


昆虫是世界上种类和数量最多的生物类群, 在生物多样性保护中具重要地位(Sanways, 1993)。鞘翅目作为昆虫纲最大的类群, 栖息地多样, 其群落结构与环境存在密切关系。许多研究表明, 栖息地理化特征、植被组成和结构、地表枯落物、演替阶段、栖息地破碎化、耕作及其他人为干扰等因素均可能影响甲虫的组成、数量和分布(Evans, 1983; Jennings et al., 1986; Epstein & Kulman, 1990; Lövei & Sunderland, 1996; Rieske & Buss, 2001; Rainio & Niemelä, 2003; Magagula, 2003; 刘云慧等, 2004; Magura et al., 2004; Negro et al., 2007; Gormley et al., 2007)。其中, 植被类型的变化常常对甲虫群落结构具有比较重要的影响(Butterfield, 1997; 于晓东等, 2001, 2002; Magagula, 2003; Yaacobi et al., 2007), 而植被类型本身又涉及植物种类、多样性、结构等多方面因素。
甲虫群落与环境的关系随研究地区和生态系统类型的不同可能呈现较大差异。目前国外研究工作涉及森林(Koivula et al., 1999; Topp et al., 2006)、草原(Irmler & Hoernes, 2003; Larsen & Work, 2003)、农田(Magagula, 2003; Yaacobi et al., 2007)和湿地(Brose, 2003; Bates et al., 2006)等多种生态系统, 而国内研究主要集中于非湿地生态系统(周红章等, 2000; 于晓东等, 2001, 2002; 刘云慧等, 2004; 张倩等, 2008), 有关湿地的研究很少。近年来, 我国湿地、尤其是广大北方湿地因受水资源匮乏的影响而呈现不同程度的退化, 对湿地生物多样性构成严重威胁, 并明显表现在湿地植被类型的变化上。湿地退化及湿地植被类型的变化对甲虫群落结构有什么影响, 主要是哪些环境因子的作用, 亟需进一步探讨。
野鸭湖湿地自然保护区是北京官厅水库延庆辖区及海拔479 m以下环湖淹没区滩涂组成的次生湿地, 孕育着丰富的生物多样性, 于2000年建立市级自然保护区。然而近年来, 由于降水量及上游来水量明显减少, 官厅水库水面缩小, 致使湿地地下水位下降, 一些区域呈现逐渐退化的趋势, 原有的芦苇(Phragmites communis)沼泽逐渐被一些中生甚至旱生植被所替代。目前除在野鸭湖围堰内部和围堰附近以及沿河等处保存着一些生长良好的芦苇群落外, 原有苇地形成大面积主要以牛鞭草(Hemarthria altissima)或猪毛菜(Salsola collina)占优势的植被类型, 存留的芦苇群落也明显退化。本文于2007年4-10月利用陷阱法对野鸭湖湿地两种主要的保存较好的湿地植被类型和3种退化植被类型中的甲虫群落开展了比较系统的调查, 对地表甲虫群落特征与植被类型的关系进行了初步研究, 以期为湿地甲虫多样性保护、湿地环境监测和评价提供科学依据。
1 研究地概况
野鸭湖湿地自然保护区位于北京延庆县西北部, 115°46′-116°00′E, 40°22′-40°31′N, 北面为燕山山脉, 西部和南部与河北怀来县接壤, 保护区总面积6,873 hm2, 拥有常年稳定水面2,000 hm2, 四周淹没区及草地约3,330 hm2 (刘玉金, 2007)。研究地点位于官厅水库东岸向东到野鸭湖度假村围堰的广阔区域, 植被自西向东呈现较明显的带状分布特征, 包括野鸭湖湿地两种主要的湿地景观保存较好的植被类型和3种主要的退化的植被类型。各植被类型的主要特征如下:
球穗莎草(Cyperus glomeratus)带: 官厅水库新近退水后形成的湿生草本植被带, 受季节性淹没和退水的影响。主要植物有球穗莎草、扁秆藨草(Scirpus planiculmis)、绵毛酸模叶蓼(Polygonum lapathifolium var. salicifolium)、稗(Echinochloa crusgalli)等, 夏季群落高度约80 cm, 盖度约80%。
猪毛菜带: 猪毛菜、软毛虫实(Corispermum puberulum)、茵陈蒿(Artemisia capillaris)和鹅绒藤(Cynanchum chinensis)占优势, 为退化的湿地植被。群落高度约25 cm, 盖度约50%。
退化芦苇带: 芦苇占优势但退化较严重。牛鞭草侵入明显。其他植物有直立黄芪(Astragalus adsurgens)、狗尾草(Setaria viridis)、鹅绒委陵菜(Potentilla anserina)等, 并有稀疏小叶杨(Populus simonii)和棉花柳(Salix linearistipularis)幼苗生长。夏季草本群落高度约75 cm, 盖度约60%。
牛鞭草带: 牛鞭草占绝对优势, 夏季蒿类植物长势良好, 其他植物主要有鹅绒委陵菜、旋复花(Inula britanica)等, 为退化的湿地植被。夏季群落高度约50 cm, 盖度约70%。
保存较好的芦苇带: 芦苇占优势且生长良好, 其他植物主要有牛鞭草、鹅绒委陵菜、扁秆藨草、光稃香茅(Hierochloe glabra)等。夏季群落高度150 cm, 盖度90-100%。
2 研究方法
2.1 地表甲虫的采集
根据上述各植被类型带状分布宽度的不同, 利用GPS定位, 在各植被类型中分别设立1-2条样带, 共8条样带(图1)。
图1
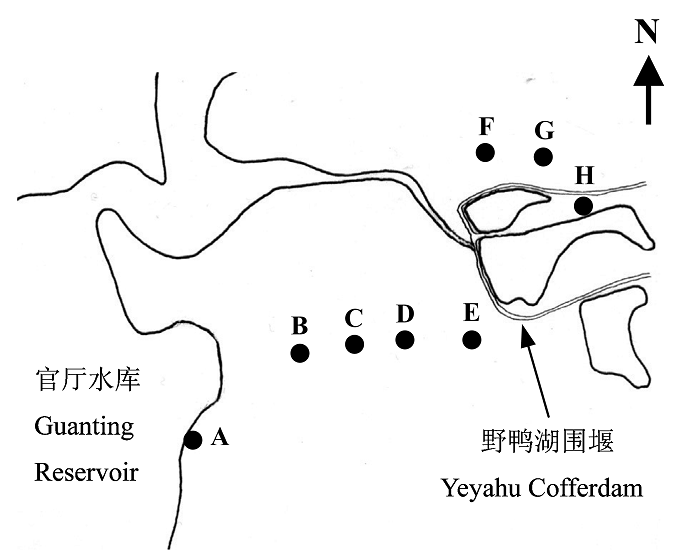
图1
野鸭湖湿地地表甲虫采集样带分布示意图。A: 球穗莎草带; B、C: 猪毛菜带; D: 退化的芦苇带; E、F: 牛鞭草带; G、H: 保存较好的芦苇带。
Fig. 1
The distribution of sampling transects for ground- dwelling beetles investigation in Yeyahu Wetland. A, Cyperus glomeratus transect; B, C, Salsola collina transects; D, Degradated Phragmites communis transect; E, F, Hemarthria altissima transects; G, H, Well-conserved Phragmites communis transects
其中样带A位于球穗莎草带, 样带B、C位于猪毛菜带, 样带D位于退化芦苇带, 样带E、F位于牛鞭草带, 样带G、H位于保存较好的芦苇带。样带间距一般300-400 m, 但样带A、B之间因存在大片耕地而相距较远。
2007年4-10月, 每月中旬以陷阱法采样一次。分别在各样带中自南向北设4个样地, 样地间距50 m。每块样地内设4个正方形排列的采样点, 采样点内设4个正方形排列的陷阱。采样点间距4 m, 采样点内陷阱间距2 m (图2)。利用一次性塑料杯作为诱杯, 上口径7.5 cm, 下口径5.3 cm, 高10 cm。按照周红章等(2000)和于晓东等(2001)的方法配制诱捕剂, 适量增加酒精比例(醋、糖、酒精、水的质量比为2:1:10:12, 并加入少量表面活性剂)以防标本腐烂。诱杯内加约150 mL诱捕剂, 置于样地3 d后收回。将样地内4个采样点共16个诱杯的捕获结果合并作为1个收集单位, 用75%酒精保存。
图2
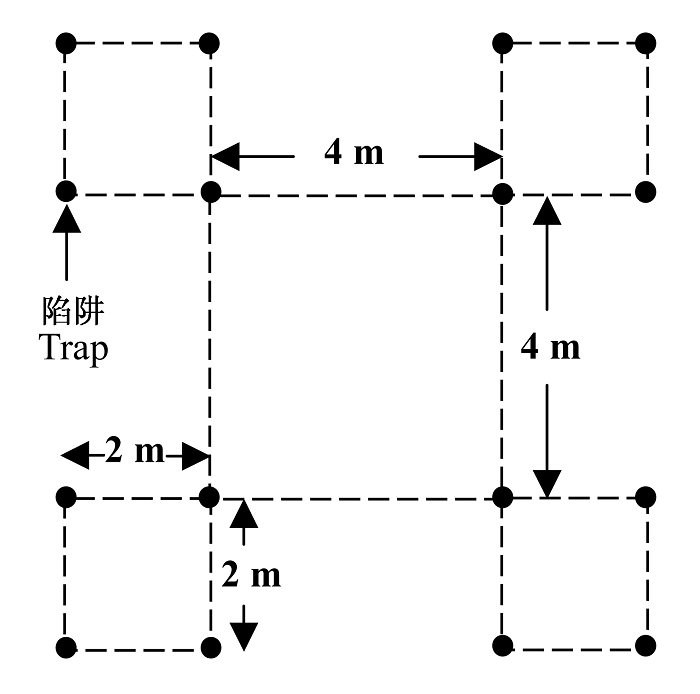
所有标本均鉴定到科水平(李鸿兴等, 1987; 郑乐怡和归鸿, 1999), 部分标本请中国科学院动物研究所相关专家鉴定。
2.2 环境变量的测定
每月采样时对各样地植物组成、盖度、高度、枯落物盖度等进行调查, 并测定各样地土壤含水量。统一于实验中期(2007年8月)在各样地设1个1 m×1 m样方, 测定样方内植物物种数、密度、盖度、生物量(干重)等数量特征, 以及地表枯落物的盖度、厚度和干重。
2.3 数据处理
甲虫数量测定: 以“活动密度”反映不同样地或不同类群地表甲虫的数量, 其含义为每个样地中16个陷阱3 d所捕获的甲虫个体数。
甲虫数量等级划分: 以特定类群个体数占所有类群总个体数的百分比(D)作为划分标准: D≥20%为优势类群, 5%≤D<20%为亚优势类群, 1%≤D<5%为常见类群, D<1%为稀有类群。
依据如下方法进行群落分析(赵志模和郭依泉, 1990; 杜荣骞, 1999; 张金屯, 2004):
群落多样性测度:
Margalef丰富度指数
Simpson优势度指数
Shannon-Wiener信息多样性指数
Pielou均匀度指数
其中S为类群数(甲虫科数或植物物种数), N为群落总个体数, pi为类群i个体数占群落总个体数的比率, H'max为最大多样性。
甲虫和环境数据的显著性检验: 对服从正态分布或转换后服从正态分布的数据, 采用单因素方差分析方法; 对转换后仍不能服从正态分布的数据, 采用非参数检验方法。
采用典范对应分析(canonical correspondence analysis, CCA)、Spearman秩相关系数和非线性回归方法分析不同植被类型地表甲虫群落组成与环境因子的关系。
采用主成分分析(principal component analysis, PCA)对环境数据进行综合分析, 获得新的综合环境变量, 进而分析主要环境因子的综合作用对不同植被类型地表甲虫群落组成特征的影响。
利用Excel、SPSS13.0统计软件、Canoco4.5分析软件和Canodraw4.0作图软件进行以上统计分析。
3 结果
本次调查共采集地表甲虫11,820号, 隶属于42科(表1)。步甲科和隐翅虫科为优势类群, 蚁形甲科、肖叶甲科和薪甲科为亚优势类群, 5个科合计占甲虫总个体数的70.82%。象甲科、蜉金龟科、叩甲科、拟步甲科、金龟科等11个科为常见类群, 合计占总个体数的24.66%。粪金龟科、小蕈甲科、鳃金龟科、瓢甲科、苔甲科等26个科为稀有类群, 合计不到总个体数的5%。
表1 野鸭湖湿地地表甲虫群落的科组成和分布(2007年4-10月)
Table 1
| 科 Family | 甲虫个体数 Individual number | 总个体数 Total | 百分比 (%) | ||||
|---|---|---|---|---|---|---|---|
| 球穗莎草带 (CGT) | 猪毛菜带 (SCT) | 退化芦苇带 (DPCT) | 牛鞭草带 (HAT) | 保存较好的芦苇带 (WPCT) | |||
| 步甲科 Carabidae | 943 | 282 | 50 | 134 | 1,132 | 2,541 | 21.50 |
| 隐翅虫科 Staphylinidae | 554 | 533 | 183 | 546 | 585 | 2,401 | 20.31 |
| 蚁形甲科 Anthicidae | 71 | 680 | 17 | 530 | 66 | 1,364 | 11.54 |
| 肖叶甲科 Eumolpidae | 61 | 46 | 1 | 15 | 977 | 1,100 | 9.31 |
| 薪甲科 Lathridiidae | 191 | 40 | 49 | 86 | 595 | 961 | 8.13 |
| 象甲科 Curculionidae | 240 | 119 | 18 | 27 | 73 | 477 | 4.04 |
| 蜉金龟科 Aphodiidae | 50 | 48 | 6 | 82 | 197 | 383 | 3.24 |
| 叩甲科 Elateridae | 50 | 44 | 2 | 0 | 280 | 376 | 3.18 |
| 拟步甲科 Tenebrionidae | 5 | 247 | 15 | 2 | 48 | 317 | 2.68 |
| 金龟科 Scarabaeidae | 8 | 201 | 20 | 53 | 10 | 292 | 2.47 |
| 叶甲科 Chrysomelidae | 62 | 37 | 2 | 4 | 159 | 264 | 2.23 |
| 埋葬甲科 Silphidae | 67 | 64 | 20 | 31 | 18 | 200 | 1.69 |
| 隐食甲科 Cryptophagidae | 28 | 108 | 7 | 19 | 28 | 190 | 1.61 |
| 丽金龟科 Rutelidae | 2 | 100 | 49 | 3 | 2 | 156 | 1.32 |
| 虎甲科 Cicindelidae | 14 | 88 | 0 | 0 | 33 | 135 | 1.14 |
| 阎甲科 Histeridae | 4 | 77 | 12 | 26 | 5 | 124 | 1.05 |
| 粪金龟科 Geotrupidae | 1 | 25 | 14 | 57 | 9 | 106 | 0.90 |
| 小蕈甲科 Mycetophagidae | 46 | 11 | 1 | 7 | 14 | 79 | 0.67 |
| 鳃金龟科 Melolonthidae | 12 | 22 | 1 | 12 | 8 | 55 | 0.47 |
| 瓢甲科 Coccinellidae | 6 | 14 | 9 | 16 | 4 | 49 | 0.41 |
| 苔甲科 Scydmaenidae | 0 | 0 | 13 | 0 | 25 | 38 | 0.32 |
| 犀金龟科 Dynastidae | 11 | 2 | 3 | 11 | 11 | 38 | 0.32 |
| 蕈坚甲科 Merycidae | 1 | 2 | 6 | 21 | 0 | 30 | 0.25 |
| 花萤科 Cantharidae | 2 | 0 | 0 | 0 | 23 | 25 | 0.21 |
| 露尾甲科 Nitidulidae | 4 | 2 | 2 | 4 | 10 | 22 | 0.19 |
| 缨甲科 Ptiliidae | 18 | 0 | 0 | 0 | 3 | 21 | 0.18 |
| 平唇水龟科 Hydraenidae | 12 | 0 | 0 | 0 | 0 | 12 | 0.10 |
| 蚁甲科 Pselaphidae | 2 | 2 | 0 | 1 | 6 | 11 | 0.09 |
| 圆泥甲科 Georyssidae | 1 | 0 | 0 | 0 | 7 | 8 | 0.07 |
| 锹甲科 Lucanidae | 4 | 0 | 0 | 0 | 4 | 8 | 0.07 |
| 花蚤科 Mordellidae | 0 | 3 | 0 | 3 | 1 | 7 | 0.06 |
| 长蠹科 Bostrychidae | 0 | 3 | 1 | 1 | 1 | 6 | 0.05 |
| 球蕈甲科 Leiodidae | 0 | 1 | 1 | 0 | 3 | 5 | 0.04 |
| 水龟甲科 Hydrophilidae | 3 | 0 | 0 | 0 | 1 | 4 | 0.03 |
| 小丸甲科 Nosodendridae | 0 | 0 | 1 | 0 | 3 | 4 | 0.03 |
| 铁甲科 Hispidae | 2 | 1 | 0 | 0 | 0 | 3 | 0.03 |
| 水缨甲科 Hydroscaphidae | 0 | 0 | 0 | 0 | 2 | 2 | 0.02 |
| 花金龟科 Cetoniidae | 0 | 2 | 0 | 0 | 0 | 2 | 0.02 |
| 天牛科 Cerambycidae | 0 | 1 | 0 | 0 | 0 | 1 | 0.01 |
| 皮蠹科 Dermestidae | 1 | 0 | 0 | 0 | 0 | 1 | 0.01 |
| 拟花萤科 Melyridae | 0 | 0 | 0 | 0 | 1 | 1 | 0.01 |
| 窃蠹科 Anobiidae | 0 | 0 | 1 | 0 | 0 | 1 | 0.01 |
| 总计 Total | 2,476 | 2,805 | 504 | 1,691 | 4,344 | 11,820 | |
CGT, Cyperus glomeratus transect; SCT, Salsola collina transect; DPCT, Degradated Phragmites communis transect; HAT, Hemarthria altissima transect; WPCT, Well-conserved Phragmites communis transect
3.1 不同植被类型地表甲虫群落的组成特征
3.1.1 群落组成的多样性
从表2可以看出, 地表甲虫群落的活动密度以球穗莎草带最高, 其次是保存较好的芦苇带, 3种较退化的植被类型明显较低。5种植被类型之间, 除保存较好的芦苇带与球穗莎草带无显著差异(P>0.05)、与猪毛菜带差异显著(P<0.05)外, 其余植被类型彼此差异均极显著(P<0.01)。
表2 野鸭湖湿地不同植被类型地表甲虫群落的活动密度、科丰富度和多样性指数(平均值±标准误)
Table 2
| 球穗莎草带 (CGT) | 猪毛菜带 (SCT) | 退化芦苇带 (DPCT) | 牛鞭草带 (HAT) | 保存较好的 芦苇带 (WPCT) | |
|---|---|---|---|---|---|
| 活动密度 Activity density (AD) | 619.0 ± 50.2d | 350.6 ± 26.4c | 126.0 ± 8.7a | 211.4 ± 20.1b | 543.0 ± 67.1d |
| 科丰富度 Family richness (R) | 23.5 ± 1.0c | 20.8 ± 0.9b | 19.0 ± 0.4b | 15.8 ± 0.7a | 21.1 ± 0.7bc |
| 丰富度指数 Margalef richness index (D) | 3.51 ± 0.19bc | 3.38 ± 0.14bc | 3.73 ± 0.13c | 2.77 ± 0.11a | 3.23 ± 0.10b |
| 多样性指数 Shannon-Wiener diversity index (H') | 2.01 ± 0.11ab | 2.35 ± 0.08c | 2.22 ± 0.02bc | 1.82 ± 0.09a | 2.02 ± 0.06ab |
| 均匀度指数 Pielou evenness index (E) | 0.64 ± 0.03a | 0.78 ± 0.02b | 0.75 ± 0.01b | 0.66 ± 0.03a | 0.66 ± 0.02a |
| 优势度指数 Simpson dominance index (C) | 0.22 ± 0.03ab | 0.15 ± 0.02a | 0.18 ± 0.01ab | 0.26 ± 0.03b | 0.20 ± 0.02ab |
同行数据间不同上标字母表示差异显著(P<0.05)
The meanings of CGT, SCT, DPCT, HAT and WPCT are the same as those in
地表甲虫群落科丰富度仍以球穗莎草带最高, 保存较好的芦苇带次之, 3种较退化的植被类型相对贫乏。两种保存较好的植被类型彼此无显著差异(P>0.05); 而保存较好的芦苇带与牛鞭草带差异极显著(P<0.01), 球穗莎草带与3种退化植被类型差异均极显著(P<0.01)或显著(P<0.05)。3种退化植被类型中, 牛鞭草带的甲虫科丰富度明显低于退化芦苇带和猪毛菜带 (P<0.01)。其余植被类型之间无明显差异(P>0.05)。
两种保存较好的湿地植被类型虽然地表甲虫群落组成丰富, 但一些类群的优势现象明显, Simpson优势度指数(C)较高而Pielou均匀度指数(E)较低, Shannon-Wiener多样性指数(H')仅高于牛鞭草带而低于猪毛菜带和较退化的芦苇带, 其中与猪毛菜带差异极显著(P<0.01), 与其他植被类型无显著差异(P>0.05)。3种退化的植被类型中, 牛鞭草带不仅组成简单且优势现象突出, 多样性指数(H')明显低于其他两种植被类型(P<0.01)。
Margalef丰富度指数(D)以退化芦苇带最高, 牛鞭草带最低。退化芦苇带与保存较好的芦苇带差异显著(P<0.05); 牛鞭草带与其他植被类型差异显著(P<0.05)或极显著(P<0.01); 其余植被类型之间无明显差异(P>0.05)。
3.1.2 优势科和亚优势科的活动密度
不同植被类型地表甲虫群落的优势科和亚优势科的活动密度见表3。步甲科表现出明显的生境选择性, 不仅在两个保存较好的湿地植被类型为优势科, 且活动密度显著高于其他植被类型(P<0.05)。隐翅虫科分布广泛, 在各个植被类型中的活动密度均较高, 为球穗莎草带、退化芦苇带和牛鞭草带的优势科, 以球穗莎草带活动密度最高, 与其他植被类型差异显著(P<0.05)或极显著(P<0.01)。蚁形甲科为猪毛菜带和牛鞭草带的优势科, 且活动密度与其他植被类型差异显著(P<0.05)或极显著(P<0.01)。肖叶甲科仅在保存较好的芦苇带活动密度较高并为其优势科, 与其他植被类型差异极显著(P<0.01)。
表3 野鸭湖湿地不同植被类型地表甲虫群落优势及亚优势科的活动密度(平均值±标准误)
Table 3
| 类群 Family | 球穗莎草带 (CGT) | 猪毛菜带 (SCT) | 退化芦苇带 (DPCT) | 牛鞭草带 (HAT) | 保存较好的芦苇带 (WPCT) |
|---|---|---|---|---|---|
| 步甲科 Carabidae | 235.8 ± 53.1**, b | 35.3 ± 3.4*, a | 12.5 ± 4.2*, a | 16.8 ± 5.8*, a | 141.5 ± 25.6**, b |
| 隐翅虫科 Staphylinidae | 138.5 ± 9.9**, b | 66.6 ± 2.6*, a | 45.8 ± 3.8**, a | 68.3 ± 11.8**, a | 73.1 ± 13.8*, a |
| 蚁形甲科 Anthicidae | 17.8 ± 4.5b | 85.0 ± 24.5**, c | 4.3 ± 0.8a | 66.3 ± 17.1**,c | 8.3 ± 4.0a |
| 肖叶甲科 Eumolpidae | 15.3 ± 7.2c | 5.8 ± 1.1bc | 0.3 ± 0.3a | 1.9 ± 1.1ab | 122.1 ± 41.5**, d |
| 薪甲科 Lathridiidae | 47.8 ± 4.2*, b | 5.0 ± 1.3a | 12.3 ± 4.0*, a | 10.8 ± 2.7*, a | 74.4 ± 16.7*, b |
| 象甲科 Curculionidae | 60.0 ± 8.6*, d | 14.9 ± 1.9c | 4.5 ± 0.9ab | 3.4 ± 1.3a | 9.1 ± 1.9bc |
| 叩甲科 Elateridae | 12.5 ± 1.7c | 5.5 ± 1.9bc | 0.5 ± 0.3b | 0.0 ± 0.0a | 35.0 ± 4.8*, d |
| 拟步甲科 Tenebrionidae | 1.3 ± 0.6ab | 30.9 ± 5.8*, d | 3.8 ± 0.6c | 0.3 ± 0.3a | 6.0 ± 2.1bc |
| 金龟科 Scarabaeidae | 2.0 ± 1.1ab | 25.1 ± 2.3*, c | 5.0 ± 1.2b | 6.6 ± 2.3b | 1.3 ± 0.8a |
| 丽金龟科 Rutelidae | 0.5 ± 0.3a | 12.5 ± 4.3b | 12.3 ± 3.5*b | 0.4 ± 0.3a | 0.3 ± 0.3a |
* 亚优势科 ** 优势科。同行数据间不同上标字母表示差异显著(P<0.05)。
* Subdominant family ** Dominant family; The meanings of CGT, SCT, DPCT, HAT and WPCT are the same as those in
此外, 薪甲科为球穗莎草带、两种芦苇带和牛鞭草带的亚优势科, 其空间分布特征与步甲科类似, 在两种湿地景观保存较好的植被类型中活动密度较高, 与3种较退化的植被类型差异极显著(P<0.01)。象甲科、叩甲科、拟步甲科和金龟科、丽金龟科分别为球穗莎草带、保存较好的芦苇带、猪毛菜带、退化芦苇带的亚优势科, 而且活动密度均显著(P<0.05)或极显著(P<0.01)高于其他植被类型, 仅退化芦苇带与猪毛菜带之间丽金龟科的活动密度无显著差异(P>0.05)。
3.2 不同植被类型地表甲虫群落与环境因子的关系
3.2.1 不同植被类型环境因子特征
由表4可以看出, 在5个植被因子中, 物种丰富度(SP)和Shannon-Wiener多样性指数(HP)仅在退化芦苇带与牛鞭草带之间差异显著(P<0.05), 密度(DP)仅在猪毛菜带与保存较好的芦苇带之间差异显著(P<0.05)。相比之下, 植物盖度(CP)和生物量(BP)变化很大, 并均在两种保存较好的湿地植被类型高而3种退化的植被类型低。两种保存较好的植被类型之间, 彼此盖度差异显著(P<0.05)、生物量差异不显著(P>0.05); 3种退化植被类型之间, 除猪毛菜带的盖度明显低于牛鞭草带(P<0.05)外, 差异均不显著(P>0.05); 而两种保存较好的植被类型与3种退化植被类型之间, 除球穗莎草带的盖度与牛鞭草带差异不显著(P>0.05)外, 其余差异均极显著(P<0.01)。
表4 野鸭湖湿地不同植被类型9个环境因子的观测值(平均值±标准误)
Table 4
| 环境因子 Environmental factors | 球穗莎草带 (CGT) | 猪毛菜带 (SCT) | 退化芦苇带 (DPCT) | 牛鞭草带 (HAT) | 保存较好的 芦苇带 (WPCT) |
|---|---|---|---|---|---|
| 植物物种数 Plant species number (SP) | 14.5 ± 1.9 ab | 13.9 ± 0.7 ab | 15.5 ± 0.9 b | 11.4 ± 0.8 a | 12.0±1.5 ab |
| 植物群落多样性指数 Shannon-Wiener diversity index of plant community (HP) | 1.12 ± 0.29 ab | 1.46 ± 0.11 ab | 1.45 ± 0.05 b | 1.36 ± 0.06 a | 1.52 ± 0.12 ab |
| 植物密度 Plant density (DP, ind. /m2) | 2,668 ± 1,600 ab | 1,268 ± 144 b | 1,088 ± 467 ab | 888 ± 56 ab | 484 ± 113 a |
| 植物盖度 Plant coverage (CP, %) | 80.0 ± 5.8 c | 48.8 ± 4.0 a | 60.0 ± 4.0 ab | 68.8 ± 4.0 bc | 95.6 ± 1.5 d |
| 植物生物量 Plant biomass (BP, g/m2) | 593.2 ± 77.5 b | 206.2 ± 17.9 a | 252.9 ± 34.2 a | 209.1 ± 15.4 a | 539.0 ± 73.6 b |
| 枯落物盖度 Coverage of litter (CL, %) | 13.8 ± 6.9 a | 35.6 ± 2.9 a | 65.0 ± 2.9 b | 76.3 ± 5.6 b | 82.5 ± 7.7 b |
| 枯落物厚度 Thickness of litter (TL, cm) | 0.2 ± 0.1 a | 0.4 ± 0.1 a | 1.1 ± 0.2 b | 1.3 ± 0.3 b | 1.3 ± 0.3 b |
| 枯落物干重 Dry weight of litter (WL, g/m2) | 50.8 ± 21.5 a | 90.7 ± 16.5 a | 222.2 ± 35.1 bc | 170.2 ± 19.3 b | 292.3 ± 47.5c |
| 土壤含水量 Soil water content (SW, %) | 25.7 ± 3.3 b | 8.1 ± 1.0 a | 7.1 ± 0.4 a | 8.0 ± 0.7 a | 27.8 ± 1.9 b |
同行数据间不同上标字母表示差异显著(P<0.05)
The meanings of CGT, SCT, DPCT, HAT and WPCT are the same as those in
土壤含水量的变化与植物盖度和生物量的变化趋势相似, 两种保存较好的植被类型明显高于3种退化的植被类型, 差异显著(P<0.05)或极显著(P<0.01); 而两种保存较好的植被类型间、以及3种退化的植被类型间均未见明显差异(P>0.05)。
5种植被类型中, 球穗莎草带的枯落物很少, 其次为猪毛菜带, 3个枯落物因子均显著(P<0.05)或极显著(P<0.01)低于其他3种植被类型。
3.2.2 地表甲虫群落组成与环境因子的典范对应分析
32个样地地表甲虫群落组成与9个环境因子的CCA排序结果见图3。前两个排序轴的特征值分别为0.388和0.106, 达到总特征值(1.106)的44.7%和总典范分析特征值(0.677)的73.1%, 而且环境因子轴与种类排序轴间的相关系数分别为0.9643和0.8519, 两个种类排序轴近似垂直(相关系数0.0257), 两个环境排序轴的相关系数为0, 表明排序结果可靠, 能够较好地反映地表甲虫群落组成变化与9个环境因子的关系。
图3
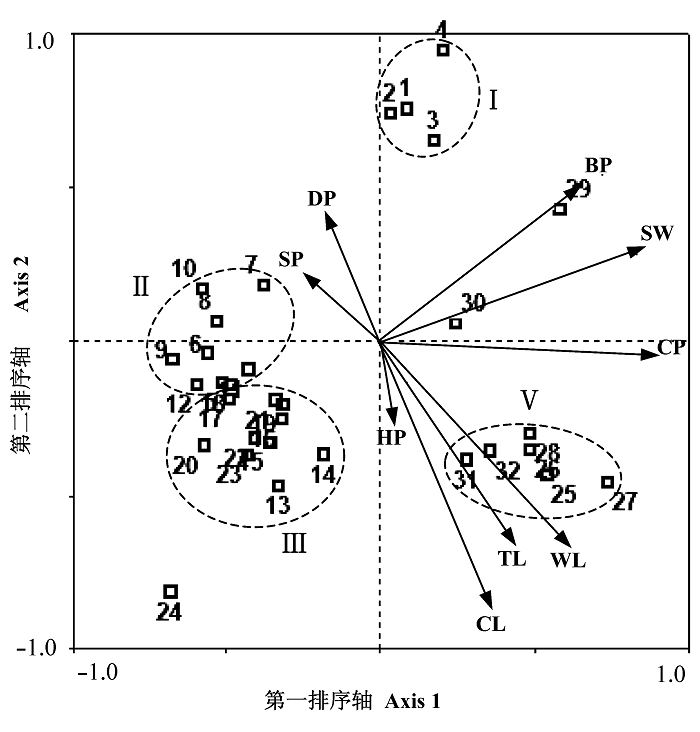
图3
野鸭湖湿地32个样地地表甲虫群落与9个环境因子的CCA排序图。1-4: 球穗莎草样地; 5-12: 猪毛菜样地; 13-16: 退化的芦苇样地; 17-24: 牛鞭草样地; 25-32: 保存较好的芦苇样地。各环境因子名称同
Fig. 3
Canonical correspondence analysis (CCA) ordination performed on ground-dwelling beetles communities and nine environmental factors in 32 sampling sites in Yeyahu Wetland. 1-4, Cyperus glomeratus sites; 5-12, Salsola collina sites; 13-16, Degradated Phragmites communis sites; 17-24, Hemarthria altissima sites; 25-32, Well-conserved Phragmites communis sites. The meanings of environmental factors are the same as those in
CCA第一排序轴与植物盖度(CP)和土壤含水量(SW)呈很强的正相关, 其次为植物生物量(BP)和枯落物干重(WL), 相关系数分别为0.8569、0.8199、0.6356 和0.5931。第二排序轴与枯落物盖度(CL)呈强负相关(相关系数-0.7482), 其次为枯落物干重(WL)和枯落物厚度(TL), 相关系数分别为-0.5585和-0.5571。在CCA排序图中(图3), 植物盖度的变化方向与第一排序轴基本一致, 从左至右增加; 土壤含水量和植物生物量的变化方向自左下向右上增加; 而3个枯落物变量则自左上向右下增加。
依据CCA排序结果, 可将32个样地划分为较明显的4组。依据甲虫群落组成与环境因子关系进行的分组与按植被类型进行的分组之间呈现明显的对应关系, 表明不同植被类型作为地表甲虫的栖息地对于甲虫群落组成特征具综合影响, 而且土壤含水量、植物盖度、生物量及枯落物盖度是影响地表甲虫群落组成的重要环境因子。
组Ⅰ: 包括球穗莎草带的4个样地, 位于两个排序轴正方向, 对应于高植物生物量和高土壤含水量及较高的植物盖度, 但因主要受季节性淹没和退水冲刷的影响, 枯落物覆盖度很低。
组Ⅱ: 包括猪毛菜带的8个样地, 位于第一排序轴负方向。栖息地中大多数环境因子如植物盖度、生物量、土壤含水量及枯落物覆盖度等均处于较低水平。
组Ⅲ: 包括退化芦苇带和牛鞭草带的各个样地, 惟样地24相距较远。本群落组与群落组Ⅱ的位置非常接近, 但在第二排序轴上向负方向偏移更多。土壤含水量和植物生物量仍然较低, 但植物盖度较猪毛菜带有所增加, 且枯落物覆盖度较高。
组Ⅳ: 包括保存较好的芦苇带的大多数样地, 位于第一排序轴正方向和第二排序轴负方向。栖息地环境中很多因子均处于较高水平, 除高植物盖度、生物量和土壤含水量外, 枯落物覆盖度也很高。
总体来看, 湿地景观保存较好的两种植被类型的各个样地均位于代表植物盖度、生物量及土壤含水量3个环境因子的矢量的正方向, 而3种较退化的植被类型的各个样地均位于相反方向, 反映在湿地植被退化的过程中, 植物盖度、生物量及土壤含水量的变化及其综合作用对于地表甲虫群落的组成具有重要影响。
3.2.3 地表甲虫群落活动密度、科丰富度及多样性指数与环境因子的关系
3.2.3.1 相关和回归分析
采用Spearman秩相关系数分析不同植被类型地表甲虫群落组成特征与各环境因子之间变化的关联程度。地表甲虫活动密度与土壤含水量 (r=0.605, P<0.01)、植物生物量 (r=0.487, P<0.01)和植物盖度 (r=0.419, P<0.05)均呈极显著或显著正相关; 科丰富度与植物生物量显著正相关(r=0.403, P<0.05), 与枯落物盖度显著负相关(r=-0.414, P<0.05); 多样性指数(H')与植物盖度极显著负相关(r=-0.449, P<0.01), 与枯落物盖度显著负相关(r=-0.421, P<0.05)。5个植被因子中, 植物种类组成的多样性(SP和HP)和植物密度与地表甲虫群落活动密度、科丰富度及多样性指数(H')之间的相关性均不显著(P>0.05)。
在地表甲虫群落活动密度、科丰富度及多样性指数(H')与9个环境因子的关系中, 土壤含水量与甲虫活动密度(AD)的回归关系最为明显(图4), 能够解释32个样地地表甲虫活动密度总方差的大约57% (AD=-15.3659+40.9446SW-0.7093SW2, R2=0.5715, F =19.3375, P<0.0001)。总体来看, 地表甲虫群落的活动密度有随土壤含水量增加而增加的趋势, 但在较高土壤含水量条件下呈现渐进饱和。
图4
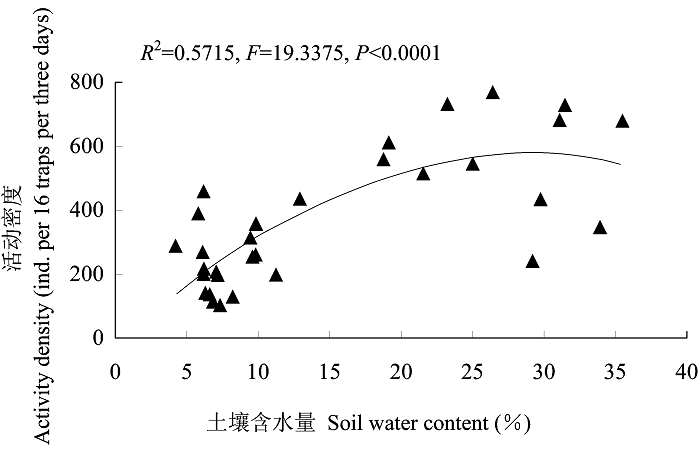
图4
地表甲虫群落活动密度与土壤含水量的非线性回归
Fig. 4
Nonlinear regression of activity density of ground- dwelling beetle communities on soil water content
3.2.3.2 土壤含水量、植物盖度和生物量综合作用分析
对32个样地的9个环境因子进行主成分分析(PCA), 利用PCA的降维功能寻找新的综合环境变量。前3个主成分累积贡献率为79.59%, 进行方差最大正交旋转(Varemax)后的因子载荷矩阵见表5。
表5 9个环境因子主成分分析因子载荷矩阵(旋转后)
Table 5
| 环境因子 Environmental factor | 主成分 Component | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| TL | 0.911 | -0.035 | -0.018 |
| CL | 0.900 | -0.020 | 0.103 |
| WL | 0.859 | 0.215 | 0.195 |
| SP | -0.553 | -0.074 | 0.432 |
| SW | 0.023 | 0.934 | -0.006 |
| BP | -0.153 | 0.917 | -0.118 |
| CP | 0.411 | 0.825 | 0.103 |
| HP | -0.062 | -0.025 | 0.946 |
| DP | -0.473 | -0.013 | -0.674 |
各环境因子名称同
The meanings of environmental factors are the same as those in
由表5可以看出, 第二主成分的生态学含义十分明确, 突出反映了土壤含水量、植物生物量和盖度3个重要环境因子的影响, 因此可将其作为一 个综合环境变量并命名为WBC (Water-Biomass- Coverage), 以各样地PCA第二主成分的因子得分作为变量WBC的值, 分析3个环境因子的综合作用对于不同植被类型地表甲虫群落组成特征的影响。分别以32个样地地表甲虫群落活动密度、科丰富度和多样性指数(H')对综合环境变量WBC作图(图5)。可以看出, 沿着综合环境变量WBC的变化梯度, 32个样地的分布明显间断, 形成相距较远的两组: 两种保存较好的湿地植被类型(组Ⅰ)对应于高WBC值(1.1939 ± 0.1375), 而3种较退化的植被类型对应于低WBC值(-0.7163 ± 0.0547), 两组差异极显著(P<0.01)。而且, 3种比较退化的植被类型的分布格局与CCA排序图非常类似, 猪毛菜带形成一组(组Ⅱ), 退化芦苇带及牛鞭草带形成另一组(组Ⅲ), 两组之间仅个别样地重叠, 表明牛鞭草带与退化芦苇带地表甲虫群落的特征比较相似, 而猪毛菜带甲虫群落则有其自身的特点。
图5
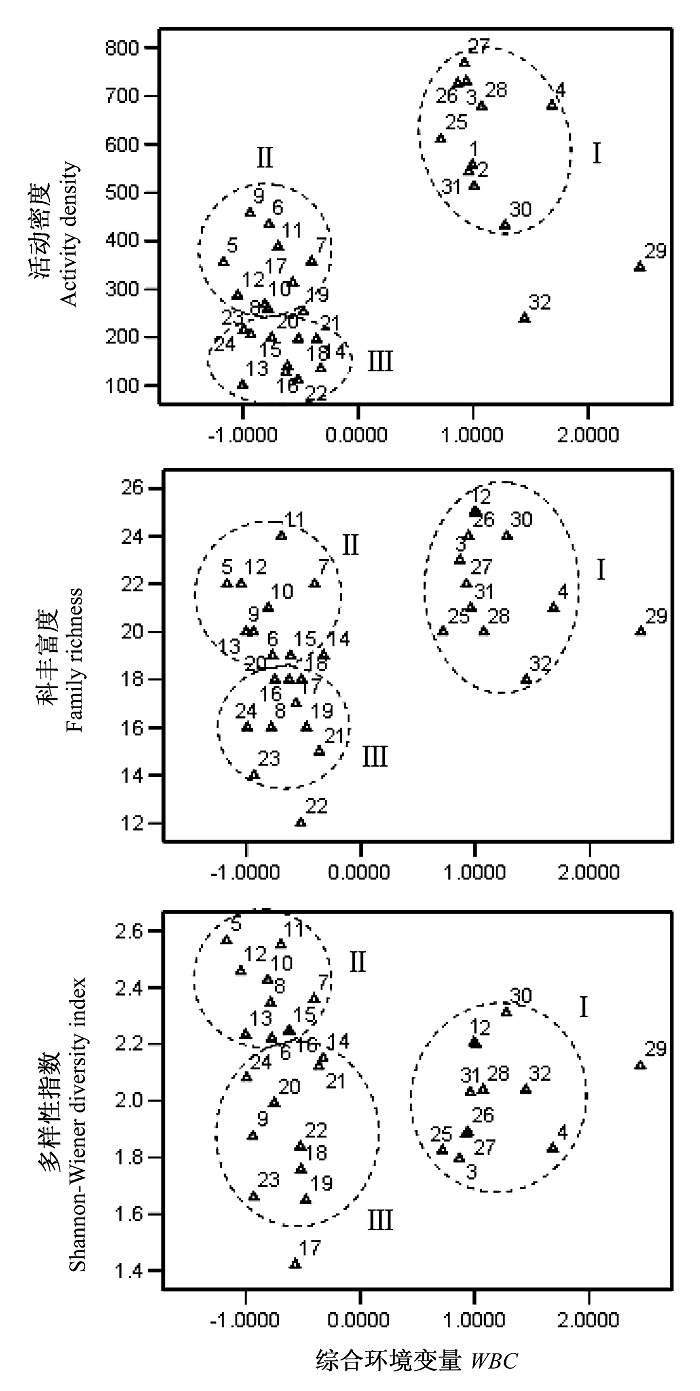
图5
野鸭湖湿地不同植被类型地表甲虫群落活动密度、科丰富度和多样性指数与综合环境变量WBC的关系。其中变量WBC为对9个环境因子进行主成分分析(PCA)得到的第二主成分, 反映了土壤含水量、植物生物量和盖度3个环境因子的综合影响。各样地编号同
Fig. 5
The relationship between activity density, family richness and diversity index of ground-dwelling beetle communities and the synthetic environmental variable (WBC) in Yeyahu Wetland. The variable WBC (Water-Biomass-Coverage), which reflects the situation of soil water content, plant biomass and coverage, is the second component obtained by PCA of nine environmental factors. The labels of sampling sites are the same as those in
在地表甲虫群落活动密度、科丰富度和多样性指数(H')3个特征中, 活动密度与综合环境变量 WBC的关系最为密切, 居于高WBC段的组Ⅰ的甲虫活动密度最高(568.33 ± 47.47), 与组Ⅱ(350.63 ± 26.39)和组Ⅲ(182.92 ± 18.03)差异均极显著(P< 0.01)。
4 讨论
4.1 不同植被类型综合环境特征对地表甲虫群落组成的影响
典范对应分析结果表明, 32个样地大致可分为4组: 球穗莎草样地、保存较好的芦苇样地、猪毛菜样地基本各形成一组, 植被和环境特征比较相似的牛鞭草样地和退化芦苇样地形成另一组, 而且保存较好与较退化的植被类型分别位于第一排序轴的两侧。地表甲虫群落与植被类型的这种对应关系在甲虫群落活动密度、科丰富度和多样性指数(H')与综合环境变量WBC的关系图中也非常明显。不仅保存较好的植被类型与退化的植被类型之间沿着WBC的变化梯度明显分离, 而且3种退化植被类型的分布格局也与CCA排序图非常类似, 牛鞭草样地和退化芦苇样地形成一组, 属于盐沼型植被的猪毛菜样地形成另一组。此外, 两条牛鞭草样带E和F比较, 芦苇残存较多的样带F在CCA排序图中也与退化芦苇带关系更为密切。依据甲虫群落组成与环境因子关系形成的分组与按植被类型进行的分组之间呈现很强的对应关系, 充分反映了不同植被类型作为地表甲虫的栖息地对甲虫群落的重要综合作用以及地表甲虫群落组成特征对于植被类型和环境变化的响应。
而且, 虽然两种保存较好的湿地植被类型在植物物种组成上存在较大差异, 但其植被结构和栖息地环境却具有许多共同特征, 尤其表现为高土壤含水量、高植物盖度和生物量, 并对生活于其内的地表甲虫群落的组成产生重要影响。两种保存较好的湿地植被类型中地表甲虫群落组成丰富, 活动密度远高于退化植被类型, 而且一些典型的湿地甲虫类群也主要栖于这两种生境, 突出反映了湿地景观的保护对于地表甲虫多样性保护的重要意义, 并再次证实了保护生物学的一条基本原理: 栖息地保护是生物多样性保护的前提。
4.2 不同环境因子的相对重要性
总体来看, 关于不同植被特征对甲虫群落的影响, 目前存在两种主要的假说, 即分类学多样性假说(taxonomic diversity hypothesis) (Siemann et al., 1998)和结构异质性假说(structural heterogeneity hypothesis) (Dennis et al., 1998), 分别强调植物种类组成的多样性和群落结构特征对甲虫群落的作用, 并均获得了一些观察结果的支持(Butterfield, 1997; Brose, 2003; Magagula, 2003; 艾尼瓦尔∙吐米尔等, 2005; Yaacobi et al., 2007)。为了进一步比较两类特征的相对重要性, Brose(2003)在东德农业景观区随机选择了30个演替早期的暂时性湿地, 对步甲群落与植被特征的关系进行了定量研究。结果表明植被结构异质性解释了步甲物种丰富度方差的55%, 而植物物种丰富度仅解释了18%, 从而支持“结构异质性假说”。 Brose指出, 植被的影响并不仅仅表现为分类学多样性的影响, 植被结构的作用可能比人们以前所认为的更加重要。
本文采用多种分析方法研究了上述两类植被特征对地表甲虫群落的影响。由CCA图可以看出, 植物盖度和生物量两个植被结构特征与第一排序轴关系非常密切, 而植物物种丰富度和Shannon- Wiener多样性指数两个表征物种组成多样性的变量及植物密度对两个排序轴的影响均较小。而且, 保存较好与较退化的植被类型明显分布于第一排序轴的两侧, 反映在湿地植被的退化过程中, 植物盖度和生物量及土壤含水量的变化及其综合作用对于地表甲虫群落组成具重要影响。甲虫群落活动密度、科丰富度和多样性指数与5个植被因子的相关分析也表明, 植物盖度和生物量与甲虫群落的活动密度相关显著或极显著、植物生物量与甲虫科丰富度相关显著, 植物盖度与甲虫多样性指数相关极显著, 但植物物种丰富度和多样性指数以及植物密度与3个甲虫群落特征的相关性均不明显。可以认为, 本文的研究结果与Brose (2003) 的研究结果类似, 更倾向于支持“结构异质性假说”。但本次研究显示植物密度对于野鸭湖湿地地表甲虫群落影响较小, 与Brose的结果有些不同。
在湿地生态系统中, 水是控制湿地环境和湿地生物多样性的主导因子, 对湿地植被类型的形成及其变化具有关键作用, 进而直接或间接影响着湿地甲虫群落的组成和分布。许多研究均表明土壤含水量对甲虫群落的影响较大。Irmler和Hoernes(2003)采用CCA研究了环境因子对德国北部草原甲虫群落的影响, 其中土壤含水量的作用最大。Boscaini等(2000)对不同环境条件下河岸交错区的结构和功能进行了分析, 发现优势步甲的Shannon-Wiener多样性指数曲线的变化与环境由潮湿到干旱生境的趋势一致。本文CCA结果表明, 土壤含水量与第一排序轴的相关系数仅略低于植物盖度, 对不同植被类型地表甲虫群落在CCA图第一排序轴方向的分布、尤其是保存较好的植被类型与退化植被类型之间的差异具有重要影响。相关和回归分析结果也显示, 土壤含水量与甲虫活动密度的关系十分密切, 二者不仅极显著正相关, 而且土壤含水量能够解释32个样地甲虫活动密度总方差的大约57%, 反映土壤含水量是影响野鸭湖湿地地表甲虫群落活动密度的十分重要的环境因子。
地表枯落物特征反映死亡植物体对以地表为主要栖息地的甲虫的直接或间接影响, 不同学者的研究结果差异较大。Magura等(2004)的研究表明, 枯落物盖度的变化与步甲多度和物种丰富度无明显关系, 但其可能改变地表湿度和温度, 由此产生的对步甲分布的影响可能比枯落物本身重要。Topp(2006)等发现, 在中欧西部特定类型原始森林中, 枯木碎屑对地表甲虫的影响比其他环境因子都大。Koivula等(1999)对北方落叶林的研究显示, 枯落物的添加使大部分步甲的个体数增加, 但对步甲群落的多样性没有显著影响。于晓东等(2006)也发现枯落物厚度对甲虫群落的个体数具重要正影响, 但枯落物盖度对甲虫科丰富度、多样性指数和均匀度有比较重要的负影响。本文CCA结果表明地表枯落物覆盖对野鸭湖湿地不同植被类型地表甲虫群落组成的影响小于土壤含水量和植物盖度及生物量, 但明显大于植物群落多样性; 相关分析显示枯落物盖度与甲虫科丰富度和多样性指数的变化显著负相关, 与于晓东等(2006)研究结果类似。但是, 对于这种负相关关系的解释尚需更多、更深入的比较研究。例如, 在野鸭湖湿地5种植被类型中选取样地数目较多的猪毛菜带、牛鞭草带和保存较好的芦苇带, 进一步分析特定植被类型中不同样地之间地表甲虫群落活动密度、科丰富度和多样性指数与3个枯落物因子的相关性。结果显示, 这些关系在枯落物覆盖程度较高的牛鞭草带和保存较好的芦苇带大多为负相关, 而在枯落物覆盖程度较低的猪毛菜带大多为正相关。上述结果表明, 枯落物覆盖与地表甲虫群落的关系比较复杂, 不仅对不同地区、不同生态系统类型和不同空间尺度的研究可能得出不同结论, 而且在特定区域、相同研究尺度但不同枯落物覆盖条件下所得结果也可能不同, 因此在对研究结果进行解释时需十分谨慎。
4.3 湿地评价指标分析
由本文结果可以看出, 在9个环境因子中, 土壤含水量、植物盖度和生物量不仅与植被类型的变化关系密切, 而且对甲虫群落的组成特征具有重要影响。3个环境因子在保存较好与比较退化的植被类型之间差异很大, 而两种保存较好的植被类型之间、3种比较退化的植被类型之间通常未见明显差异, 反映其对于湿地环境及湿地植被的退化具有比较明显的生境指示作用。而且, 保存较好与退化的植被类型之间沿着综合环境变量WBC的变化梯度明显分离, 在CCA图中也明显分布于第一排序轴的两侧, 进一步表明将土壤含水量、植物盖度和植物生物量3个因子结合成一个综合环境变量, 可能 成为反映湿地环境和湿地植被退化的重要评价指 标。
本文分析结果还显示, 在地表甲虫群落的各特征中, 活动密度对栖息地环境变化更为敏感, 也有可能成为湿地生态环境的综合评价指标。而且, 以甲虫群落总活动密度作为评价指标, 大大减少了分类鉴定的困难, 极大方便了实际应用。
此外, 一些学者指出, 一些甲虫可成为监测环境和生物多样性变化的指示类群(Boscaini et al., 2000; 李巧等, 2006)。本文研究结果也表明, 在野鸭湖湿地生态系统中, 某些地表甲虫的分布表现出比较明显的倾向性。如步甲科在湿地景观较好、土壤水分含量较高的球穗莎草带和保存较好的芦苇带显著较多, 反映出对高湿度生境的选择性, 与其他一些学者的研究结果类似(Luff et al., 1989; Irmler, 2000)。另外, 水龟甲科、平唇水龟科等湿生类群仅分布在湿地景观保存较好的植被类型; 肖叶甲科在保存较好的芦苇带数量高而退化芦苇带极少; 叩甲科在牛鞭草带未采到。但这些类群是否可以筛选出来作为环境指示生物, 对湿地生态环境变化具有怎样的指示作用, 以及如何筛选等, 是下一步研究的重点。
致谢
衷心感谢中科院动物所梁宏斌老师帮助鉴定部分甲虫标本, 首都师范大学生命科学学院胡东老师帮助鉴定植物标本, 北京师范大学张金屯先生和山西大学张峰先生对统计分析进行悉心指导, 北京野鸭湖湿地自然保护区管理处对野外调查工作给予大力支持和热情帮助。同时对协助标本鉴定的仲雨霞师姐深表谢意!
参考文献
Condition-dependent dispersal of a patchily distributed riparian ground beetle in response to disturbance
River ecotones: carabid beetles as a tool for quality assessment
Bottom-up control of carabid beetle communities in early successional wetlands: mediated by vegetation structure or plant diversity?
DOI:10.1007/s00442-003-1222-7 URL [本文引用: 4]
Carabid community succession during the forestry cycle in conifer plantations
Distribution and abundance of small insects and arachnids in relation to structural heterogeneity of grazed, indigenous grasslands
Habitat distribution and seasonal occurrence of carabid beetles in East-central Minnesota
Habitat selection in the Carabidae
Distribution of ground-dwelling beetles in fragmented tropical habitats
Environmental characteristics of ground beetle assemblages in northern German forests as basis for an expert system
Assignment and evaluation of ground beetle (Coleoptera: Carabidae) assemblages to sites on different scales in a grassland landscape
Carabid beetles (Coleoptera: Carabidae) associated with strip clearcut and dense spruce-fir forests of Maine
Leaf litter and the small-scale distribution of carabid beetles (Coleoptera, Carabidae) in the boreal forest
Differences in ground beetles (Coleoptera: Carabidae) of original and reconstructed tallgrass prairies in northeastern Iowa, USA, and impact of 3-year spring burn cycles
Arthropod used as bio-indicator to assess the success of ecological restoration
Temporal and spatial structure of carabid community in agricultural landscape of Dongbeiwang, Beijing
Ecology and behavior of ground beetles (Coleoptera: Carabidae)
Classification and ordination of habitats of ground beetles (Coleoptera, Carabidae) in north-east England
Changes in carabid beetle diversity within a fragmented agricultural landscape
Changes in carabid beetle assemblages along an urbanisation gradient in the city of Debrecen, Hungary
The effect of local anthropogenic habitat heterogeneity on assemblages of carabids (Coleoptera, Caraboidea) endemic to the Alps
Ground beetles (Coleoptera: Carabidae) as bioindicators
Influence of site on diversity and abundance of ground- and litter-dwelling coleoptera in Appalachian oak-hickory forests
Insects in biodiversity conservation: some perspectives and directives
Experimental tests of the dependence of arthropod diversity on plant diversity
DOI:10.1086/286204
URL
PMID:18811348
[本文引用: 1]

Because a diversity of resources should support a diversity of consumers, most models predict that increasing plant diversity increases animal diversity. We report results of a direct experimental test of the dependence of animal diversity on plant diversity. We sampled arthropods in a well-replicated grassland experiment in which plant species richness and plant functional richness were directly manipulated. In simple regressions, both the number of species planted (log2 transformed) and the number of functional groups planted significantly increased arthropod species richness but not arthropod abundance. However, the number of species planted was the only significant predictor of arthropod species richness when both predictor variables were included in ANOVAs or a MANOVA. Although highly significant, arthropod species richness regressions had low R2 values, high intercepts (24 arthropod species in monocultures), and shallow slopes. Analyses of relations among plants and arthropod trophic groups indicated that herbivore diversity was influenced by plant, parasite, and predator diversity. Furthermore, herbivore diversity was more strongly correlated with parasite and predator diversity than with plant diversity. Together with regression results, this suggests that, although increasing plant diversity significantly increased arthropod diversity, local herbivore diversity is also maintained by, and in turn maintains, a diversity of parasites and predators.
Litter-dwelling beetles in primeval forests of Central Europe: does deadwood matter?
Beetles communities structure differences between farmland and ecological compensation area
Effects of interactive scale-dependent variables on beetle diversity patterns in a semi-arid agricultural landscape
Diversity of ground-dwelling beetles (Coleoptera) in larch plantation with different stages of reforestation in Wolong Natural Reserve, Southwestern China
Composition and seasonal dynamics of litter-layer beetle community in the Dongling Mountain region, North China
Species diversity of insects on the Ordos Plateau, Northwest China
Diversity of litter-layer beetles in Wuyishan Nature Reserve, Jiangxi Province
Insect abundance and environmental effects in Shennongjia Natural Reserve, Hubei Province

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}