


太湖是我国第三大淡水湖, 位于长江三角洲太湖平原上, 介于30°55′40″-31°32′58″N, 119°52′32″- 120°36′10″E之间, 面积2,338 km2, 平均水深1.9 m(秦伯强等, 2004)。太湖又是我国富营养化水平严重, 蓝藻频频暴发的湖泊, 是国家重点治理的富营养水体之一。在20世纪60年代太湖尚属Ⅰ-Ⅱ类水体, 70年代发展至Ⅱ类, 80年代初平均为Ⅱ-Ⅲ类, 80年代末全面进入Ⅲ类, 局部地区Ⅳ类和Ⅴ类, 北部的梅梁湾开始频繁暴发蓝藻水华, 到2000年太湖85%的湖面已为Ⅳ类水, 蓝藻水华的暴发也从以前的局部区域向全太湖扩展(黄漪平, 2001; 秦伯强, 2002; Qin et al., 2007)。2007年的监测结果显示太湖湖心已出现严重的蓝藻水华(Duan, 2009)。近几年来, 太湖水华暴发的趋势有增无减(朱广伟, 2008), 严重阻碍了周边地区的生产生活活动, 也对湖泊生态系统生物类群产生极大的影响。
底栖动物作为湖泊生态系统的重要类群, 一直为淡水生态学家所关注。众多研究表明大型底栖动物在湖泊生态系统中发挥着重要的功能, 它可以加速水底碎屑的分解, 促进泥水界面的物质交换和水体的自净, 在生态系统物质循环和能量流动中起着重要作用(Lindegaard, 1994; Covich et al., 1999; Vanni, 2002)。因此, 研究大型底栖动物的种类组成、群落结构、时空变化以及物种多样性等特征, 对合理利用湖泊资源, 改善湖泊水质具有重要意义。
以往对于太湖大型底栖动物群落有过几次调查(中国科学院南京地理研究所, 1965; 孙超白和周凤帆, 1983; 黄漪平, 2001; 秦伯强等, 2004), 但调查频率低且多局限于少数湖湾, 未能全面系统地阐述太湖大型底栖动物的群落结构和空间分布特征。我们于2007年2月至2008年11月对太湖大型底栖动物进行了为期两年的调查, 分析各类群底栖动物及优势种在太湖的空间分布特征, 初步探讨导致各湖区大型底栖动物群落结构差异的原因。
1 材料与方法
1.1 采样点布设与样品采集
均匀布设30个采样点(图1), 其中TH0、TH6、TH10位于太湖3条主要河流入湖口。于2007年2月至2008年11月每季度采样一次, 分别为每年的2月、5月、8月和11月; 样品定量采集用1/40 m2改良彼得逊采泥器, 每个点视情况采集1-2次。泥样过60目尼龙筛洗净后, 在解剖盘中逐一将底栖动物拣出。标本用10%福尔马林溶液保存。在实验室中将标本鉴定至尽可能低的分类单元(刘月英等, 1979; Morse et al., 1994; 王洪铸, 2002), 并确定其功能摄食类群 (Morse et al., 1994; 梁彦龄和王洪铸, 2000; 刘学勤, 2006), 然后用滤纸吸去表面固定液, 置于电子天平上称重, 并将结果折算成单位面积的密度和生物量。
图1
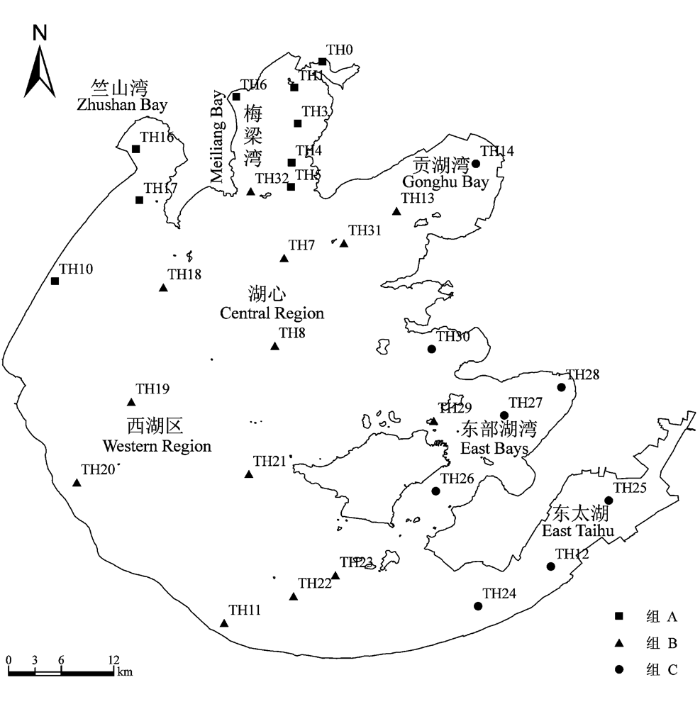
图1
太湖大型底栖动物采样点及聚类结果分布图
Fig. 1
Locations of the macrozoobenthos sampling sites in Lake Taihu and showing the spatial distribution of affinity groups
1.2 数据分析
采用相对重要性指数(index of relative importance, IRI)确定底栖动物优势种类(韩洁等, 2004):
式中W为相对生物量, 即某一物种的生物量占大型底栖动物总生物量的百分比; N为相对丰度, 即该物种的丰度占大型底栖动物总丰度的百分比; F为该物种出现的频率。
采用ArcGIS 9.2软件模拟底栖动物的空间分布。寡毛纲颤蚓科是典型的耐污种, 其对环境具有重要的指示作用(梁彦龄和王洪铸, 2000), 数据分析时发现其在太湖密度较高, 因此在分析各类群底栖动物空间分布时将其单独作为一个类群, 另外将软体动物双壳类和腹足类也分开计算。
基于各采样点底栖动物的Bray-Curtis相似性测度系数, 采用非加权成对平均法(unweighted pair group method using arithmetic averages)对30个采样点底栖动物密度进行聚类分析。由于不同种类密度差异较大, 分析时将密度对数标准化以消除量纲的差别。基于聚类分析结果, 利用ANOSIM (one-way analysis of similarities)检验各聚类组群落结构的差异性, 并利用SIMPER分析(Similarity percentages)找出对组内相似性贡献最大的种类, 分析软件用PRIMER 5.0 (Clarke, 1993; 周红和张志南, 2002)。
用K-优势曲线来评估各组大型底栖动物的多样性(Warwick, 1986), 同时结合以下2个多样性指数比较各聚类组的多样性:
Shannon-Wiener多样性指数 (Shannon & Wiener, 1949):
Pielou均匀度指数(Pielou, 1975):
式中S为群落内的种类总数, ni为第i个种的个体数, N表示所有种类总个体数。
2 结果
2.1 种类组成
本次调查共鉴定大型底栖动物40种(表1), 隶属于3门7纲19科。其中寡毛类2科8属10种(25.0%), 摇蚊幼虫10属11种(27.5%), 软体动物腹足类5科7属7种(17.5%)、双壳类4科5属5种(12.5%), 其他类7种(17.5%)。平均每个采样点采集到大型底栖动物13种, 最小值出现在TH8(7种), 最大值出现在TH26 (23种)。按功能摄食类群分, 其中直接收集者19种(占总物种数的47.5%), 捕食者和刮食者各7种(分别占17.5%), 过滤收集者5种(12.5%), 撕食者2种(5%)。
表1 太湖大型底栖动物种类组成及相对重要性指数
Table 1
| 物种 Species | 相对重要性指数 IRI | 物种 Species | 相对重要性指数 IRI |
|---|---|---|---|
| 寡毛纲 Oligochaeta | 椎实螺科 Lymnaeidae | ||
| 仙女虫科 Naididae | 椭圆萝卜螺 Radix swinhoei | <1 | |
| 七鳃管盘虫 Aulophorus heptabranchiata | 17 | 扁蜷螺科 Planorbidae | |
| 指鳃尾盘虫 Dero digitata | 21 | 凸旋螺 Gyraulus convexiusculus | <1 |
| 参差仙女虫 Nais variabilis | 10 | 黑螺科 Melaniidae | |
| 颤蚓科 Tubificidae | 方格短沟蜷 Semisulcospira cancellata | 12 | |
| 苏氏尾鳃蚓 Branchiura sowerbyi | 36 | 田螺科 Viviparidae | |
| 霍甫水丝蚓 Limnodrilus hoffmeisteri | 2,077 | 铜锈环棱螺 Bellamya aeruginosa | 535 |
| 巨毛水丝蚓 L. grandisetosus | 6 | 摇蚊幼虫 Chironomidae | |
| 水丝蚓属一种 Limnodrilus sp. | 7 | 半折摇蚊 Chironomus semireductus | 8 |
| 中华河蚓 Rhyacodrilus sinicus | 109 | 羽摇蚊 C. plumosus | <1 |
| 嫩丝蚓属一种 Teneridrilus sp. | <1 | 褐斑菱跗摇蚊 Clinotanypus sugiyamai | <1 |
| 颤蚓科一种 Tubificidae sp. | 三带环足摇蚊 Cricotopus trifasciatus | <1 | |
| 软体动物 Mollusca | 指突隐摇蚊 Cryptochironomus digitatus | <1 | |
| 双壳纲 Bivalvia | 侧叶雕翅摇蚊 Glyptotendipes lobiferus | <1 | |
| 蚬科 Corbiculidae | 小摇蚊属一种 Microchironomus sp. | 1 | |
| 河蚬 Corbicula fluminea | 1,788 | 多足摇蚊属一种 Polypedilum sp. | <1 |
| 截蛏科 Solecurtidae | 花翅前突摇蚊 Proclakius choreus | 2 | |
| 中国淡水蛏 Novaculina chinensis | <1 | 中国长足摇蚊 Tanypus chinensis | 46 |
| 球蚬科 Sphaeriidae | 长跗摇蚊属一种 Tanytarsus sp. | <1 | |
| 湖球蚬 Sphaerium lacustre | 4 | 其他 Others | |
| 蚌科 Unionidae | 端足目一种 Amphipoda sp. | 4 | |
| 背角无齿蚌 Anodonta woodiana woodiana | * | 虫怱属一种 Caenagrion sp. | <1 |
| 背瘤丽蚌 Lamprotula leai | * | 扁舌蛭 Glossiphonia complanata | <1 |
| 腹足纲 Gastropod | 钩虾属一种 Gammarus sp. | 137 | |
| 觿螺科 Hydrobiidae | 寡鳃齿吻沙蚕 Nephtys oligobranchia | 7 | |
| 长角涵螺 Alocinma longicornis | 13 | 多毛纲一种 Polychatea sp.1 | 41 |
| 纹沼螺 Parafossarulus striatulus | 6 | 多毛纲一种 Polychatea sp.2 | 3 |
| 光滑狭口螺 Stenothyra glabra | 6 |
* 计算相对重要性指数时, 在240份样品中出现次数小于8的种类被剔除。
* When calculated index of relative importance (IRI), the species occurred less than 8 samples in the 240 samples were excluded.
2.2 空间分布格局
图2
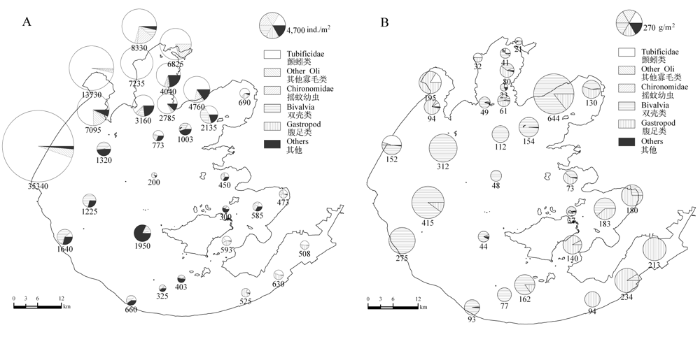
图2
太湖大型底栖动物平均密度(A, ind./m2)和生物量(B, g/m2)空间分布及各类群底栖动物所占比例。Other_Oli代表除颤蚓科之外的寡毛类。
Fig. 2
Spatial distribution of mean density (A, ind./m2) and biomass (B, g/m2) and the proportion of different groups in 30 sampling sites. Other_Oli: species of Oligochaeta excluding Tubificidae.
图3
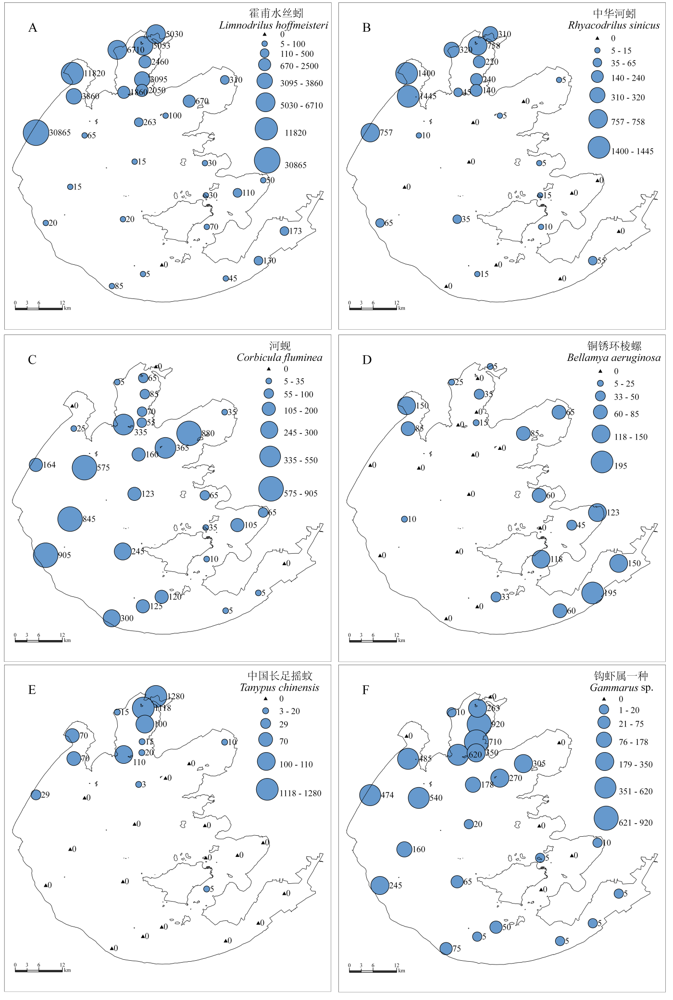
图3
太湖底栖动物优势种密度(ind./m2)空间分布格局
Fig. 3
Spatial distribution patterns of density (ind./m2) for six dominant species in Lake Taihu
软体动物主要分布在贡湖湾、西湖区、东太湖及东部湖湾(图2B), 河蚬(Corbicula fluminea)和铜锈环棱螺(Bellamya aeruginosa)是软体动物的优势种(共占软体动物总密度的79.1%), 但空间分布格局差异较大(图3C, D)。河蚬在贡湖湾和西湖区密度最高, 在东太湖和竺山湾最低(< 25 ind./m2), 而铜锈环棱螺在这两个区域的密度却最高(平均128 ind./m2), 同时在东部湖湾及贡湖湾密度也较高(83 ind./m2), 而湖心和西湖区基本无铜锈环棱螺分布(< 10 ind./m2)。摇蚊幼虫在太湖大型底栖动物总密度所占比例较低(3.6%), 且主要分布在梅梁湾、竺山湾及东太湖和东部湖湾。其中梅梁湾、竺山湾种类较少,但密度较高, 其优势种为中国长足摇蚊(Tanypus chinensis), 占摇蚊总密度的72.3%, 平均密度336 ind./m2(图3E); 东太湖和东部湖湾摇蚊幼虫密度 较低(<34 ind./m2), 但种类丰富度相对较高, 且出 现了对水质要求较高的种类如长跗摇蚊属一种(Tanytarsus sp.)和指突隐摇蚊(Cryptochironomus digitatus), 而这两种摇蚊在梅梁湾和竺山湾均未发现。其他种类中优势度较高的种类是钩虾属一种(Gammarus sp.), 平均密度达到192 ind./m2, 主要分布在梅梁湾、贡湖湾及西湖区, 而在东部湖湾和东太湖密度很低(< 10 ind./m2) (图3F)。调查中还发现一些河口种类, 如多毛类的寡鳃齿吻沙蚕(Nephtys oligobranchia), 其在西湖区密度较高, 最高值可达170 ind./m2(TH11), 其在太湖的分布说明太湖与长江之间有着物种的交流。
2.3 聚类及多样性分析
在50%的相似性水平上, 聚类分析将太湖30个采样点分成3组(图4)。组A包括9个采样点, 结合采样点分布图发现其全部位于梅梁湾、竺山湾以及河口(图1), 其优势种为霍甫水丝蚓、中华河蚓、钩虾属一种、指鳃尾盘虫(Dero digitata)、中国长足摇蚊、苏氏尾鳃蚓(Branchiura sowerbyi)和半折摇蚊(Chironomus semireductus), SIMPER分析表明这7个种类对组内相似性贡献率最大(累积达到91.2%) (表2)。组B包括的13个采样点位于贡湖湾、湖心及西湖区, 其优势种主要是河蚬、钩虾属一种、多毛类一种(Polychaeta sp.1)、霍甫水丝蚓和苏氏尾鳃蚓, 其对组内相似性的累积贡献率达80.3%。组C的8个采样点除TH14外全部位于东太湖和东部湖湾, 其优势种为霍甫水丝蚓、苏氏尾鳃蚓、河蚬以及腹足纲螺类的一些种类(表2), 其对组内相似性的贡献率为84.8%。值得注意的是, 虽然霍甫水丝蚓在组B和组C也为优势种之一, 但其密度与组A相比却低很多, 且由于个体较小, 在总生物量上所占比例较低。
图4

图4
基于30个采样点大型底栖动物密度的Bray-Curtis相似性系数的聚类分析树状图
Fig. 4
Dendrogram showing the hierarchical clustering of 30 sampling sites in Lake Taihu based on Bray-Curtis similarity coefficients
表2 各聚类组大型底栖动物优势种平均密度(ind./m2)及其对组内相似性贡献百分比
Table 2
| 物种 Species | Group A | Group B | Group C |
|---|---|---|---|
| 寡毛类 Oligochaeta | |||
| 霍甫水丝蚓 Limnodrilus hoffmeisteri | 7,883(50.29%) | 242(10.08%) | 115(18.46%) |
| 中华河蚓 Rhyacodrilus sinicus | 621(14.6%) | ||
| 苏氏尾鳃蚓 Branchiura sowerbyi | 51(4.97%) | 23(5.69%) | 44(10.12%) |
| 指鳃尾盘虫 Dero digitata | 211(5.79%) | ||
| 软体动物 Mollusca | |||
| 河蚬 Corbicula fluminea | 386(30.34%) | 36(6.8%) | |
| 铜锈环棱螺 Bellamya aeruginosa | 102(19.73%) | ||
| 方格短沟蜷 Semisulcospira cancellata | 19(5.43%) | ||
| 光滑狭口螺 Stenothyra glabra | 11(3.56%) | ||
| 长角涵螺 Alocinma longicornis | 50(14.13%) | ||
| 纹沼螺 Parafossarulus striatulus | 31(6.57%) | ||
| 摇蚊幼虫 Chironomidae | |||
| 中国长足摇蚊 Tanypus chinensis | 302(5.31%) | ||
| 半折摇蚊 Chironomus semireductus | 46(3.04%) | ||
| 其他 Others | |||
| 钩虾属一种 Gammarus sp. | 357(7.22%) | 195(16.39%) | |
| 多毛类一种 Polychaeta sp.1 | 200(17.80%) | ||
| 合计 Total | 9,471(91.20%) | 1,046(80.30%) | 408(84.80%) |
比较各聚类组底栖动物功能摄食类群发现, 密度方面, 直接收集者最重要, 其在各聚类组所占比重分别达到94.73%, 57.91%和51.09%, 此外过滤收集者和刮食者分别是组B(37.58%)和组C(38.73%)重要的摄食类群(表3)。生物量方面, 由于直接收集者个体较小, 仅在组A占据较大比重(24.35%), 而在组B和组C所占比例较低, 这进一步说明了直接收集者(主要是颤蚓类)在组A的绝对优势, 组B在生物量上被过滤收集者主导(92.40%), 组C的摄食类群主要为刮食者(76.97%)和过滤收集者(22.27%) (表3)。One-way ANOSIM分析结果表明3组底栖动物群落之间具有显著性差异, 不相似性百分比分别为DisAB = 73.7%, DisAC = 75.3%, DisBC = 68.5% (P< 0.001)。
表3 各聚类组不同功能摄食类群大型底栖动物平均密度(ind./m2)、生物量(g/m2)及其所占比例
Table 3
| 密度 Density | 生物量 Biomass | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group A | Group B | Group C | Group A | Group B | Group C | ||||
| 直接收集者 Gatherers | 8,771(94.73%) | 604(57.91%) | 306(51.09%) | 18.14(24.35%) | 1.64(0.83%) | 1.08(0.69%) | |||
| 过滤收集者 Filterers | 138(1.49%) | 392(37.58%) | 37(6.18%) | 21.55(28.92%) | 182.58(92.40%) | 34.71(22.27%) | |||
| 捕食者 Predators | 294(3.18%) | 27(2.59%) | 20(3.34%) | 0.83(1.11%) | 0.13(0.07%) | 0.09(0.06%) | |||
| 刮食者 Scrapers | 44(0.48%) | 20(1.92%) | 232(38.73%) | 33.99(45.62%) | 13.24(6.70%) | 119.95(76.97%) | |||
| 撕食者 Shredders | 12(0.13%) | 0 | 4(0.67%) | 0 | 0 | 0.01(0.01%) | |||
图5
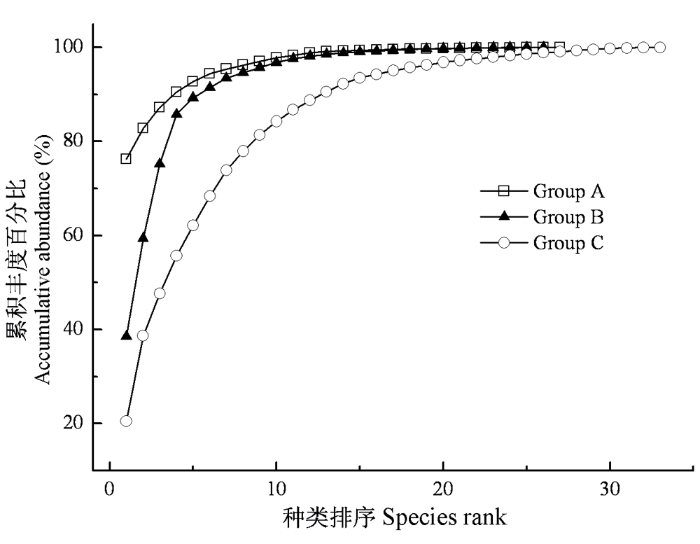
图5
各聚类组大型底栖动物K-优势曲线
Fig. 5
K-dominant curves of macrozoobenthos for each affinity group in Lake Taihu
图6
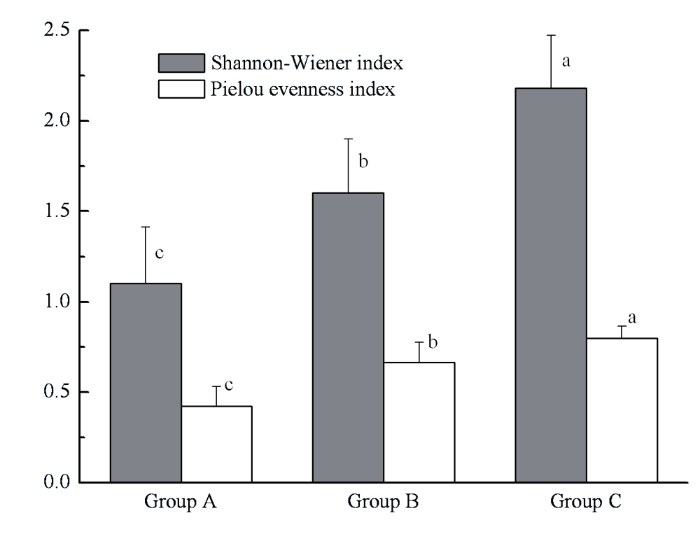
图6
各聚类组大型底栖动物Shannon-Wiener及Pielou多样性指数。各聚类组间不同字母表示具有显著差异(one-way ANOVA, Tukey’s post-hoc test, P < 0.05)
Fig. 6
Shannon-Wiener index and Pielou index of macrozoobenthos for each affinity group in Lake Taihu. Different letters indicate significant differences among the groups (one-way ANOVA, Tukey’s post-hoc test, P < 0.05)
3 讨论
3.1 太湖大型底栖动物空间格局及生境特征分析
本次调查结果表明, 太湖大型底栖动物群落在空间上具有显著差异, 且优势种空间分布差异较大, 聚类分析将30个采样点划分为3组, 结合采样点的生境特征发现, 各聚类组所处区域生境差异显著。组A采样点位于梅梁湾、竺山湾及河口, 根据已有的资料及现场调查结果, 太湖北部是沉积物的主要蓄积区, 属于淤泥黏土底质(黄漪平, 2001; 秦伯强等, 2004), 梅梁湾和竺山湾有泥区平均泥深分别可达1.51 m、1.36 m (秦伯强等, 2004), 显著大于其他湖区。同时该区域也是污染最严重、水华暴发最频繁的区域。其沉积物中的有机质、总氮、总磷都显著高于其他湖区(除东太湖) (秦伯强等, 2004; 李江等, 2007)。野外现场测定发现该区域沉积物氧化还原电位较低(-310至-150 mV), 呈还原状态, 含氧量低。TH0、TH6、TH10三个采样点位于河口, 受入湖污水影响较大, 其污染更加严重。这样的生境状况更有利于耐污能力较强的种类生存, 从而使其在竞争中处于优势地位, 直接表现为寡毛纲颤蚓类在该区域的绝对优势, 同时耐污能力较强的中国长足摇蚊和摇蚊属幼虫也集中分布在该区域。
组B的代表优势种为河蚬, 按摄食功能类群划分河蚬为过滤收集者, 主要滤食悬浮碎屑、细菌和浮游植物。组B采样点位于贡湖湾、湖心及西湖区, 该区沉积物属于粉砂、黏土质粉砂类型, 粒径范围较广(黄漪平, 2001; 秦伯强等, 2004), 其受风浪影响较大, 底泥扰动强烈, 含氧量较高, 这些特征更有利于河蚬的滤食, 其密度高值也出现在该区(图3C)。北部梅梁湾、竺山湾内细粒径的淤泥可能阻碍了河蚬的滤食活动, 较低的氧浓度也降低了其存活率, 其密度较低(< 85 ind./m2)。东太湖和东部湖湾河蚬密度也较低, 可能是因为该区域水生植物丰富, 水体透明度高, 浮游植物的密度相对较低, 可供滤食的有机质较少。
组C的采样点主要位于东太湖和东部湖湾(TH14除外), 其优势种为腹足纲螺类。根据采样现场记录以及谷孝鸿等(2005)、刘伟龙等(2007)的研究, 该区域是太湖水生植物的主要分布区, 而其他湖区水生植物分布很少。关于螺类与水生植物之间关系的研究较多, 传统观点认为螺类主要以有机碎屑及附着藻类等为食, 且几乎不牧食水生植物本身(Reavell, 1980), Thomas (1990)基于自己及前人的研究工作提出了螺—草互利理论(mutualistic theory), 该理论认为螺类、附生生物与水生植物之间形成了一个互利的子系统, 这一理论一直被许多实验结果所支持(BrÖnmark, 1985, 1990; Jones & Sayer, 2003), 比较太湖螺类与水生植物分布发现, 其高值出现在水生植物最为丰富的东太湖和东部湖湾, 这也进一步印证了螺—草的互惠关系。
3.2 太湖大型底栖动物多样性及影响因子
传统生态学观点认为, 空间异质性程度越高, 意味着有更加多样的小生境, 能允许更多的物种共存(Tews et al., 2004; Shostell & Williams, 2007), 从而具有更高的生物多样性。国内外关于生境与底栖动物多样性关系的研究较多(Tolonen et al., 2001; Galuppo et al., 2007; Shostell & Williams, 2007), 认为生境的复杂性是决定底栖动物多样性的关键因子。太湖大型底栖动物多样性大小可依次排列为组C > 组B > 组A。组C是水生植物的主要分布区域, 其为底栖动物提供了更加多样的栖息、繁殖场所, 也有利于底栖动物逃避其他动物的捕食, 从而增加了可供更多底栖动物生存的生境空间, 底栖动物多样性较高。闫云君等(2005)在研究草型和藻型湖泊底栖动物群落结构时也发现草型湖泊底栖动物多样性显著高于藻型湖泊。组A和组B基本无水草分布, 因此很多依靠水草生存及庇护的种类在该区域的密度和生物量很低甚至未出现。多样性分析结果表明组B均匀度显著高于组A, 组B为砂质淤泥, 其粒径范围较广的底质不仅为附生生物提供了很大的附着面积, 其形成的众多小生境也为底栖动物提供了生存空间和庇护场所, 从而增加了某些种类的生存机会。相比之下, 组A沉积物粒径偏细, 黏土(<0.01 mm)比重达到40%以上(黄漪平, 2001; 秦伯强等, 2004), 粒径范围较窄使得底质生境的异质性较低, 难以支持更多种类的生存, 而且有机质丰富, 分解作用强烈导致该区域溶氧较低, 限制了敏感种类的生存, 而耐污能力较强的颤蚓类则大量繁殖, 成为绝对优势种, 从而物种均匀度相对较低。段学花等(2007)研究底质对底栖动物多样性的影响时也发现, 颗粒粒径大、孔隙率大的底质具有更高的生物多样性。
3.3 富营养化对太湖大型底栖动物群落的影响
太湖是典型的富营养化湖泊, 且各湖区营养水平差异较大(朱广伟, 2008), 各湖区大型底栖动物群落结构的差异在一定程度上反映了富营养化对底栖动物群落结构的影响。一般来说, 中等营养水平下生物群落结构最复杂, 物种多样性最高, 而极端贫营养和超富营养的条件下群落结构简单, 多样性低(Frouin, 2000)。Gong & Xie (2001)研究水体富营养化对武汉东湖大型底栖动物多样性影响时发现多样性与水体营养水平呈相反趋势, 富营养化导致多样性明显降低。熊金林等(2003)在研究湖北4个不同营养水平湖泊大型底栖动物群落结构时也发现, 大型底栖动物种类数、物种多样性与湖泊富营养程度呈负相关。
梅梁湾是太湖水体富营养化过程的典型湖湾, 几十年来不断受生活工业污水影响, 从20世纪60至90年代, 伴随着水质由II-III类不断恶化至劣V类, 梅梁湾水生植被分布面积不断缩减直至消亡。同时大型底栖动物群落也发生了较大的变化, 20世纪60年代以河蚬、环棱螺为主要种类(88 ind./m2、26 ind./ m2), 环节动物和水生昆虫密度仅为37 ind./m2(中国科学院南京地理研究所, 1965); 80年代除了河蚬和环棱螺外, 出现了较多的颤蚓科种类, 特别是在0号点附近水丝蚓密度达到2,148 ind./m2(黄漪平, 2001); 90年代梅梁湾底栖动物优势种已变成了摇蚊属幼虫和水丝蚓(秦伯强等, 2004), 而本次调查结果显示寡毛类水丝蚓已成为绝对优势种(占总密度的94.6%)。
本研究显示, 现阶段太湖北部梅梁湾、竺山湾及河口以寡毛纲颤蚓类为绝对优势种, 底栖动物多样性最低, 到了贡湖湾、湖心及西湖区优势种变为瓣鳃纲河蚬, 在水生植物丰富的东部湖湾和东太湖多样性最高, 优势种为腹足纲螺类。分析结果表明营养水平、底质类型以及水生植被的分布是决定太湖大型底栖动物群落结构及多样性的关键因子。
参考文献
Interactions between macrophytes, epiphytes and herbivores: an experimental approach
How do herbivorous freshwater snails affect macrophyte? A comment
Non-parametric multivariate analyses of changes in community structure
The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling
Two-decade reconstruction of algal blooms in China’s Lake Taihu
Field experiment on the effect of streambed substrate on macroinvertebrate diversity
Effects of anthropogenic disturbances of tropical soft-bottom benthic communities
Habitat types and distribution of benthic macroinvertebrates in a transitional water ecosystem: Alimini Grande (Puglia, Italy)
Impact of eutrophication on biodiversity of the macrozoobenthos community in a Chinese shallow lake
Evolution of community structure of aquatic macrophytes in East Taihu Lake and its wetlands
Macrobenthic community structure in the southern and central Bohai Sea, China
Does the fish-invertebrate-periphy- ton cascade precipitate plant loss in shallow lakes?
Vertical changes of physiochemical characteristics and phosphorus concentrations of sediments from different trophic regions of Taihu Lake
The role of zoobenthos in energy flow in two shallow lakes
Temporal and spatial variation of aquatic macrophytes in West Taihu Lake
Approaches to mechanisms and control of eutrophication of shallow lakes in the middle and lower reaches of the Yangtze River
Environmental issues of Lake Taihu, China
A study on the diets of some British freshwater gastropods
Habitat complexity as a determinate of benthic macroinvertebrate community structure in cypress tree reservoirs
Benthic assessment
Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures
Mutualistic interactions in freshwater modular systems with molluscan components
Influences of habitat type and environmental variables on littoral macroinvertebrate communities in a large lake system
Nutrient cycling by animals in freshwater ecosystems
A new method for detecting pollution effects on marine macrobenthic communities
Comparative study on the community structure and biodiversity of zoobenthos in lakes of different pollution states
A comparative study on community structure of macrozoobenthos between macrophtic and algal lakes
Rationale of the multivariate statistical software primer and its application in benthic community ecology
Eutrophic status and causing factors for a large, shallow and subtropical Lake Taihu, China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}