生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 )。生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 )。在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 )。但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的。
群落系统发育(community phylogeny)研究将系统发育生物学的方法运用到群落生态学中, 利用群落内物种的系统发育状况来推测历史因素对现有群落的影响, 通过分析群落内物种间的亲缘关系探究影响群落构建的主要原因(Webb, 2000 ; Webb et al ., 2002 )。群落系统发育研究主要是通过两个群落系统发育关系指数, 即净亲缘指数(net relatedness index, NRI)和最近亲缘指数(nearest taxon index, NTI), 将群落内物种间的系统发育距离和标准化零模型(即假设物种随机分布)下的系统发育距离进行比较, 统计二者间的差异, 用于检测群落是否存在系统发育结构。如果群落系统发育关系指数与零模型假设没有显著差异, 说明群落不存在系统发育结构或系统发育随机(phylogenetically randomness); 如果显著大于零模型假设, 说明群落具有系统发育结构, 表现为系统发育聚集(phylogeneticcally clustering); 相反, 如果显著小于零模型假设, 说明群落系统发育发散(phylogenetically over-dispersion)。达尔文早在1859年就曾说过, 和属外的物种相比, 同属内的物种在生活习性和结构上具有更近的相似性, 当它们相互之间发生竞争时, 也比与属外物种的竞争激烈得多(Darwin, 1859 )。这一观点与群落系统发育研究中生态位保守的假设(即亲缘关系越近的物种, 生态特征越相似)是一致的。在一个群落里, 如果是生境筛选作用占主导地位, 那么相同生境将筛选出适应能力相似、亲缘关系偏近的物种, 表现为系统发育聚集; 相反, 竞争排斥作用会使生态位相似的物种无法共存于同一环境, 则群落内物种亲缘关系较远, 表现为系统发育发散。据此, 在生态性状保守的前提下, 我们就可以从群落现有的系统发育表现形式, 推测群落构建过程中的主导因素(表1 )。
在最初的群落系统发育研究中, 多数学者主要是通过Phylomatic方法(Webb & Donoghue, 2005 )来探究群落内所有物种间的系统发育关系, 但这种方法也存在一些不足。首先, 采用Phylomatic方法所构建的系统发育树往往具有明显的多分支结构(polytomy), 尤其是当同一科属所含物种数目较多时这种现象更加明显; 其次, 基于APG系统构建的发育树缺乏裸子植物的信息, 而有些裸子植物也是群落建群种, 如云南的云南松(Pinus yunnanensis )林和长白山的阔叶红松(P. koraiensis )林等(裴男才等, 2011b )。
DNA条形码的快速兴起, 极大地促进了群落系统发育生态学的发展, 为精确地构建群落系统发育关系提供了新的技术。其中比较典型的研究, 如Kress等(2009)发现由DNA条形码建立的系统发育树为末端的低阶分类阶元提供了更为准确的系统位置, 并且得到的物种系统发育关系增加了拒绝零假设的概率; Gonzalez等(2010)认为, DNA条形码所构建的物种间精确的系统发育关系, 可以提高有关群落结构计算的统计力; Pei等(2011)和裴男才(2012)综合使用进化速率不同的DNA条形码片段, 相对准确地构建出鼎湖山南亚热带森林植物群落的系统发育关系。然而, Kress等(2010)在亚热带地区波多黎各森林样地的研究发现, 单独利用DNA 条形码序列所构建的群落系统发育关系与APG系统(APG III, 2009 )相比, 在较高的分类阶元即目水平上存在较大的出入, 需要利用APG系统的大框架作为约束树(constraint tree), 并对各分支末端的类群配合使用DNA条形码的建树方法, 才能获得群落物种间精确的系统发育关系(裴男才等, 2011 )。
生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 )。早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 )。但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 )。海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 )。研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义。
现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 )。其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 )。如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在。虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据。
哀牢山地处云贵高原、横断山脉和青藏高原三大自然地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地(牛跃等, 2008 )。哀牢山由于山体高大宽厚, 高差悬殊较大, 具有明显的垂直气候特征, 且海拔梯度升高引起的气候因子变化效应使得动物、植物以及土壤微生物等也相应呈现垂直分带现象(吴征镒等, 1983 )。哀牢山西坡的森林植被类型多样, 根据物种组成、植被结构、气候以及土壤环境等特征, 从海拔1,300-2,750 m大致可以划分为3个植被带: 山体中下部的亚热带季风常绿阔叶林(海拔高度1,300-2,100 m)、山体中部的中山湿性常绿阔叶林(海拔高度2,100-2,600 m)和山顶部的苔藓矮林(海拔高度2,600-2,750 m)。这种显著的山体垂直分布格局是开展生物多样性沿海拔梯度变化研究的理想场所。本研究以对温度变化极为敏感的山地植被为研究对象(Walter, 1979 ; Cavender-Bares et al ., 2009 ), 开展哀牢山亚热带森林群落系统发育研究, 分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化, 从而推测群落构建的生态学过程。
1 方法
1.1 野外样方建立与样品采集
以哀牢山西坡作为主要研究区域, 在沿海拔梯度分布的3种植被类型中选取16个群落样点(表2 )。在每个样点上分别建立20 m×30 m的群落样方, 调查和记录样方内所有胸径大于1 cm的乔木树种的名称、树高、胸径等信息, 同时采集样方内每个树种用于DNA提取的叶片或树皮材料、凭证标本以及照片。哀牢山西坡16个样点共采集到144个树种, 涉及49科(按APG III系统), 其中包含两种裸子植物。
1.2 DNA提取、扩增及排序
使用天根公司的试剂盒完成所有植物样品的总DNA提取。选择叶绿体rbc L、mat K、基因间隔区trn H-psb A与核基因ITS等4个分子标记作为研究的扩增片段。PCR参照国际生命DNA条形码联盟植物工作组推荐的反应体系(CBOL Plant Working Group, 2009 ), 并作出优化和调整。扩增产物送北京六合华大基因科技有限公司(BGI)完成测序。得到的序列使用软件Sequencher 4.14(Gene Codes Corporation, Ann Arbor, Mich.)进行拼接和校对, rbc L、mat K和ITS等3个DNA片段在Geneious 6.1.5(Biomatters Ltd.)中选择Clustal X(Larkin et al. , 2007 )排序, 而trn H-psb A由于序列长度变化大, 用软件SATé(Liu et al. , 2009 )进行排序。
1.3 哀牢山森林群落系统发育树构建
构建方法主要参考Kress等(2010)的研究, 以APG III系统为大框架作“约束树”, 用4个分子标记得到的DNA分子序列来解决末端分类单元确切的系统位置。具体步骤是: 先将APG认可的物种名录提交到Phylomatic在线软件上运行, 获得哀牢山森林群落的简易系统发育关系, 以科为分类单元, 把科内的所有物种平行化(collapse), 形成APG系统的“约束树”; 再将4个DNA片段分别排序后合并成一个联合矩阵, 即DNA序列超矩阵; 然后把DNA超矩阵和“约束树”在CIPRES超级远程计算机(Miller et al ., 2010 )的RAxML程序上(Stamatakis, 2006 )进行最大似然方法ML分析, 并通过500次的重复来获得拓扑结构上的节点支持率, 将各节点的支持率附在拓扑结构最优的系统发育树上; 最后用r8s软件包(Sanderson, 2003 )非参数速率平滑法(NPRS)对系统发育树进行处理, 进而获得用于群落系统发育结构分析的进化树。
1.4 群落物种多样性和系统发育多样性分析
依据样方调查材料, 采用物种丰富度(S )、Shannon-Wiener指数(H )、Simpson优势度指数(D )和Pielou均匀度指数(E )(马克平, 1994 ), 比较和评价哀牢山3种典型植被森林群落多样性的差异。
S = 样地内的植物物种总数
( Shannon, 1949)
( Simpson, 1949)
( Pielou, 1969)
对群落系统发育多样性的分析采用系统发育多样性Faith’s PD指数(Faith, 1992 )和系统发育beta多样性PhyloSor指数来量化, 在R软件(R Development Core Team, 2005)中完成计算。系统发育多样性Faith’s PD指数是指某一样地中分类单元系统发育分支长度之和(Faith, 1992 ; Kemble et al. , 2010 ), 值越小, 表明该样地内物种的系统发育多样性在整个区域的系统发育多样性占有的比重越小, 在R程序的软件包picante中计算得到。系统发育beta多样性PhyloSor 指数是指两个群落共有物种的枝长和占两个群落所有物种的枝长和的比例, 计算公式为:
PhyloSor ij = 2BL ij /(B Li +BL j )
其中BL i、 BL j 分别为群落i和群落j内所有物种系统发育结构的枝长和, BL ij 为两个群落内共有物种系统发育结构的枝长和(Bryant et al ., 2008 )。PhyloSor比值越大, 表明这两个群落间的相似性越高。
1.5 群落系统发育结构
为了测量群落内物种之间的亲缘关系, 使用R软件中的picante软件包计算平均系统发育距离(MPD)和平均最近系统发育类群的距离(MNPD)。这两个参数可转换成净亲缘关系指数(NRI)和最近种间亲缘关系指数(NTI)。采用的方法是: 先建立代表森林群落物种库的超级系统发育树, 再将实际的每个群落与标准化的零模型作比较, 统计实际观测值与零假设期望值间的差异。零模型的构建采用群落中所有物种于系统进化树末端随机置换999次的方法, 保持每次零模型和生成的系统进化树具有相同的物种数目。如果量化后的群落指数显著大于零模型, 则反映出该群落系统发育聚集(物种之间的亲缘关系比预期的近); 反之, 则表明该群落系统发育发散(物种之间的亲缘关系比预期的远), 从而可推断群落装配的关键过程。NRI 和NTI 计算公式如下:
(1)
(2)
本研究借鉴了Bryant等(2008) 和Ghosh-Harihar等(2014)的研究策略, 将调查得到的所有树种作为物种库, 然后计算群落内物种间的谱系距离, 通过统计方法检测其与随机模型下的谱系距离是否有显著差异来获得谱系结构。使用Student t- 检验, 确定不同海拔梯度(低、中、高)的群落系统发育结构是否显著不同于零模型。所有的NRI值和NTI值计算时未进行多度加权。
2 结果
2.1 物种多样性和系统发育多样性沿海拔梯度的变化
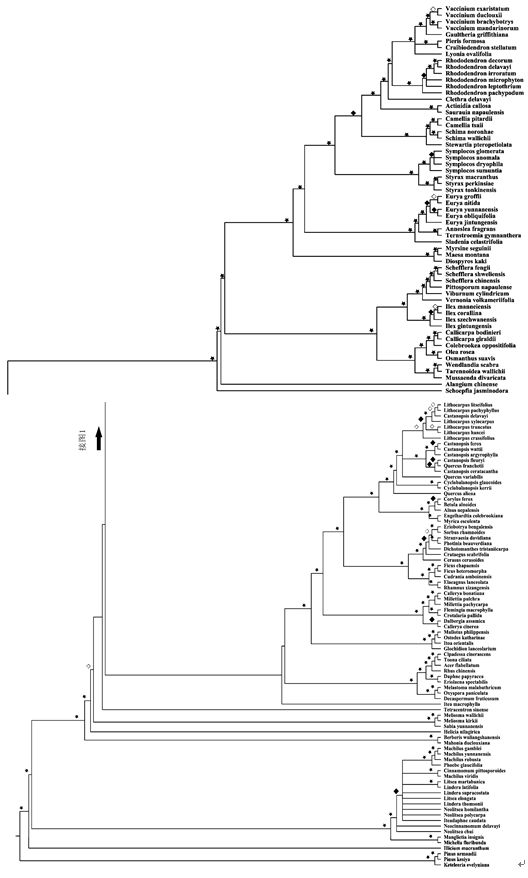
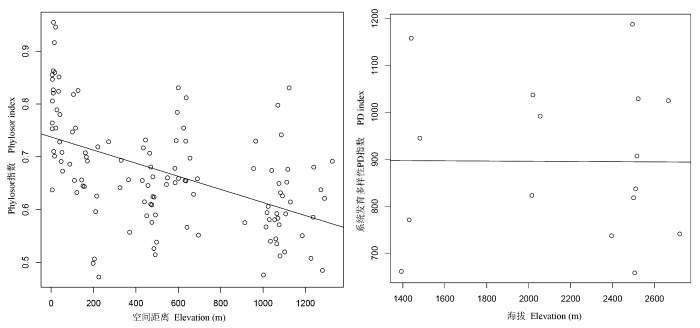
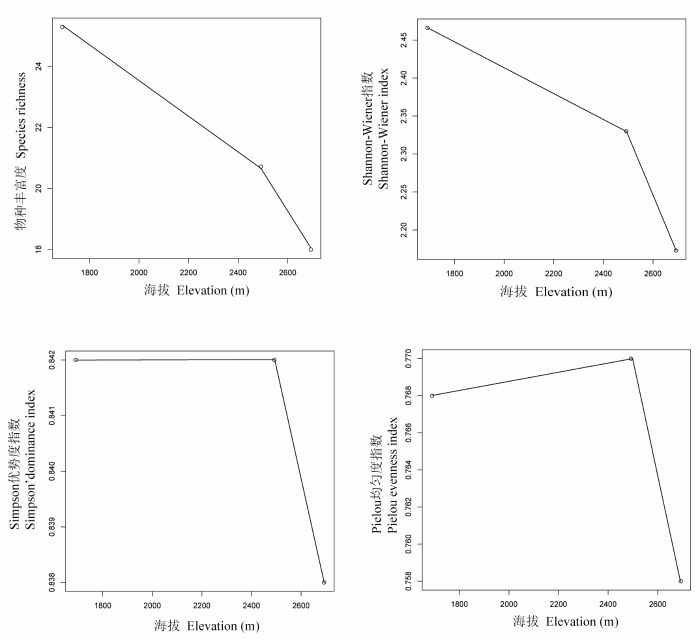
对于哀牢山垂直分布的3种植被类型而言, 低海拔的季风常绿阔叶林的丰富度和Shannon-Wiener指数最高, 并随着海拔的升高呈现逐渐降低的趋势; 而Simpson优势度指数和Pielou均匀度指数变化不明显(附图1 )。应用DNA条形码并结合APG III系统构建的包含144个物种的群落系统发育树如图1 所示。基于该发育关系得到的系统发育beta多样性PhyloSor指数与空间距离呈明显的负相关关系(图2 )。而群落系统发育多样性PD指数在不同的植被类型间没有显著的变化, 但海拔相近的样方之间差异较为明显(图2 )。
图1 利用DNA条形码与APG III系统相结合得到的哀牢山森林群落的16个样地144个物种系统发育进化树。每个节点靴带支持率数值用星号(≥ 85%, 高度支持)、菱形(空白)(70-84%, 中度支持)、菱形(黑色填充)(50-69%, 较弱支持)表示。
Fig. 1 A community phylogeny of 144 species from 16 plots along the elevational gradients in Ailao Mountains. The community phylogeny is constructed based on a maximum likelihood analysis of rbc L, mat K, psb A-trn H and ITS sequence data with APG III as a constraint tree. Nodes with strong (≥ 85%), moderate (70-84%) and/or weak (50-69%) bootstrap support are indicated by an asterisk, a diamond filled with blank and a diamond filled with black, respectively.
图2 系统发育多样性PhyloSor指数和PD指数随海拔梯度变化的趋势
Fig. 2 Variation in community phylogenetic diversity along the elevation gradient from two indices: PhyloSor and PD
2.2 群落系统发育结构沿海拔梯度的变化规律
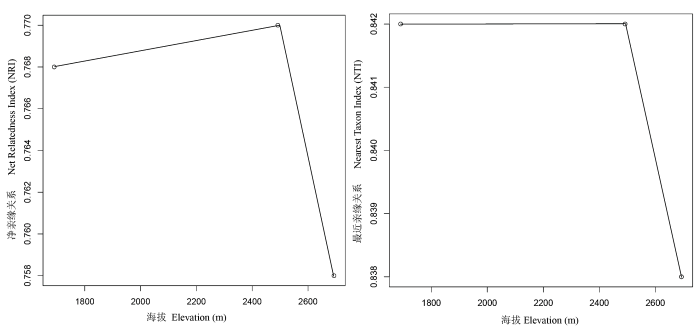
在整体的变化趋势上, 两个群落系统发育指数NRI和NTI都支持哀牢山群落系统发育结构随着海拔梯度的上升呈现出聚集程度逐渐降低至随机或发散的变化规律, 但统计学上没有达到显著性(R 2 = 0.08, P = 0.294)。考虑到本研究在高海拔地区仅有两个样方, 如去除这两个样方进行分析, 则发现由NRI反映的聚集程度降低的趋势具有统计学上的显著性(R 2 = 0.34, P = 0.03)。在相同植被类型的不同样方之间, 群落系统发育结构也存在一定的变化(图3 )。在低海拔季风常绿阔叶林中, 绝大多数群落NRI和NTI显著大于零, 表现为系统发育聚集, 但随海拔梯度的升高, 聚集程度降低, 个别样方甚至表现为系统发育随机(如plot 7); 在中海拔2,400-2,550 m上的中山湿性常绿阔叶林群落中, 出现了多种群落系统发育结构, 一部分样地仍然是系统发育聚集, 另一部分表现为系统发育发散; 在高海拔的山顶苔藓矮林中, NRI指数大于零, 表现为系统发育聚集, 但NTI指数接近或小于零, 表现为系统发育随机或发散, 两个指数得出了相反的结果。
图3 两个群落系统发育指数(NRI和NTI)随海拔梯度的变化趋势
Fig. 3 Variation in community phylogenetic relatedness along the elevation gradient as measured with the NRI and NTI
3 讨论
3.1 群落多样性和系统发育多样性沿海拔梯度的变化
本研究中选取的4个群落多样性指数更多地是表现出了随海拔升高逐渐降低的变化趋势。这与施济普等(2005)对该地区的研究结果一致, 但与牛跃等(2008)得出的“单峰”格局不符。造成这一差异的原因可能是, 本研究样方设置中对低海拔地区群落样方的选取主要参考了施济普等(2005)的研究地点, 而未选择牛跃等(2008)研究中所涉及的位于山体下部(1,100-1,300m)、具有较低生物多样性的季风常绿阔叶林这一海拔区域, 因为在野外调查时我们发现该区域的季风常绿阔叶林破坏严重, 已经难以找到保存完好的原生林。PhyloSor指数显示, 群落间的相似性随着海拔差异的增加逐渐而降低的规律在哀牢山亚热带森林群落中依然适用, 与之前诸多研究结果也是一致的(如Soininen et al ., 2007 ; Bryant et al ., 2008 ), 表明群落系统发育beta多样性会随空间距离增大而降低。
3.2 哀牢山森林群落沿海拔梯度的构建过程和驱动因素
哀牢山森林群落系统发育结构随着海拔梯度的上升由系统发育聚集走向发散或聚集程度降低, 呈现出非随机的状态, 说明中性理论预测群落随机的系统发育结构在哀牢山地区并不适用, 暗示在该地区森林中生态位过程对于群落的组成和维持更加重要。在生态位理论中, 生境过滤和竞争排斥是两个相反的过程, 前者使得物种间的系统发育结构相对聚集, 而后者使得亲缘关系近的物种不能共存, 系统发育结构相对发散(牛红玉等, 2011 )。哀牢山森林群落系统发育结构在低海拔表现出系统发育聚集, 说明生境过滤作用起着主导作用。对此有两种可能的解释: 其一, 低海拔地区受温度和水分等的影响比高海拔地区小得多, 地形异质性和生境异质性都较小, 促使进化程度相似的类群得以共存; 其二, 低海拔地区的森林群落受到的人为干扰较多(非破坏性的干扰), 干扰(如砍伐、火烧等)作为一种环境过滤器会促使群落系统发育聚集(Mo et al ., 2012), 这可能也是造成低海拔群落系统发育结构聚集的一个重要因子(Dinnage, 2009 ; 牛红玉等, 2011 )。中海拔地区出现了多种系统发育结构(聚集、发散和随机), 我们推测可能是由于随着海拔的上升, 中海拔的生境异质性要比低海拔复杂得多, 群落物种间会更倾向于采取不同的资源利用策略, 物种间的竞争作用会加剧, 同时存在生境过滤和竞争排斥两种生态过程的相互作用, 从而表现出了更多的系统发育结构。
两个群落系统发育指数NRI和NTI反映出的高海拔地区群落(山顶苔藓矮林)的系统发育结构相反, NRI表现出系统发育聚集, 而NTI却表现为随机或发散。NRI和NTI是两个相互联系却又有不同的系统发育指数, 前者基于群落整体水平的亲缘关系, 反映发生在整个系统树上的系统发育模式, 而后者仅仅基于最近的系统发育关系, 主要针对系统进化树的末端(Kraft et al. , 2007 )。NRI和NTI不一致或相反的结果也出现在其他的研究中, 例如Bryant等(2008)发现这两个指数在检测高海拔地区的植物群落系统发育时表现不一致, 通过NRI指数的变化趋势, 认为在高海拔地区系统发育结构更趋向发散; Kress等(2009)在检测不同生境下不成熟森林的系统发育时出现了NRI小于零, 而NTI大于零的现象, 只是NTI的统计结果不显著(P = 0.17)。
有研究显示, 哀牢山高海拔的山顶苔藓矮林由于承受较强的紫外线辐射和大风等环境胁迫, 小叶所占的比例要比中山湿性常绿阔叶林高(①施济普 (2007) 云南山顶苔藓矮林群落生态学与生物地理学研究. 硕士论文, 中国科学院西双版纳热带植物园, 昆明。); 同时, You等(2013) 对哀牢山的山顶苔藓矮林开展了微气象学研究, 发现植物在高海拔地区承受了更强的干旱胁迫。以上研究结果反映在高强度的环境压力下, 高海拔地区的物种可能发生了趋同进化。然而, 生境过滤后虽聚集了性状相似的物种, 但它们之间的亲缘关系并不相近, 表现为谱系发散; 而另外一些因竞争排斥作用留下的具有不同性状的物种间亲缘关系可能会没有规律, 表现为谱系随机或者谱系聚集, 正如本研究NRI指数所反映的结果。
鉴于NRI和NTI反映出来的趋势存在差异, 加之本研究在高海拔地区样方过少, 我们认为进一步探讨高海拔地区群落的系统发育结构及其驱动因素, 还需要通过增加更多的取样, 并结合植物功能性状进行综合研究。
3.3 物种库和零模型对群落系统发育的影响
物种库和零模型的选择对群落系统发育生态学研究的结果有着很大影响(Kooyman et al. , 2011 )。物种库是指一个地区可进入某一特定群落的潜在物种数目(方精云等, 2009 ), 但在实际的研究中很难验证一个群落的实际物种库, 本研究中选取群落内所有物种作为小尺度物种库是目前来说相对合理的一种选择。Swenson等(2006)研究发现, 物种库的空间尺度越大, 越有可能发生系统发育聚集。在未来的研究中, 针对不同的研究区域范围和类群, 如何更为合理地选择物种库, 需要进一步改进。同时, 目前选择的零模型也不完善, 应该全面考虑群落中的物种丰富度和多度以及生态学特征, 并将进化特征和群落生态学性状特征结合起来(Kraft et al ., 2007 )。因此, 将来需要更多的研究来验证NRI和NTI指数的准确性并寻找更加合理的模型, 增加统计分析和解决问题的能力。
本研究揭示了哀牢山森林群落的系统发育结构随海拔梯度的升高呈现由聚集走向发散或聚集程度降低的变化规律, 表明在亚热带森林中生态位过程较中性过程而言对群落的构建发挥着更为重要的作用。生境过滤在低海拔群落构建中起到主要作用, 海拔梯度和较多的人为干扰可能是造成该地区系统发育聚集的原因; 生境过滤和竞争排斥在中海拔森林群落的构建中共同发挥作用; 高海拔群落的系统发育结构及其驱动因素尚需进一步的研究。利用群落系统发育方法来理解和探讨群落装配的过程, 对全球气候变化下生物多样性的保护与维持有着非常重要的意义, 但考虑到本研究还存在不足之处, 如受客观条件的限制在高海拔地区选取的群落样方过少, 以及缺乏小生境的环境因子和群落的功能性状特征数据等, 在今后的研究中需要开展综合的多维度研究来进一步深入探讨森林群落构建的机制。
附图1 哀牢山亚热带不同植被类型的群落丰富度、优势度、均匀度和多样性指数
Fig. S1 Species richness, evenness and species diversity of different vegetation types in Ailao Mountainshttp://www.biodiversity-science.net/fileup/PDF/w2014-011-1.pdf
(责任编委: 米湘成 责任编辑: 周玉荣)
致谢: 野外考察得到了云南省林业厅、哀牢山国家级自然保护区景东管理局、中国科学院西双版纳热带植物园哀牢山生态站的大力支持; 物种鉴定工作由中国科学院昆明植物研究所李锡文研究员完成; 采样工作得到了中国科学院西双版纳热带植物园植物系统发育与保护生物学实验室李朗博士、李恒、刘志芳等人的协助, 图片绘制工作得到了中国科学院热带森林生态学重点实验室(西双版纳热带植物园)杨洁博士的帮助, 在此一并表示感谢。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
APG III 2009 ) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
Botanical Journal of the Linnean Society , 161 , 105 -121 .
[本文引用: 1]
[2]
Bryant JA Lamanna C Morlon H Kerkhoff AJ Enquist BJ Green JL 2008 ) Microbes on mountain sides: contrasting elevational patterns of bacterial and plant diversity
. Proceedings of the National Academy of Sciences,USA , 105 , 11505 -11511 .
[本文引用: 4]
[3]
Cavender-Bares J Kozak KH Fine PVA Kembel SW 2009 ) The merging of community ecology and phylogenetic biology.
Ecology Letters , 12 , 693 -715 .
[本文引用: 1]
[4]
CBOL Plant Working Group 2009 ) A DNA barcode for land plants.
Proceedings of the National Academy of Sciences, USA , 106 , 12794 -12797 .
[本文引用: 1]
[5]
Darwin C 1859 ) On the Origin of Species by Means of Natural Selection
. John Murray, London.
[本文引用: 1]
[6]
Dinnage R 2009 ) Disturbance alters the phylogenetic composition and structure of plant communities in an old field system.
PLoS ONE , 4 , e7071 .
[本文引用: 1]
[7]
Emerson BC Gillespie RG 2008 ) Phylogenetic analysis of community assembly and structure over space and time.
Trends in Ecology and Evolution , 23 , 619 -630 .
[本文引用: 1]
[8]
Faith DP 1992 ) Systematics and conservation: on predicting the feature diversity of subsets of taxa.
Cladistics , 8 , 361 -373 .
[本文引用: 2]
[9]
Fang JY 方精云 ), Wang XP 王襄平 ), Tang ZY 唐志尧 ) (2009 ) Local and regional processes control species richness of plant communities: the species pool hypothesis.
Biodiversity Science (生物多样性 ), 17 , 605 -612 . (in Chinese with English abstract)
[本文引用: 1]
[10]
Gaston KJ 2000 ) Global patterns in biodiversity.
Nature , 405 , 220 -226 .
[本文引用: 1]
[11]
Ghosh-Harihar M 2014 ) Phylogenetic and ecomorphological structure of assemblages of breeding leaf warblers (Phylloscopidae) along Himalayan elevational gradients.
Journal of Biogeography , 41 , 1193 -1203 .
[12]
Graham CH Parra JL Rahbekb C McGuire JA 2009 ) Phylogenetic structure in tropical hummingbird communities.
Proceedings of the National Academy of Sciences, USA , 106 , 19673 -19678 .
[本文引用: 1]
[13]
Goldberg DE Miller TE 1990 ) Effects of different resource additions on species diversity in an annual plant community.
Ecology , 71 , 213 -225 .
[本文引用: 1]
[14]
Gonzalez MA Roger A Courtois EA Jabot F Norden N Paine CET Baraloto C Thébaud C Chave J 2010 ) Shifts in species and phylogenetic diversity between sapling and tree communities indicate negative density dependence in a lowland rain forest.
Journal of Ecology , 98 , 137 -146 .
[15]
Johnson MTJ Stinchcombe JR 2007 ) An emerging synthesis between community ecology and evolutionary biology.
Trends in Ecology and Evolution , 22 , 250 -257 .
[本文引用: 1]
[16]
Kembel SW Cowan PD Helmus MR Cornwell WK Morlon H Ackerly DD Blomberg SP Webb CO 2010 ) Picante: R tools for integrating phylogenies and ecology.
Bioinformatics , 26 , 1463 -1464 .
[本文引用: 1]
[17]
Kikkawa J Williams EE 1971 ) Altitude distribution of land birds in New Guinea.
Search , 2 , 64 -65 .
[本文引用: 1]
[18]
Kraft NJB Cornwell WK Webb CO Ackerly DD 2007 ) Trait evolution, community assembly, and the phylogenetic structure of ecological communities.
The American Naturalist , 170 , 271 -283
[本文引用: 4]
[19]
Kress WJ Erickson DL Jones FA Swenson NG Perez R Sanjur O Bermingham E 2009 ) Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama.
Proceedings of the National Academy of Sciences, USA , 106 , 18621 -18626 .
[20]
Kress WJ Erickson DL Swenson NG Thompson J Uriarte M Zimmerman JK 2010 ) Advances in the use of DNA barcodes to build a community phylogeny for tropical trees in a Puerto Rican forest dynamics plot.
PLoS ONE , 5 , e15409 .
[21]
Kooyman R Rossetto M Cornwell W Westoby M 2011 ) Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests.
Global Ecology and Biogeography , 20 , 707 -716 .
[本文引用: 1]
[22]
Larkin MA Blackshields G Brown NP Chenna R McGettigan PA McWilliam H Valentin F Wallace IM Wilm A Lopez R Thompson JD Gibson TJ Higgins DG 2007 ) Clustal W and Clustal X version 2.0.
Bioinformatics , 23 , 2947 -2948 .
[本文引用: 1]
[23]
Liu K Raghavan S Nelesen S Linder CR Warnow T 2009 ) Rapid and accurate large-scale coestimation of sequence alignments and phylogenetic trees.
Science , 324 , 1561 -1564 .
[本文引用: 1]
[24]
Loya Y 1972 ) Community structure and species diversity of hermatypic corals at Eilat, Red Sea.
Marine Biology , 13 , 100 -123 .
[本文引用: 1]
[25]
Ma KP 马克平 ) (1994 ) The measurement of community diversity. In: Principles and Methodologies of Biodiversity Studies (生物多样性研究的原理与方法) (eds Qian YQ (钱迎倩), Ma KP (马克平)) , pp. 141 -165 . China Science and Technology Press , Beijing . (in Chinese)
[本文引用: 1]
[26]
Machac A Janda M Dunn RR Sanders N 2011 ) Elevational gradients in phylogenetic structure of ant communities reveal the interplay of biotic and abiotic constraints on diversity.
Ecography , 34 , 364 -371 .
[本文引用: 1]
[27]
McCain CM 2005 ) Elevational gradients in diversity of small mammals.
Ecology , 86 , 366 -372 .
[本文引用: 1]
[28]
Miller MA Pfeiffer W Schwartz T 2010 ) Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE) , pp.1 -8 , New Orleans , LA .
[本文引用: 1]
[29]
Mo XX Shi LL Zhang YJ Zhu H Slik JWF 2013 ) Change in phylogenetic community structure during succession of traditionally managed tropical rainforest in Southwest China.
PLoS ONE , 8 , e71464 .
[30]
Monk CD Child GI Nicholson SA 1969 ) Species diversity of a stratified oak-hickory community.
Ecology , 50 , 468 -470 .
[本文引用: 1]
[31]
Niu HY 牛红玉 ), Wang ZF 王峥峰 ), Lian JY 练琚愉 ), Ye WH 叶万辉 ), Shen H 沈浩 ) (2011 ) New progress in community assembly: community phylogenetic structure combining evolution and ecology.
Biodiversity Science (生物多样性 ), 19 , 275 -283 . (in Chinese with English abstract)
[本文引用: 3]
[32]
Niu KC 牛克昌 ), Liu YN 刘怿宁 ), Shen ZH 沈泽昊 ), He FL 何芳良 ), Fang JY 方精云 ) (2009 ) Community assembly: the relative importance of neutral theory and niche theory.
Biodiversity Science (生物多样性 ), 17 , 579 -593 . (in Chinese with English abstract)
[本文引用: 1]
[33]
Niu Y 牛跃 ), Liu Y 刘洋 ), Zhang YP 张一平 ), Gao JB 高进波 ) (2008 ) Vertical distribution patterns of plant species diversity in Ailao Mountains.
Journal of West China Forestry Science (西部林业科学 ), 37 , 27 -34 . (in Chinese with English abstract)
[本文引用: 2]
[34]
Oomen MA Shanker K 2005 ) Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants.
Ecology , 86 , 3039 -3047 .
[本文引用: 1]
[35]
Pei NC Lian JY Erickson DL Swenson NG Kress WJ Ye WH Ge XJ 2011 ) Exploring tree-habitat associations in a Chinese subtropical forest plot using a molecular phylogeny generated from DNA barcode loci.
PLoS ONE , 6 , e21273 .
[本文引用: 1]
[36]
Pei NC 裴男才 ), Zhang JL 张金龙 ), Mi XC 米湘成 ), Ge XJ 葛学军 ) (2011 ) Plant DNA barcodes promote the development of phylogenetic community ecology.
Biodiversity Science (生物多样性 ), 19 , 284 -294 . (in Chinese with English abstract)
[本文引用: 1]
[37]
Pei NC 裴男才 ) (2012 ) Building a subtropical forest community phylogeny based on plant DNA barcodes from Dinghushan plot.
Plant Diversity and Resources (植物分类与资源学报 ), 34 , 263 -270 . (in Chinese with English abstract)
[38]
Pielou EC 1969 ) Introduction to Mathematical Ecology. Wiley-Interscience
, New York.
[39]
Rahbek C 1997 ) The relationship among area, elevation, and regional species richness in Neotropical birds.
The American Naturalist , 149 , 875 -902 .
[本文引用: 1]
[40]
R Development Core Team 2013 ) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2013. URL
.
URL
[41]
Sanderson MJ 2003 ) r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock.
Bioinformatics , 19 , 301 -302 .
[本文引用: 1]
[42]
Shannon CE Weaver W 1949 ) A Mathematical Theory of Communication . University of Illinois Press , Urbana, IL .
[43]
Shi JP 施济普 ), Zhao CJ 赵崇奖 ), Zhu H 朱华 ) (2005 ) Characteristics and species composition of main vegetation types on west slope of the Ailao Mountains in Yunnan.
Chinese Journal of Applied and Environmental Biology (应用与环境生物学报 ), 11 , 1 -7 . (in Chinese with English abstract)
[44]
Simpson EH 1949 ) Measurement of diversity.
Nature , 163 , 688 .
[45]
Soininen J McDonald R Hillebrand H 2007 ) The distance decay of similarity in ecological communities.
Ecography , 30 , 3 -12 .
[本文引用: 1]
[46]
Stamatakis A 2006 ) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models.
Bioinformatics , 22 , 2688 -2690 .
[本文引用: 1]
[47]
Stevens RD 2006 ) Historical processes enhance patterns of diversity along latitudinal gradients.
Proceedings of the National Academy of Sciences, USA , 273 , 2283 -2289 .
[本文引用: 1]
[48]
Swenson NG Enquist BJ Pither J Thompson J Zimmerman JK 2006 ) The problem and promise of scale dependency in community phylogenetics.
Ecology , 87 , 2418 -2424 .
[49]
Terborgh J 1977 ) Bird species diversity on an Andean elevation gradient.
Ecology , 58 , 1007 -1019 .
[本文引用: 1]
[50]
Tunnicliffe V 1981 ) High species diversity and abundance of the epibenthic community in an oxygen-deficient basin.
Nature , 294 , 354 -356 .
[本文引用: 1]
[51]
You GY Zhang YP Liu YH Schaefer D Gong HD Gao JB Lu ZY Song QH Zhao JB Wu CS Yu L Xie YN 2013 ) Investigation of temperature and aridity at different elevations of Mt. Ailao, SW China.
International Journal of Biometeorology , 57 , 487 -492 .
[本文引用: 1]
[52]
Vamosi SM Heard SB Vamosi JC Webb CO 2009 ) Emerging patterns in the comparative analysis of phylogenetic community structure.
Molecular Ecology , 18 , 572 -592 .
[本文引用: 1]
[53]
Walter H 1979 ) Vegetation of the Earth. Springer
, New York.
[本文引用: 2]
[54]
Wang GH 王国宏 ) (2002 ) Species diversity of plant communities along an altitudinal gradient in the middle section of northern slopes of Qilian Mountains, Zhangye, Gansu.
Biodiversity Science (生物多样性 ), 10 , 7 -14 . (in Chinese with English abstract)
[本文引用: 1]
[55]
Webb CO 2000 ) Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
The American Naturalist , 156 , 145 -155 .
[本文引用: 1]
[56]
Webb CO Ackerly DD McPeek MA Donoghue MJ 2002 ) Phylogenies and community ecology.
Annual Review of Ecology and Systematics , 33 , 475 -505 .
[本文引用: 3]
[57]
Webb CO Donoghue MJ 2005 ) Phylomatic: tree assembly for applied phylogenetics.
Molecular Ecology Notes , 5 , 181 -183 .
[本文引用: 1]
[58]
Wiens JJ Parra-Olea G Garcia-Paris MM Wake DB 2007 ) Phylogenetic history underlies elevational biodiversity patterns in tropical salamanders.
Proceedings of the Royal Society B , 274 , 919 -928 .
[本文引用: 1]
[59]
Wu ZY 吴征镒 ), Qu ZX 曲仲湘 ), Jiang HQ 姜汉侨 ) (1983 ) Research of Forest Ecosystem on Ailao Mountains, Yunnan (云南哀牢山森林生态系统研究 ). Yunnan Science and Technology Press , Kunming . (in Chinese)
[本文引用: 1]
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III.
1
2009
... DNA条形码的快速兴起, 极大地促进了群落系统发育生态学的发展, 为精确地构建群落系统发育关系提供了新的技术.其中比较典型的研究, 如Kress等(2009)发现由DNA条形码建立的系统发育树为末端的低阶分类阶元提供了更为准确的系统位置, 并且得到的物种系统发育关系增加了拒绝零假设的概率; Gonzalez等(2010)认为, DNA条形码所构建的物种间精确的系统发育关系, 可以提高有关群落结构计算的统计力; Pei等(2011)和裴男才(2012)综合使用进化速率不同的DNA条形码片段, 相对准确地构建出鼎湖山南亚热带森林植物群落的系统发育关系.然而, Kress等(2010)在亚热带地区波多黎各森林样地的研究发现, 单独利用DNA 条形码序列所构建的群落系统发育关系与APG系统(APG III, 2009 )相比, 在较高的分类阶元即目水平上存在较大的出入, 需要利用APG系统的大框架作为约束树(constraint tree), 并对各分支末端的类群配合使用DNA条形码的建树方法, 才能获得群落物种间精确的系统发育关系(裴男才等, 2011 ). ...
Microbes on mountain sides: contrasting elevational patterns of bacterial and plant diversity
4
2008
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
... 其中BL i、 BL j 分别为群落i和群落j内所有物种系统发育结构的枝长和, BL ij 为两个群落内共有物种系统发育结构的枝长和(Bryant et al ., 2008 ).PhyloSor比值越大, 表明这两个群落间的相似性越高. ...
... 本研究借鉴了Bryant等(2008) 和Ghosh-Harihar等(2014)的研究策略, 将调查得到的所有树种作为物种库, 然后计算群落内物种间的谱系距离, 通过统计方法检测其与随机模型下的谱系距离是否有显著差异来获得谱系结构.使用Student t- 检验, 确定不同海拔梯度(低、中、高)的群落系统发育结构是否显著不同于零模型.所有的NRI值和NTI值计算时未进行多度加权. ...
... 本研究中选取的4个群落多样性指数更多地是表现出了随海拔升高逐渐降低的变化趋势.这与施济普等(2005)对该地区的研究结果一致, 但与牛跃等(2008)得出的“单峰”格局不符.造成这一差异的原因可能是, 本研究样方设置中对低海拔地区群落样方的选取主要参考了施济普等(2005)的研究地点, 而未选择牛跃等(2008)研究中所涉及的位于山体下部(1,100-1,300m)、具有较低生物多样性的季风常绿阔叶林这一海拔区域, 因为在野外调查时我们发现该区域的季风常绿阔叶林破坏严重, 已经难以找到保存完好的原生林.PhyloSor指数显示, 群落间的相似性随着海拔差异的增加逐渐而降低的规律在哀牢山亚热带森林群落中依然适用, 与之前诸多研究结果也是一致的(如Soininen et al ., 2007 ; Bryant et al ., 2008 ), 表明群落系统发育beta多样性会随空间距离增大而降低. ...
The merging of community ecology and phylogenetic biology.
1
2009
... 哀牢山地处云贵高原、横断山脉和青藏高原三大自然地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地(牛跃等, 2008 ).哀牢山由于山体高大宽厚, 高差悬殊较大, 具有明显的垂直气候特征, 且海拔梯度升高引起的气候因子变化效应使得动物、植物以及土壤微生物等也相应呈现垂直分带现象(吴征镒等, 1983 ).哀牢山西坡的森林植被类型多样, 根据物种组成、植被结构、气候以及土壤环境等特征, 从海拔1,300-2,750 m大致可以划分为3个植被带: 山体中下部的亚热带季风常绿阔叶林(海拔高度1,300-2,100 m)、山体中部的中山湿性常绿阔叶林(海拔高度2,100-2,600 m)和山顶部的苔藓矮林(海拔高度2,600-2,750 m).这种显著的山体垂直分布格局是开展生物多样性沿海拔梯度变化研究的理想场所.本研究以对温度变化极为敏感的山地植被为研究对象(Walter, 1979 ; Cavender-Bares et al ., 2009 ), 开展哀牢山亚热带森林群落系统发育研究, 分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化, 从而推测群落构建的生态学过程. ...
A DNA barcode for land plants.
1
2009
... 使用天根公司的试剂盒完成所有植物样品的总DNA提取.选择叶绿体rbc L、mat K、基因间隔区trn H-psb A与核基因ITS等4个分子标记作为研究的扩增片段.PCR参照国际生命DNA条形码联盟植物工作组推荐的反应体系(CBOL Plant Working Group, 2009 ), 并作出优化和调整.扩增产物送北京六合华大基因科技有限公司(BGI)完成测序.得到的序列使用软件Sequencher 4.14(Gene Codes Corporation, Ann Arbor, Mich.)进行拼接和校对, rbc L、mat K和ITS等3个DNA片段在Geneious 6.1.5(Biomatters Ltd.)中选择Clustal X(Larkin et al. , 2007 )排序, 而trn H-psb A由于序列长度变化大, 用软件SATé(Liu et al. , 2009 )进行排序. ...
On the Origin of Species by Means of Natural Selection
1
1859
... 群落系统发育(community phylogeny)研究将系统发育生物学的方法运用到群落生态学中, 利用群落内物种的系统发育状况来推测历史因素对现有群落的影响, 通过分析群落内物种间的亲缘关系探究影响群落构建的主要原因(Webb, 2000 ; Webb et al ., 2002 ).群落系统发育研究主要是通过两个群落系统发育关系指数, 即净亲缘指数(net relatedness index, NRI)和最近亲缘指数(nearest taxon index, NTI), 将群落内物种间的系统发育距离和标准化零模型(即假设物种随机分布)下的系统发育距离进行比较, 统计二者间的差异, 用于检测群落是否存在系统发育结构.如果群落系统发育关系指数与零模型假设没有显著差异, 说明群落不存在系统发育结构或系统发育随机(phylogenetically randomness); 如果显著大于零模型假设, 说明群落具有系统发育结构, 表现为系统发育聚集(phylogeneticcally clustering); 相反, 如果显著小于零模型假设, 说明群落系统发育发散(phylogenetically over-dispersion).达尔文早在1859年就曾说过, 和属外的物种相比, 同属内的物种在生活习性和结构上具有更近的相似性, 当它们相互之间发生竞争时, 也比与属外物种的竞争激烈得多(Darwin, 1859 ).这一观点与群落系统发育研究中生态位保守的假设(即亲缘关系越近的物种, 生态特征越相似)是一致的.在一个群落里, 如果是生境筛选作用占主导地位, 那么相同生境将筛选出适应能力相似、亲缘关系偏近的物种, 表现为系统发育聚集; 相反, 竞争排斥作用会使生态位相似的物种无法共存于同一环境, 则群落内物种亲缘关系较远, 表现为系统发育发散.据此, 在生态性状保守的前提下, 我们就可以从群落现有的系统发育表现形式, 推测群落构建过程中的主导因素(表1 ). ...
Disturbance alters the phylogenetic composition and structure of plant communities in an old field system.
1
2009
... 哀牢山森林群落系统发育结构随着海拔梯度的上升由系统发育聚集走向发散或聚集程度降低, 呈现出非随机的状态, 说明中性理论预测群落随机的系统发育结构在哀牢山地区并不适用, 暗示在该地区森林中生态位过程对于群落的组成和维持更加重要.在生态位理论中, 生境过滤和竞争排斥是两个相反的过程, 前者使得物种间的系统发育结构相对聚集, 而后者使得亲缘关系近的物种不能共存, 系统发育结构相对发散(牛红玉等, 2011 ).哀牢山森林群落系统发育结构在低海拔表现出系统发育聚集, 说明生境过滤作用起着主导作用.对此有两种可能的解释: 其一, 低海拔地区受温度和水分等的影响比高海拔地区小得多, 地形异质性和生境异质性都较小, 促使进化程度相似的类群得以共存; 其二, 低海拔地区的森林群落受到的人为干扰较多(非破坏性的干扰), 干扰(如砍伐、火烧等)作为一种环境过滤器会促使群落系统发育聚集(Mo et al ., 2012), 这可能也是造成低海拔群落系统发育结构聚集的一个重要因子(Dinnage, 2009 ; 牛红玉等, 2011 ).中海拔地区出现了多种系统发育结构(聚集、发散和随机), 我们推测可能是由于随着海拔的上升, 中海拔的生境异质性要比低海拔复杂得多, 群落物种间会更倾向于采取不同的资源利用策略, 物种间的竞争作用会加剧, 同时存在生境过滤和竞争排斥两种生态过程的相互作用, 从而表现出了更多的系统发育结构. ...
Phylogenetic analysis of community assembly and structure over space and time.
1
2008
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
Systematics and conservation: on predicting the feature diversity of subsets of taxa.
2
1992
... 对群落系统发育多样性的分析采用系统发育多样性Faith’s PD指数(Faith, 1992 )和系统发育beta多样性PhyloSor指数来量化, 在R软件(R Development Core Team, 2005)中完成计算.系统发育多样性Faith’s PD指数是指某一样地中分类单元系统发育分支长度之和(Faith, 1992 ; Kemble et al. , 2010 ), 值越小, 表明该样地内物种的系统发育多样性在整个区域的系统发育多样性占有的比重越小, 在R程序的软件包picante中计算得到.系统发育beta多样性PhyloSor 指数是指两个群落共有物种的枝长和占两个群落所有物种的枝长和的比例, 计算公式为: ...
... )和系统发育beta多样性PhyloSor指数来量化, 在R软件(R Development Core Team, 2005)中完成计算.系统发育多样性Faith’s PD指数是指某一样地中分类单元系统发育分支长度之和(Faith, 1992 ; Kemble et al. , 2010 ), 值越小, 表明该样地内物种的系统发育多样性在整个区域的系统发育多样性占有的比重越小, 在R程序的软件包picante中计算得到.系统发育beta多样性PhyloSor 指数是指两个群落共有物种的枝长和占两个群落所有物种的枝长和的比例, 计算公式为: ...
Local and regional processes control species richness of plant communities: the species pool hypothesis.
1
2009
... 物种库和零模型的选择对群落系统发育生态学研究的结果有着很大影响(Kooyman et al. , 2011 ).物种库是指一个地区可进入某一特定群落的潜在物种数目(方精云等, 2009 ), 但在实际的研究中很难验证一个群落的实际物种库, 本研究中选取群落内所有物种作为小尺度物种库是目前来说相对合理的一种选择.Swenson等(2006)研究发现, 物种库的空间尺度越大, 越有可能发生系统发育聚集.在未来的研究中, 针对不同的研究区域范围和类群, 如何更为合理地选择物种库, 需要进一步改进.同时, 目前选择的零模型也不完善, 应该全面考虑群落中的物种丰富度和多度以及生态学特征, 并将进化特征和群落生态学性状特征结合起来(Kraft et al ., 2007 ).因此, 将来需要更多的研究来验证NRI和NTI指数的准确性并寻找更加合理的模型, 增加统计分析和解决问题的能力. ...
Global patterns in biodiversity.
1
2000
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
Phylogenetic and ecomorphological structure of assemblages of breeding leaf warblers (Phylloscopidae) along Himalayan elevational gradients.
2014
Phylogenetic structure in tropical hummingbird communities.
1
2009
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
Effects of different resource additions on species diversity in an annual plant community.
1
1990
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
Shifts in species and phylogenetic diversity between sapling and tree communities indicate negative density dependence in a lowland rain forest.
2010
An emerging synthesis between community ecology and evolutionary biology.
1
2007
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
Picante: R tools for integrating phylogenies and ecology.
1
2010
... 对群落系统发育多样性的分析采用系统发育多样性Faith’s PD指数(Faith, 1992 )和系统发育beta多样性PhyloSor指数来量化, 在R软件(R Development Core Team, 2005)中完成计算.系统发育多样性Faith’s PD指数是指某一样地中分类单元系统发育分支长度之和(Faith, 1992 ; Kemble et al. , 2010 ), 值越小, 表明该样地内物种的系统发育多样性在整个区域的系统发育多样性占有的比重越小, 在R程序的软件包picante中计算得到.系统发育beta多样性PhyloSor 指数是指两个群落共有物种的枝长和占两个群落所有物种的枝长和的比例, 计算公式为: ...
Altitude distribution of land birds in New Guinea.
1
1971
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
Trait evolution, community assembly, and the phylogenetic structure of ecological communities.
4
2007
... 不同生态性状进化特征和不同群落构建过程下的群落期望系统发育结构(参考Webb et al ., 2002 ; Kraft et al ., 2007 ) ...
... Patterns of community phylogenetic dispersion predicted to be produced by various community assembly processes and different evolutionary characteristic of ecological traits (From Webb et al ., 2002 ; Kraft et al ., 2007 ) ...
... 两个群落系统发育指数NRI和NTI反映出的高海拔地区群落(山顶苔藓矮林)的系统发育结构相反, NRI表现出系统发育聚集, 而NTI却表现为随机或发散.NRI和NTI是两个相互联系却又有不同的系统发育指数, 前者基于群落整体水平的亲缘关系, 反映发生在整个系统树上的系统发育模式, 而后者仅仅基于最近的系统发育关系, 主要针对系统进化树的末端(Kraft et al. , 2007 ).NRI和NTI不一致或相反的结果也出现在其他的研究中, 例如Bryant等(2008)发现这两个指数在检测高海拔地区的植物群落系统发育时表现不一致, 通过NRI指数的变化趋势, 认为在高海拔地区系统发育结构更趋向发散; Kress等(2009)在检测不同生境下不成熟森林的系统发育时出现了NRI小于零, 而NTI大于零的现象, 只是NTI的统计结果不显著(P = 0.17). ...
... 物种库和零模型的选择对群落系统发育生态学研究的结果有着很大影响(Kooyman et al. , 2011 ).物种库是指一个地区可进入某一特定群落的潜在物种数目(方精云等, 2009 ), 但在实际的研究中很难验证一个群落的实际物种库, 本研究中选取群落内所有物种作为小尺度物种库是目前来说相对合理的一种选择.Swenson等(2006)研究发现, 物种库的空间尺度越大, 越有可能发生系统发育聚集.在未来的研究中, 针对不同的研究区域范围和类群, 如何更为合理地选择物种库, 需要进一步改进.同时, 目前选择的零模型也不完善, 应该全面考虑群落中的物种丰富度和多度以及生态学特征, 并将进化特征和群落生态学性状特征结合起来(Kraft et al ., 2007 ).因此, 将来需要更多的研究来验证NRI和NTI指数的准确性并寻找更加合理的模型, 增加统计分析和解决问题的能力. ...
Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama.
2009
Advances in the use of DNA barcodes to build a community phylogeny for tropical trees in a Puerto Rican forest dynamics plot.
2010
Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests.
1
2011
... 物种库和零模型的选择对群落系统发育生态学研究的结果有着很大影响(Kooyman et al. , 2011 ).物种库是指一个地区可进入某一特定群落的潜在物种数目(方精云等, 2009 ), 但在实际的研究中很难验证一个群落的实际物种库, 本研究中选取群落内所有物种作为小尺度物种库是目前来说相对合理的一种选择.Swenson等(2006)研究发现, 物种库的空间尺度越大, 越有可能发生系统发育聚集.在未来的研究中, 针对不同的研究区域范围和类群, 如何更为合理地选择物种库, 需要进一步改进.同时, 目前选择的零模型也不完善, 应该全面考虑群落中的物种丰富度和多度以及生态学特征, 并将进化特征和群落生态学性状特征结合起来(Kraft et al ., 2007 ).因此, 将来需要更多的研究来验证NRI和NTI指数的准确性并寻找更加合理的模型, 增加统计分析和解决问题的能力. ...
Clustal W and Clustal X version 2.0.
1
2007
... 使用天根公司的试剂盒完成所有植物样品的总DNA提取.选择叶绿体rbc L、mat K、基因间隔区trn H-psb A与核基因ITS等4个分子标记作为研究的扩增片段.PCR参照国际生命DNA条形码联盟植物工作组推荐的反应体系(CBOL Plant Working Group, 2009 ), 并作出优化和调整.扩增产物送北京六合华大基因科技有限公司(BGI)完成测序.得到的序列使用软件Sequencher 4.14(Gene Codes Corporation, Ann Arbor, Mich.)进行拼接和校对, rbc L、mat K和ITS等3个DNA片段在Geneious 6.1.5(Biomatters Ltd.)中选择Clustal X(Larkin et al. , 2007 )排序, 而trn H-psb A由于序列长度变化大, 用软件SATé(Liu et al. , 2009 )进行排序. ...
Rapid and accurate large-scale coestimation of sequence alignments and phylogenetic trees.
1
2009
... 使用天根公司的试剂盒完成所有植物样品的总DNA提取.选择叶绿体rbc L、mat K、基因间隔区trn H-psb A与核基因ITS等4个分子标记作为研究的扩增片段.PCR参照国际生命DNA条形码联盟植物工作组推荐的反应体系(CBOL Plant Working Group, 2009 ), 并作出优化和调整.扩增产物送北京六合华大基因科技有限公司(BGI)完成测序.得到的序列使用软件Sequencher 4.14(Gene Codes Corporation, Ann Arbor, Mich.)进行拼接和校对, rbc L、mat K和ITS等3个DNA片段在Geneious 6.1.5(Biomatters Ltd.)中选择Clustal X(Larkin et al. , 2007 )排序, 而trn H-psb A由于序列长度变化大, 用软件SATé(Liu et al. , 2009 )进行排序. ...
Community structure and species diversity of hermatypic corals at Eilat, Red Sea.
1
1972
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
1
1994
... 依据样方调查材料, 采用物种丰富度(S )、Shannon-Wiener指数(H )、Simpson优势度指数(D )和Pielou均匀度指数(E )(马克平, 1994 ), 比较和评价哀牢山3种典型植被森林群落多样性的差异. ...
Elevational gradients in phylogenetic structure of ant communities reveal the interplay of biotic and abiotic constraints on diversity.
1
2011
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
Elevational gradients in diversity of small mammals.
1
2005
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
1
2010
... 构建方法主要参考Kress等(2010)的研究, 以APG III系统为大框架作“约束树”, 用4个分子标记得到的DNA分子序列来解决末端分类单元确切的系统位置.具体步骤是: 先将APG认可的物种名录提交到Phylomatic在线软件上运行, 获得哀牢山森林群落的简易系统发育关系, 以科为分类单元, 把科内的所有物种平行化(collapse), 形成APG系统的“约束树”; 再将4个DNA片段分别排序后合并成一个联合矩阵, 即DNA序列超矩阵; 然后把DNA超矩阵和“约束树”在CIPRES超级远程计算机(Miller et al ., 2010 )的RAxML程序上(Stamatakis, 2006 )进行最大似然方法ML分析, 并通过500次的重复来获得拓扑结构上的节点支持率, 将各节点的支持率附在拓扑结构最优的系统发育树上; 最后用r8s软件包(Sanderson, 2003 )非参数速率平滑法(NPRS)对系统发育树进行处理, 进而获得用于群落系统发育结构分析的进化树. ...
Change in phylogenetic community structure during succession of traditionally managed tropical rainforest in Southwest China.
2013
Species diversity of a stratified oak-hickory community.
1
1969
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
New progress in community assembly: community phylogenetic structure combining evolution and ecology.
3
2011
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
... 哀牢山森林群落系统发育结构随着海拔梯度的上升由系统发育聚集走向发散或聚集程度降低, 呈现出非随机的状态, 说明中性理论预测群落随机的系统发育结构在哀牢山地区并不适用, 暗示在该地区森林中生态位过程对于群落的组成和维持更加重要.在生态位理论中, 生境过滤和竞争排斥是两个相反的过程, 前者使得物种间的系统发育结构相对聚集, 而后者使得亲缘关系近的物种不能共存, 系统发育结构相对发散(牛红玉等, 2011 ).哀牢山森林群落系统发育结构在低海拔表现出系统发育聚集, 说明生境过滤作用起着主导作用.对此有两种可能的解释: 其一, 低海拔地区受温度和水分等的影响比高海拔地区小得多, 地形异质性和生境异质性都较小, 促使进化程度相似的类群得以共存; 其二, 低海拔地区的森林群落受到的人为干扰较多(非破坏性的干扰), 干扰(如砍伐、火烧等)作为一种环境过滤器会促使群落系统发育聚集(Mo et al ., 2012), 这可能也是造成低海拔群落系统发育结构聚集的一个重要因子(Dinnage, 2009 ; 牛红玉等, 2011 ).中海拔地区出现了多种系统发育结构(聚集、发散和随机), 我们推测可能是由于随着海拔的上升, 中海拔的生境异质性要比低海拔复杂得多, 群落物种间会更倾向于采取不同的资源利用策略, 物种间的竞争作用会加剧, 同时存在生境过滤和竞争排斥两种生态过程的相互作用, 从而表现出了更多的系统发育结构. ...
... ; 牛红玉等, 2011 ).中海拔地区出现了多种系统发育结构(聚集、发散和随机), 我们推测可能是由于随着海拔的上升, 中海拔的生境异质性要比低海拔复杂得多, 群落物种间会更倾向于采取不同的资源利用策略, 物种间的竞争作用会加剧, 同时存在生境过滤和竞争排斥两种生态过程的相互作用, 从而表现出了更多的系统发育结构. ...
Community assembly: the relative importance of neutral theory and niche theory.
1
2009
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
Vertical distribution patterns of plant species diversity in Ailao Mountains.
2
2008
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
... 哀牢山地处云贵高原、横断山脉和青藏高原三大自然地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地(牛跃等, 2008 ).哀牢山由于山体高大宽厚, 高差悬殊较大, 具有明显的垂直气候特征, 且海拔梯度升高引起的气候因子变化效应使得动物、植物以及土壤微生物等也相应呈现垂直分带现象(吴征镒等, 1983 ).哀牢山西坡的森林植被类型多样, 根据物种组成、植被结构、气候以及土壤环境等特征, 从海拔1,300-2,750 m大致可以划分为3个植被带: 山体中下部的亚热带季风常绿阔叶林(海拔高度1,300-2,100 m)、山体中部的中山湿性常绿阔叶林(海拔高度2,100-2,600 m)和山顶部的苔藓矮林(海拔高度2,600-2,750 m).这种显著的山体垂直分布格局是开展生物多样性沿海拔梯度变化研究的理想场所.本研究以对温度变化极为敏感的山地植被为研究对象(Walter, 1979 ; Cavender-Bares et al ., 2009 ), 开展哀牢山亚热带森林群落系统发育研究, 分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化, 从而推测群落构建的生态学过程. ...
Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants.
1
2005
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
Exploring tree-habitat associations in a Chinese subtropical forest plot using a molecular phylogeny generated from DNA barcode loci.
1
2011
... DNA条形码的快速兴起, 极大地促进了群落系统发育生态学的发展, 为精确地构建群落系统发育关系提供了新的技术.其中比较典型的研究, 如Kress等(2009)发现由DNA条形码建立的系统发育树为末端的低阶分类阶元提供了更为准确的系统位置, 并且得到的物种系统发育关系增加了拒绝零假设的概率; Gonzalez等(2010)认为, DNA条形码所构建的物种间精确的系统发育关系, 可以提高有关群落结构计算的统计力; Pei等(2011)和裴男才(2012)综合使用进化速率不同的DNA条形码片段, 相对准确地构建出鼎湖山南亚热带森林植物群落的系统发育关系.然而, Kress等(2010)在亚热带地区波多黎各森林样地的研究发现, 单独利用DNA 条形码序列所构建的群落系统发育关系与APG系统(APG III, 2009 )相比, 在较高的分类阶元即目水平上存在较大的出入, 需要利用APG系统的大框架作为约束树(constraint tree), 并对各分支末端的类群配合使用DNA条形码的建树方法, 才能获得群落物种间精确的系统发育关系(裴男才等, 2011 ). ...
Plant DNA barcodes promote the development of phylogenetic community ecology.
1
2011
... 在最初的群落系统发育研究中, 多数学者主要是通过Phylomatic方法(Webb & Donoghue, 2005 )来探究群落内所有物种间的系统发育关系, 但这种方法也存在一些不足.首先, 采用Phylomatic方法所构建的系统发育树往往具有明显的多分支结构(polytomy), 尤其是当同一科属所含物种数目较多时这种现象更加明显; 其次, 基于APG系统构建的发育树缺乏裸子植物的信息, 而有些裸子植物也是群落建群种, 如云南的云南松(Pinus yunnanensis )林和长白山的阔叶红松(P. koraiensis )林等(裴男才等, 2011b ). ...
Building a subtropical forest community phylogeny based on plant DNA barcodes from Dinghushan plot.
2012
Introduction to Mathematical Ecology. Wiley-Interscience
1969
The relationship among area, elevation, and regional species richness in Neotropical birds.
1
1997
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2013. URL
2013
r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock.
1
2003
... 构建方法主要参考Kress等(2010)的研究, 以APG III系统为大框架作“约束树”, 用4个分子标记得到的DNA分子序列来解决末端分类单元确切的系统位置.具体步骤是: 先将APG认可的物种名录提交到Phylomatic在线软件上运行, 获得哀牢山森林群落的简易系统发育关系, 以科为分类单元, 把科内的所有物种平行化(collapse), 形成APG系统的“约束树”; 再将4个DNA片段分别排序后合并成一个联合矩阵, 即DNA序列超矩阵; 然后把DNA超矩阵和“约束树”在CIPRES超级远程计算机(Miller et al ., 2010 )的RAxML程序上(Stamatakis, 2006 )进行最大似然方法ML分析, 并通过500次的重复来获得拓扑结构上的节点支持率, 将各节点的支持率附在拓扑结构最优的系统发育树上; 最后用r8s软件包(Sanderson, 2003 )非参数速率平滑法(NPRS)对系统发育树进行处理, 进而获得用于群落系统发育结构分析的进化树. ...
Characteristics and species composition of main vegetation types on west slope of the Ailao Mountains in Yunnan.
2005
Measurement of diversity.
1949
The distance decay of similarity in ecological communities.
1
2007
... 本研究中选取的4个群落多样性指数更多地是表现出了随海拔升高逐渐降低的变化趋势.这与施济普等(2005)对该地区的研究结果一致, 但与牛跃等(2008)得出的“单峰”格局不符.造成这一差异的原因可能是, 本研究样方设置中对低海拔地区群落样方的选取主要参考了施济普等(2005)的研究地点, 而未选择牛跃等(2008)研究中所涉及的位于山体下部(1,100-1,300m)、具有较低生物多样性的季风常绿阔叶林这一海拔区域, 因为在野外调查时我们发现该区域的季风常绿阔叶林破坏严重, 已经难以找到保存完好的原生林.PhyloSor指数显示, 群落间的相似性随着海拔差异的增加逐渐而降低的规律在哀牢山亚热带森林群落中依然适用, 与之前诸多研究结果也是一致的(如Soininen et al ., 2007 ; Bryant et al ., 2008 ), 表明群落系统发育beta多样性会随空间距离增大而降低. ...
RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models.
1
2006
... 构建方法主要参考Kress等(2010)的研究, 以APG III系统为大框架作“约束树”, 用4个分子标记得到的DNA分子序列来解决末端分类单元确切的系统位置.具体步骤是: 先将APG认可的物种名录提交到Phylomatic在线软件上运行, 获得哀牢山森林群落的简易系统发育关系, 以科为分类单元, 把科内的所有物种平行化(collapse), 形成APG系统的“约束树”; 再将4个DNA片段分别排序后合并成一个联合矩阵, 即DNA序列超矩阵; 然后把DNA超矩阵和“约束树”在CIPRES超级远程计算机(Miller et al ., 2010 )的RAxML程序上(Stamatakis, 2006 )进行最大似然方法ML分析, 并通过500次的重复来获得拓扑结构上的节点支持率, 将各节点的支持率附在拓扑结构最优的系统发育树上; 最后用r8s软件包(Sanderson, 2003 )非参数速率平滑法(NPRS)对系统发育树进行处理, 进而获得用于群落系统发育结构分析的进化树. ...
Historical processes enhance patterns of diversity along latitudinal gradients.
1
2006
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
The problem and promise of scale dependency in community phylogenetics.
2006
Bird species diversity on an Andean elevation gradient.
1
1977
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
High species diversity and abundance of the epibenthic community in an oxygen-deficient basin.
1
1981
... 生物多样性维持机制即群落构建(community assembly)的成因, 一直以来都是群落生态学研究领域的热点和焦点之一(牛克昌等, 2009 ).生物群落的形成和维持是一个长期的演化过程, 是进化过程与生态过程共同作用的结果(Johnson & Stinchcombe, 2007 ; 牛红玉等, 2011 ).在20世纪90年代之前, 生态学家主要从群落的环境条件和群落内的生态学过程(如捕食、竞争、互利共生、干扰等)来解释现有群落的结构、组成和多样性等特征, 几乎不考虑群落构建过程中的历史因素(Monk et al. , 1969 ; Loya, 1972 ; Tunnicliffe, 1981 ; Goldberg & Miller, 1990 ).但是, 缺乏对群落构建历程具有重要影响的历史因素的研究, 去探讨并解释群落构建的成因显然是不全面的. ...
Investigation of temperature and aridity at different elevations of Mt. Ailao, SW China.
1
2013
... 有研究显示, 哀牢山高海拔的山顶苔藓矮林由于承受较强的紫外线辐射和大风等环境胁迫, 小叶所占的比例要比中山湿性常绿阔叶林高(①施济普 (2007) 云南山顶苔藓矮林群落生态学与生物地理学研究. 硕士论文, 中国科学院西双版纳热带植物园, 昆明.); 同时, You等(2013) 对哀牢山的山顶苔藓矮林开展了微气象学研究, 发现植物在高海拔地区承受了更强的干旱胁迫.以上研究结果反映在高强度的环境压力下, 高海拔地区的物种可能发生了趋同进化.然而, 生境过滤后虽聚集了性状相似的物种, 但它们之间的亲缘关系并不相近, 表现为谱系发散; 而另外一些因竞争排斥作用留下的具有不同性状的物种间亲缘关系可能会没有规律, 表现为谱系随机或者谱系聚集, 正如本研究NRI指数所反映的结果. ...
Emerging patterns in the comparative analysis of phylogenetic community structure.
1
2009
... 现在越来越多的研究采用群落系统发育的方法, 较好地解释了群落成分的空间变化(Stevens, 2006 ; Emerson & Gillespie, 2008 ; Vamosi et al ., 2009 ).其中一些研究从生物群落的系统发育角度来推测多样性沿海拔变化的驱动因素(Bryant et al ., 2008 ; Graham et al ., 2009 ; Machac et al ., 2011 ).如Bryant等(2008)检测了北美落基山脉沿海拔梯度5个地点的微生物和植物群落的系统发育结构, 发现微生物群落全部表现为系统发育聚集, 而植物群落却是随着海拔的升高由聚集走向发散, 并且发散程度越来越显著; 而Graham等(2009)在厄瓜多尔的安第斯山脉对189个蜂鸟的研究、Machac等(2011)对3个温带山地蚂蚁群落的研究却表明, 群落系统发育结构在高海拔地区表现为系统发育聚集, 在低海拔地区表现为系统发育发散, 揭示了物种间竞争作用的存在.虽然上述研究得出了截然相反的结论, 但是均证实群落系统发育结构沿海拔梯度存在某种显著的变化, 说明在生态群落的构建过程中有非随机过程起着促进乃至关键作用, 这就为探讨生物多样性格局的形成与维持机制、物种的共存与环境适应以及物种灭绝的后果提供了新的角度和依据. ...
Vegetation of the Earth. Springer
2
1979
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
... 哀牢山地处云贵高原、横断山脉和青藏高原三大自然地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地(牛跃等, 2008 ).哀牢山由于山体高大宽厚, 高差悬殊较大, 具有明显的垂直气候特征, 且海拔梯度升高引起的气候因子变化效应使得动物、植物以及土壤微生物等也相应呈现垂直分带现象(吴征镒等, 1983 ).哀牢山西坡的森林植被类型多样, 根据物种组成、植被结构、气候以及土壤环境等特征, 从海拔1,300-2,750 m大致可以划分为3个植被带: 山体中下部的亚热带季风常绿阔叶林(海拔高度1,300-2,100 m)、山体中部的中山湿性常绿阔叶林(海拔高度2,100-2,600 m)和山顶部的苔藓矮林(海拔高度2,600-2,750 m).这种显著的山体垂直分布格局是开展生物多样性沿海拔梯度变化研究的理想场所.本研究以对温度变化极为敏感的山地植被为研究对象(Walter, 1979 ; Cavender-Bares et al ., 2009 ), 开展哀牢山亚热带森林群落系统发育研究, 分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化, 从而推测群落构建的生态学过程. ...
Species diversity of plant communities along an altitudinal gradient in the middle section of northern slopes of Qilian Mountains, Zhangye, Gansu.
1
2002
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
Exploring the phylogenetic structure of ecological communities: an example for rain forest trees.
1
2000
... 群落系统发育(community phylogeny)研究将系统发育生物学的方法运用到群落生态学中, 利用群落内物种的系统发育状况来推测历史因素对现有群落的影响, 通过分析群落内物种间的亲缘关系探究影响群落构建的主要原因(Webb, 2000 ; Webb et al ., 2002 ).群落系统发育研究主要是通过两个群落系统发育关系指数, 即净亲缘指数(net relatedness index, NRI)和最近亲缘指数(nearest taxon index, NTI), 将群落内物种间的系统发育距离和标准化零模型(即假设物种随机分布)下的系统发育距离进行比较, 统计二者间的差异, 用于检测群落是否存在系统发育结构.如果群落系统发育关系指数与零模型假设没有显著差异, 说明群落不存在系统发育结构或系统发育随机(phylogenetically randomness); 如果显著大于零模型假设, 说明群落具有系统发育结构, 表现为系统发育聚集(phylogeneticcally clustering); 相反, 如果显著小于零模型假设, 说明群落系统发育发散(phylogenetically over-dispersion).达尔文早在1859年就曾说过, 和属外的物种相比, 同属内的物种在生活习性和结构上具有更近的相似性, 当它们相互之间发生竞争时, 也比与属外物种的竞争激烈得多(Darwin, 1859 ).这一观点与群落系统发育研究中生态位保守的假设(即亲缘关系越近的物种, 生态特征越相似)是一致的.在一个群落里, 如果是生境筛选作用占主导地位, 那么相同生境将筛选出适应能力相似、亲缘关系偏近的物种, 表现为系统发育聚集; 相反, 竞争排斥作用会使生态位相似的物种无法共存于同一环境, 则群落内物种亲缘关系较远, 表现为系统发育发散.据此, 在生态性状保守的前提下, 我们就可以从群落现有的系统发育表现形式, 推测群落构建过程中的主导因素(表1 ). ...
Phylogenies and community ecology.
3
2002
... 群落系统发育(community phylogeny)研究将系统发育生物学的方法运用到群落生态学中, 利用群落内物种的系统发育状况来推测历史因素对现有群落的影响, 通过分析群落内物种间的亲缘关系探究影响群落构建的主要原因(Webb, 2000 ; Webb et al ., 2002 ).群落系统发育研究主要是通过两个群落系统发育关系指数, 即净亲缘指数(net relatedness index, NRI)和最近亲缘指数(nearest taxon index, NTI), 将群落内物种间的系统发育距离和标准化零模型(即假设物种随机分布)下的系统发育距离进行比较, 统计二者间的差异, 用于检测群落是否存在系统发育结构.如果群落系统发育关系指数与零模型假设没有显著差异, 说明群落不存在系统发育结构或系统发育随机(phylogenetically randomness); 如果显著大于零模型假设, 说明群落具有系统发育结构, 表现为系统发育聚集(phylogeneticcally clustering); 相反, 如果显著小于零模型假设, 说明群落系统发育发散(phylogenetically over-dispersion).达尔文早在1859年就曾说过, 和属外的物种相比, 同属内的物种在生活习性和结构上具有更近的相似性, 当它们相互之间发生竞争时, 也比与属外物种的竞争激烈得多(Darwin, 1859 ).这一观点与群落系统发育研究中生态位保守的假设(即亲缘关系越近的物种, 生态特征越相似)是一致的.在一个群落里, 如果是生境筛选作用占主导地位, 那么相同生境将筛选出适应能力相似、亲缘关系偏近的物种, 表现为系统发育聚集; 相反, 竞争排斥作用会使生态位相似的物种无法共存于同一环境, 则群落内物种亲缘关系较远, 表现为系统发育发散.据此, 在生态性状保守的前提下, 我们就可以从群落现有的系统发育表现形式, 推测群落构建过程中的主导因素(表1 ). ...
... 不同生态性状进化特征和不同群落构建过程下的群落期望系统发育结构(参考Webb et al ., 2002 ; Kraft et al ., 2007 ) ...
... Patterns of community phylogenetic dispersion predicted to be produced by various community assembly processes and different evolutionary characteristic of ecological traits (From Webb et al ., 2002 ; Kraft et al ., 2007 ) ...
Phylomatic: tree assembly for applied phylogenetics.
1
2005
... 在最初的群落系统发育研究中, 多数学者主要是通过Phylomatic方法(Webb & Donoghue, 2005 )来探究群落内所有物种间的系统发育关系, 但这种方法也存在一些不足.首先, 采用Phylomatic方法所构建的系统发育树往往具有明显的多分支结构(polytomy), 尤其是当同一科属所含物种数目较多时这种现象更加明显; 其次, 基于APG系统构建的发育树缺乏裸子植物的信息, 而有些裸子植物也是群落建群种, 如云南的云南松(Pinus yunnanensis )林和长白山的阔叶红松(P. koraiensis )林等(裴男才等, 2011b ). ...
Phylogenetic history underlies elevational biodiversity patterns in tropical salamanders.
1
2007
... 生物多样性在大陆(continent)或地区(region)尺度上随海拔的升高会出现较大的变化(Wiens et al ., 2007 ).早期的研究认为, 这种海拔梯度变化格局与纬度梯度变化格局相似, 即随着海拔的上升生物多样性降低(Kikkawa & Williams, 1971 ; Terborgh, 1977 ).但是之后在不同地区开展的大量研究却发现, 随着海拔高度的不断升高, 物种丰富度呈现“单峰”分布的格局(Rahbek, 1997 ; 王国宏, 2002 ; McCain, 2005 ; Oomen & Shanker, 2005 ; 牛跃等, 2008 ).海拔作为影响山地森林群落结构和物种组成的重要因素之一, 集合了温度、水分、风力、光照和土壤等诸多生态因子的综合影响(Gaston, 2000 ), 并且在海拔梯度上温度的变化速率要比在纬度梯度上快1,000倍(Walter, 1979 ).研究生物在不同海拔梯度下的变化规律, 对于揭示植物群落的环境梯度变化规律、了解植物适应性和物种多样性沿海拔梯度的变化趋势等具有重要意义. ...
1
1983
... 哀牢山地处云贵高原、横断山脉和青藏高原三大自然地理区域的结合部, 是生物多样性较丰富及植物区系地理成分荟萃之地(牛跃等, 2008 ).哀牢山由于山体高大宽厚, 高差悬殊较大, 具有明显的垂直气候特征, 且海拔梯度升高引起的气候因子变化效应使得动物、植物以及土壤微生物等也相应呈现垂直分带现象(吴征镒等, 1983 ).哀牢山西坡的森林植被类型多样, 根据物种组成、植被结构、气候以及土壤环境等特征, 从海拔1,300-2,750 m大致可以划分为3个植被带: 山体中下部的亚热带季风常绿阔叶林(海拔高度1,300-2,100 m)、山体中部的中山湿性常绿阔叶林(海拔高度2,100-2,600 m)和山顶部的苔藓矮林(海拔高度2,600-2,750 m).这种显著的山体垂直分布格局是开展生物多样性沿海拔梯度变化研究的理想场所.本研究以对温度变化极为敏感的山地植被为研究对象(Walter, 1979 ; Cavender-Bares et al ., 2009 ), 开展哀牢山亚热带森林群落系统发育研究, 分析不同植被类型的森林群落系统发育结构和沿海拔梯度的变化, 从而推测群落构建的生态学过程. ...

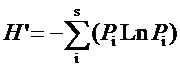
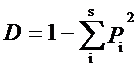
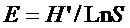
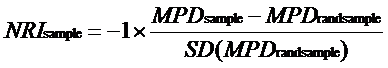
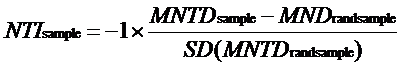


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}