).(4)不能测定多样性对单个生态系统功能的影响.(5)未考虑功能之间的 相关性(Dooley et al, 2015 ).(6)未考虑不同功能的权重. ...
Higher levels of multiple ecosystem services are found in forests with more tree species.
1
2013
... EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识.与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 ).单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
Linking soil bacterial diversity to ecosystem multifunctionality using backward- elimination boosted trees analysis.
1
2009
... 然而, 因为物种库中的物种对不同的功能有不同的作用, 而多样性对多功能性的影响取决于正负效应之间的权衡.若正效应大于负效应, 则随着多样性的增加, 多功能性将增加; 反之则降低.Hector和Bagchi (2007)的方法目前仅被用于各物种对生态系统功能具有正效应的情况(He et al, 2009 ; Isbell et al, 2011 ), 而在很多情况下并非如此.当将负效应考虑进来时, 就可以通过检验物种正效应是否强于、等于或者弱于负效应以及是否有物种主导整体功能的正负效应, 来更好地揭示BEMF的潜在关系.为此, Byrnes等(2014a)提出了2个额外的分析步骤来完善该方法: ...
Biodiversity and ecosystem multifunctionality.
10
2007
... 传统的生物多样性与生态系统功能(biodiver- sity and ecosystem functioning, BEF)研究多关注植物物种丰富度和生态系统净初级生产力(NPP)之间的关系.显然, 单个生态系统功能指标(如NPP)并不能完全反映多样性对生态系统功能的影响(Hector & Bagchi, 2007 ; Gamfeldt et al, 2008 ; Zavaleta et al, 2010 ), 因为一个生态系统具有同时提供多种功能和服务的能力, 这被称为生态系统多功能性(ecosystem multifunctionality, EMF) (Hector & Bagchi, 2007 ). ...
... ), 因为一个生态系统具有同时提供多种功能和服务的能力, 这被称为生态系统多功能性(ecosystem multifunctionality, EMF) (Hector & Bagchi, 2007 ). ...
... EMF概念的提出, 极大地丰富了人们对生物多样性与生态系统功能的认识.与单个生态系统功能相比, 一个生态系统需要更高的物种丰富度才能同时维持多个生态系统功能(Hector & Bagchi, 2007 ; Zavaleta et al, 2010 ).单个生态系统功能的研究往往会忽略不同功能之间的权衡(trade-off)关系, 而EMF的提出引发研究者对这一问题进行了新的探索(徐炜等, 2016 ).不同功能之间的权衡会导致生态系统同时提供多个功能能力的降低(Zavaleta et al, 2010 ; Gamfeldt et al, 2013 ), 因为多个生态系统功能很难同时达到较高水平. ...
... 第一步, 通过模型模拟每个物种的作用, 以此来量化物种对EMF的影响.依据最小AIC (Akaike Information Criterion)标准来选择模型, 用反向-剔除多元回归分析(backward-elimination multiple regression analysis)将物种依次从模型中剔除, 计算相应的AIC值.通过比较各物种剔除前后的AIC值, 将导致AIC值减小最多的物种彻底剔除, 不断重复此过程, 直至剔除任意一个物种都会使AIC值增加, 这时便得到了最优模型.未被剔除的物种集合就是影响该生态系统功能的最简物种组合(Hector & Bagchi, 2007 ).该模型对数据的要求较高, 通常要求不同多样性梯度下都有很好的可重复性(Byrnes et al, 2014a ). ...
... 利用上述分析得到的物种集合, 通过模型拟合以检测对某一功能组合有影响的物种, 从而判断支持不同功能的物种重叠度, 以此获取功能数与影响一个或一个以上功能的最小物种数之间的关系(Hector & Bagchi, 2007 ).因不同功能间物种的替代率与重叠度成反比, 故物种的替代率可由重叠度来计算.影响每一对功能的物种, 其重叠度可用Sørenson指数o 表示, 计算公式为: ...
... 根据上式可以定量预测一个生态系统中, 随着所考虑的生态系统功能数的增加, 维持这些功能所需的平均物种数.当对所考虑的功能与维持这些功能所需的物种数进行回归时, 有2种极端的情况: (1)如果一个物种只影响一个功能( ) 之间存在正向线性关系, 其斜率为1 (Hector & Bagchi, 2007 ); (2)如果生态系统所有的功能都受同一物种集合的影响( Hector & Bagchi, 2007 ).在实际研究中, 结果往往介于两者之间, 表明物种和功能间存在一定的替代(Hector & Bagchi, 2007 ; Isbell et al, 2011 ). ...
... = 1), 那么斜率就变为0, 截距即是维持相应生态系统功能所需的物种数目(Hector & Bagchi, 2007 ).在实际研究中, 结果往往介于两者之间, 表明物种和功能间存在一定的替代(Hector & Bagchi, 2007 ; Isbell et al, 2011 ). ...
... ).在实际研究中, 结果往往介于两者之间, 表明物种和功能间存在一定的替代(Hector & Bagchi, 2007 ; Isbell et al, 2011 ). ...
... 物种替代法主要关注不同的功能是否由不同物种驱动以及维持一定数量的功能需要多少个物种, 这和其他方法探讨在不同多样性水平下生态系统能维持的功能数或整体功能水平是不同的.其最大的优点是,能揭示不同的物种是否驱动不同的生态系统功能, 并能检测每个物种对不同功能的重要性.同时, 该方法还能对某一生态系统中维持特定数量的功能所需的平均物种数进行定量预测(Hector & Bagchi, 2007 ).但是, 该方法同样存在缺点: (1)未直接测定多功能性, 故不能用多功能性指数来定量估计多样性改变对多功能性的影响.(2)需要大量数据支持, 且对数据有较高的要求; 分析每个物种对各个功能的作用, 需要一系列的多样性梯度, 且每个梯度都要有很好的可重复性.(3)数据分析步骤较繁琐.(4)未考虑不同功能的权重. ...
... 根据对各方法原理和特点的分析, 研究者应根据研究问题的需要以及实验所得的数据选择合适的多功能性测度方法.事实上, 由于目前的每种方法都存在相应的缺点, 大多数分析方法会损失关键信息, 多元模型法虽然损失的信息较少, 但不适合功能数量较多时的情况, 而常规的多功能性研究往往功能数较多(Hector & Bagchi, 2007 ; Isbell et al, 2011 ; Jing et al, 2015 ).所以,仅靠一种方法很难对BEMF进行全面分析. ...

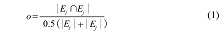
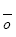
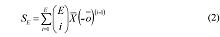
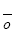
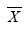
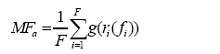
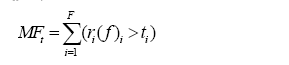
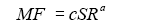
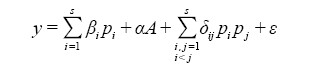
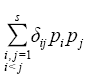
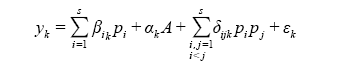


{kind=link}
{kind=link}
{kind=link}
{kind=link}